Timeline human development
| Embryology - 21 Jun 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Introduction
This page is organised to show a week by week human timeline of development features and approximate timing of key events with more detailed information about specific events in different systems. For a less detailed timeline see week by week.
| From a single cell | to a newborn infant | in 9 months. |
|---|---|---|
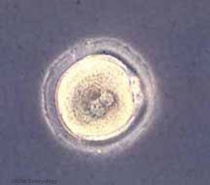
|
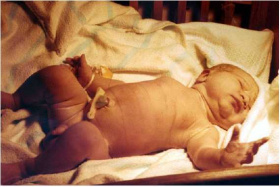
|

|
- "Weeks" refer to embryonic development from fertilization.
- Clinical weeks (shown in brackets) or Gestational Age GA) is from the first day of the Last Menstrual Period (LMP).
- "Stages" refer to the Carnegie stages of development.
- "Timing" refers to days from fertilization or post conception age (PC), not the clinical or gestational age (GA) calculated from LMP (add 2 weeks).
- Dates and staging are also "ideal", and there is significant biological variability in the general timing of events.
- Week 1 to Week 8 (GA 10)are considered the embryonic period of development.
- Week 9 to week 37 (GA 11-39) or birth are considered the fetal period of development.
- First month (4 weeks) after birth is the neonatal period of development.
Each developmental feature is linked to online content with more detailed information and resources such as images and movies. The superscript numbers are the original source references. There are similar "timelines" for other species shown below.
First Trimester
| Week: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| Carnegie stage: | 1 2 3 4 | 5 6 | 7 8 9 | 10 11 12 13 | 14 15 | 16 17 | 18 19 | 20 21 22 23 |
| Links: human timeline | first trimester timeline | second trimester timeline | third trimester timeline | ||||
| Event | ||||
|---|---|---|---|---|
| Menstrual Phase |  menstrual cycle changes: uterus endometrium (loss), ovary (follicle development) | |||

| ||||
| Proliferative Phase |   menstrual cycle changes: uterus endometrium (proliferation), ovary (Follicle Development) menstrual cycle changes: uterus endometrium (proliferation), ovary (Follicle Development)
| |||
| Proliferative Phase | ||||
    menstrual cycle - Mid proliferative menstrual cycle - Mid proliferative
| ||||
  menstrual cycle - Late Proliferative menstrual cycle - Late Proliferative
| ||||
| ovulation
Capacitation |
  | |||
| Event | ||||
| Secretory Phase |    fertilization, zygote, Secretory Phase fertilization, zygote, Secretory Phase
| |||
| Stage 2 |  | |||
| Stage 3 |  blastocyst Hatching (zona pellucida lost) blastocyst Hatching (zona pellucida lost)
| |||
  Late Secretory, blastocyst (free floating) Late Secretory, blastocyst (free floating)
| ||||
| Stage 4 | Adplantation | |||
| Stage 5 | 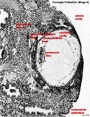
| |||
| Stage 6 | 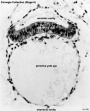 | |||
| Event | ||||
| Stage 7 |   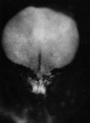 gastrulation, ectoderm, mesoderm, endoderm gastrulation, ectoderm, mesoderm, endoderm
| |||
| Stage 8 | 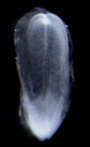 | |||
 | ||||
| Stage 9 | 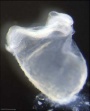 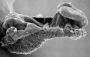 Musculoskeletal somitogenesis, first somites form and continue to be added in sequence caudally (1 - 3 somite pairs). Musculoskeletal somitogenesis, first somites form and continue to be added in sequence caudally (1 - 3 somite pairs).
neural the three main divisions of the brain, which are not cerebral vesicles, can be distinguished while the neural groove is still completely open Neural Crest mesencephalic neural crest is visible[1] | |||
| heart cardiogenesis, week 3 begins as paired heart tubes. | ||||
| Event | ||||
| Stage 10 |  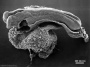 Neural Crest differentiation at spinal cord level from day 22 until day 26 neural folds begin to fuse near the junction between brain and spinal cord, when Neural Crest cells are arising mainly from the neural ectoderm Neural Crest trigeminal, facial, and postotic ganglia components visible[1] Neural Crest migration of vagal level neural crest cells begins (7-10 somite stage) neural rostral neural tube forms 3 primary brain vesicles (week 4) respiratory Week 4 - laryngotracheal groove forms on floor foregut. | |||
| heart begins to beat in Humans by day 22-23, first functioning embryonic organ formed. | ||||
| Stage 11 | 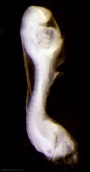
thyroid - thyroid median endodermal thickening in the floor of pharynx neural rostral (or cephalic) neuropore closes within a few hours; closure is bidirectional, it takes place from the dorsal and terminal lips and may occur in two areas simultaneously. The two lips, however, behave differently. ventricular Optic ventricle appears and the neural groove/tube space is initially filled with amniotic fluid.[2] | |||
| Stage 12 |  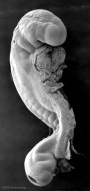
pituitary Week 4 hypophysial pouch, Rathke's pouch, diverticulum from roof liver septum transversum forming liver stroma and hepatic diverticulum forming hepatic trabeculae[3] neural caudal neuropore takes a day to close (closure is approximately at future somitic pair 31/sacral vertebra 2) neural secondary neurulation begins ventricular onset of the ventricular system and separates the ependymal from the amniotic fluid.[2] neural crest cardiac crest, neural crest from rhombomeres 6 and 7 that migrates to pharyngeal arch 3 and from there the truncus arteriosus[1] neural crest vagal neural crest enter the foregut (20-25 somite stage) | |||
| Stage 13 | 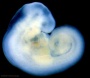 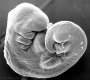 neural the neural tube is normally completely closed, ventricular system now separated from amniotic fluid. Neural crest at spinal level is segregating, and spinal ganglia are in series with the somites. Spinal cord ventral roots beginning to develop.[4] neural the neural tube is normally completely closed, ventricular system now separated from amniotic fluid. Neural crest at spinal level is segregating, and spinal ganglia are in series with the somites. Spinal cord ventral roots beginning to develop.[4]
telencephalon cavity appears Neural - Vascular Development - hindbrain is supplied by two parallel neural arteries (or channels) that obtain their blood supply from carotid-vertebrobasilar anastomoses given by the pharyngeal arch arteries; trigeminal artery, the otic artery, hypoglossal artery, and the proatlantal artery.[5] liver epithelial cord proliferation enmeshing stromal capillaries[3] smell Crest comes from the nasal plates[6] integumentary 4 weeks - simple ectoderm epithelium over mesenchyme integumentary 1-3 months ectoderm- germinative (basal) cell repeated division of generates stratified epithelium; mesoderm- differentiates into connective tissue and blood vessels vision Optic vesicle lies close to the surface ectoderm. The surface ectoderm overlying the optic vesicle, in response to this contact, has thickened to form the lens placode.[7] Diaphragm - pleuroperitoneal fold (PPF) first discernible in human embryos (CRL 6mm).[8] | |||
| pituitary Week 5 elongation, contacts infundibulum, diverticulum of diencephalon
heart Week 5 septation starts, atrial and ventricular respiratory Week 5 left and right lung buds push into the pericardioperitoneal canals (primordia of pleural cavity) Respiratory Week 5 to 17 lung histology - pseudoglandular hearing Week 5 cochlear part of otic vesicle elongates (humans 2.5 turns) | ||||
| Stage 14 | 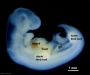  Placodes sensory placodes, lens pit, otocyst, nasal placode, primary/secondary vesicles, fourth ventricle of brain Placodes sensory placodes, lens pit, otocyst, nasal placode, primary/secondary vesicles, fourth ventricle of brain
somite continued segmentation of paraxial mesoderm (somite pairs), heart prominence head 1st, 2nd and 3rd pharyngeal arch, forebrain, site of lens placode, site of otic placode, stomodeum Body - heart, liver, umbilical cord, mesonephric ridge visible externally as bulges. limb upper and lower limb buds growing. Abdominal Wall mesoderm of the primary body wall coalesced in the ventral midline to create the abdominal cavity.[9] neural first appearance of the future cerebral hemispheres. Cerebellar plate differentiated to an intermediate layer, and future rhombic lip identifiable[10] Neural - Vascular Development - basilar artery forms from the consolidation of the neural arteries.[5] ventricular Subarachnoid space initially as irregular spaces on the ventral surface of the spinal cord.[11] liver hepatic gland and its vascular channels enlarge, hematopoietic function appears[3] | |||
| Stage 15 | 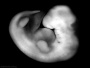
neural cranial nerves (except olfactory and optic) are identifiable in more advanced embryos[12] Neural - Vascular Development - vertebral arteries form from transverse anastomoses between cervical intersegmental arteries, beginning with the proatlantal artery and proceeding downward to the 6th intersegmental artery,[5] vision lens pit is closed. The lens vesicle and optic cup lie close to the surface ectoderm and appear to press against the surface.[7] | |||
| vision 35 to 37 days retinal pigment present | ||||
| pituitary Week 6 - connecting stalk between pouch and oral cavity degenerates
parathyroid Week 6 - diverticulum elongate, hollow then solid, dorsal cell proliferation thymus Week 6 - diverticulum elongate, hollow then solid, ventral cell proliferation adrenal Week 6 - fetal cortex forms from mesothelium adjacent to dorsal mesentery, medulla neural crest cells from adjacent sympathetic ganglia respiratory Week 6 - descent of heart and lungs into thorax. Pleuroperitoneal foramen closes tongue Week 6 - gustatory papilla, caudal midline near the foramen caecum (week 6 to 7 - nerve fibers approach the lingual epithelium) | ||||
| Stage 16 | 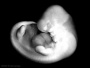 Neural first parasympathetic ganglia, submandibular and ciliary, are identifiable[13] Neural first parasympathetic ganglia, submandibular and ciliary, are identifiable[13]
Neural - Vascular Development - development of the middle cerebral artery is first identified as small buds originating proximal to the anterior cerebral artery on the anterior division of the primitive internal carotid artery.[5] limb upper limb bud nerves median nerve, radial nerve and ulnar nerve entered into hand plate, myoblasts spindle shaped and oriented parallel to limb bud axis. Abdominal Wall muscle cell migration about 25% of the hemicircumference of the abdominal cavity, the lateral plate mesoderm has become more condensed and thicker in the area around the myoblasts.[9] heart outflow tract elliptical configuration with four cushions, the two larger fusing at this stage. Semilunar valve leaflets form at the downstream end of the cushions head lip and palate components of the upper lip, medial nasal prominence and maxillary process present, median palatine process appears. Eyelid prior to the development of the eyelids, one small sulcus or groove forms above the eye (eyelid groove) and another below it.[7] | |||
| Stage 17 | 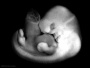
| |||
| heart separation of common cardiac outflow (aortic arch and pulmonary aorta) | ||||
| Event | ||||
| pancreas Week 7 to 20 pancreatic hormones secretion increases, small amount maternal insulin
respiratory Week 7 - enlargement of liver stops descent of heart and lungs | ||||
| Stage 18 | 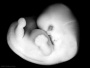
limb bone forms by endochondrial ossification and throughout embryo replacement of cartilage with bone (week 5-12). neural smell vomeronasal fibres and nervus terminalis[6] liverobturation due to epithelial proliferation, bile ducts became reorganized, continuity between liver cells and gut[3] ventricular duramater appears and spaces surround the circumference of the spinal cord, which coalesce and contain many blood vessels.[15] Female uterus opening of the paramesonephric (Müllerian) duct to the coelomic cavity formed as an invagination of the coelomic epithelium[16] Abdominal Wall separation of the myoblasts into distinct inner and outer layers, with unidirectional orientation. Abdominal wall thicker in the region where secondary structures were forming compared with the primary body wall region, dorsally outermost layer of connective tissue approximately half of this thickness.[9] | |||
|
liver (stage 18 to 23) biliary ductules developed in periportal connective tissue produces ductal plates that receive biliary capillaries[3] | ||||
| Stage 19 | 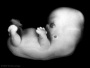
| |||
| Stage 20 | 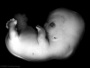
Head scalp vascular plexus visible limb upper limbs begin to rotate ventrally neural amygdaloid body has at least four individual nuclei[24] oculomotor nerve shows a dorsolateral and a ventromedial portion rhombic lip (rhombencephalon) formation of the cerebellum (intermediate layer) and of the cochlear nuclei cerebellum cell layer (future Purkinje cells) develops choroid plexuses of the fourth and lateral ventricles | |||
| gastrointestinal tract anal membrane perforates | ||||
| Stage 21 | 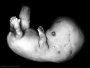
neural cortical plate appears in the area of future insula[25] Neural - Vascular Development - formation of the anterior communicating artery.[5] limb upper and lower limbs rotate Intraembryonic Coelom pericardioperitoneal canals close Abdominal Wall Myoblasts have reached the ventral midline and myotubes were present and oriented uniformly within all muscle groups. The rectus abdominis formed distinct bundles of muscle. Connective tissue layers comprised the majority of the thickness of the abdominal wall, outermost layer of connective tissue accounted for the majority of this thickness.[9] | |||
| Stage 22 | 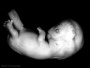 neural neocortical fibres project to epithalamus, to dorsal thalamus, and to mesencephalon[25] neural neocortical fibres project to epithalamus, to dorsal thalamus, and to mesencephalon[25]
limb fingers and toes lengthen smell Stage 22 to early fetal period - migratory streams of neurons from the subventricular zone of the olfactory bulb towards the future claustrum[6] Uterus Vagina fused duct (uterovaginal canal) bifurcated at the caudal portion at Carnegie stages 22 and 23[16] | |||
| Genital 8 Weeks Testis - mesenchyme, interstitial cells (of Leydig) secrete testosterone, androstenedione
Genital 8 to 12 Weeks - hCG stimulates testosterone production Tongue Week 8 - nerves penetrate epitheilai basal lamina and synapse with undifferentiated, elongated, epithelial cells (taste bud progenitor cell)[26] | ||||
| Stage 23 | 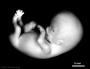 Stage 23 defines the end of the embryonic (organogenesis) period Stage 23 defines the end of the embryonic (organogenesis) period
Mesoderm heart prominence, ossification continues Head nose, eye, external acoustic meatus, eyelids, external ears, rounded head Body - straightening of trunk, umbilical cord, intestines herniated at umbilicus limb upper limbs longer and bent at elbow, hands and feet turned inward, foot with separated digits, wrist, hand with separated digits Extraembryonic Coelom chorionic cavity is now lost by fusion with the expanding amniotic cavity neural rhombencephalon, pyramidal decussation present, nuclei and tracts similar to those present in the newborn cerebellum present as only a plate connected to midbrain and hindbrain through fibre bundles[27] Axial Skeleton vertebral column 33 or 34 cartilaginous vertebrae (20-33 mm in total length), vertebral pedicles, articular and transverse processes identifiable (no spinous processes)[28] Abdominal Wall Rectus muscle forms 2 or 3 distinct layers with myotube orientation uniform in all muscles. The external oblique and internal oblique started to expand in thickness, transversus a thin layer of muscle.[9] | |||
| Week 8 | Stomach Week 8 - Gastrin containing cells in stomach antrum. Somatostatin cells in both the antrum and the fundus.
Genital - Female Development paired paramesonephric (Müllerian) ducts contact each other and are fused into a single tube that separates again and returns to the mesonephric (Wolffian) ducts. The paramesonephric ducts have not yet reached the urogenital sinus.[16] | |||
| Week 9 | Beginning of Fetal Development
CRL 43 mm, femur length 6 mm 9 weeks CRL 50 mm - genital genitalia in both sexes look identical[30] uterus - paramesonephric ducts come into apposition with the urorectal septum and begin to fuse | |||
| Event | ||||
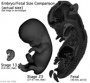
Gastrointestinal Tract Week 10 intestines in abdomen Pituitary growth hormone and ACTH detectable Pancreas Week 10 glucagon (alpha) differentiate first, somatostatin (delta), insulin (beta) cells differentiate, insulin secretion begins Tongue Week 10 shallow grooves above the taste bud primordium Stomach Week 10 - Glucagon containing cells in stomach fundus. Nail Development fingernails appear outer ear Week 10 - Meatal plug extends in a disc-like fashion, the meatus is boot-shaped with a narrow neck and the sole of the meatal plug spreading widely to form the future tympanic membrane medially. Proximal portion of the neck starts to be resorbed. inner ear Week 10 - neural-crest-derived melanocytes migrate into the cochlea. They penetrate the basement membrane of the lateral wall epithelium and develop into the intermediate cells of the stria vascularis.[31] | ||||
| Week 10 - CRL 55 mm, femur length 9 mm, biparietal diameter 17 mm | ||||
| Event
neural - Cerebrum appearance of the first sulcus (week 11-15, GA 13-17 weeks)[32] | ||||
|
Thyroid colloid appearance in thyroid follicles, iodine and thyroid hormone (TH) synthesis Stomach Week 11 - Serotonin containing cells in both the antrum and the fundus. | ||||
| Week 11 - CRL 68 mm, femur length 12 mm, biparietal diameter 20 mm | ||||
| ||||

Second Trimester
(GA Clinical Week 14) Second Trimester
| Links: human timeline | first trimester timeline | second trimester timeline | third trimester timeline | ||||
| Event | ||||
| Clinical second trimester |  Week 12 - CRL 85 mm, femur length 15 mm, biparietal diameter 25 mm Week 12 - CRL 85 mm, femur length 15 mm, biparietal diameter 25 mm
Hearing Week 12-16 - Capsule adjacent to membranous labrynth undegoes vacuolization to form a cavity (perilymphatic space) around membranous labrynth and fills with perilymph
Respiratory Month 3-6 - lungs appear glandular, end month 6 alveolar cells type 2 appear and begin to secrete surfactant Tongue Week 12 - first differentiated epithelial cells (Type II and III) Genital female genital canal (80 days) formed with absorption of the median septum | |||
| tongue Week 12 to 13 - maximum synapses between cells and afferent nerve fibers
hearing outer ear Week 13 - Meatal plug disc-like, innermost surface in contact with the primordial malleus, contributes to the formation of the tympanic membrane. | ||||
| tongue Week 14 to 15 - taste pores develop, mucous
ovary 100 days - primary follicles present nail toenails appear Head Development facial skeleton remodelling begins Hearing - Inner Ear Development Week 14 GA 16 - neural-crest-derived melanocytes, now intermediate cells of the stria vascularis, tightly integrate with Na+ /K+ -ATPase-positive marginal cells, which started to express KCNQ1 in their apical membrane.[31] | ||||
| Pancreas glucagon detectable in fetal plasma.
spleen Week 15 -alpha-smooth muscle actin (alpha-SMA)-positive reticulum cells scattered around the arterioles.[33] | ||||
| 14 cm | 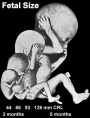 Hearing Week 16-24 - Centres of ossification appear in remaining cartilage of otic capsule form petrous portion of temporal bone. Continues to ossify to form mastoid process of temporal bone. Hearing Week 16-24 - Centres of ossification appear in remaining cartilage of otic capsule form petrous portion of temporal bone. Continues to ossify to form mastoid process of temporal bone.
pituitary adenohypophysis fully differentiated respiratory Week 16 to 25 lung histology - canalicular Hearing - Outer Ear Development Week 16.5 - External auditory meatus is fully patent throughout its length, lumen is still narrow and curved. Hearing - Inner Ear Development Week 16 GA 18 - cells in the outer sulcus express KCNJ10 and gap junction proteins GJB2/CX26 and GJB6/CX30, but these are not expressed in the spiral ligament.[31] gap junction cartoon neural - Cerebrum development of the periinsular sulci (week 16-17, GA 18-19 weeks)[32]
primary follicles begin to form in the ovary and are characterized by an oocyte glandular urethra forms and skin folds present | |||
 Neural - Brain development histology week 17 Neural - Brain development histology week 17
Cerebellum Magnetic Resonance Imaging (MRI) can study the developing cerebellum from 17 to 18 weeks (GA 19 to 20 weeks). tooth Week 17 - First papilla of the permanent dentition appear (first molar) immediately behind the second milk molar, milk teeth are well advanced (Fetus 180 mm). | ||||
 tongue Week 18 - substance P detected in dermal papillae, not in taste bud primordia tongue Week 18 - substance P detected in dermal papillae, not in taste bud primordia
integumentary vernix caseosa covers skin spleen Week 18 - alpha-SMA-positive reticulum cells increase in number and began to form a reticular framework. An accumulation of T and B lymphocytes occurred within the framework, and a primitive white pulp was observed around the arterioles.[33] Hearing - Outer Ear Development week 18 - External auditory meatus is already fully expanded to its complete form. neural - Cerebrum central sulci and opercularization of the insula (week 18-20, GA 20-22 weeks)[32] | ||||
| neural week 19 neuronal migration ends and the radial glial cells that aided the migration now become transformed into astrocytes and astrocytic precursors.[34] | ||||
| pituitary week 20 to 24 growth hormone levels peak, then decline
integumentary lanugo, skin hair integumentary 5 months - Hair growth initiated at base of cord, lateral outgrowths form associated sebaceous glands; Other cords elongate and coil to form sweat glands; Cords in mammary region branch as they elongate to form mammary glands. touch pacinian corpuscle begin to develop[35] | ||||
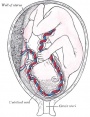 Neural brain cortical sulcation - sylvian fissure, interhemispheric fissure, callosal sulcus, parietooccipital fissure, and hippocampic fissures present[36] Neural brain cortical sulcation - sylvian fissure, interhemispheric fissure, callosal sulcus, parietooccipital fissure, and hippocampic fissures present[36]
spleen - Week 22 - antigenic diversity of the reticular framework was observed, and T and B lymphocytes were segregated in the framework. T lymphocytes were sorted into the alpha-smooth muscle actin-positive reticular framework, and the periarteriolar lymphoid sheath (PALS) was formed around the arteriole. B lymphocytes aggregated in eccentric portions to the PALS and formed the lymph follicle (LF). The reticular framework of the LF was alpha-SMA-negative. [33] neural - Cerebrum covering of the posterior insula (week 22-24, GA 24-26 weeks)[32] | ||||
| respiratory Week 24 to 40 lung histology - terminal sac
spleen Week 24 - marginal zone appeared in the alpha-smooth muscle actin-positive reticular framework around the white pulp.[33] tooth Week 24 - Permanent incisors and canines appear. Earliest potential survival expected if born ovary follicles can consist of growing oocytes surrounded by several layers of granulosa cells | ||||
| respiratory end month 6 alveolar cells type 2 appear and begin to secrete surfactant
neural - Cerebrum closure of the laeteral sulcus (Sylvian fissure or lateral fissure) (week 25-26, GA 27-28 weeks)[32] | ||||
| touch pacinian corpuscle well developed[35] | ||||
| ||||
Third Trimester
| Links: human timeline | first trimester timeline | second trimester timeline | third trimester timeline | ||||
| Event | ||||
| Clinical third trimester | hearing 3rd Trimester - vibration acoustically of maternal abdominal wall induces startle respone in fetus.
| |||
| respiratory Month 7 - respiratory bronchioles proliferate and end in alveolar ducts and sacs | ||||
|
tooth Week 29 - Permanent premolars (correspond to the milk molars) appear. | ||||
|
Genital male gonad (testes) descending | ||||
| nail fingernails reach digit tip | ||||
| neural brain cortical sulcation - primary sulci present[36] | ||||
| neural brain cortical sulcation - insular, cingular, and occipital secondary sulci present[36] | ||||
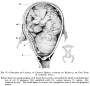 Nail Development toenails reach digit tip Nail Development toenails reach digit tip
Lens Development - lens growth and interocular distance plateaus after 36 weeks of gestation[37] | ||||
| Birth | 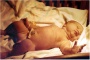 Clinical Week 40 Clinical Week 40
Heart pressure difference closes foramen ovale leaving a fossa ovalis thyroid TSH levels increase, thyroxine (T3) and T4 levels increase to 24 h, then 5-7 days postnatal decline to normal levels adrenal - zona glomerulosa, zona fasiculata present | |||
| ||||
Postnatal
| Event | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| vision - eye globe growth plateaus after 42 weeks of gestation[37] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| testis | spermatozoa - about 2 months of age, primordial germ cells (gonocytes) are replaced by adult dark (Ad) and pale (Ap) spermatogonia forming the spermatogonial stem cell (SSC) population that at puberty will commence differentiation into spermatozoa. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| neural Hearing (6 months to 5 years) thalamocortical afferents to the deeper cortical layers mature and are the first source of input to the auditory cortex[38]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Adrenal - Year 3 zona reticularis present.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| neural hearing - (5 to 12 years) commissural and association axons in the superficial cortical layers allows communication between subdivisions of the auditory cortex[38] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| puberty - Female | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| puberty - Male | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
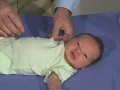
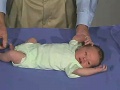
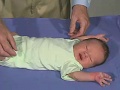



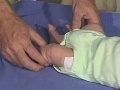









{kind=link}
{kind=link}
Human Systems
| Human Systems: bone timeline | eye neural crest timeline | hearing timeline | heart abnormality timeline | hearing EAM timeline | muscle timeline | ovary timeline | placental villi timeline | shoulder timeline | smell timeline | spleen timeline | ventricular timeline | comparitive |
Somite
| Week | Days | Carnegie Stage | Somite Number (pairs) |
|---|---|---|---|
| Week 3 | 19 - 21 | 9 image | 1 - 3 |
| Week 4 | 22 - 23 | 10 image | 4 - 12 |
| Week 4 | 23 - 26 | 11 image | 13 - 20 |
| Week 4 | 26 - 30 | 12 image | 21 - 29 |
| Week 5 | 28 - 32 | 13 image | 30 |
| Week 5 | 31 - 35 | 14 image | 30+ |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Limb
| Carnegie stages for limb elements appearance | ||||
|---|---|---|---|---|
| Skeletal Element | Condensed Mesenchyme |
Cartilage | Bone | |

|
Humerus | 16 | 16 — 17 | 21 — 22 |
| Radius | 16 | 17 | 21 — 23 | |
| Ulna | 16 | 17 — 18 | 17 — 23 | |
| Hand | 17 | 17 — 21 | In fetus1and after birth | |
| Femur | 17 | 17—18 | 22 — 23 | |
| Patella | 20 | 21 | After birth | |
| Tibia | 17 | 17 — 23 | 22 — 23 | |
| Fibula | 17 | 17 — 18 | In fetus | |
| Foot | 17 — 18 | 18 — 23 or later | in fetus and after birth | |
| 1 Intramembranous ossification at the tips of the distal phalanges of the hand may be in Stage 23. | ||||
| Reference - O'Rahilly R. Gray DI. and Gardner E. Chondrification in the hands and feet of staged human embryos. (1957) Carnegie Instn. Wash. Publ. 611, Contrib. Embryol., 36: | ||||
Neural
| Early Neural Timeline | |
|---|---|
| Carnegie Stage | Event |
| 8 | (about 18 postovulatory days) neural groove and folds are first seen |
| 9 | three main divisions of the brain, which are not cerebral vesicles, can be distinguished while the neural groove is still completely open. |
| 10 | (two days later) neural folds begin to fuse near the junction between brain and spinal cord, when neural crest cells are arising mainly from the neural ectoderm |
| 11 | (about 24 days) the rostral (or cephalic) neuropore closes within a few hours; closure is bidirectional, it takes place from the dorsal and terminal lips and may occur in several areas simultaneously. The two lips, however, behave differently. |
| 12 | (about 26 days) The caudal neuropore takes a day to close. The level of final closure is approximately at future somitic pair 31 (corresponds to the level of sacral vertebra 2). Secondary neurulation begins, is the differentiation of the caudal part of the neural tube from the caudal eminence (or end-bud) without the intermediate phase of a neural plate. |
| 13 | (4 weeks) the neural tube is normally completely closed. |
| Links: neural | Week 3 | Week 4 | |

Neural timeline overview[39]
Sensorimotor Cortex
| Sensorimotor Cortex Timeline | ||
|---|---|---|
| Gestational Age GA weeks | Fertilization Age FA weeks | Event |
| 18-19 | 16-17 | appearance of the inferior part of the central cerebral sulcus |
| 20-22 | 18-20 | development of the pericentral lateral regions and the beginning of opercularization |
| 24-26 | 22-24 | development of parietal and temporal cortices and the covering of the postcentral insular region |
| 27-28 | 25-26 | maturation of the central cerebral regions |
| Table data.[40] Links: sensorimotor cortex | cerebral cortex | second trimester | Sensory System Development | ||
Notochord
| Carnegie Stage | Days | Event |
|---|---|---|
| 7 | 15 to 17 | notochordal process directly rostral to the area of gastrulation. |
| 8 to 10 | 17 to 23 | group of undifferentiated cells that briefly persists. Within the notochordal process, a connection between the amniotic cavity and yolk-sac (neurenteric canal). |
| 8 to 11 | 17 to 26 | notochordal process incorporates entirely into the endoderm, forming the epithelial notochordal plate. |
| 10 to 11 | 21 to 26 | notochordal plate then acquires an “inverted U-shape”, relatively lengthens during these stages and remains intimately associated with the neural tube until stage 12 (26–30 days). |
| 12 | 26 to 30 | notochordal plate detaches completely from the endoderm to form the definitive notochord. |
| 12 to 14 | 26 to 35 | definitive notochord formation then allows the paired dorsal aortae to fuse in-between the notochord and the roof of the gut. |
| Table data[41] Links: notochord | timeline | Week 3 | Week 4 |
Ventricular
| Week | Carnegie Stage | Event | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Week 4 | 11 | appearance of the optic ventricle. The neural groove/tube space is initially filled with amniotic fluid. | ||||||||||||||||||
| 12 | closure of the caudal neuropore, onset of the ventricular system and separates the ependymal from the amniotic fluid. | |||||||||||||||||||
| 13 | cavity of the telencephalon medium is visible. | |||||||||||||||||||
| Week 5 | 14 | cerebral hemispheres and lateral ventricles begin, rhomboid fossa becomes apparent. | ||||||||||||||||||
| 15 | medial and lateral ventricular eminences cause indentations in the lateral ventricle | |||||||||||||||||||
| Week 6 | 16 | hypothalamic sulcus is evident. | ||||||||||||||||||
| 17 - 18 | interventricular foramina are becoming relatively smaller, and cellular accumulations indicate the future choroid villi of the fourth and lateral ventricles. | |||||||||||||||||||
| Week 7 | 18 | areae membranaceae rostralis and caudalis are visible in the roof of the fourth ventricle, and the paraphysis is appearing. | ||||||||||||||||||
| 19 | choroid villi are visible in the fourth ventricle, and a mesencephalic evagination (blindsack) is visible | |||||||||||||||||||
| Week 8 | 20 | choroid villi are visible in the lateral ventricle. | ||||||||||||||||||
| 21 | olfactory ventricle is visible. | |||||||||||||||||||
| 21 - 23 | lateral ventricle has become C-shaped (anterior and inferior horns visible). Recesses develop in the third ventricle (optic, infundibular, pineal). | |||||||||||||||||||
| ||||||||||||||||||||
| Links: ventricular | neural | timeline | Category:Timeline Table Data Reference[42] | ||||||||||||||||||||
Neural Crest
| Carnegie Stage | Event |
|---|---|
| 9 | an indication of mesencephalic neural crest |
| 10 | trigeminal, facial, and postotic components |
| 11 | crest-free zones are soon observable in rhombomere 1, 3, and 5 |
| 12 | rhombomeres 6 and 7 neural crest migrate to pharyngeal arch 3 and then rostrad to the truncus arteriosus |
| 13 | nasal crest and the terminalis-vomeronasal complex are last of the cranial crest to appear |
| 9 to 14 | otic vesicle primordium descends |
| Week: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| Carnegie stage: | 1 2 3 4 | 5 6 | 7 8 9 | 10 11 12 13 | 14 15 | 16 17 | 18 19 | 20 21 22 23 |
Data from a study of 185 serially sectioned staged (Carnegie) human embryos.[1] Links: vision | neural crest | timeline | Category:Timeline
Hearing
| Developmental Time | Event |
|---|---|
| Week 9 | Mesenchyme surrounding membranous labryinth (otic capsule) chondrifies. |
| Week 12-16 | Capsule adjacent to membranous labryinth undegoes vacuolization to form a cavity (perilymphatic space) around membranous labrynth and fills with perilymph. |
| Week 18 | ectodermal plug in external auditory meatus breaks down. |
| Week 16 - 24 | Centres of endochondral ossification appear in remaining cartilage of otic capsule form petrous portion of temporal bone. Continues to ossify to form mastoid process of temporal bone. |
| Week 18 - 22 | Organ of corti structural elements develop. (GA 20 - 24 weeks) |
| 26 weeks | human brainstem auditory pathway is anatomically formed. |
| 28 weeks | AABR can be recorded. |
| 3rd Trimester | Vibration acoustically of maternal abdominal wall induces startle response in fetus. |
| Links: hearing | timeline | |
| Carnegie Stage |
CRL (mm) | Description |
|---|---|---|
| 13 | 6 | Presumptive stapedial area |
| 14 | 7 | Appearance of the stapedial anlage |
| 16 | 9 | Relationship between the stapedial artery and the stapedial anlage. Appearance of the interhyale |
| 17 | 12 | Delimitation of the parts of the stapedial anlage |
| 18 | 16 | Chondrogenesis phase. Start of involution of the stapedial artery |
| 20 | 18.5 | Delimitation of the ossicular anlages. Cartilaginous phase. Disappearance of the stapedial artery |
| 22 | 26 | Delimitation of the interhyale |
| 23 | 28 | Anlage of the stapedial muscle tendon |
| Data from Table 1[43] Links: middle ear | ||
| Period | Week | Description |
| Embryo | week 8 | Funnel-shaped tube continues medially into mesenchymal tissue, forms a curved path. |
| Fetus (first trimester) | week 9 | Ectodermal cells proliferate, fill the meatus lumen and form the "meatal plug". |
| Fetus (first trimester) | week 10 | Meatal plug bottom extends in a disc-like fashion, so that in the horizontal plane the meatus is boot-shaped with a narrow neck and the sole of the meatal plug spreading widely to form the future tympanic membrane medially. At the same time, the plug in the proximal portion of the neck starts to be resorbed. |
| Fetus (second trimester) | week 13 | Meatal plug disc-like, innermost surface in contact with the primordial malleus, contributes to formation of tympanic membrane. |
| Fetus (second trimester) | week 15 | Meatal plug innermost portion splits, leaving a thin ectodermal cell layer of immature tympanic membrane. The neck of the boot forms the border between the primary and secondary meatus, and is the last part to split. |
| Fetus (second trimester) | week 16.5 | The meatus is fully patent throughout entire length. Lumen is still narrow and curved. Epithelium cornification commences. |
| Fetus (second trimester) | week 18 | The meatus is now fully expanded to its complete form. |
| Links: outer ear | hearing | timeline Reference[44] | ||
Vision
| Carnegie Stage | Event | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10 | optic primordia appear. | ||||||||||||||||||
| 11 | Right and left optic primordia meet at the optic chiasma forming a U-shaped rim. | ||||||||||||||||||
| 12 | optic neural crest reaches its maximum extent and the optic vesicle becomes covered by a complete sheath, | ||||||||||||||||||
| 13 | By the end of the fourth week the optic vesicle lies close to the surface ectoderm. Optic evagination differentiation allows identification of optic part of retina, future pigmented layer of retina, and optic stalk. The surface ectoderm overlying the optic vesicle, in response to this contact, has thickened to form the lense placode. | ||||||||||||||||||
| 14 | (about 32 days) Lens placode is indented by the lens pit, cup-shaped and still communicates with the surface by a narrowing pore. | ||||||||||||||||||
| 15 | (about 33 days) Lens pit is closed. The lens vesicle and optic cup lie close to the surface ectoderm and appear to press against the surface. | ||||||||||||||||||
| 16 | (37 days) Growth of the lens body results in a D-shaped lens cavity. Perilental blood vessels (tunica vasculosa lentis) are visible. Prior to the development of the eyelids, one small sulcus or groove forms above the eye (eyelid groove) and another below it. | ||||||||||||||||||
| 17 - 19 | Retinal pigment is visible and the retinal fissure is largely closed. Eyelids grooves deepen, eyelid folds develop, first below, and then above, the eye. | ||||||||||||||||||
| 18 | Mesenchyme invades the region between the lens epithelium and the surface ectoderm. | ||||||||||||||||||
| 19 - 22 | Eyelid folds develop into the eyelids and cover more of the eye as the palpebral fissure takes shape. The upper and the lower eyelids meet at the outer canthus in Stage 19. | ||||||||||||||||||
| 20 | Lens cavity is lost and a lens suture begins to form. The inner canthus is established. | ||||||||||||||||||
| 23 | retina comprises the pigmented layer, external limiting membrane, proliferative zone, external neuroblastic layer, transient fiber layer, internal neuroblastic layer, nerve fiber layer, and internal limiting membrane. Eyelids closure is complete (Note - shown as still open in the Kyoto embryo). | ||||||||||||||||||
Data from a study of human embryonic carnegie stages[7] and other sources.
| |||||||||||||||||||
| Carnegie Stage | Event |
|---|---|
| 9 | an indication of mesencephalic neural crest |
| 10 | trigeminal, facial, and postotic components |
| 11 | crest-free zones are soon observable in rhombomere 1, 3, and 5 |
| 12 | rhombomeres 6 and 7 neural crest migrate to pharyngeal arch 3 and then rostrad to the truncus arteriosus |
| 13 | nasal crest and the terminalis-vomeronasal complex are last of the cranial crest to appear |
| 9 to 14 | otic vesicle primordium descends |
| Week: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| Carnegie stage: | 1 2 3 4 | 5 6 | 7 8 9 | 10 11 12 13 | 14 15 | 16 17 | 18 19 | 20 21 22 23 |
Data from a study of 185 serially sectioned staged (Carnegie) human embryos.[1] Links: vision | neural crest | timeline | Category:Timeline
| Carnegie Stage | Event |
|---|---|
| 10 | optic primordia appear |
| 13 | By the end of the fourth week the optic vesicle lies close to the surface ectoderm. The surface ectoderm overlying the optic vesicle, in response to this contact, has thickened to form the lense placode. |
| 14 | (about 32 days) the lens placode is indented by the lens pit. |
| 15 | (about 33 days) the lens pit is closed. The lens vesicle and optic cup lie close to the surface ectoderm and appear to press against the surface. |
| 16 | (37 days) Prior to the development of the eyelids, one small sulcus or groove forms above the eye (eyelid groove) and another below it. |
| 17 - 19 | grooves deepen, eyelid folds develop, first below, and then above, the eye. |
| 19 - 22 | eyelid folds develop into the eyelids and cover more of the eye as the palpebral fissure takes shape. The upper and the lower eyelids meet at the outer canthus in Stage 19. |
| 20 | the inner canthus is established. |
| 23 | closure of the eyelids is complete (Note - shown as still open in some Kyoto embryo). |
| Data - from Kyoto embryos with Carnegie staging.[7] Links: eyelid |
Smell
| Week FA (GA) | Carnegie Stage | Event |
|---|---|---|
| week 4 (GA 6) | 11 | nasal epiblastic thickening appears |
| 12 | nasal field is well outlined | |
| week 5 (GA 7) | 15 | continuous cellulovascular strand seen between the nasal groove and the olfactory field |
| week 6 (GA 8) | 16 | vomeronasal groove appears. |
| 17 | olfactory nerve is organized into two plexuses, lateral and medial, the latter mingled with the terminal-vomeronasal complex. | |
| week 7 (GA 9) | 18 | olfactory bulb begins to appear |
| 19 | individualization of the olfactory bulb and nuclei, distinction between olfactory structures and terminal and vomeronasal ones begins to be clear. | |
| week 8 (GA 10) | 21 | structure of the olfactory bulb is evident. |
| 23 | olfactory strands are well individualized, and olfactory and terminal-vomeronasal fibers are easily distinguishable. | |
| Links: smell | sensory | timeline | Category:Timeline Table Data Reference[45] | ||
Cardiovascular
Heart
| Characteristic | Carnegie stage: | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Septum primum | ||||||||||||
| Foramen primum | ||||||||||||
| Atrioventricular bundle | ||||||||||||
| Atrioventricular cushions | ||||||||||||
| Conotruncal ridges | ||||||||||||
| Foramen secundum | ||||||||||||
| Semilunar cusps | ||||||||||||
| Conotruncal septum; atria | ||||||||||||
| Closure primum foramen | ||||||||||||
| Fusion atrioventricular cushions | ||||||||||||
| Septum secundum and foramen ovale | ||||||||||||
| Closure secondary interventricular foramen | ||||||||||||
| Chordae tendineae | ||||||||||||
| Colour Coding: | beginning to appear | present | Table data[46] Links: heart | Madrid Collection | |||||||||
| Table 1. Comparison of Human and Murine Development | |||||
|---|---|---|---|---|---|
| Human | Mouse | ||||
| Carnegie Stage |
Post-ovulatory Days |
Size (mm) Greatest Length |
Theiler Stage |
Post-conception Days |
Size (mm) Crown-rump |
| 11 | 24 | 2.5 - 4.5 | 14 | 9 - 9.5 | 2.1 |
| 12 | 26 | 3 - 5 | 15 | 9.5 - 10.25 | 2.5 |
| 13 | 28 | 4 - 6 | 16 | 10.25 - 10.5 | 3.6 |
| 14 | 32 | 5 - 7 | 17 | 10.5 | 4.1 |
| 15 | 33 | 7 - 9 | 18 | 11 | 4.6 |
| 16 | 37 | 8 - 11 | 19 | 11.5 | 6 - 7 |
| 17 | 41 | 11 - 14 | 20 | 12 | 7 |
| 18 | 44 | 13 - 17 | 21 | 12.5 - 13 | 8.2 |
| 19 | 47.5 | 16 - 18 | 21 | 12.5 - 13 | 8.2 |
| 20 | 50.5 | 18 - 22 | 22 | 13.5 - 14 | 9.1 |
| 21 | 52 | 22 - 24 | 22 | 13.5 - 14 | 9.1 |
| 22 | 54 | 23 - 28 | 22 | 13.5 - 14 | 9.1 |
| 23 | 56.5 | 27 - 31 | 22 | 13.5 - 14 | 9.1 |
| 26 | 17.5 - 18 | 17.8 | |||
|
Notes: Human Embryonic Development | Carnegie Stages | Mouse Development | Theiler Stage | Post-conception Days | Carnegie Stage Comparison Reference: Anderson RH. Teratogenecity in the setting of cardiac development and maldevelopment. (2016) | |||||
{kind=link}
Cerebral Arterial
| Carnegie Stage | CRL (mm) | Event |
|---|---|---|
| 13 | 4 - 5 | hindbrain (future posterior fossa) is supplied by two parallel neural arteries (or channels). These arteries obtain their blood supply from carotid-vertebrobasilar anastomoses given by the trigeminal artery (TA), the otic artery (OA), hypoglossal artery (HA), and the proatlantal artery (ProA) |
| 14 | 5 - 8 | basilar artery (BA) forms from the consolidation of the neural arteries. |
| 15 | 7 - 12 | vertebral arteries (VA) forms from transverse anastomoses between cervical intersegmental arteries, beginning with the ProA and proceeding downward to the 6th intersegmental artery, |
| 16 | 11 - 12 | (35 days) development of the middle cerebral artery (MCA) is first identified as small buds originating proximal to the anterior cerebral artery (ACA) on the anterior division of the primitive internal carotid artery (ICA). |
| 19 | 16 - 18 | middle cerebral artery (MCA) becomes more prominent, the plexi fuse into a single artery and further branches pierce the cerebral hemisphere. |
| 20 | 18 | stem of the ACA gives rise to the olfactory artery. |
| 21 | 21-24 | formation of the anterior communicating artery (ACOMM). |
| Early development - the posterior circulation relies almost entirely from blood supply coming from the anterior circulation through carotid-vertebrobasilar anastomoses. | ||
| Later development - as the posterior fossa structures and the occipital lobe grow, the posterior circulation becomes progressively independent from the anterior circulation with obliteration of the anterior-posterior anastomoses from caudal to rostral maintaining in the majority of adult only one connection between the distal basilar arteries with the carotid artery via the posterior communicating artery. | ||
| Data source[5]) Links: neural vascular | ||
Placenta
| Fertilization Age
(weeks) |
Gestational Age
(weeks) |
Vessel Lumen Diameter
(range in microns, µm) |
Features |
| 3 to 4 | 5 and 6 | 10 - 15 |
|
| 5 to 6 | 7 and 8 | 10 - 26 |
|
| 7 to 8 | 9 and 10 | 60 - 75 two central vessels
26 - 34 capillary network |
|
| 9 to 10 | 11 and 12 | 70 - 90 two central vessels
26 - 34 capillary network |
|
| Vill development data based upon immunochemistry confocal laser scanning microscope (CLSM) study[47] with clinical gestational age (GA) from last menstrual period (LMP) and has been corrected for post-conception (fertilization) age, approximately 14 days later.
CD31 - (PECAM-1, Platelet Endothelial Cell Adhesion Molecule) is a cluster of differentiation molecule found on endothelial and other blood cells. | |||
Liver
| Feature | ||
|---|---|---|
| hepatic diverticulum development (ductal plate) | ||
| cell differentiation
septum transversum forming liver stroma hepatic diverticulum forming hepatic trabeculae | ||
| epithelial cord proliferation enmeshing stromal capillaries | ||
| hepatic gland and its vascular channels enlarge
hematopoietic function appeared | ||
| obturation due to epithelial proliferation
bile ducts became reorganized (continuity between liver cells and gut) | ||
| biliary ductules developed in periportal connective tissue
produces ductal plates that receive biliary capillaries | ||
| Human data[3], see also liver development in the rat embryonic period (Carnegie stages 15-23).[48] (More? Detailed Timeline | Timeline human development) | ||
| Carnegie Stage | Age (days) | CRL (mm) | Biliary system | Vascular | Hepatic parenchyma |
|---|---|---|---|---|---|
| 14 | 33 | 7 |
|
|
|
| 18 | 46 | 15 |
|
|
|
| 21 | 53 | 22.5 | Bile duct morphology as earlier stage. Common bile duct empties at the level of the proximal duodenum. |
|
Hepatic parenchyma a large rounded mass. |
| 23 | 58 | 27 | Bile duct morphology as earlier stage. |
|
|
| Data from a recent human study[49] Links: liver | Carnegie stage 14 | 18 | 21 | 23 | simple embryonic timeline | Timeline human development | |||||
Pancreas
| Carnegie Stage | Days | Event |
|---|---|---|
| 10 | 25-27 | distal endoderm foregut |
| 12 | 29-31 | pancreatic duodenal endoderm
extra-hepatic billiard duct |
| 13 | 30-33 | pancreatic bud |
| 19 | 47 | "trunk" progenitor
"tip" progenitor |
| 23 | 8 weeks + | fetal beta cell
ductal cell |
| fetal | 14 weeks | acinar cell |
| Table data[50] Links: pancreas | exocrine pancreas | pancreas molecular timeline | timeline | ||
| Human Pancreas Timeline | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||
| Carnegie Stage | Days | Molecular | Event |
|---|---|---|---|
| 10 | 25-27 | FOXA2, SOX17 (dorsal), SHH (ventral) | distal endoderm foregut |
| 12 | 29-31 | PDX1, FOXA2, SOX17, GATA4, SOX9 (weak)
SOX9, FOXA2, PDX1 (weak) |
pancreatic duodenal endoderm
extra-hepatic billiard duct |
| 13 | 30-33 | PDX1, FOXA2, GATA4, NKX6.1, SOX9 | pancreatic bud |
| 19 | 47 | PDX1, FOXA2, GATA4, NKX6.1, SOX9
PDX1, FOXA2, GATA4, NKX6.1, SOX9 |
"trunk" progenitor
"tip" progenitor |
| 23 | 8 weeks + | PDX1, FOXA2, GATA4, NKX6.1, NKX2.2, ISL1
SOX9, FOXA2, PDX1 (weak) |
fetal beta cell
ductal cell |
| fetal | 14 weeks | GATA4 | acini cell |
| Table data[50] Links: pancreas | exocrine pancreas | timeline | |||
| Human Pancreas Molecular Timeline | |||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||
Spleen
| Feature | ||
|---|---|---|
| Carnegie stage 14 to 17
appears as a bulge in the dorsal mesogastrium. Mesothelium pseudostratified. | ||
| Mesothelium (pseudostratified} replaced with high columnar cells and then low columnar cells. | ||
| Basement membrane present after this stage. | ||
| Hematopoietic cells detected. | ||
| Spleen is now apparent. Mesenchymal cells differentiated from cells in dorsal mesogastrium. Sinus and hilus formation after this stage. | ||
| Arteries and veins parallel entries at this stage. | ||
| Feature | ||
| reticulum cells (alpha-smooth muscle actin-positive) scattered around the arterioles | ||
| reticulum cells increase in number and began to form a reticular framework. Accumulation of T and B lymphocytes within this framework, and a primitive white pulp was observed around the arterioles. | ||
| reticular framework antigenic diversity, and T and B lymphocytes were segregated in the framework. T lymphocytes were sorted into the alpha-SMA-positive reticular framework, and the periarteriolar lymphoid sheath (PALS) was formed around the arteriole. B lymphocytes aggregated in eccentric portions to the PALS and formed the lymph follicle (LF, reticular framework alpha-SMA-negative). | ||
| marginal zone appears in reticular framework around the white pulp. | ||
| Human embryonic data[51] and fetal data [33] Links: spleen | Kyoto Collection | human timeline | ||
Respiratory
| Respiratory Stages - Species Comparison - Stages Gestational age (days) | |||||
|---|---|---|---|---|---|
| Species | Term | Embryonic | Pseudoglandular | Canalicular | Saccular |
| human | 280 | < 42 | 52 - 112 | 112 - 168 | 168 |
| primate | 168 | < 42 | 57 - 80 | 80 - 140 | 140 |
| sheep | 150 | < 40 | 40 - 80 | 80 - 120 | 120 |
| rabbit | 32 | < 18 | 21 - 24 | 24 - 27 | 27 |
| rat | 22 | < 13 | 16 - 19 | 19 - 20 | 21 |
| mouse | 20 | < 9 | 16 | 18 | 19 |
| Data modified from[52]
Links: respiratory | Respiratory Comparison | Mouse Human Respiratory | Mouse respiratory stages | mouse | rat | rabbit | Timeline Comparisons | |||||
| Age (months) | Alveoli (million) | Respiratory Airways (million) | Generations of Airways |
| Birth | 24 | 1.5 | |
| 3 | 86 | 1.8 | |
| 3 | 77 | 2.5 | 21 |
| 3 | 73 | 2.0 | |
| 7 | 112 | 3.7 | |
| 13 | 129 | 4.5 | 22 |
| 16 | 127 | 4.7 | |
| 22 | 160 | 7.1 | |
| 48 | 257 | 7.9 | |
| 98 | 280 | 14.0 | 23 |
| Adult | 296 | 14.0 | 23 |
| Data modified from [53]
Links: respiratory | Respiratory Comparison | Mouse Human Respiratory | Mouse respiratory stages | mouse | rat | rabbit | Timeline Comparisons | |||
Genital
| Week | Fetal CRL mm | Event |
|---|---|---|
| 13 | 80 | seminal vesicles appear as lateral evaginations or out-pocketings from the lower portion of the mesonephric ducts (Wolffian ducts). At this stage their lateral diameter is always greater than the anteroposterior length. |
| 14 | 100 | sacculations or diverticula of the vesicles as evaginations of the walls of the vesicles. At this time 3 small but distinct sacculations may be counted at a given level in each vesicle. |
| 19 | 170 | dilatation of the mesonephric ducts in their lower portion, to form the ampullae of the deferent ducts. Sacculations of the ampullae in the form of slight but definite irregularities of their lumina. Each vesicle at this stage shows from 3 to 8 distinct sacculations at a given level. The lateral and antero—posterior diameters are now about equal. |
| 21 | 180 | sacculations of the mesonephric ducts have become well developed and the dilatation of the ducts to form the ampullae have assumed very nearly their proportions at birth. |
| 25 | 220 | vesicles and ampullae have assumed practically their adult form as regards their general topography and arrangement of sacculations. At a single level may be counted from~nine to twelve separate diverticula. |
| 25 to 31 | 220 to 275 | prostatic utricle opens into the posterior urethra. At the latter date has a bifid lumen showing the union of two partially fused tubes. At this stage 5 to 7 distinct sacculations in each vesicle at a given level. |
| Birth | vesicles show at a given level 7 distinct sacculations at one time. At this stage many of the diverticula are traversed by a network of fine trabeculae. | |
| Data[54] Links: seminal vesicle | timeline | ||
Ovary
| Time | Carnegie Stage | Event |
|---|---|---|
| 24 days | 11 | intermediate mesoderm, pronephros primordium |
| 28 days | 12 | mesonephros and mesonephric duct |
| 35 days | 14 | uteric bud, metanephros, urogenital ridge |
| 42 days | 17 | cloacal divison, gonadal primordium (indifferent) |
| 49 days | 19 | paramesonephric duct, gonadal differentiation |
| 6-7 weeks (GA 8-9 weeks) | primordial germ cell mitosis proliferation commences | |
| 56 days | 23 | paramesonephric duct fusion (female) |
| 12-14 weeks (GA 14-16 weeks) | fetal | primordial germ cell meiosis germ cell differentiation, formation of syncitial clusters of oogonia |
| 15-18 weeks (GA 17–20 weeks) | fetal | breakdown of syncitial clusters and assembly of primordial follicles |
| Links: ovary | oocyte | timeline | Category:Timeline | ||
Human Ovary Follicle Number Timeline
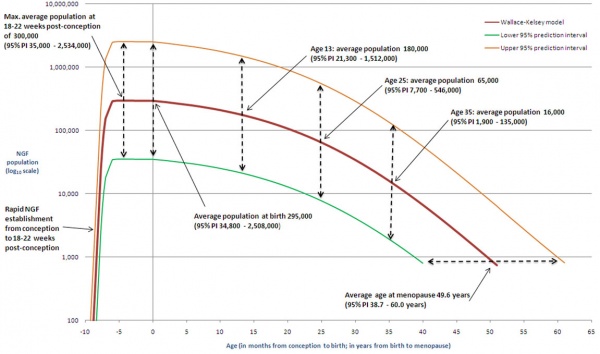
Bone
| Carnegie Stage | Event |
|---|---|
| 17 | chondrogenic progenitor of the humerus and the medial border of the scapula can be observed. |
| 18 | chondrogenic progenitor for rest of the scapula appears. |
| 19 | glenohumeral joint will begin delaminating and showing a looser central band (interzone). Denser lateral bands will join the humeral head (caput humeri) and the margins of the articular surface of the scapula, thus forming the glenoid labrum (glenoid ligament). |
| 21 | long head of the biceps tendon present |
| 22 | glenoid labrum (glenoid ligament) present |
| 23 | coracohumeral ligament present |
| Week | |
| Fetal Week 10 | osteogenic process begins in the humeral head. Primitive glenohumeral ligament present |
| Fetal Week 11 | osteogenic process begins in the scapula |
| Links: shoulder | joint | limb | timeline Data from human histological study.[55] | |
| Carnegie Stage | Event |
|---|---|
| 18 | chondrification centres of the ilium, ischium, and pubis first appears. Located around the acetabulum and grew radially. |
| 20 | iliac crest formed while the iliac body's central part remained chondrified. |
| 22 | Sacroiliac joint forms. |
| 22 | Iliac body is discoid. The growth rate was greater in the ilium than in the sacrum-coccyx, pubis, and ischium. |
| 23 | Articulation of the pubic symphysis, connection of the articular column in the sacrum, and Y-shape connection of the three parts of the hip bones to the acetabulum. |
| Early Fetal | connection of the ischium and pubic ramus. |
| Data from human CT and MRI study.[56] | |
| Links: pelvis | pelvis timeline | joint | limb | timeline | |
| Bone | Centres | Time of appearance of centre | Union of primary and secondary centres; remarks. |
|---|---|---|---|
| Clavicle | Diaphysis | 6th week | There are two centres in the shaft, a medial and a lateral. These blend on the 45th day (Mall). Shaft and epiphysis unite between the 20th and 25th years. |
| Sternal epiphysis | 18th to 20th year | ||
| Scapula | Primary centres: | The chief centre appears near the lateral angle. The subcoracoid centre appears at the base of the coracoid process and also gives rise to a part of the superior margin of the glenoid fossa. The coracoid process joins the body about the age of puberty. The acromial epiphysis centres (two or three in number) fuse with one another soon after their appearance and with the spine between the 22nd and 25th years (Quain); 20th year (Wilms). The subcoracoid and the epiphysis of the coracoid process, the glenoid fossa, the inferior angle, and the vertebral margin join between the 18th and 24th years in the order mentioned (Sappey). | |
| 1. That of the body, the spine, and the base of the glenoid cavity. | 8th week (Mall) 1 | ||
| 2. Goraooid process | 1st year | ||
| 3. Subcoracoid | 10th to 12th year | ||
| Epiphyses: | |||
| Acromial epiphyses | 15th to 18th year | ||
| Epiphysis of the inferior angle. | 16 to 18th year | ||
| Epiphyses of the vertebral border. | 18th to 20th year | ||
| Epiphyses of upper surface of coracoid. | 16th to 18th year. | ||
| Epiphysis of surface of glenoid fossa. | 16th to 18th year. | ||
| Humerus | Diaphysis | 6th to 7th week (Mall) | The epiphyses of the head, the tuberculum majus and the tuberculum minus (the last is inconstant) unite with one another in 4th-6th year and with the shaft in 20th-25th year. The epiphyses of the capitulum, lateral epicondyle, and trochlea unite with one another and then in the 16th-17th year join the shaft. The epiphysis of the medial epicondyle joins the shaft in the 18th year. |
| Epiphyses: | |||
| Head | 1st to 2d year | ||
| Tuberculum majus | 2d to 3d year | ||
| Tuberculum minus | 3d to 5th year | ||
| Capitulum | 2d to 3d year | ||
| Epioondylus med | 5th to 8th year | ||
| Lateral margin of trochlea | 11th to 12th year | ||
| Epicondylus lat | 12th to 14th year | ||
| Radius | Diaphysis | 7th week (Mall) | The superior epiphysis and shaft unite between the 17th and 20th years. The inferior epiphysis and shaft about the 21st year (Pryor); M 21st year, F 21st-25th year (Sappey). Sometimes an epiphysis is found m the tuberosity (R. and K.) and in the styloid process (Sappey). |
| Epiphyses: | |||
| Carpal end | F 8th month - M 15th month (Pryor) | ||
| Humeral end | 6th-7th year | ||
| Ulna | Diaphysis | 7th week | The centre for the shaft of the ulna arises a few days later than that for the radius. The proximal epiphysis is united to the shaft about the 17th year; the inferior epiphysis between the 18th and 20th years; F 20th - 21st years, M 21st - 24th years (Sappey). There is sometimes an epiphysis in the styloid process (Sohwegel) and in the tip of the olecranon process (Sappey). |
| Epiphyses: | |||
| Carpal end | F 6th-7th year - M 7th-8th year (Pryor) | ||
| Humeral end | 10th year | ||
| Carpus | Os capitatum | F 3d-6th month M 4th-10th month | The navicular sometimes has two centres of ossification (Serres. Rambaud and Renault). Serres and Pryor have described two centres of ossification in the lunatum. Debierre has described two centres in the pisiform, one in a girl of eleven, the other in a boy of twelve. The OS hamatum may have a special centre for the hamular process. Pryor has found two centres in the triquetrum. Pryor (1908), describes the centres of ossification of the carpal bones as assuming shapes characteristic of each bone at an early period. |
| Os hamatum | F 5th-10th month M 6th-12th month | ||
| Os triquetrum | F 2d-3d year M about 3 years | ||
| Os lunatum | F 3rd-4th year M about 4 years | ||
| Os naviculare | F at 4 years, or early in 5th year M about 5 years | ||
| Os mult. maj. | F 4th-5th year M 5th-6th year | ||
| Osmult. min. | F 4th-5th year M 6th-6th year | ||
| Os pisiforme | F 9th-10th year M 12th-3th year | ||
| Metacarpals | Diaphyses | 9th week (Mall) | The centres for the shafts of the second and third metacarpals are the first to appear. There may be a distal epiphysis for the first metacarpal and a proximal epiphysis for the second. Pryor (1906). found the distal epiphysis of the first metacarpal in about 6 per cent, of cases. It is a family characteristic. It arises before the 4th year and unites later. Pryor found the proximal epiphysis of the second metacarpal in six out of two hundred families. It unites with the shaft between the 4th and 6th-7th year; sometimes, however, not until the 14th year. In the seal and some other animals all the metacarpals have proximal and distal epiphyses (Quain). The epiphyses join the shafts between the 15th and 20th years. There may bean independent epiphysis for the styloid process of the 5th metacarpal. The epiphysis of the metacarpal of the index finger appears first. This is followed by those of the 3d, 4th, 5th, and 1st digits. |
| Proximal epiphysis of the first metacarpal | 3d year | ||
| Distal epiphyses of the metacarpals | 2d year | ||
| Phalanges | Diaphyses | 9th week (Mall) | |
| First row | Proximal epiphyses | 1st-3rd year (Pryor) | The shafts of the phalanges of the second and third fingers are the first to show centres of ossification. The phalanges of the little finger are the last, the epiphysis in the middle finger is the first to appear. This is followed by those of the 4th, 2d, 5th, and 1st digits. |
| Middle row | Diaphyses | 11th-12th week (Mall) | The centres in the shafts of this row are the last to appear. The epiphysis of the phalanx of the middle finger is the first to appear. This is followed by those of the ring, index, and little finger (Pryor). |
| Proximal epiphyses | 2nd-3rd year | ||
| Terminal row | Diaphyses | 7th-8th week | The terminal phalanx of the thumb is the first to show a centre of ossification in the shaft. This is the first centre of ossification in the hand. It is developed in connective tissue while the centres of the other phalanges are developed in cartilage (Mall). The epiphysis of the ungual phalanx of the thumb is followed by those of the middle, ring, index, and little fingers. The fusion of the epiphyses of the phalanges with the diaphyses takes place in the 18th-20th year. |
| Proximal epiphyses | 2nd-3rd year | ||
| Sesamoid bones | Ossification begins generally in the 13th - 14th years, and may not take place until after middle life (Thilenius). For table of relative frequency in the embryo and adult see p. 385. | ||
| Days and weeks refer to the prenatal, years to the postnatal period. M = male F = female. According to Poirier, Traite d'Anatomie, p. 138, two centres appear in the eighth week, and unite in the third month to form a centre of ossification for the body of the scapula. | |||
| Links: limb | bone | upper limb ossification timeline | lower limb ossification timeline | Historic - Chapter 11 Development of the Skeleton | timeline | Category:Timeline Table Data Reference[57] | |||
| Bone | Centres | Time of appearance of centre | Time of fusion: general remarks |
|---|---|---|---|
| Os coxae | Os ilium | 56th day (Mall) | The rami of the ischium and the pubis are united by bone in the 7th or 8th year (Quain) ( 12-14 year Sappey). In the acetabulum the three hip bones are separated by a Y-shaped cartilage until after puberty. In this cartilage between the ilium and pubis the "os acetabuli" appears between the ninth and twelfth years. This bone, variable in size, forms a greater or less part of the pubic portion of the articular cavity. Leche (1884). Krause (1885), and many others consider it primarily an independent bone. About puberty between the ilium and ischium and over the acetabular surfaces of these bones small irregular epiphyseal centres appear. The os acetabuli becomes imited to the pubic bone about puberty and soon afterwards the acetabular portions of the ilium and ischium and the ischium and pubis begin to become united by bone. The acetabular portions of the pubis and ilium are unite a little later. Osseous union takes place earlier on the pelvic than on the articular surface of the acetabulum. The union of the several primary centres and the epiphyses is usually completed about the twentieth year. |
| Os ischii | 105th day (Mall) | ||
| Os pubis | 4th to 5th fetal month | ||
| Os acetabuli. | 9th to 12th year | ||
| Epiphyses:
Those of the acetabulum |
Soon after puberty | ||
| Crest of ilium | Soon after puberty | Fuses with main bone 20th to 25th year | |
| Tuberosity of ischium | Soon after puberty | Fusion begins in the 17th year and is completed between the 20th and 24th years (Sappey) | |
| Ischial spine | Soon after puberty | 18th to 20th year (Poirier). | |
| Ant. inf. spine of ilium | Soon after puberty | 18th to 20th year (Poirier) | |
| Symphysis end of os pubis (1 or 2 centres) | 18th to 20th year (Sappey) | After the 20th year | |
| Femur | Diaphysis | 43d day (Mall) | |
| Epiphyses:
Distal end |
Shortly before birth1 | 20th to 24th year | |
| Head | 1st year | 18th to 19th year | |
| Great trochanter | 3d to 4th year (Osseous granules soon after birth, (Poirier) | 18th year | |
| Small trochanter | 13th to 14th year
8th year (Sappey) |
17th year (Quain)
Proximal epiphysis 18th to 22d year (Poirier) | |
| Patella | 3d to 5th year | The osseous patella reaches its definitive form soon before puberty | |
| Tibia | Diaphysis | 44th day (Mall) | |
| Epiphyses:
Proximal end |
About birth | 19th to 24th year (Sappey) | |
| Distal end | 2d year | 16th to 19th year | |
| Tubercle (occas.) | 13th year | Fuses with epiphysis of the proximal end and then with this to the diaphysis | |
| Fibula | Diaphysis | 55th day (Mall). | |
| Epiphyses:
Distal end |
2d year | 20th to 22d year | |
| Proximal end | 3d to 5th year | 22d to 24th year | |
| Calcaneus | Chief centre | 6th fetal month | The chief nucleus is endochondral. A periosteal nucleus appears frequently in the 4-5 fetal month (Hasselwander) |
| Epiphysis (distal end) | 10th year (Quain)
7th-8th year ( Sappey) |
15th-16th year (Quain)
16th-18th year (Poirier) M 17-21, average 20 years F 13-17, average 16 years (Hasselwander) | |
| Talus | 6th fetal month (Hasselwander) | In the 7th-8th year the posterior part of the talus, the os trigonum, is frequently ossified from a special centre (v. Bardeleben). It fuses about the 18th year. | |
| Cuboid | About birth | ||
| Cuneiform III | 1st year | ||
| Cuneiform I | 2d-3d year | ||
| Cuneiform II | 3d-4th year | ||
| Navicular | 4th-5th year | ||
| Metatarsals | Diaphyses | 8th-10th week | According to v. Bardeleben a second centre of ossincation appears much later than the primary in the navicular, and finally about the time of puberty a medial epiphyseal centre arises. |
| Epiphyses | 3d-8th year | The centre for the 2d metatarsal usually appears first, then come the 3rd, 4th, 1st and 5th. The epiphysis of the 1st metatarsal appears at the proximal end of the bone: the other epiphyses arise at the distal ends of the metatarsals. There may be a distal epiphysis in the first metatarsal also.2 In some instances a proximal epiphysis is formed on the tuberosity of the fifth metatarsal (Gruber). The epiphyses unite with the shafts in the 17-21 year in males and in the 14-19 year in females. (Hasselwander). | |
| Phalanges: | |||
| Terminal row | Diaphyses | 58th day (Mall) | |
| Epiphyses (distal) | 4th year | M 13-23, average 16-21 year.
F 13-17, average 14-17 year (Hasselwander). | |
| Middle row | Diaphyses | 4th-10th fetal month | |
| Epiphyses | 3d year | M 15-19 year
F 13-16 year (Hasselwander) | |
| Proximal row | Diaphyses | 3d fetal month | |
| Epiphyses | 3d year | M 15-17 year.
F 14-15 year (Hasselwander) The centres for the shafts of the phalanges often appear double, one for the dorsal and one for the plantar surface. The centres for the medial phalanges in each row usually appear before the more laterally placed centres. The centre for the 5th terminal phalanx appears much later than the other centres in this row (Mall). According to Rambaud and Renault the epiphyses arise each from two centres which fuse together. In the terminal phalanx of the great toe the ossification centre of the epiphysis often appears as early as the second or even the first year. (Hasselwander) | |
| Sesamoid bones of the great toe | M 14th year
F 12th-13th year |
Ossification may begin in the 8th year in females, in the 11th in males (Hasselwander). | |
| |||
| Days and weeks refer to the prenatal, years to the postnatal period. M = male F = female. According to Poirier, Traite d'Anatomie, p. 138, two centres appear in the eighth week, and unite in the third month to form a centre of ossification for the body of the scapula. | |||
| Links: limb | bone | upper limb ossification timeline | lower limb ossification timeline | Historic - Chapter 11 Development of the Skeleton | timeline | Category:Timeline Table Data Reference[57] | |||
| Before birth: | ||
| Both sexes | Appearance | Head of humerus, distal femur, proximal tibia, calcaneum, talus |
| Female | Appearance | Cuboid |
| During first year: | ||
| Both sexes | Appearance | Hamate, capitate, head of femur, third cuneiform |
| Female | Appearance | Capitulum, distal radius, distal tibia, distal fibula |
| Male | Appearance | Cuboid |
| During second year: | ||
| Both sexes | Appearance | Proximal phalanges of inner four fingers |
| Female | Appearance | First metacarpal, distal phalanges of thumb, middle and ring fingers,
tarsal navicular, first and second cuneiforms |
| Male | Appearance | Capitulum, distal epiphysis of radius, distal fibula |
| At age of 2: | ||
| Both sexes | Appearance | Innerfourmetacarpals,firstmetatarsal,proximalphalangesoftoes, distal phalanx of hallux |
| Female | Appearance | Proximal phalanx of thumb, middle row of phalanges of fingers |
| Male | Appearance | First metacarpal, distal phalanx of thumb, and distal phalanx of
index,firstcuneiform |
| At age of 3: | ||
| Female | Appearance | Patella, proximal fibula, second metatarsal, third metatarsal, middle phalangesof second, third and fourth toes, distal phalanges of third and fourth toes |
| Male | Appearance | Triquetrum, proximal phalanx of thumb, middle phalanges of middle and ring fingers, tarsal navicular, second cuneiform |
| At age of 4: | ||
| Both sexes | Appearance | Fourth metatarsal |
| Female | Appearance Fusion |
Head of radius, fifth metatarsal Greater tubercle to head of humerus |
| Male | Appearance | Lunate, middle phalanges of index and litle fingers, distal phalanges
of middle and ring fingers, second metatarsal, third metatarsal, middle phalanx of second toe |
| At age of 5: | ||
| Both sexes | Appearance | Navicular (carpal), multangulum majus, greater trochanter, distal
phalanx of second toe |
| Female | Appearance | Medial epicondyle, distal ulna, lunate, triquetrum, multangulum
minus, distal phalanx of index |
| Male | Appearance |
Head of radius, distal phalanx of litle finger, patella, proximal
fibula, fifth metatarsal, middle phalanges of third and fourth toes, distal phalanges of third and fourth toes |
| Fusion | Greater tubercle to head of humerus | |
| At age of 6: | ||
| Male | Appearance | Medial epicondyle, distal ulna, multangulum minus |
| At age of 7: | ||
| Female | Appearance | Distal phalanx of litle finger |
| Fusion | Rami ofischium and pubis | |
| At age of 8: | ||
| Both sexes | Appearance | Apophysis of calcaneus |
| Female | Appearance | Olecranon |
| At age of 9: | ||
| Female | Appearance | Trochlea, pisiform |
| Male | Fusion | Rami of ischium and pubis |
| At age of 10: | ||
| Male | Appearance | Trochlea, olecranon |
| At age of 11: | ||
| Female | Appearance | Lateral epicondyle |
| Male | Appearance | Pisiform |
| At age of 12: | ||
| Male | Appearance | Lateral epicondyle |
| At age of 13: | ||
| Female | Appearance | Proximal sesamoid of thumb |
| Fusion | Lower conjoint epiphysis of humerus, distal phalanx of thumb, bodies ilium, ischium and pubis | |
| Male | Fusion | Capitulum to trochlea and lateral epicondyle |
| At age of 14: | ||
| Female | Appearance | Acromion, iliac crest, lesser trochanter |
| Fusion | Olecranon, upper radius, proximal phalanx of ring finger, distal phalanx of thumb, head of femur, greater trochanter, distal tibia and fibula, apophysis calcaneus, first metatarsal, proximal phalanges oftoes | |
| Male | Appearance | Proximal sesamoid of thumb, base of fifth metatarsal |
| At age of 15: | ||
| Both sexes | Appearance | Sesamoid of little finger |
| Fusion | Distal phalanges of second, third and fourth toes | |
| Female | Appearance | Sesamoid of index and little fingers |
| Fusion | Medial epicondyle, first metacarpal, proximal phalanx of thumb, distal phalanges of inner four fingers, proximal tibia, outer four metatarsals, middle phalanx of second toe, distal phalanges of innerfourtoes | |
| Male | Appearance | Acromion |
| Fusion | Ilium, ischium and pubis | |
| At age of 16: | ||
| Female | Appearance | Distal sesamoid of thumb, tuber ischii |
| Fusion | Inner four metacarpals, proximal phalanges of index, middle and litle fingers, middle phalanges of fingers | |
| Male | Fusion | Lower conjoint epiphysis of humerus, medial epicondyle, olecranon, head of radius, distal phalanx of middle finger, apophysis of calcaneus |
| At age of 17: | ||
| Both sexes | Fusion | Acromion |
| Female | Fusion | Upper conjoint epiphysis of humerus, distal ulna, distal femur, proximal fibula |
| Male | Appearance | Distalsesamoidofthumb |
| Fusion | First metacarpal, proximal phalanges of thumb and ring finger, middle phalanges, index, middle and ring fingers, distal phalanges of thumb, index, ring and little fingers, head of femur, greater trochanter,distal tibia and fibula, metatarsals, proximal phalanges of toes, middle phalanx of second toe, distal phalanx of hallux | |
| At age of 18: | ||
| Female | Fusion | Distal radius |
| Male | Fusion | Inner four metacarpals, proximal phalanges of index, middle and litle fingers, middle phalanges of litle finger, proximal tibia |
| At age of 19: | ||
| Male | Appearance | Sesamoid of index, tuber ischii |
| Fusion | upper conjoint epiphysis of humerus, distal radius and ulna, distal femur, proximal fibula | |
| At age of 20: | ||
| Both sexes | Fusion | Iliac crest |
| Male | Fusion | Tuber ischii |
| At age of 21: | ||
| Both sexes | Appearance | Clavicle |
| Female | Fusion | Tuber ischii |
| At age of 22: | ||
| Both sexes | Fusion | Clavicle |
| Table Data: Flecker (1932)[58] | Original table | bone timeline | ||
| Bone - Appearance and Fusion of Epiphyses Timeline | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Muscle
| Carnegie Stage | Day | Mouse | Event |
|---|---|---|---|
| 14 | 33 | E10.5 | mesoderm of primary body wall non-compact, coalesced in the ventral midline to create the abdominal cavity. Liver and stomach present. Dermomyotomes that are derived from somites have been formed. |
| 16 | 40 | Migration distance about 25% of the hemicircumference of the abdominal cavity. Lateral plate mesoderm has become more condensed and thicker in the area around the myoblasts. Primary abdominal wall ventral to this region was thinner and less dense. Suggests both myoblasts and connective tissue may migrate into the primary body wall or active cell proliferation. | |
| 17 | 42 | E11.5 | cells now migrated about 50% of the distance to the ventral midline. Inner and outer layers not yet not discernible. |
| 18 | 44 | E12.5 | Separation of myoblasts into distinct inner and outer layers. Myoblasts in both inner and outer layers began to exhibit unidirectional orientation. Abdominal wall thicker (500 μm) in region where secondary structures forming compared with primary body wall region (260 μm). More dorsally positioned regions, outermost layer of connective tissue comprised approximately half of this thickness. |
| 19 | 48 | Segregation of the myoblasts into four distinct muscle groups with unidirectional orientation of myoblasts. Myoblasts migrated over half of the distance to the ventral midline. Abdominal wall remains thickest in the area where the muscles migrated and again the outermost layer of connective tissue comprises approximately half of the total thickness of the abdominal wall in this region. Primary abdominal wall that is ventral to the migrating myoblasts noticeably thinner. Human rectus completely separated after migrating over half the distance to the midline. In mouse, the rectus is not segregated from the other muscles until reaching the midline. | |
| 21 | 54 | E14.5 | Myoblasts have reached the ventral midline and myotubes were present and oriented uniformly within all muscle groups. Rectus abdominis formed distinct bundles of muscle indicating that development and differentiation of this muscle were more prominent in humans than in mice. Connective tissue layers form majority of the thickness of the abdominal wall, outermost layer of connective tissue majority of thickness. |
| 23 | 58 | E15.5 | Rectus muscle forms 2 or 3 distinct layers and myotube orientation remained uniform in all muscles. External oblique and internal oblique expand in terms of thickness. Transversus remained a thin layer of muscle. Thickness of the connective tissue was reduced. The orientation of connective tissue layers in the obliques and transversus abdominis was dorsal to ventral. |
| Table Data[9] | |||
For additional muscle timelines see the page - Muscle Development Timeline
Adipose
| Anatomical Region | Specific Location | Start (weeks) |
CRL (mm) | Complete (weeks) |
CRL (mm) |
|---|---|---|---|---|---|
| Head | Buccal pad | 14 | 100 | 17 | 153 |
| Cheek | 14.5 | 103 | 17 | 150 | |
| Chin | 14.5 | 103 | 17 | 150 | |
| Ocular pad | 15 | 113 | 19.5 | 170 | |
| Neck | Neck | 15 | 113 | 19.5 | 170 |
| Thorax | Anterior wall | 16 | 135 | 19.5 | 170 |
| Posterior wall | 15 | 113 | 20.5 | 190 | |
| Mammary | 14.5 | 106 | 17.5 | 156 | |
| Abdomen | Abdominal wall | 14.5 | 106 | 20.5 | 190 |
| Perirenal | 15 | 113 | 20.5 | 190 | |
| Upper limb | Shoulder | 15 | 113 | 23.5 | 216 |
| Forearm | 16 | 131 | 20.5 | 190 | |
| Arm | 16 | 131 | 20.5 | 190 | |
| Hand | 16 | 131 | l9.5 | 172 | |
| Lower limb | Gluteal | 16 | 131 | 20.5 | 190 |
| Thigh | 16.5 | 141 | 22.5 | 212 | |
| Leg | 16 | 131 | 22.5 | 212 | |
| Foot | 16 | 131 | 19.5 | 170 | |
| Table Notes - weeks are fertilization age (FA), not GA, both male and female data are combined. Table Data source[59] Links: adipose | Second Trimester | |||||
Nerve Endings
| Striped muscle and tendon | Sensory nerve endings | Motor nerve endings | Human (approximate age) weeks | Chick (Tello) (approximate age) days |
|---|---|---|---|---|
| Beginning differentiation | — | — | 7 | 5 |
| Myofibrils appear | Exploratory fibres with bulbous endings penetrate into muscles | 8 | 6 | |
| Great increase of connective tissue. Vast numbers of nuclei | - | - | - | 6-7 |
| Isolated “fat” fibres | - | — | 10-12 | 7 |
| Varicose endings in contact with myotubes | - | 10 | — | |
| Tendon fibres diflerentiate | — | — | 12 | 10 |
| Beginning pacinian corpuscle | — | 20 | 11 | |
| Multiplication of myotubes | — | - | 12-22 | 9-13 |
| Spindles differentiate | - | 20 | 12 | |
| Dissociation of myotubes from one another. Nuclei become peripheral | Spindles become complex | — | 22-26 | 13 |
| pacinian corpuscle well developed | — | 26-28 | 15 | |
| Plates begin to differentiate | 26-28 (very rudimentary except tongue) | 18 | ||
| Data: Hewer (1935)[35] (human weeks appear to be fertilization age based on CRL) Links: timeline | chicken | skeletal muscle | ||||
Tongue
| Fertilisation Age (weeks) |
Gestational Age GA (weeks) |
Mean Circumference (mm) |
|---|---|---|
| 12 | 14 | 28 |
| 13 | 15 | 33 |
| 14 | 16 | 36 |
| 15 | 17 | 37 |
| 16 | 18 | 43 |
| 17 | 19 | 48 |
| 18 | 20 | 51 |
| 19 | 21 | 55 |
| 20 | 22 | 58 |
| 21 | 23 | 62 |
| 22 | 24 | 64 |
| 23 | 25 | 70 |
| 24 | 26 | 73 |
| Table data[60] Tongue circumference measured by transvaginal ultrasonography between 14 and 17 GA weeks, and by abdominal ultrasound between 18 and 26 GA weeks of gestation. Links: tongue | fetal | ultrasound | ||
Tooth
| Milk Dentition | |
|---|---|
| Median incisors | 6th to 8th month |
| Lateral incisors | 8th to 12th month |
| First molars | 12th to 16th month |
| Canines 1 | 7th to 20th month |
| Second molars | 20th to 24th month |
| Permanent Dentition | |
| First molars | 7th year |
| Median incisors | 8th year |
| Lateral incisors | 9th year |
| First premolars | 10th year |
| Second premolars | 11th year |
| Canines | 13th to 14th year |
| Second molars | 13th to 14th year |
| Third molars | 17th to 40th year |
| Approximate timings only, vary considerably according to racial, climatic, and nutritive conditions. Data source: Keibel and Mall (1912)[61] The teeth Links: tooth | |
Integumentary
| Carnegie Stage | Event |
|---|---|
| 10 | optic primordia appear |
| 13 | By the end of the fourth week the optic vesicle lies close to the surface ectoderm. The surface ectoderm overlying the optic vesicle, in response to this contact, has thickened to form the lense placode. |
| 14 | (about 32 days) the lens placode is indented by the lens pit. |
| 15 | (about 33 days) the lens pit is closed. The lens vesicle and optic cup lie close to the surface ectoderm and appear to press against the surface. |
| 16 | (37 days) Prior to the development of the eyelids, one small sulcus or groove forms above the eye (eyelid groove) and another below it. |
| 17 - 19 | grooves deepen, eyelid folds develop, first below, and then above, the eye. |
| 19 - 22 | eyelid folds develop into the eyelids and cover more of the eye as the palpebral fissure takes shape. The upper and the lower eyelids meet at the outer canthus in Stage 19. |
| 20 | the inner canthus is established. |
| 23 | closure of the eyelids is complete (Note - shown as still open in some Kyoto embryo). |
| Data - from Kyoto embryos with Carnegie staging.[7] Links: eyelid |
References
- ↑ 1.0 1.1 1.2 1.3 1.4 O'Rahilly R & Müller F. (2007). The development of the neural crest in the human. J. Anat. , 211, 335-51. PMID: 17848161 DOI.
- ↑ 2.0 2.1 O'Rahilly R & Müller F. (1990). Ventricular system and choroid plexuses of the human brain during the embryonic period proper. Am. J. Anat. , 189, 285-302. PMID: 2285038 DOI.
- ↑ 3.0 3.1 3.2 3.3 3.4 3.5 Godlewski G, Gaubert-Cristol R, Rouy S & Prudhomme M. (1997). Liver development in the rat and in man during the embryonic period (Carnegie stages 11-23). Microsc. Res. Tech. , 39, 314-27. PMID: 9407542 <314::AID-JEMT2>3.0.CO;2-H DOI.
- ↑ Müller F & O'Rahilly R. (1988). The development of the human brain from a closed neural tube at stage 13. Anat. Embryol. , 177, 203-24. PMID: 3354839
- ↑ 5.0 5.1 5.2 5.3 5.4 5.5 5.6 Menshawi K, Mohr JP & Gutierrez J. (2015). A Functional Perspective on the Embryology and Anatomy of the Cerebral Blood Supply. J Stroke , 17, 144-58. PMID: 26060802 DOI.
- ↑ 6.0 6.1 6.2 6.3 Müller F & O'Rahilly R. (2004). Olfactory structures in staged human embryos. Cells Tissues Organs (Print) , 178, 93-116. PMID: 15604533 DOI.
- ↑ 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 7.8 7.9 Pearson AA. (1980). The development of the eyelids. Part I. External features. J. Anat. , 130, 33-42. PMID: 7364662
- ↑ 8.0 8.1 Clugston RD, Zhang W & Greer JJ. (2010). Early development of the primordial mammalian diaphragm and cellular mechanisms of nitrofen-induced congenital diaphragmatic hernia. Birth Defects Res. Part A Clin. Mol. Teratol. , 88, 15-24. PMID: 19711422 DOI.
- ↑ 9.0 9.1 9.2 9.3 9.4 9.5 9.6 9.7 Nichol PF, Corliss RF, Yamada S, Shiota K & Saijoh Y. (2012). Muscle patterning in mouse and human abdominal wall development and omphalocele specimens of humans. Anat Rec (Hoboken) , 295, 2129-40. PMID: 22976993 DOI.
- ↑ Müller F & O'Rahilly R. (1988). The first appearance of the future cerebral hemispheres in the human embryo at stage 14. Anat. Embryol. , 177, 495-511. PMID: 3377191
- ↑ Patelska-Banaszewska M & Woźniak W. (2005). The subarachnoid space develops early in the human embryonic period. Folia Morphol. (Warsz) , 64, 212-6. PMID: 16228957
- ↑ Müller F & O'Rahilly R. (1988). The development of the human brain, including the longitudinal zoning in the diencephalon at stage 15. Anat. Embryol. , 179, 55-71. PMID: 3213956
- ↑ Müller F & O'Rahilly R. (1989). The human brain at stage 16, including the initial evagination of the neurohypophysis. Anat. Embryol. , 179, 551-69. PMID: 2751117
- ↑ Müller F & O'Rahilly R. (1989). The human brain at stage 17, including the appearance of the future olfactory bulb and the first amygdaloid nuclei. Anat. Embryol. , 180, 353-69. PMID: 2802187
- ↑ 15.0 15.1 Patelska-Banaszewska M & Woźniak W. (2004). The development of the epidural space in human embryos. Folia Morphol. (Warsz) , 63, 273-9. PMID: 15478101
- ↑ 16.0 16.1 16.2 16.3 Hashimoto R. (2003). Development of the human Müllerian duct in the sexually undifferentiated stage. Anat Rec A Discov Mol Cell Evol Biol , 272, 514-9. PMID: 12740945 DOI.
- ↑ Keibel F. and Mall FP. Manual of Human Embryology II. (1912) J. B. Lippincott Company, Philadelphia.
- ↑ Congdon ED. Transformation of the aortic-arch system during the development of the human embryo. (1922) Contrib. Embryol., Carnegie Inst. Wash. Publ 277, 14:47-110.
- ↑ Teal SI., Moore GW. and Hutchins GM. Development of aortic and mitral valve continuity in the human embryonic heart. (1986) Amer. J. Anat., 176:447-460.
- ↑ Wells LJ. Development of the human diaphragm and pleural sacs. (1954) Contrib. Embryol., Carnegie Inst. Wash. Publ. 603, 35: 107-134.
- ↑ Jirásek JE. Development of the Genital System and Male Pseudohermaphroditism. (1971) Johns Hopkins Press, Baltimore.
- ↑ Wilson KM. Origin and development of the rete ovarii and the rete testis in the human embryo. (1926) Carnegie Instn. Wash. Publ. 362, Contrib. Embryol., Carnegie Inst. Wash., 17:69-88.
- ↑ Gasser RL. Atlas of Human Embryos. (1975) Harper & Row, Hagerstown, Maryland.
- ↑ 24.0 24.1 Müller F & O'Rahilly R. (1990). The human brain at stages 18-20, including the choroid plexuses and the amygdaloid and septal nuclei. Anat. Embryol. , 182, 285-306. PMID: 2268071
- ↑ 25.0 25.1 Müller F & O'Rahilly R. (1990). The human brain at stages 21-23, with particular reference to the cerebral cortical plate and to the development of the cerebellum. Anat. Embryol. , 182, 375-400. PMID: 2252222
- ↑ Witt M & Reutter K. (1996). Embryonic and early fetal development of human taste buds: a transmission electron microscopical study. Anat. Rec. , 246, 507-23. PMID: 8955790 <507::AID-AR10>3.0.CO;2-S DOI.
- ↑ Müller F & O'Rahilly R. (1990). The human rhombencephalon at the end of the embryonic period proper. Am. J. Anat. , 189, 127-45. PMID: 2244584 DOI.
- ↑ O'Rahilly R, Muller F & Meyer DB. (1980). The human vertebral column at the end of the embryonic period proper. 1. The column as a whole. J. Anat. , 131, 565-75. PMID: 7216919
- ↑ Streeter GL. Developmental horizons in human embryos (fourth issue). A review of the histogenesis of cartilage and bone. (1949) Carnegie Instn. Wash. Publ. 583, Contrib. Embryol., 33: 149-169. PMID: 18144445
- ↑ Wünsch L & Schober JM. (2007). Imaging and examination strategies of normal male and female sex development and anatomy. Best Pract. Res. Clin. Endocrinol. Metab. , 21, 367-79. PMID: 17875485 DOI.
- ↑ 31.0 31.1 31.2 Locher H, de Groot JC, van Iperen L, Huisman MA, Frijns JH & Chuva de Sousa Lopes SM. (2015). Development of the stria vascularis and potassium regulation in the human fetal cochlea: Insights into hereditary sensorineural hearing loss. Dev Neurobiol , 75, 1219-40. PMID: 25663387 DOI.
- ↑ 32.0 32.1 32.2 32.3 32.4 Afif A, Bouvier R, Buenerd A, Trouillas J & Mertens P. (2007). Development of the human fetal insular cortex: study of the gyration from 13 to 28 gestational weeks. Brain Struct Funct , 212, 335-46. PMID: 17962979 DOI.
- ↑ 33.0 33.1 33.2 33.3 33.4 Satoh T, Sakurai E, Tada H & Masuda T. (2009). Ontogeny of reticular framework of white pulp and marginal zone in human spleen: immunohistochemical studies of fetal spleens from the 17th to 40th week of gestation. Cell Tissue Res. , 336, 287-97. PMID: 19255788 DOI.
- ↑ Kadhim HJ, Gadisseux JF & Evrard P. (1988). Topographical and cytological evolution of the glial phase during prenatal development of the human brain: histochemical and electron microscopic study. J. Neuropathol. Exp. Neurol. , 47, 166-88. PMID: 3339373
- ↑ 35.0 35.1 35.2 Hewer EE. The development of nerve endings in the human foetus. (1935) J Anat. 69(3):369-79. PMID 17104543
- ↑ 36.0 36.1 36.2 Garel C, Chantrel E, Brisse H, Elmaleh M, Luton D, Oury JF, Sebag G & Hassan M. (2001). Fetal cerebral cortex: normal gestational landmarks identified using prenatal MR imaging. AJNR Am J Neuroradiol , 22, 184-9. PMID: 11158907
- ↑ 37.0 37.1 Paquette LB, Jackson HA, Tavaré CJ, Miller DA & Panigrahy A. (2009). In utero eye development documented by fetal MR imaging. AJNR Am J Neuroradiol , 30, 1787-91. PMID: 19541779 DOI.
- ↑ 38.0 38.1 Moore JK. (2002). Maturation of human auditory cortex: implications for speech perception. Ann Otol Rhinol Laryngol Suppl , 189, 7-10. PMID: 12018354
- ↑ Evrard P. Gadisseux J.-F. and Gressens P. Hypoxia Opportunism During Brain Development. (1993) from: Report of the Workshop on Acute Perinatal Asphyxia in Term Infants, U.S. Department of Health and Human Services, Public Health Service, NIH, National Institute of Child Health and Human Development. August 30-31, 1993.
- ↑ Afif A, Trouillas J & Mertens P. (2015). Development of the sensorimotor cortex in the human fetus: a morphological description. Surg Radiol Anat , 37, 153-60. PMID: 24972575 DOI.
- ↑ de Bree K, de Bakker BS & Oostra RJ. (2018). The development of the human notochord. PLoS ONE , 13, e0205752. PMID: 30346967 DOI.
- ↑ O'Rahilly R. and Müller F. Ventricular system and choroid plexuses of the human brain during the embryonic period proper. (1990) Amer. J Anat. 189(4): 285-302. PMID 2285038
- ↑ Rodríguez-Vázquez JF, Mérida-Velasco JR & Verdugo-López S. (2010). Development of the stapedius muscle and unilateral agenesia of the tendon of the stapedius muscle in a human fetus. Anat Rec (Hoboken) , 293, 25-31. PMID: 19899117 DOI.
- ↑ Nishimura Y & Kumoi T. (1992). The embryologic development of the human external auditory meatus. Preliminary report. Acta Otolaryngol. , 112, 496-503. PMID: 1441991
- ↑ Bossy J. Development of olfactory and related structures in staged human embryos. (1980) Anat. Embryol., 161(2);225-36 PMID 7469043
- ↑ Arráez-Aybar LA, Turrero-Nogués A & Marantos-Gamarra DG. (2008). Embryonic cardiac morphometry in Carnegie stages 15-23, from the Complutense University of Madrid Institute of Embryology Human Embryo Collection. Cells Tissues Organs (Print) , 187, 211-20. PMID: 18057862 DOI.
- ↑ Lisman BA, van den Hoff MJ, Boer K, Bleker OP, van Groningen K & Exalto N. (2007). The architecture of first trimester chorionic villous vascularization: a confocal laser scanning microscopical study. Hum. Reprod. , 22, 2254-60. PMID: 17545656 DOI.
- ↑ Godlewski G, Gaubert-Cristol R, Rouy S & Prudhomme M. (1997). Liver development in the rat during the embryonic period (Carnegie stages 15-23). Acta Anat (Basel) , 160, 172-8. PMID: 9718390
- ↑ Lhuaire M, Tonnelet R, Renard Y, Piardi T, Sommacale D, Duparc F, Braun M & Labrousse M. (2015). Developmental anatomy of the liver from computerized three-dimensional reconstructions of four human embryos (from Carnegie stage 14 to 23). Ann. Anat. , 200, 105-13. PMID: 25866917 DOI.
- ↑ 50.0 50.1 50.2 50.3 Jennings RE, Berry AA, Kirkwood-Wilson R, Roberts NA, Hearn T, Salisbury RJ, Blaylock J, Piper Hanley K & Hanley NA. (2013). Development of the human pancreas from foregut to endocrine commitment. Diabetes , 62, 3514-22. PMID: 23630303 DOI.
- ↑ Endo A, Ueno S, Yamada S, Uwabe C & Takakuwa T. (2015). Morphogenesis of the spleen during the human embryonic period. Anat Rec (Hoboken) , 298, 820-6. PMID: 25403423 DOI.
- ↑ Pinkerton KE & Joad JP. (2000). The mammalian respiratory system and critical windows of exposure for children's health. Environ. Health Perspect. , 108 Suppl 3, 457-62. PMID: 10852845
- ↑ Dunnill MS. Postnatal growth of the lung. Thorax 1962;17:329–333.
- ↑ Watson EM. The development of the seminal vesicles in man. (1918) Amer. J Anat. 24(4): 395 - 439.
- ↑ Hita-Contreras F, Sánchez-Montesinos I, Martínez-Amat A, Cruz-Díaz D, Barranco RJ & Roda O. (2018). Development of the human shoulder joint during the embryonic and early fetal stages: anatomical considerations for clinical practice. J. Anat. , 232, 422-430. PMID: 29193070 DOI.
- ↑ Okumura M, Ishikawa A, Aoyama T, Yamada S, Uwabe C, Imai H, Matsuda T, Yoneyama A, Takeda T & Takakuwa T. (2017). Cartilage formation in the pelvic skeleton during the embryonic and early-fetal period. PLoS ONE , 12, e0173852. PMID: 28384153 DOI.
- ↑ 57.0 57.1 Keibel F. and Mall FP. Manual of Human Embryology I. (1910) J. B. Lippincott Company, Philadelphia.
- ↑ 58.0 58.1 Flecker H. (1932). Roentgenographic observations of the times of appearance of epiphyses and their fusion with the diaphyses. (1932) J Anat. 67: 118-164.3 PMID 17104405
- ↑ Poissonnet CM, LaVelle M & Burdi AR. (1988). Growth and development of adipose tissue. J. Pediatr. , 113, 1-9. PMID: 3290412
- ↑ Achiron R, Ben Arie A, Gabbay U, Mashiach S, Rotstein Z & Lipitz S. (1997). Development of the fetal tongue between 14 and 26 weeks of gestation: in utero ultrasonographic measurements. Ultrasound Obstet Gynecol , 9, 39-41. PMID: 9060129 DOI.
- ↑ Grosser O. Lewis FT. and McMurrich JP. The Development of the Digestive Tract and of the Organs of Respiration. (1912) chapter 17, vol. 2, in Keibel F. and Mall FP. Manual of Human Embryology II. (1912) J. B. Lippincott Company, Philadelphia.
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2026, Haziran 21) Embryology Timeline human development. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Timeline_human_development
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G