Sensory - Smell Development
| Embryology - 12 Jul 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Introduction

These notes introduce the development of the sense of smell or olfaction and the associated structures including the nasal placode, olfactory epithelium, olfactory bulb, and vomeronasal organ. Recent research has shown a relationship between what the receptive epithelium is exposed too and how the central neural pathway develops, similar to that shown earlier for the vision system. See also a review on the cell biology of smell.[1]
A French research group has recently been investigating the development of smell in the fetus and in neonates. The nasal epithelium has also been a research "hot topic" as it is one of the few easily accessible sites of adult neural stem cells.
Note the different spellings "odour" (UK) or "odor" (USA). Anosmia is the term used to describe having no sense of smell. Anosmia/hyposmia is related to the absence or hypoplasia of the olfactory bulbs and tracts.
| Smell Links: Introduction | placode | Rhinencephalon | head | respiratory | Student project | taste | sensory | Category:Smell | ||
|
| Senses Links: Introduction | placode | Hearing and Balance hearing | balance | vision | smell | taste | touch | Stage 22 | Category:Sensory |
Some Recent Findings
|
| More recent papers |
|---|
 This table allows an automated computer search of the external PubMed database using the listed "Search term" text link.
More? References | Discussion Page | Journal Searches | 2019 References | 2020 References Search term: Olfactory Development | Smell Development | Smell Embryology | nasal placode | olfactory epithelium | olfactory bulb | vomeronasal organ | accessory olfactory bulb | anosmia |
| Older papers |
|---|
| These papers originally appeared in the Some Recent Findings table, but as that list grew in length have now been shuffled down to this collapsible table.
See also the Discussion Page for other references listed by year and References on this current page.
|
Nasal Placode
(Nasal)
The olfactory epithelium develops from the paired nasal placodes, each develop initially with two components, a medial and lateral region.
Carnegie stage 13

| Carnegie stage 13 - serial sections nasal placode sections | |||
|---|---|---|---|

|

|

|

|
|
|

|

|

|
| Nasal Placode - Human Embryo stage 14 |

|
In the mouse, gonadotropin-releasing hormone-1 neurones control the release of gonadotropins from the anterior pituitary and were thought to originate from the adenohypophyseal placed. A recent study has shown that the real origin is associated with the formation of the nasal placode.[10]
- Links: placode | Search PubMed
Olfactory Epithelium
Adult - pseudostratified columnar epithelium overlying a lamina propria
Five basic cell types:
- horizonatal basal cells - not present embryonically.
- globose basal cells - transit amplifying progenitors of the olfactory epithelium.
- sustentacular cells - aligned on the surface with thin cytoplasmic projections terminating at the basal lamina.
- olfactory receptor neurons - located in an intermediate zone between basal and apical layers, form the bulk of the epithelium.
- olfactory gland cells (Bowman's gland/duct complex) - extend from the glands in the lamina propria to the ducts within the epithelium. Function to carry secretions to the apical epithelial surface.
Olfactory Ensheathing Cells
- neural crest in origin.[7]
- accompany and ensheath the small olfactory axons of the nonmyelinated olfactory nerves.
- compartmentalize the small olfactory axons into fascicles.
- allow regenerating olfactory nerves to cross the peripheral/central nerve threshold.
- Links: neural crest
Olfactory Receptors
Odours bind to and activate olfactory receptors located on the dendrites of sensory neurons in the nose and how the mitral cells of the olfactory bulb (OB) process olfactory information. What has yet to be thoroughly described is how the piriform cortex receives and transforms information arriving from the OB via the lateral olfactory tract (LOT). Although the cell types present in the piriform cortex are known (Shepherd, 2004), previous work has failed to differentiate between disparate electrophysiological profiles and synaptic contacts made between principal cells.
Timeline
| Week FA (GA) | Carnegie Stage | Event |
|---|---|---|
| week 4 (GA 6) | 11 | nasal epiblastic thickening appears |
| 12 | nasal field is well outlined | |
| week 5 (GA 7) | 15 | continuous cellulovascular strand seen between the nasal groove and the olfactory field |
| week 6 (GA 8) | 16 | vomeronasal groove appears. |
| 17 | olfactory nerve is organized into two plexuses, lateral and medial, the latter mingled with the terminal-vomeronasal complex. | |
| week 7 (GA 9) | 18 | olfactory bulb begins to appear |
| 19 | individualization of the olfactory bulb and nuclei, distinction between olfactory structures and terminal and vomeronasal ones begins to be clear. | |
| week 8 (GA 10) | 21 | structure of the olfactory bulb is evident. |
| 23 | olfactory strands are well individualized, and olfactory and terminal-vomeronasal fibers are easily distinguishable. | |
| Links: smell | sensory | timeline | Category:Timeline Table Data Reference[11] | ||
Week 5 to 8 - Stage 15, 17, 20

Development of the Human Olfactory System (Carnegie Stage 15, 17 and 20)[12]
Abbreviations: (cal) calretinin immunostaining, (CS) Carnegie stage, (HE) hematoxylin-eosin staining, (MZ) marginal zone, (OB) olfactory bulb, (OP) olfactory placode, (Tel) telencephalon.
Week 8 - Stage 22


Week 15 - Second Trimester

Human 15 weeks - terminal nerve and vomeronasal organ nerves[13]
Immunohistochemistry of S100 protein (S100) (A, C) or calretinin (cal) (D). (B) Hematoxylin and eosin staining. Panels C and D display the vomeronasal organ (VNO) and VNON in the other side of panels A and B. Intervals between panels are 0.2 mm (A–B), 0.6 mm (B–C), and 0.1 mm (C–D). In panel A, the terminal nerve (NT) runs along the posterior aspect of the cartilaginous nasal bone, while the VNON run along the palate near the VNO. Thus, these nerves are separated by a developing nasal septum. Scale bars=1 mm (A–D).
(text from figure legend)
Olfactory Pathway
| Olfactory receptor neurons (ORNs) - odours bind and activate olfactory receptors located on dendrites of sensory neurons in the nose.
Olfactory Bulb (OB) - Mitral cells process olfactory information (encoded in a chemotopic map). Lateral olfactory tract (LOT) - pathway to cortex. Primary Olfactory Cortex (= Piriform cortex) - receives and transforms information.
|

Mouse olfactory nerve pathway development[15] |
Olfactory Bulb
Vomeronasal Organ
The vomeronasal organ (VNO, vomeronasal accessory olfactory system, Jacobson's organ) is involved in detecting and transfering pheromone information through the vomeronasal nerve (nervus vomeronasalis) to the neuroendocrine hypothalamus. This signaling pathway can be used for mating and as olfactory cues for nocturnal animals. There are two families of receptors located in the vomeronasal organ the V1Rs and the V2Rs, are thought to detect pheromonal signals.[16]
|
|
Human Embryo (week 8, Carnegie stage 22)
Transverse section through the embryo head nasal region showing the developing Vomeronasal Organ (VNO) or Jacobson's organ. This contains sensory neurons that detect chemical stimuli (pheromones) Accessory olfactory system
|

|
Adult Mouse VMO anatomical position and showing also solitary chemosensory cells[17]
Human embryos also have a vomeronasal organ, though later many structures such as the vomeronasal nerve, accessory olfactory bulb and chemoreceptor cells within the organ are lost. Therefore it is not clear whether there is any postnatal neuroendocrine role for this structure.[18][19] |
Search PubMed: vomeronasal organ development | vomeronasal organ
Accessory Olfactory Bulb
The Accessory Olfactory Bulb (AOB) is the first neural integrative centre of the vomeronasal system.
Search PubMed: accessory olfactory bulb development | accessory olfactory bulb
Grueneberg Ganglion
The Grueneberg (Grüneberg) ganglion[22] "Within the nasal epithelium of mammals, there are several compartments which are populated with neuronal cells. One of them - the so-called Grueneberg ganglion - is composed of ciliated neurons residing in the anterior region of the nose."
Identified mainly in rodents, first identified in 1973 by Hans Grüneberg.
Histology
 |
| 
Animal Models
Mouse
The nasal mucosa features four separate olfactory areas:
- main olfactory epithelium (MOE)
- septal organ (SO)
- ganglion of Grüneberg (GG)
- vomeronasal sensory epithelium (VNsE) forms a part of the vomeronasal organ (VNO)
- Links: mouse
Molecular
Retinoid acid

Retinoic acid model in olfactory development[23]
- Links: retinoic acid
Abnormalities
Anosmia
Term used to describe having no sense of smell. Anosmia/hyposmia is related to the absence or hypoplasia of the olfactory bulbs and tracts.
Kallmann Syndrome
- A developmental disease affecting both the hormonal reproductive axis and the sense of smell.
- Affected individuals have mutations in either of two different genes KAL1 and FGFR1 (20%) and prokineticin receptor-2 (PROKR2) or prokineticin-2 (PROK2) genes (10%).
Choanal Atresia
[[File:Choanal atresia computed tomography 01.jpg|thumb|Choanal atresia computed tomography[24]
- Choanal atresia is the most common form of congenital nasal obstruction, usually diagnosed at birth.[24]
- failure of the posterior nasal cavity (choanae) to communicate with the nasopharynx.
- unilateral or bilateral bony membranous septum located between the nose and the pharynx.
- occurs in approximately 1 in 5000 to 7000 live births.
- Thought to be secondary to an abnormality during the rupture of the buccopharyngeal membrane in the embryological period.
Additional Images
Historic
Keith, A. (1902) Human Embryology and Morphology. London: Edward Arnold.
- 1902 The Nasal Cavities and Olfactory Structures
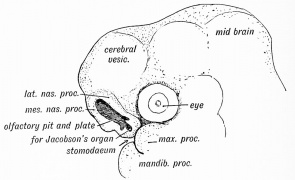
Fig. 16. Embryo Olfactory Pit and Nasal Processes (4 week)
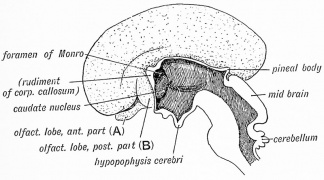
Fig. 17. Fetal Olfactory Lobe (3.5 months)
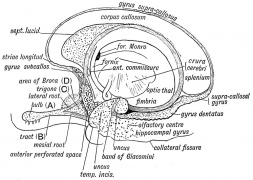
Fig. 18. Adult Olfactory Lobe
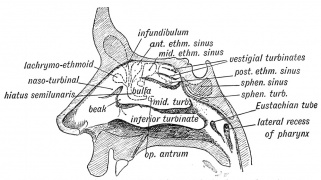
Fig. 19. Lateral Wall Nasal Cavity Air Sinus
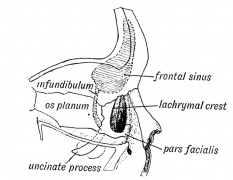
Fig. 20. Orbit and Infundibulum





References
- ↑ 1.0 1.1 DeMaria S & Ngai J. (2010). The cell biology of smell. J. Cell Biol. , 191, 443-52. PMID: 21041441 DOI.
- ↑ Nasu M, Shimamura K, Esumi S & Tamamaki N. (2020). Sequential pattern of sublayer formation in the paleocortex and neocortex. Med Mol Morphol , 53, 168-176. PMID: 32002665 DOI.
- ↑ Matsumoto M, Sawada M, García-González D, Herranz-Pérez V, Ogino T, Bang Nguyen H, Quynh Thai T, Narita K, Kumamoto N, Ugawa S, Saito Y, Takeda S, Kaneko N, Khodosevich K, Monyer H, Manuel García-Verdugo J, Ohno N & Sawamoto K. (2019). Dynamic changes in ultrastructure of the primary cilium in migrating neuroblasts in the postnatal brain. J. Neurosci. , , . PMID: 31685650 DOI.
- ↑ Sarnat HB & Flores-Sarnat L. (2019). Development of the human olfactory system. Handb Clin Neurol , 164, 29-45. PMID: 31604554 DOI.
- ↑ Oprych K, Cotfas D & Choi D. (2017). Common olfactory ensheathing glial markers in the developing human olfactory system. Brain Struct Funct , 222, 1877-1895. PMID: 27718014 DOI.
- ↑ Secundo L, Snitz K & Sobel N. (2014). The perceptual logic of smell. Curr. Opin. Neurobiol. , 25, 107-15. PMID: 24440370 DOI.
- ↑ 7.0 7.1 Katoh H, Shibata S, Fukuda K, Sato M, Satoh E, Nagoshi N, Minematsu T, Matsuzaki Y, Akazawa C, Toyama Y, Nakamura M & Okano H. (2011). The dual origin of the peripheral olfactory system: placode and neural crest. Mol Brain , 4, 34. PMID: 21943152 DOI.
- ↑ Weiss J, Pyrski M, Jacobi E, Bufe B, Willnecker V, Schick B, Zizzari P, Gossage SJ, Greer CA, Leinders-Zufall T, Woods CG, Wood JN & Zufall F. (2011). Loss-of-function mutations in sodium channel Nav1.7 cause anosmia. Nature , 472, 186-90. PMID: 21441906 DOI.
- ↑ Ohsawa S, Hamada S, Kuida K, Yoshida H, Igaki T & Miura M. (2010). Maturation of the olfactory sensory neurons by Apaf-1/caspase-9-mediated caspase activity. Proc. Natl. Acad. Sci. U.S.A. , 107, 13366-71. PMID: 20624980 DOI.
- ↑ Metz H & Wray S. (2010). Use of mutant mouse lines to investigate origin of gonadotropin-releasing hormone-1 neurons: lineage independent of the adenohypophysis. Endocrinology , 151, 766-73. PMID: 20008041 DOI.
- ↑ Bossy J. Development of olfactory and related structures in staged human embryos. (1980) Anat. Embryol., 161(2);225-36 PMID 7469043
- ↑ Antal MC, Samama B, Ghandour MS & Boehm N. (2015). Human Neural Cells Transiently Express Reelin during Olfactory Placode Development. PLoS ONE , 10, e0135710. PMID: 26270645 DOI.
- ↑ Jin ZW, Cho KH, Shibata S, Yamamoto M, Murakami G & Rodríguez-Vázquez JF. (2019). Nervus terminalis and nerves to the vomeronasal organ: a study using human fetal specimens. Anat Cell Biol , 52, 278-285. PMID: 31598357 DOI.
- ↑ Tsuji T, Tsuji C, Lozic M, Ludwig M & Leng G. (2019). Coding of odors in the anterior olfactory nucleus. Physiol Rep , 7, e14284. PMID: 31782263 DOI.
- ↑ Miller AM, Maurer LR, Zou DJ, Firestein S & Greer CA. (2010). Axon fasciculation in the developing olfactory nerve. Neural Dev , 5, 20. PMID: 20723208 DOI.
- ↑ Karn RC, Young JM & Laukaitis CM. (2010). A candidate subspecies discrimination system involving a vomeronasal receptor gene with different alleles fixed in M. m. domesticus and M. m. musculus. PLoS ONE , 5, . PMID: 20844586 DOI.
- ↑ Ogura T, Krosnowski K, Zhang L, Bekkerman M & Lin W. (2010). Chemoreception regulates chemical access to mouse vomeronasal organ: role of solitary chemosensory cells. PLoS ONE , 5, e11924. PMID: 20689832 DOI.
- ↑ Witt M & Hummel T. (2006). Vomeronasal versus olfactory epithelium: is there a cellular basis for human vomeronasal perception?. Int. Rev. Cytol. , 248, 209-59. PMID: 16487792 DOI.
- ↑ Dénes L, Pap Z, Szántó A, Gergely I & Pop TS. (2015). Human vomeronasal epithelium development: An immunohistochemical overview. Acta Microbiol Immunol Hung , 62, 167-81. PMID: 26132837 DOI.
- ↑ Mahdy EAA, El Behery EI & Mohamed SKA. (2019). Comparative morpho-histological analysis on the vomeronasal organ and the accessory olfactory bulb in Balady dogs (Canis familiaris) and New Zealand rabbits (Oryctolagus cuniculus). J Adv Vet Anim Res , 6, 506-515. PMID: 31819879 DOI.
- ↑ Villamayor PR, Cifuentes JM, Quintela L, Barcia R & Sanchez-Quinteiro P. (2019). Structural, morphometric and immunohistochemical study of the rabbit accessory olfactory bulb. Brain Struct Funct , , . PMID: 31802255 DOI.
- ↑ Fleischer J & Breer H. (2010). The Grueneberg ganglion: a novel sensory system in the nose. Histol. Histopathol. , 25, 909-15. PMID: 20503179 DOI.
- ↑ Paschaki M, Cammas L, Muta Y, Matsuoka Y, Mak SS, Rataj-Baniowska M, Fraulob V, Dollé P & Ladher RK. (2013). Retinoic acid regulates olfactory progenitor cell fate and differentiation. Neural Dev , 8, 13. PMID: 23829703 DOI.
- ↑ 24.0 24.1 Al-Noury K & Lotfy A. (2011). Role of multislice computed tomography and local contrast in the diagnosis and characterization of choanal atresia. Int J Pediatr , 2011, 280763. PMID: 21772853 DOI.
Reviews
Sarnat HB & Flores-Sarnat L. (2019). Development of the human olfactory system. Handb Clin Neurol , 164, 29-45. PMID: 31604554 DOI.
Sarnat HB & Flores-Sarnat L. (2017). Olfactory Development, Part 2: Neuroanatomic Maturation and Dysgeneses. J. Child Neurol. , 32, 579-593. PMID: 28424008 DOI.
Sarnat HB, Flores-Sarnat L & Wei XC. (2017). Olfactory Development, Part 1: Function, From Fetal Perception to Adult Wine-Tasting. J. Child Neurol. , 32, 566-578. PMID: 28424010 DOI.
Sarnat HB & Yu W. (2016). Maturation and Dysgenesis of the Human Olfactory Bulb. Brain Pathol. , 26, 301-18. PMID: 26096058 DOI.
DeMaria S & Ngai J. (2010). The cell biology of smell. J. Cell Biol. , 191, 443-52. PMID: 21041441 DOI.
Gane S. (2010). What we do not know about olfaction. Part 1: from nostril to receptor. Rhinology , 48, 131-8. PMID: 20502748 DOI.
Yuan TF. (2010). Smell with new neurons. Cell Tissue Res. , 340, 211-4. PMID: 20387075 DOI.
Schoppa NE. (2009). Making scents out of how olfactory neurons are ordered in space. Nat. Neurosci. , 12, 103-4. PMID: 19172161 DOI.
Munger SD, Leinders-Zufall T & Zufall F. (2009). Subsystem organization of the mammalian sense of smell. Annu. Rev. Physiol. , 71, 115-40. PMID: 18808328 DOI.
Wilson RI. (2008). Neural and behavioral mechanisms of olfactory perception. Curr. Opin. Neurobiol. , 18, 408-12. PMID: 18809492 DOI.
Articles
Nakano H, Iida Y, Murase T, Oyama N, Umemura M, Takahashi S & Takahashi Y. (2019). Co-expression of C/EBPγ and ATF5 in mouse vomeronasal sensory neurons during early postnatal development. Cell Tissue Res. , 378, 427-440. PMID: 31309319 DOI.
Taroc EZM, Naik A, Lin JM, Peterson NB, Keefe DL, Genis E, Fuchs G, Balasubramanian R & Forni PE. (2019). Gli3 regulates vomeronasal neurogenesis, olfactory ensheathing cell formation and GnRH-1 neuronal migration. J. Neurosci. , , . PMID: 31767679 DOI.
Jin ZW, Cho KH, Shibata S, Yamamoto M, Murakami G & Rodríguez-Vázquez JF. (2019). Nervus terminalis and nerves to the vomeronasal organ: a study using human fetal specimens. Anat Cell Biol , 52, 278-285. PMID: 31598357 DOI.
Sarnat HB, Flores-Sarnat L & Wei XC. (2017). Olfactory Development, Part 1: Function, From Fetal Perception to Adult Wine-Tasting. J. Child Neurol. , 32, 566-578. PMID: 28424010 DOI.
Sarnat HB & Flores-Sarnat L. (2017). Olfactory Development, Part 2: Neuroanatomic Maturation and Dysgeneses. J. Child Neurol. , 32, 579-593. PMID: 28424008 DOI.
Dénes L, Pap Z, Szántó A, Gergely I & Pop TS. (2015). Human vomeronasal epithelium development: An immunohistochemical overview. Acta Microbiol Immunol Hung , 62, 167-81. PMID: 26132837 DOI.
Bossy J. (1980). Development of olfactory and related structures in staged human embryos. Anat. Embryol. , 161, 225-36. PMID: 7469043
Search PubMed
Search Pubmed: Smell Development | olfactory receptors | Olfactory bulb development | anosmia
Historic
Humphrey T. The development of the olfactory and the accessory olfactory formations in human embryos and fetuses. (1940) J. Comp. Neurol. 431-468.
External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
Terms
- glomerulus - the functional unit of the olfactory bulb.
- odorant - a compound that elicits the perception of smell.
- odorant receptor - (OR) a receptor expressed by an olfactory sensory neuron. Receptor belongs to the G protein–coupled receptors (GPCR) superfamily. There are multiple families of odorant receptors, which include the OR (the largest family), TAAR, V1R, V2R, and formyl peptide-like receptors.
- olfactory cortex - refers to the brain regions that receive direct input from the olfactory bulb are responsible for the perception of smell and for generating odor-evoked behaviours. The five brain regions are the: piriform cortex, anterior olfactory nucleus, olfactory tubercle, entorhinal cortex, and amygdala.
- olfactory glomeruli - spherically shaped regions of neuropil where information is passed from sensory neurons to postsynaptic neurons.
- vomeronasal organ (VNO, Jacobson's organ) A neural structure forming part of olfactory system that functions in the detection of pheromones. In humans, the vomeronasal nerve, accessory olfactory bulb and chemoreceptor cells within the organ are lost. Named after Ludwig Lewin Jacobson (1783 – 1843) a Danish surgeon who identified it in 1813.
Additional Images
Historic Images
| Historic Disclaimer - information about historic embryology pages |
|---|
|
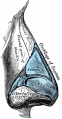
852 Cartilages of the nose Side view
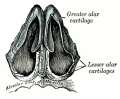
853 Cartilages of the nose, seen from below
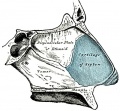
854 Bones and cartilages of septum of nose Right side
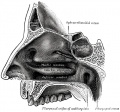
855 Lateral wall of nasal cavity
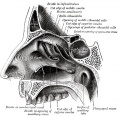
856

857
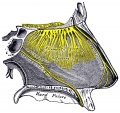
858
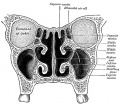
859 Coronal section of nasal cavities
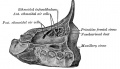
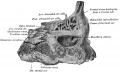
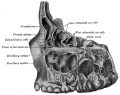











Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2026, July 12) Embryology Sensory - Smell Development. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Sensory_-_Smell_Development
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G