Cardiovascular System - Blood Vessel Development: Difference between revisions
mNo edit summary |
mNo edit summary |
||
| Line 32: | Line 32: | ||
|-bgcolor="F5FAFF" | |-bgcolor="F5FAFF" | ||
| | | | ||
* '''The relationship between human placental morphometry and ultrasonic measurements of utero-placental blood flow and fetal growth'''<ref name=PMID26907381><pubmed>26907381</pubmed></ref> "Placental area and weight are associated with uterine and umbilical blood flow, respectively, and both are associated with fetal growth rate." | |||
* '''Cell-matrix signals specify bone endothelial cells during developmental osteogenesis'''<ref name=PMID28218908><pubmed>28218908</pubmed></ref> “Blood vessels in the mammalian skeletal system control bone formation and support haematopoiesis by generating local niche environments. Here, we report that embryonic and early postnatal long bone contains a specialized endothelial cell subtype, termed type E, which strongly supports osteoblast lineage cells and later gives rise to other endothelial cell subpopulations. The differentiation and functional properties of bone endothelial cells require cell-matrix signalling interactions." [[Musculoskeletal_System_-_Bone_Development|Bone Development]] | [https://omim.org/entry/135630 INTEGRIN BETA-1] | * '''Cell-matrix signals specify bone endothelial cells during developmental osteogenesis'''<ref name=PMID28218908><pubmed>28218908</pubmed></ref> “Blood vessels in the mammalian skeletal system control bone formation and support haematopoiesis by generating local niche environments. Here, we report that embryonic and early postnatal long bone contains a specialized endothelial cell subtype, termed type E, which strongly supports osteoblast lineage cells and later gives rise to other endothelial cell subpopulations. The differentiation and functional properties of bone endothelial cells require cell-matrix signalling interactions." [[Musculoskeletal_System_-_Bone_Development|Bone Development]] | [https://omim.org/entry/135630 INTEGRIN BETA-1] | ||
* '''Endothelium in the pharyngeal arches 3, 4 and 6 is derived from the second heart field'''<ref name="PMID27955943"><pubmed>27955943</pubmed></ref> "Oxygenated blood from the heart is directed into the systemic circulation through the aortic arch arteries (AAAs). The AAAs arise by remodeling of three symmetrical pairs of pharyngeal arch arteries (PAAs), which connect the heart with the paired dorsal aortae at mid-gestation. Aberrant PAA formation results in defects frequently observed in patients with lethal congenital heart disease. How the PAAs form in mammals is not understood. The work presented in this manuscript shows that the second heart field (SHF) is the major source of progenitors giving rise to the endothelium of the pharyngeal arches 3 - 6, while the endothelium in the pharyngeal arches 1 and 2 is derived from a different source. During the formation of the PAAs 3 - 6, endothelial progenitors in the SHF extend cellular processes toward the pharyngeal endoderm, migrate from the SHF and assemble into a uniform vascular plexus. This plexus then undergoes remodeling, whereby plexus endothelial cells coalesce into a large PAA in each pharyngeal arch." | * '''Endothelium in the pharyngeal arches 3, 4 and 6 is derived from the second heart field'''<ref name="PMID27955943"><pubmed>27955943</pubmed></ref> "Oxygenated blood from the heart is directed into the systemic circulation through the aortic arch arteries (AAAs). The AAAs arise by remodeling of three symmetrical pairs of pharyngeal arch arteries (PAAs), which connect the heart with the paired dorsal aortae at mid-gestation. Aberrant PAA formation results in defects frequently observed in patients with lethal congenital heart disease. How the PAAs form in mammals is not understood. The work presented in this manuscript shows that the second heart field (SHF) is the major source of progenitors giving rise to the endothelium of the pharyngeal arches 3 - 6, while the endothelium in the pharyngeal arches 1 and 2 is derived from a different source. During the formation of the PAAs 3 - 6, endothelial progenitors in the SHF extend cellular processes toward the pharyngeal endoderm, migrate from the SHF and assemble into a uniform vascular plexus. This plexus then undergoes remodeling, whereby plexus endothelial cells coalesce into a large PAA in each pharyngeal arch." | ||
* '''Review - The Molecular Regulation of Arteriovenous Specification and Maintenance'''<ref name="PMID25641373"><pubmed>25641373</pubmed></ref> "The formation of a hierarchical vascular network, composed of arteries, veins and capillaries, is essential for embryogenesis and is required for the production of new functional vasculature in the adult. Elucidating the molecular mechanisms that orchestrate the differentiation of vascular endothelial cells into arterial and venous cell fates is requisite for regenerative medicine, as the directed formation of perfused vessels is desirable in a myriad of pathological settings, such as in diabetes and following myocardial infarction. Additionally, this knowledge will enhance our understanding and treatment of vascular anomalies, such as arteriovenous malformations (AVMs). From studies in vertebrate model organisms, such as mouse, zebrafish and chick, a number of key signaling pathways have been elucidated that are required for the establishment and maintenance of arterial and venous fates. These include the Hedgehog, Vascular Endothelial Growth Factor (VEGF), Transforming Growth Factor-β (TGF-β), Wnt and Notch signaling pathways. In addition, a variety of transcription factor families acting downstream of-or in concert with-these signaling networks play vital roles in arteriovenous (AV) specification. These include Notch and Notch-regulated transcription factors (e.g. HEY and HES), SOX factors, Forkhead factors, β-Catenin, ETS factors and COUP-TFII. It is becoming apparent that AV specification is a highly coordinated process that involves the intersection and carefully orchestrated activity of multiple signaling cascades and transcriptional networks." | * '''Review - The Molecular Regulation of Arteriovenous Specification and Maintenance'''<ref name="PMID25641373"><pubmed>25641373</pubmed></ref> "The formation of a hierarchical vascular network, composed of arteries, veins and capillaries, is essential for embryogenesis and is required for the production of new functional vasculature in the adult. Elucidating the molecular mechanisms that orchestrate the differentiation of vascular endothelial cells into arterial and venous cell fates is requisite for regenerative medicine, as the directed formation of perfused vessels is desirable in a myriad of pathological settings, such as in diabetes and following myocardial infarction. Additionally, this knowledge will enhance our understanding and treatment of vascular anomalies, such as arteriovenous malformations (AVMs). From studies in vertebrate model organisms, such as mouse, zebrafish and chick, a number of key signaling pathways have been elucidated that are required for the establishment and maintenance of arterial and venous fates. These include the Hedgehog, Vascular Endothelial Growth Factor (VEGF), Transforming Growth Factor-β (TGF-β), Wnt and Notch signaling pathways. In addition, a variety of transcription factor families acting downstream of-or in concert with-these signaling networks play vital roles in arteriovenous (AV) specification. These include Notch and Notch-regulated transcription factors (e.g. HEY and HES), SOX factors, Forkhead factors, β-Catenin, ETS factors and COUP-TFII. It is becoming apparent that AV specification is a highly coordinated process that involves the intersection and carefully orchestrated activity of multiple signaling cascades and transcriptional networks." | ||
Revision as of 12:39, 6 October 2017
| Embryology - 26 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Introduction

Blood develops initially within the core of "blood islands" in the mesoderm. During development, there follows a series of "relocations" of the stem cells to different organs within the embryo. In the adult, these stem cells are located in the bone marrow. At the time when blood first forms, there are no bones!
Note that blood vessel development is tightly coupled to development of other systems for example: osteogenesis (bone formation) that is dependent upon early capillary formation; endocrine development that requires blood vessels for hormone distribution.
| Vasculogenesis | Angiogenesis |
|---|---|
| formation of new blood vessels (endothelium from mesoderm) |
formation of blood vessels from pre-existing vessels (occurs in development and adult) |
Angioblasts initially form small cell clusters (blood islands) within the embryonic and extraembryonic mesoderm. These blood islands extend and fuse together making a primordial vascular network. Within these islands the peripheral cells form endothelial cells while the core cells form blood cells (haemocytoblasts).
Recent work has shown that the formation of the initial endothelial tube is by a process of coalescence of cellular vacuoles within the developing endothelial cells, which fuse together without cytoplasmic mixing to form the blood vessel lumen.
See also the related pages Arterial Development, Venous Development, Placental Villi Blood Vessels and Coronary Circulation Development.
Some Recent Findings

|
| More recent papers |
|---|
 This table allows an automated computer search of the external PubMed database using the listed "Search term" text link.
More? References | Discussion Page | Journal Searches | 2019 References | 2020 References Search term: Blood Vessel Embryology <pubmed limit=5>Blood Vessel Embryology</pubmed> |
| Older papers |
|---|
|
Endothelial Progenitors
Recent work has shown that the formation of the initial endothelial tube is by a process of coalescence of cellular vacuoles within the developing endothelial cells, which fuse together without cytoplasmic mixing to form the blood vessel lumen. [8]
Endothelial Tube Formation

Vessel Specification

The following data is from a recent review.[5]
Arterial Specification
| Factor | Function |
| Shh | Loss of Shh results in lack of arterial identity in zebrafish. Shh acts upstream of VEGF. |
| VEGF | VEGF acts downstream of Shh signaling to activate Notch via the PLCγ/ERK pathway in zebrafish. Mutant mice expressing only VEGF188 lack arterial differentiation. |
| Nrp1 | Null mice display impaired arterial differentiation. Nrp1 is involved in a positive feedback loop of VEGF signaling. |
| Notch | Notch acts downstream of Shh and VEGF signaling in zebrafish. Notch1; Notch4 mutant mice have abnormal vascular development. |
| Dll4 | Null mice lack arterial specification. |
| Dll1 | Null mice fail to maintain arterial identity. |
| Hey1/2 (Grl) | Null mice lack arterial specification. Lack of grl in zebrafish results in loss of arterial specification. |
| Foxc1/c2 | Foxc1; Foxc2 mutant mice lack arterial specification. Foxc1 and Foxc2 directly regulate Dll4 and Hey2 expression. Foxc1 and Foxc2 are also involved in lymphatic vessel development. |
| Sox7/18 | Lack of Sox7/18 results in loss of arterial identity in zebrafish. |
| Snrk-1 | Snrk-1 acts downstream or parallel to Notch signaling in zebrafish. |
| Dep1 | Dep1 acts upstream of PI3K in arterial specification in zebrafish. |
| Crlr | Shh regulates VEGF activity by controlling crlr expression in zebrafish. |
| EphrinB2 | Null mice lack boundaries between arteries and veins. EphrinB2 is involved in lymphatic vascular remodeling and maturation. |
Venous Specification
| Factor | Function |
| COUP-TFII | COUP-TFII suppresses arterial cell fate by inhibiting Nrp1 and Notch. COUP-TFII also interacts with Prox1 to regulate lymphatic gene expression. |
| EphB4 | Null mice lack boundaries between arteries and veins. |
Lymphatic Specification
| Factor | Function |
| Sox18 | Null mice fail to specify lymphatic endothelial cells. Sox18 induces Prox1 expression. |
| Prox1 | Prox1 induces lymphatic markers and maintains lymphatic cell identity. |
Vascular Endothelial Growth Factor
Growing blood vessels follow a gradient generated by tagret tissues/regions of Vascular Endothelial Growth Factor (VEGF) to establish a vascular bed. Recent findings suggest that Notch signaling acts as an inhibitor for this system, preventing sprouting of blood vessels.
Notch is a transmembrane receptor protein involved in regulating cell differentiation in many developing systems.

|

|
| Notch and yolk sac blood vessels model[9] | Vasculogenesis and angiogenesis[10] |
Links: OMIM - VEGFA | OMIM - Notch
Regulators of Growth
The following data is from a review article on ovary vascular development.[11]
Stimulators of Angiogenisis
|
Inhibitors of Angiogenisis
|
Histology
Vein Light Microscopy

The entire developing and adult cardiovascular system (blood vessels and heart) is lined by a simple squamous epithelium. (Stain - Haematoxylin Eosin)
Capillaries
Type H
A developmental capillary endothelial cell subtype associated with osteogenesis, located at the metaphysis and endosteum of postnatal long bone, that couples angiogenesis with osteogenesis. This endothelial cell subtype expresses the markers CD31/PECAM1 and endomucin (CD31hi Emcnhi).
Type E
A newly identified endothelial cell subtype similar to type H in function, supporting osteoblast lineage cells and then gives rise to other endothelial cell subpopulations, but this subtype is found in embryonic and early postnatal long bone.[2]
Electron Micrographs


Capillary 1 large labeled
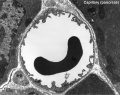
Capillary 1 large unlabelled

Capillary 1 small labeled
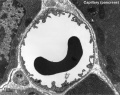
Capillary 1 small unlabelled
![endothelium detail[12]](/embryology/images/thumb/6/6a/Blood_capillary_EM_01.jpg/120px-Blood_capillary_EM_01.jpg)
endothelium detail[12]
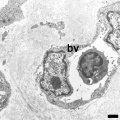
Containing white blood cell



![endothelium detail[12]](/embryology/index.php?title=File:Blood_capillary_EM_01.jpg)

Arteries
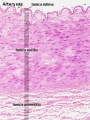
Artery overview
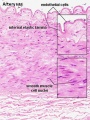
Artery detail

Artery elastin
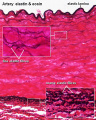
Artery elastin detail
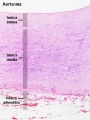
Aorta overview
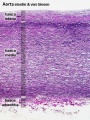
Aorta elastin
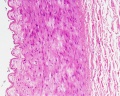
Artery overview

Artery elastin

Artery tunica media elastin

Artery elastin detail
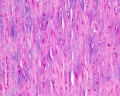
Aorta overview
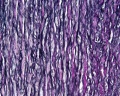
Aorta elastin












Cardiac Blood Vessels
Earliest vessels in the heart wall develop subepicardially (beneath the outside surface of the heart) near the apex at Carnegie stage 15, which then extends centripetally and at stage 17 coronary arterial stems communicate with the aortic lumen.[13]
Abnormalities
Due to the extensive embryonic, and ongoing, remodelling of the vascular system, there are many different vascular variations and anomalies.
Neural

Persistent trigeminal and hypoglossal arteries[14]
- Links: Cerebrum Development | Head Development
References
- ↑ <pubmed>26907381</pubmed>
- ↑ 2.0 2.1 <pubmed>28218908</pubmed>
- ↑ <pubmed>27955943</pubmed>
- ↑ <pubmed>25641373</pubmed>
- ↑ 5.0 5.1 <pubmed>20238301</pubmed>| PMC2899674
- ↑ <pubmed>18417617</pubmed>
- ↑ <pubmed>18245384</pubmed>
- ↑ <pubmed>11827993</pubmed>
- ↑ <pubmed>21352545</pubmed>| BMC Dev Biol.
- ↑ <pubmed>21537463</pubmed>
- ↑ <pubmed>11141338</pubmed>
- ↑ <pubmed>21702933</pubmed>| PMC3141733 | BMC Cell Biol.
- ↑ <pubmed>8915616</pubmed>
- ↑ <pubmed>26060802</pubmed>| J Stroke.
Reviews
<pubmed></pubmed> <pubmed></pubmed> <pubmed>25641373</pubmed>
Articles
<pubmed></pubmed> <pubmed></pubmed> <pubmed>10948449</pubmed> <pubmed>12406884</pubmed>
Search Pubmed
Click on the listed keywords below (used to search the external database) the most current references on Medline will be displayed.
Search Pubmed: Blood Vessel Development | Blood Vessel embryology | Blood Vessel smooth muscle Development | Blood Vessel smooth muscle Development
Terms
- cerebroplacental ratio - (CPR) measured by doppler ultrasound, the ratio between the middle cerebral artery pulsatility index (PI) MoM and the umbilical artery PI. A potential predictor of adverse pregnancy outcome.
- pulsatility index - (PI) systolic peak velocity/diastolic peak velocity)/velocity time integral
- resistance index - (RI) systolic peak velocity/diastolic peak velocity)/systolic peak velocity
- S/D - systolic/diastolic ratio
| Placenta Terms (expand to view) |
|---|
with an incidence of about 2.8 per 1,000 pregnancies, there is also a rarer form of extra-abdominal varices.PMID 24883288
with an incidence of about 2.8 per 1,000 pregnancies, there is also a rarer form of extra-abdominal varices. PMID 24883288
|
| Other Terms Lists |
|---|
| Terms Lists: ART | Birth | Bone | Cardiovascular | Cell Division | Endocrine | Gastrointestinal | Genital | Genetic | Head | Hearing | Heart | Immune | Integumentary | Neonatal | Neural | Oocyte | Palate | Placenta | Radiation | Renal | Respiratory | Spermatozoa | Statistics | Tooth | Ultrasound | Vision | Historic | Drugs | Glossary |
| Cardiovascular Terms |
|---|
Cardiovascular System Development See also Heart terms, Immune terms and Blood terms.
|
| Other Terms Lists |
|---|
| Terms Lists: ART | Birth | Bone | Cardiovascular | Cell Division | Endocrine | Gastrointestinal | Genital | Genetic | Head | Hearing | Heart | Immune | Integumentary | Neonatal | Neural | Oocyte | Palate | Placenta | Radiation | Renal | Respiratory | Spermatozoa | Statistics | Tooth | Ultrasound | Vision | Historic | Drugs | Glossary |
External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2024, April 26) Embryology Cardiovascular System - Blood Vessel Development. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Cardiovascular_System_-_Blood_Vessel_Development
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G