Paper - The development of the adrenal gland in man (1957)
| Embryology - 13 May 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Crowder RE. The development of the adrenal gland in man, with special reference to origin and ultimate location of cell types and evidence in favor of the "cell migration" theory. (1957) Contrib. Embryol., Carnegie Inst. Wash. 36, 193-210.
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
The Development of the Adrenal Gland in Man, with Special Reference to Origin and Ultimate Location of Cell Types and Evidence in Favor of the “Cell Migration” Theory
by Roy E. Crowder
With 8 plates and 7 text figures.
- The ideas herein expressed are those of the author and do not reflect those of the United States Navy.
Introduction
A voluminous literature has accumulated on the development of the adrenal gland. In 1902, Iosef \Viesel apologized for adding one more article. Others, including the present author, have not been so restrained.
In a study of the sources of certain of the adrenal hormones, a review of the literature on the embryology and histology of the gland was undertaken. In spite of the number of articles published on the subject, including extensive reviews (Parkes, I945; Lanman, 1953), there was no consecutive life history of the source and ultimate distribution of the various cell types found in the gland. and, therefore, it was not possible to associate the synthesis of the adrenal hormones with specific cell types.
The wealth of closely seriated, well preserved, and adequately stained material available in the Carnegie Embryological Collection, in Baltimore, Maryland, made a complete histologic and embryologic study possible.
The study was begun with the idea of determining the types of cells in the adrenal cortex. It was soon found that, to do this adequately, the whole embryologic history must be reviewed. The results of this undertaking are set forth herein. It is hoped not only that they will assist in an understanding of the sequence of events in the histogenesis of the adrenal cortex and clarify a few of the points of controversy, but also that the descriptions of cell types in the cortex may be of value in determining the oligoplasia or hypcrplasia of certain of these cell types in relation to many clinical entities.
Definitions
The following terms are defined to clarify the presentation:
Fetal cortex: The fetal adrenal cortex is composed of the same cell types as the adrenal cortex of the adult human, but the proportions and arrangements differ.
Medullary cells: The cells of neural origin which take up a location in the central portion of the gland during development. The process of localization begins in the embryonic period and continues until childhood.
Cell types: Cell types in the cortex are differentiated by the structure of their nuclei; they are numbered according to the order of their appearance in the primordium, as C-I (fig. 38, pl. 8), C-II (fig. 39, pl. 8), and C-III (fig. 40, pl. 8). As will be made clear, three distinct cell types may be recognized in the development of the medulla; these cells, designated M-I, M-II, and M-III, are described beginning on page 202.
Coelomic epithelial cells: The cells of the adrenal cortex which have their origin in the celomic epithelium. They are of two types: Type C-I (fig. 38, pl. 8), the first cells to enter the primordium, are large polyhedral cells, each of which contains a large round nucleus with little chromatin, a large round nucleolus, and a spherical, refractile inclusion. In a later stage, type C-III cells (Hg. 40, pl. 8) begin to emerge from the celomic epithelium, together with type C-I. Type C-III cells are smaller than type C-1, and each contains a smaller round nucleus which stains more deeply with hematoxylin. In the nucleus. there is a large amount of chromatin, which tends to aggregate into four to five major clumps and one or two globules or vacuoles.
Cells of mesonephric origin: These cells arise from Bowman's capsule of the mesoncphros. They are designated as type C-II cells (fig. 39, pl. 8), and are first seen streaming from the glomerular capsule across the ventral portion of the early primordium (figs. 14, 15, pl. 2). The type C-II cells and their successors form most of the adrenal capsule, and, during horizon xxiii, are diverted inward by the arteriolar capillaries. Thereafter, they are found in the cords of cells within the cortex. Their nucleus is round, and stains a little more deeply than does that of type C-I. The chromatin is distributed in fine strands in a spider-like manner just beneath the nuclear membranes, the nucleolus corresponding to the body of the spider. The center of the nucleus is occupied by one or more large globules or vacuoles.
Immigration and migration: The passage of cells from one region to another; the addition of cells by movement from an adjacent region. Ameboid movement is not implied.
Invasion: The entrance of cells after destruction or disruption of pre-existing tissue, and the assumption of position without rcference to former relations.
Staging of embryos: The dating of the embryos follows the horizons described and graphed by Streeter (1951). This method of seriation affords a much better concept of the progress of development than does measurement of the length. Frequently, the organogenesis of the adrenal gland was found to be in a much further advanced stage in an embryo with a shorter crown-rump length than in a longer one, particularly in the earlier stages when the dorsal curvature varies so remarkably among embryos in the same horizon. This finding speaks well for the method, since Streeter did not use the adrenal among his criteria for staging. The age in weeks is the estimated conception age. To obtain menstrual age, two weeks must be added. There are enough stages to provide a consecutive account of the organogenesis of the adrenal.
Materials and Methods
The following embryos of the Carnegie Collection were selected for the study of the embryonic and early fetal development of the adrenal gland.
| Serial no. | Crown-Rump Length (mm) | Plane of section | Histological condition | Stain | |
|---|---|---|---|---|---|
| 836 | 4.0 | Trans. | Exc. A1. coch. | ||
| 7433 | 5.2 | Frontal | Exc. | (Stain - Haematoxylin Eosin) | |
| 8066 | 5.3 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 8119 | 5.3 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 3805 | 5.9 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 6502 | 6.7 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 7324 | 6.6 | Trans. | Good | (Stain - Haematoxylin Eosin) | |
| 7829 | 7.0 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 7370 | 7.2 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 8314 | 8.2 | Trans. | Exc. | Azan | |
| 2 | 7.0 | Trans. | Good | Al. carm. | |
| 721 | 9.0 | Trans. | lixc. | ||
| 3385 | 8.3 | Trans. | Exc. | (Stain - Haematoxylin Eosin) or G. | |
| 3441 | 8.0 | Sag. | Exc. | Al. coch. | |
| 3512 | S 5 | Trans. | Exc. | Al. coch. | |
| 6504 | 7 5 | Sag. | Exc. | Al. coch. | |
| 6508 | 7.3 | Trans. | Good | Al. coch. | |
| 6595. | 3.0 | Sag. | Good | I-1.-17.. | |
| 7199 | 8.-'1 | Frontal | Good | Al. coch. phloxine | |
| 7364 | 9 5 | Frontal | Poor | 1-1.-17.. | |
| 8929 | 6 3 | Frontal | Exc. | Azan | |
| 8997 | 9 0 | Trans. | Exc. | Azan | |
| 9140 | 7 0 | Trans. | Exc. | Azan | |
| 792 | 8 0 | Trans. | Good | Al. coch. | |
| 6511 | 8.1 | Sag. | Good | Al. coch. iron H. | |
| 6512 | 70 | Trans. | Exc. | Al. coch. | |
| 6517 | 10.5 | Trans. | Exc. | Al. coch. | |
| 7115 | 9.7 | Frontal | Exc. | H. and pltloxinc | |
| 7804 | 9.5 | Trans. | Good | (Stain - Haematoxylin Eosin) | |
| 7897 | 12 2 | Trans. | Good | (Stain - Haematoxylin Eosin) | |
| 8773 | ll 0 | Frontal | Exc. | Azan | |
| 940 | 14.0 | Trans. | Good | (Stain - Haematoxylin Eosin) or G. | |
| 5893 | 13 2 | Trans. | Good | Al. coch. | |
| 6253 | 1-10 | Trans. | Good | (Stain - Haematoxylin Eosin) | |
| 6519 | 0 3 | Sag. | Exc. | Al. coch. | |
| 6520 | 14 2 | Trans. | Exc. | Al. coch. | |
| 6742 | 11 0 | Trans. | Good | H.-phloxine | |
| 6758 | 12 8 | Trans. | Good | H.-phloxine | |
| 8101 | 13.0 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 811$ | 12.6 | Frontal | Exc. | (Stain - Haematoxylin Eosin) | |
| 8789 | ll 7 | Sag. | Exc. | Azan | |
| 3998 | 11 0 | Frontal | Exc. | Azan | |
| -192 | 16.3 | Frontal | Exc. | Al. coch. | |
| 1909 | 14.6 | Frontal | Good | Al. coch. or G. | |
| 4430 | 4.0 | Trans. | Exc. | Al. coch. or G. | |
| 6524 | 11.7 | Trans. | Exc. | Al. coch. | |
| 6525 | 3.8 | Sag. | Exc. | Al. coch. | |
| 6527 | 4.4 | Trans. | Exc. | Al. coch. | |
| 6528 | 3.4 | Frontal | Exc. | Al. coch. | |
| 7707 | 4 5 | Trans. | Exc. | 1-1.—F.. phloxine | |
| 8172 | 16 5 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 8312 | 2.2 | Trans. | Exc. | I-1.717.. | |
| 9247 | 5.0 | Sag. | Exc. | Azan | |
| 1909 | 15 5 | Trans. | Good | Coch. l\1allor_\-' | |
| 4501 | 80 | Trans. | Exc. | Coclt. or G. | |
| 5609. . 18 0 | Frontal | Exc. | Al. coch. | ||
| 6150 | 17.0 | Trans. | Good | ||
| 632-l | 8.5 | Sag. | Good | (Stain - Haematoxylin Eosin) | |
| 8092 | 16 3 | Trans. | Exc. | I-1.-E. phlo.\;ine | |
| 9113 | 18 5 | Trans. | Exc. | Azan | |
| 966 | 23.0 | Frontal | Exc. | Al. coch. | |
| 5537 | 22.0 | Trans. | Fxc. | Al. coch. | |
| 6202. | 21.0 | Sag. | Exc. | (Stain - Haematoxylin Eosin) | |
| 7274 | 8 3 | Trans. | Exc. | (Stain - Haematoxylin Eosin) phloxinc | |
| 7906 | 19.5 | Frontal | Exc. | (Stain - Haematoxylin Eosin) | |
| 8226 | 8.0 | Sag. | Exc. | Azan | |
| 5596 | 21.5 | Sag. | Good | I-1.-Ii. | |
| 7251 | 22.5 | Trans. | Exc. | ||
| 7392 | 22.7 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 7864 | 24 0 | Frontal | Exc. | (Stain - Haematoxylin Eosin) | |
| 8553 | 22.0 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 1458 | 27.5 | Sag. | Exc. | or G | |
| 430-1 | 25 0 | Trans. | Good | (Stain - Haematoxylin Eosin) | |
| 4638 | 23.4 | Trans. | Exc. | Al. coch | |
| 6701 | 24.0 | Frontal | Poor | (Stain - Haematoxylin Eosin) | |
| 6832 | 25.8 | Frontal | Exc. | I-1..F_. | |
| 339-1 | 25 3 | Trans. | Exc. | I1.-I’. | |
| 4205 | 29.5 | Trans. | Good | Al coch | |
| 4289 | 32.2 | Trans. | Good | Al coch | |
| 4570 | 30.7 | Trans. | Exc. | (Stain - Haematoxylin Eosin) | |
| 5122 | 27.0 | Sag. | Good | (Stain - Haematoxylin Eosin) | |
| 5621 | 27.5 | Trans. | Good | ||
| 6573 | 31.5 | Trans. | Good | (Stain - Haematoxylin Eosin) | |
| 6719 | 30.1 | Trans. | Good | Chrom.alum hem. | |
| 7425 | 27.0 | Frontal | Exc. | Chrom.alum hem. | |
| 9226 | 30.1 | Trans. | Exc. | Azan | |
In addition to the materials listed, a number of glands from late embryos and fetuses (too-mm. crown-rump length upwards) and newborn children, through the first month of postnatal life, were collected by the author, fixed in Bouin’s fluid, and stained by various means, including hematoxylin and eosin, Mallory-azan, and
chrome alum hematoxylin counterstained with phloxine or Mallory trichrome in which the aniline blue was replaced with light green SF, C. I. No. 670. Portions of some of the glands were fixed in Bouin’s and then placed in a saturated solution of potassium dichromate at 37”’ C. for 72 hours, after fixation, to preserve the lipids, according to the method suggested by Thomas (1948) for spheroid bodies in nerve cells. Frozen sections were prepared and stained for lipids with Sudan III, as were some of the post-chromated sections.
Adrenals of children and adults were selected from the pathology collections in the United States Naval Hospitals in St. Albans, New York, and Camp Lejeune, North Carolina. All were fixed in neutral formalin and stained with hematoxylin and eosin or hematoxylin and phloxine.
Adrenal glands in the macaque embryos in the Carnegie Collection were examined, as were glands from adult cats, hamsters, and guinea pigs, to see whether cell types in the human were species specific.
Development of the Adrenal Cortex
The adrenal primordium is first recognizable in embryos early in horizon xv (fig. T; fig. 8, pl. 1). A change in the characteristics of the cells of the celomic epithelium over the area between the gastric mesentery and the lateral extremity of the mesonephros can be seen late in horizon xiv, but not until horizon xv is a new type of cell (type C-1) found in the mesenchyme dorsal to the epithelium in this region. It can be traced by continuity of cords from the celomic epithelium (fig. IO, pl. 1). The portion of the celomic epithelium medial to the mesonephros produces cells destined for the adrenal cortex, whereas the region ventral to the mesonephros produces cells destined for the gonad. In the earliest stages, cells from the two areas are morphologically similar.
Mitotic figures are seen in profusion in the celomic epithelium, and cells in various stages of differentiation are present immediately dorsal to it. The orientation of the mitoses is such that one daughter cell is seen in the developing primordium while the other remains in the epithelium. It is suggested that, in this manner, as cells are formed, the epithelium moves ventrally, leaving columns of cells in the primordium (figs. 9, 10, pl. 1).
In the epithelium, the cells, which are cuboidal to low columnar, are crowded together. The chromatin of the nuclei is fairly dense, and there is little cytoplasm. As the cells move into the adrenal (and the gonadal) area, they increase in size and assume the form of large polyhedral epithelioid elements with light-colored, eosinophilic cytoplasm. The nucleus also enlarges; it becomes spheroidal, with a relatively small amount of chromatin loosely arranged in fine strands and small knots. The nucleolus is somewhat eccentrically placed, and the center of the nucleus is occupied by a large spherical refractile body which displaces about one—tenth of the nuclear volume (fig. 10, pl. 1; fig. 38, pl. 8).
In horizon xv, a change appears in the characteristics of the cells in the medial wall of the capsule of the mesonephric glomerulus (fig. 2; fig. 11, pl. 1; fig. 12. pl. 2). Until this time, the cells of Bowman’s capsule are large and squamous, having biconvex nuclei. As the new cells (type C-II; fig. 23, pl. 4) are produced, they are columnar, with their long axes at right angles to the capsular surface. In figure 13 (pl. 2), some of these cells have already migrated into the adrenal primordium. During horizon xvi, there is an enormous immigration of cells originally derived from Bowman’s capsule. For the most part, they leave the medial surface of the glomerular capsule and pass medially just beneath the celomic epithelium as a layer four or five cells deep (fig. 3; figs. I4, 15, pl. 2). Toward the dorsal portion of the medial surface of the actively proliferating Bowman’s capsule, near the hilum of the glomerulus, are located cells which send their progeny into the dorsolateral part of the primordium. At first, these cells are comparatively few, but they form a nidus which sends forth a great shower of cells to the adrenal and the gonad in horizon xviii (figs. 22, 23, pl. 4).
The cytoplasm of these type C-11 cells is inconspicuous in horizon xvi. The nuclei, however, are distinctive, for they are long ellipsoids. The chromatin is scanty and is distributed in very fine strands that weave a coarse, netlike pattern beneath and close to the nuclear membrane. Vvhere the strands intersect, there are small knots. A few of the strands touch the nucleolus, which is an irregular, small, flat mass lying in contact with the nuclear membrane, usually near the middle of the long axis. The nucleus is translucent. Its center is occupied by one or two ellipsoid globules or vacuoles. Figure 39 (pl. 8) shows a type C-II cell in the cortical cords of an embryo in horizon xxiii.
In horizon xvi, also, the first evidence of a change in the cell types in the cclomic epithelium is seen. Cell type C-I is still present and continues to multiply in the celomic epithelium, together with another cell type having a smaller, darker nucleus, type C-III of the adrenal cortex (fig. 40, pl. 8). The cells emerge and enter the gland primordium in columns in juxtaposition to, but discrete from, the type C-I cell columns. The diFference between the two types can be seen clearly after they enter the primordium. The new cell is smaller; its nucleus is threefifths to three—fourths the size of the nucleus of type C-I. The chromatin appears clotted. There are four to five fairly large clumps of deeply staining basophilie material connected by ragged strands. The nucleolus is erratically placed and rarely central. As a rule, two or more small globules or vacuoles can be made out in the center of the nucleus. The cytoplasm is more basophilie than in types C-I or C-II.
From the first appearance of the primordium until well into horizon xviii, mitotic figures are seen in the central portion of the gland. Most of these mitoses are in blood cells, others in the capillary endothelium, but some are in cells of types C-I or C-III. As far as can be seen, type C-II cells do not enter the body of the gland until horizon xxiii.
In horizon xvi, nerve tissue in and around the sympathetic ganglion begins to grow and extend, passing ventrally between the gland and the aorta along the caudal third to half of the gland. Cells of neural origin have already begun to migrate into the gland, disrupting its more or less homogeneous pattern (fig. 3). The neural elements tend to pass between the cells of the primordium, separating them into islands rather than dcstroying them (fig. 4; fig. 15, pl. 2).
During horizon xvii, the first neural migration is at its height. This period is marked by the development of the great para-aortic complex of nervous tissue that eventually forms the abdominal plexuses. The growth is extensive; most of the tissues in the path of the invading nerves are penetrated. In the adrenal area, nerve fibers, accompanied by large “paraganglion” (type M-III, to be discussed later) cells, pass into the cortical primordium (fig. 15, pl. 2). As a result, by the end of the horizon, the cortical cells are divided into clusters or islands in the dorsal part of the gland. The structure of the ventrolateral portion of the gland, however, is not disturbed by the invading nerves. In this area, the cells streaming medially from Bowman’s capsule appear to act as a barrier to further progress of the nerves (fig. 18, pl. 3). The line of demarcation is striking. The celomic epithelium, which is still actively proliferating, and the layer of mesonephric cells are left undisturbed. This is the stage in which Keene and I-Iewer (1927) postulated a “permanent cortex” (figs. 17, 18, pl. 3). As will be seen in this horizon and in horizons xviii and xix, the pause is merely a temporary interruption in the migration of cells from the celomic epithelium directly into the primordium in certain areas.
The caudal end is the first portion of the cortex disturbed by the neural migration. All levels have been involved by the end of horizon xvii. The neural elements increase the volume of the primordium in the human embryo, so that a cross section late in horizon xvii is more than 2% times the area at a similar level at the end of the previous horizon. Throughout the remainder of the fetal period, the cortical cells increase in number at a steady and rapid rate, whereas the neural elements have periods of relative quiescence and periods of rapid growth. During the growth periods, the relation of the cortical cells to the nerve cells may be altered by the relative volume of the two elements (figs. 6, 7; figs. 16, 17, p .3; fig. 28, pl. 5). The nerve fibers and neuroblasts are not seen in the body of the gland until horizon xxiii, except in those areas where they are entering the gland or where they are destined to become definitive nerves. From evidence found in later stages, it appears that the distinctive cells (M-III) destined for the medulla are derived from the large cells that accompany the sympathetic nerves during the primary immigration in horizons
Figures 1 to 5, drawn to the same scale, X 100, illustrate the relative size of the adrenal in horizons xiv to xvii, compared with surrounding structures. Figures 6 and 7 represent the disruption of the cortex by the neural elements in horizon xxiii.

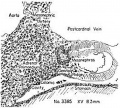
Fig. 1
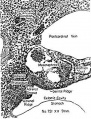
Fig. 2
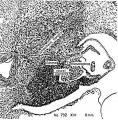
Fig. 3
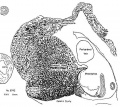
Fig. 4
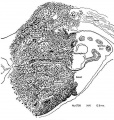
Fig. 5
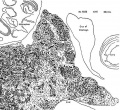
Fig. 6

Fig. 7







Fig. 1. Earliest evidence of adrenal primordium and change in character of cells in celomic epithelium over adrenal and g0n:I(_lal areas.
Fig. 2. Genital and adrenal ridges are formed, with adrenal groove separating them. The two groups of cells in the medial wall of Bowman's capsule have tlilierentiated preliminary to migration shown in figure 3. Active proliferation in the sympathetic ganglion is beginning.
Fig. 3. Active migration of cells from I:'»owman's capsule. Beginning invasion of primordium by sympathetic neural elements.
Fig. 4. Massive growth of sympathetic elements. So-called “permanent" cortex. Cell types C-I, C-ll, C-III are present in it.
Fig. 5. Height of sympathetic invasion. Cortex pattern is disrupted by sympathetic elements. Rim of cortex is still intact along the celomic border. Cells from Bowman's capsule and celomic epithelium are still being added to the primordium. Islands of cortical cells are scattered among the neural elements.
Fig. 6. Conditions near the caudal pole of the adrenal at the beginning of the second impetus to growth of sympathetic nerve elements. Much of the cortex is displaced. No capsule is yet formed in this area.
Fig. 7. Condition of the gland when proliferation of sympathetic neural elements is most active. The cortical cords are pushed aside to make room for the nerves and chromaflin elements. The retiform pattern of the sympathetic is well illus. trated. I’ o 1%} .2 '3;
xvi and xvii (see “Development of the Adrenal Medulla, below).
From horizon xix until horizon xxii, and particularly during xxiii, the character of the cytoplasm of the cortical cells differs only slightly in the peripheral and central zones of the gland. The cells are more densely packed in the outer zone of the gland, where there are fewer capillaries than in the center. Cells similar in all respects to types C-1 and C-11], formerly found in the celomic epithelium, are closely packed just central to the capsule, which is composed of type C-II cells. More centrally, they are expanded into the three cortical cell types and are grouped into cords of varying thickness by the sinusoids (fig. 25, pl. 5). The cord formation started in horizon xviii with the beginning growth of the sinusoids. At first, there are only a few thick cords near the center of the gland. As the sinusoids increase in number and ramify toward the periphery of the gland, the thickness of the cords is reduced; i.e., the number of cell columns per cord diminishes. The gland continues to expand peripherally, and the type C-I and type C-III cells are found in the cords, supported by the endothelium of the sinusoids.
The characteristics of the cells of the adrenal groove do not seem to differ at any time during its existence from the cells in other parts of the celomic epithelium which pass into the adrenals and gonads (fig. 22, pl. 4).
The type C-II cells from the nidus (figs. 22, 23, pl. 4) are identical with those that came from Bowman’s capsule during horizons xvi and xvii, described above. During horizon xix, these cells come to lie on the surface of the gland on all sides, forming the capsule of the gland. Where its surface is in contact with the celomic cavity, the capsule assumes a position in direct contact with the peritoneum, either by migrating there or as a result of migration of the epithelial cells inward, leaving the mesonephric cells to take their place. The former seems more likely, since many groups of cells of types C-1 and C-III are found outside the capsule as accessory adrenals. Also, in figure 23 (pl. 4), the cells from the nidus are seen to be migrating between the cells in the celomic epithelium.
By horizon xx, the layer of cells of type C-II is found around the periphery of the gland as a loosely organized capsule (Hg. 25, pl. 5). During horizons xxi and xxii (fig. 26, pl. 5), the capsule becomes better organized and is coveretl with a layer of fibrous tissue, and arteries appear over the surface. The celomic surface is now covered by peritoneum. VVith further development of the fetus and growth in the perirenal area, the gland soon becomes completely separated from the peritoneum.
During horizon xviii, the cortical elements are restored to order (fig. 24, pl. 5), and the succession of events proceeds. The reorganization of the gland is brought about early in the horizon by a rapid migration of cells from the celomic epithelium and multiplication of the cells in the islands and around the periphery of the gland. Since most mitoses are at or near the surface, the gland grows centrifugally; i.e., as cells form they remain in place, enlarging and differentiating, and new ones are added at the surface. The gland is not yet separated from the celomic epithelium except caudally. In the caudal region, it expands retroperitoneally on the left late in horizon xvii; on the right, it expands dorsal to the liver. Cells continue to proliferate from the celomic epithelium until, in horizon xix, the gland is separated from it by the spreading type C-II cells.
The separation begins in horizon xviii. ‘When the nidus formed by cells from the dorsal portion of the mesonephric glomerular capsule during horizon xvi and early xvii (fig. 22, pl. 4) springs into activity, the cells from this source spread around the periphery of the gland, some dorsally but most ventrally. Those passing ventrally are split by the prominent adrenal groove, part going to the adrenal and part to the gonad.
In horizon xxiii, the capsule, which is now completely formed, is penetrated in many areas by arterial capillaries which join the sinusoids. Where they enter, the capsule appears to be invaginated, so that the surface of an enlarged model of the gland would have a cobblestone appearance (Hg. 27, pl. 5).
The glomerular zone is now formed (Hg. 27, pl. 5). It is composed of nests of types C-1 and C-III cells, similar in all respects to their ancestors in the celomic epithelium during the embryonic period, held in place by the canopies of capsular (type C-II) cells between the penetrating arterial capillaries. The cells of types C-1 and C-III in the glomcruli are closely packed, with little cytoplasm and with compact nuclei. The number of each cell type varies widely from one glomcrulus to another, the distribution apparently depending on the random location of the two types in the periphery of the gland when the glomeruli were formed in horizon xxiii. The nuclei of the C-II type cells in the capsule, which changed from plump, translucent ellipsoids to compact spindle shape during horizon xxi, resemble nuclei of fibrous connective tissue. If embryos of the transitional period had not been available, the identity of these cells would have been lost.
As new cells of types C-1 and C-III are formed in the glomeruli, they migrate, or are forced by the vi: a tergo of multiplying cells, toward the center of the gland in cords between the capillary tributaries of the sinusoids. During the exit from the glomcrulus, they are joined by the type C-II cells from the capsule, which are maturing and moving from the surface toward the center of the gland on the surface of the cords and along the arterial capillaries and sinusoids (Hg. 27, pl. 5; fig. 36, pl. 8). All three types of cortical cells are now found in the cell cords in a more or less random pattern. W/here the cords are straight enough, however, it is possible to see each type of cell in a chainlike arrangement side by side with one or both other types of cells. By the time the fetus is 40 mm. in crown-rump length, the cells of type C-II are as numerous as those of the other types and are found in proportionate numbers in relation to them and well down the length of the cords.
At about 35 mm. crown-rump length, the cytoplasm of all cells in the central region of the gland becomes finely granular and markedly eosinophilic, and some of the cells near the large venous channels in the center seem to be vacuolated.
Between horizon xx and 30-mm. crown-rump length, the gland develops a pyramidal form with an overhanging lip on the medioanterior border because of the adjustment to the enlarging metanephros posterolaterally. During the next 10 to 15 mm. of growth, the adrenal vein and its three tributaries are formed (see “Development of Blood Supply of the Adrenal”). At the same time, the cords of cells from the periphery of the gland appear to disintegrate in the central portion (figs. 32-34, pl. 7). After this disintegration, the walls of the gland begin to collapse around the veins. The collapse is not complete, however, since the cells in the glomerular zone are still multiplying rapidly and going into the cords. The growth is about the same on all sides; therefore, the cords from the periphery to the three tributary veins are approximately equal in length. The gland then assumes a trilobate form, with a vein as the axis of each arm. Under normal conditions, there is little folding of the capsuleduring fetal life such as is found after the first postnatal month. The rapid replacement of the cells in the cords during fetal life keeps the gland plump and the capsule relatively smooth.
When the cells emerge from the glomerular zone into the cords, and the cytoplasm expands, they assume polyhedral shapes, owing to their compaction. In the peripheral portion of the fasciculoreticular zone, there are six to twelve or even more cells in a cross section of a cord. The cytoplasm has a lightly homogeneous basephilic appearance.
As the cells progress farther toward the center of the gland, the cytoplasm of some cells, without reference to type, becomes acidophilic and then granular. Inclusions in the cytoplasm of most of the cells will now stain with Sudan III. Next, each such cell swells to two to three times its previous size, rounds out, and crowds its neighbors. The nucleus, in the meantime, also enlarges; the globules within it increase in size and break up into numerous smaller ones. Finally, the parts of the nucleoplasm appear to undergo autolysis, and the nuclear membrane gives way, spilling its contents into the cytoplasm. The cell in turn collapses and disintegrates, and the intracellular material disappears. The surrounding cells close in and fill the space, narrowing the cord by that much, so that the volume of all of the cords is constantly being reduced. Many of the cells retain their homogeneous appearance well down the cord, but the vast majority disintegrate, as described above.
It is not in one cell, in a cord, or at any one level that the process goes on, but in many cells at all levels of the cords at the same time. Thus, in the central part of the gland there is considerable variation in the appearance of the cords. In the fetal gland, the whole of the central part of the gland has a light, mottled appearance because of the contrast between the “light” cells, which are actively maturing and disintegrating, and the “dark” cells, which have not yet begun to disintegrate.
Many of the cells in the adult gland approach the center of the gland in cords one or two cells thick. The support for these cords is slight, either from neighboring cords or from the sinusoid walls. The cords become erratic in their direction, and form a much more reticular pattern here than farther toward the periphery. Any one or all cell types may be found in these attenuated cords.
The presence of fascicular and reticular zones, per se, during fetal life, is not a normal condition. The one exception is at the 50- to 60-mm. stage, just previous to the degeneration of the central portion of the gland during the formation of the central veins, when there may be an ill-defined fascieular zone. After 75' mm., the growth of the gland is very rapid in a limited space, and the cords take an erratic course and have a reticular appearance regardless of the plane in which the gland is sectioned. The rapid destruction of cells in the cortex, as described above, leaves few cords intact to reach the veins, and, furthermore, since the medulla, with its interstices between the groups of chromaffin cells, has not yet been formed, there is no place for the residual cords to grow to form a reticular zone. For these reasons, the whole central portion of the gland can be designated the fascicu— loreticular zone. It is this portion of the fetal gland that is now, by general usage, called the fetal cortex. Because it degenerates beginning immediately after birth and is eventually replaced by the fascicular and reticular zones and the medulla, the term is acceptable, particularly because of its long usage in the literature, provided that it is understood that the “fetal cortex” is composed of cells derived from the same sources as the “permanent cortex.”
Beginning a few hours after birth, degenerative changes and marked hyperemia are present in the fasciculoreticular zone. By the end of a week or 10 days, there is almost complete disorganization of the cortical elements central to a relatively wide band of basophilic cells, which includes the capsule and glomerular zone. The gland has shrunk in size to compensate for its loss of substance. By 4 to 6 weeks, the shrinking is almost completed.
The components of the chromaflin system, which had been distributed throughout the fasciculoreticular zone in a retiform manner during fetal life, have become concentrated about the central veins; they will be described under “Development of the Adrenal Medulla.”
The histologic description of the postnatal degeneration of the fasciculoreticular zone was presented by Kern (1911), and has been confirmed by the author. Morphologic and volumetric changes have been elucidated by Lewis and Pappenheimer (I916), Scammon (1925-1926), Keene and Hewer (1927), Benner (1940), and Hunter (1954) The fascicular and reticular zones of the postnatal gland develop from cells in the capsule and glomerular zone during the regenerative period. When the fascicular zone is formed, not all the cells in its cords disintegrate during their passage from the glomerular zone; attenuated cords one to two cells thick continue on into the interstices between the nests of chromaffin cells around the veins. These attenuated cords form the reticular zone. Surrounding the medulla there is an area in which the cords become tortuous, and the intervening spaces are filled with large blood sinuses. The reticular zone hecomes a constant, although a rather poorly organized, feature of the adrenal during the second year of life, and assumes its permanent characteristics by the end of the twelfth year.
In the adult gland, the cells of the fascieular zone and the peripheral part of the reticular zone degenerate by swelling and disintegration. A few of the cells in the cords in the terminal portion of the reticular zone may also degenerate. In general, the cells in the reticular zone have a homogeneous or finely granular cytoplasm similar to that of the cells in the outermost fascicular zone. They may even have a hyalinized appearance. In older individuals, some of the cells near the veins contain brown pigment, the nature of which has not been established to the author’s satisfaction. The usual fate of the cells in the adult reticular zone is a characteristic shrinking of the cell, pycnosis of the nucleus, and phagocytosis of the remains. Except as stated above, these conditions are not found in a normal fetal gland.
Development of the Adrenal Medulla
The development of the adrenal medulla has been the subject of much controversy since the beginning of the century. Wiesel (1902) and Zuckerkandl (1912) have given the classic descriptions. The findings of the present study are consistent with the descriptions by those authors, but a few additional observations may be helpful for a further understanding of the developmental process.
During horizon xiv, the paravertebral sympathetic ganglia begin to increase in size as a result of cell division and the addition of nerve fibers from the rami communicantes. At this stage, there are three types of cells in the ganglia, as differentiated by the character of the nuclei. One type of cell, M-I, is small, with a small, round, compact nucleus, and tends to be located at the periphery of the ganglion. In a second type of cell, M-II, which is not illustrated, the nucleus is larger, and the chromatin is decidedly basophilic and coarsely clumped. A few of these cells have a teardrop nucleus and may have developing axonal processes. A third type of cell, M-III (E, fig. 37, pl. 8), has a large ellipsoid nucleus. The chromatin is scanty, and is evenly distributed in fine particles. The nucleoplasm has a pale lilac tint when stained with Mallory-azan, a characteristic of these cells until they mature into chromaffin cells. They are the “sympathetic formative cells" of \Viesel (1902), the “parasympathetic cells” of ZLlCl(Cl'l{£lI](ll (1912).
During horizons xv and early xvi, all types of cells increase in number. Mitoses are present in the ganglia, markedly increasing in horizon xvi. Late in horizon xvi, nerve fibers begin to appear ventrally and laterally from the ganglia, accompanied by the M-I and M—III cells. The M-I cells are arranged around the nerve fibers; the M-III are sparsely scattered among the fibers. The M-II cells remain in the ganglia and become the sympathetic ganglion cells. The nuclei of the M-I cells change their shape from spherical to compact ellipsoid when they leave the ganglia.
During horizons xv through xviii, the adrenal primordium is a cigar-shaped structure extending from the 6th thoracic to the 1st lumbar segment, lateral to the aorta and mesogastrium. The bundles of nerve fibers and cells, which pass laterally, enter the adrenal primordium as finger-like processes and pass between the future cortical cells, splitting them into islands (figs. 3-5; fig. 15, pl. 2; figs. 16—18, pl. 3). The nerves leaving the ganglia tend to pass caudally and ventrally before branching laterally into the adrenal primordium. As a result, relatively few processes enter the cephalic pole of the primordium; a larger number enter the caudal pole, owing to the presence of the 11th and 12th ganglionic nerve fibers. Most of the nerve fibers and their accompanying cells, from ganglia 6 to 10, take a ventrocaudal course along the lateral side of the aorta, between it and the adrenal primordium, and eventually become the splanchnic nerves. The splanehnic nerves, combined with the fibers from the 11th and 12th ganglia, form a relatively huge mass in the caudal end of the adrenal primordium.
By the end of horizon xvii, the neural tissue has invaded the gland in such quantity that the volume of the adrenal primordium has been greatly increased. In the caudal half of the gland, the cross—sectional area is more than twice that in the previous horizon (figs. 16. 1"-, pl. 3; also p. 198), whereas the cephalic end is only slightly enlarged. The whole plexiform complex is derived from the 6th to the 12th thoracic and usually the 1st lumbar paravertebral sympathetic ganglia. Included in it are the primordia of the celiac, the superior mesenteric, and the renal plexuses, as well as the adrenal medulla.
There is a quiescent period during horizons xviii through xxii in which the increase in cells and nerve fibers is limited principally to the definitive nerves. Between the aorta and the adrenal, the splanchnic nerves are beginning to take form.
During horizon xxiii and continuing through embryos of 50- to 60—mm. crown-rump length, there is a very rapid multiplication of the M-III cells that accompanied the nerve fibers during the initial neural growth in horizons xvi and xvii. These paraganglion cells, as they are called in the literature, are scattered like seeds along the course of the nerve tracts. At first, and continuing until about 80-mm. crown-rump length, they multiply as embryonic cells, but, from 30-mm. crown-rump length until birth, at least, there are always some that are differentiating into chromaffin cells (fig. 37, pl. 8). This transformation is found in all parts of nerve plexuses derived from the primary neural invasion, which began in horizon xvi.
Within the confines of the capsule of the adrenal, terminal nerve fibers have not been seen to penetrate or extend through it, once the capsule has been formed. Primary nerve tracts entering the gland, before encapsulation, particularly in the hilar area (fin. 28, pl. 5), as well as a few other isolated areas scattered over the surface of the gland (fig. 29, pl. 6), maintain their place of entry indefinitely.
In the adrenal, the pattern of distribution of the sympathetic nerves, with the accompanying paraganglion cells, is established by the end of horizon xvii. It consists of a plexus of nerves similar in all respects to the periaortic plexuses. Its extent is, as was stated above, circumscribed by the adrenal capsule. “(hen the rapid multiplication of the paraganglion cells begins in horizon xxiii, the few cells which migrated into the area originally act as nidi from which islands of these cells develop. They are scattered throughout the cortex, without definite organization. During the fetal period, the distribution of the neural and chromaffin elements is in all respects similar to the adult condition in birds (as described by Hartman and Brownell, 1949). They are not oriented with reference to the central veins during the fetal period (fig. 35, 91- 7) The maturation of the paraganglion cells into chromaffin cells begins in horizon xxiii (fig. 37, pl. 8) and continues into the neonatal period.
The degeneration of the central portion of the gland during the first postnatal month of life, so beautifully described by Kern (1911), results in the loss of support for the plexus of nerves which has, up to now, been held in place by the reticulum and cords of the cortex. \Vith this support gone, the neural elements migrate to the nearest stable .structure—the central veins—where they remain.
Development of Blood Supply of the Adrenal
Throughout the late somite stages and through the first half of horizon xvi, a small branch of the postcardinal vein runs in a cephalocaudal direction through the region where the adrenal will develop. It is this vein that Velican (1950) states will become the central vein of the adrenal. He says, also, that the adrenal primordium forms about it as a muff. Carnegie embryo no. 8066, section 9-2-3, horizon xiii, 5.3 mm. (not illustrated), exemplifies the condition of which he speaks. However, under higher magnification, these cells are seen to be no different from those entering the mesenchyme from the celomic epithelium in general. They do not have the characteristics of the cells found in the celomic epithelium between the mesogastrium and the lateral border of the mesonephros during horizon xiv and in the primordium thereafter (fig. I; fig. 8, pl. 1). They may appear massed here because of the mechanical impediment offered by the vein.
This vessel and its branches drain the area until cells from the first celomic invasion become differentiated in the region. The vein loses its identity by the middle of horizon xvi. From then until horizon xviii, the venous drainage of the developing gland is by way of segmental veins paired with the mesonephric arteries. During late horizon xiv, and horizons xv, xvi, and early xvii, the blood enters the gland at the dorsolateral corner from a branch of each mesonephric artery, which breaks up into a globular arborization of capillaries. The capillaries pass between the cords of cells and collect to form a vein for each body segment, which begins near the ventro— medial border of the primordium. These veins pass laterally and dorsally, receiving the bulk of the blood from the adrenal primordium through a few larger capillaries, and enter the postcardinal vein together with the veins from the mesonephros and genital ridge. These veins are the forerunners of the subcardinal—postcardinal anastomosis. The condition is well illustrated in Carnegie embryo no. 8101, slides 25 and 26, horizon xvii (not illustrated).
After the invasion of the neural elements, this vascular arrangement is disorganized, and in late horizon xvii and early xviii a new method of supply and drainage is initiated. At this time the central portion of the gland is made up of capillary blood sinuses and cords of the cells that formerly occupied the area. Nerve cells M-I and M-III are irregularly dispersed among the cortical cells and along the walls of the blood sinuses (figs. 3-5). The vascular channels anastomose freely, and soon become the sinusoids of the fetal cortex.
During horizon xviii (It to 14.5 mm.; Carnegie embryos nos. 6524, 6527, 7707), by proliferation of the cortical cells in the central area and by a rapid accretion from the celomic epithelium and the mesonephric cell nidus, the cortex recovers from the effects of the neural invasion. Reticular cords of cells separated by the sinusoids have formed in the central portion of the gland.
Carnegie embryo no. 7707 (not illustrated) shows the two methods of drainage at this stage; i.e., in each segment blood flows both into the postcardinal veins and into the subcardinals. By comparison with previous horizons, it will be noted in figure 21 (pl. 4) that the subcardinal vein has grown to approximately the size of the postcardinal vein, which, in relation to the other structures, is now smaller. These efferent veins are the beginning of the adrenal veins in the adult. They empty into those portions of the subcardinal veins that will become the inferior vena cava on the right and the adrenal vein on the left. As the gland grows, the vein of exit is located more laterally (compare fig. 24, pl. 5; fig. 31, pl. 7). The blood supply to the sinusoids is still from branches of the mesonephric arteries; the venous return, however, is tending to the now burgeoning subcardinal veins. At first, there are numerous efferent veins, remnants of the postcardinal-subcardinal anastomosis, emptying into the subcardinal veins. The number is rapidly reduced so that by the end of horizon xxiii there are only two or three at the most, but usually only one vein of exit (fig. 31, pl. 7).
By the time this has happened, large central sinuses form an irregular confluence into the vein of exit. A central vein, such as exists in the mammals usually studied, is not found in the human gland, which is trilobate. There are really three central veins and one vein of exit: the adrenal vein. In an embryo of 37 mm. (Carnegie no. 4415, 1061-3; fig. 31, pl. 7), the number of sinuses has been reduced to three main sinuses, one of which runs from posterolaterally, a second from the cephalic pole, and the third from the caudomcdial pole. They coalesce into the large vein of exit. These sinuses are only irregttlar channels until the definitive arrangement has been accomplished as seen in embryo no. 84, 51-2-1, 54 mm. (fig. 32, pl. 7).
The adrenal veins empty into the hepatic portion of the developing vena cava on the right side, as encountered in embryo no. 8553, C3-4-5, horizon xxi (not illustrated); they enter at a much higher level than that described and diagrammed by McClure and Butler (1925). It is the more cephalic branches that are retained by both glands.
In horizon xviii, during the reconstitution of the cortex and the beginning formation of the permanent blood supply and drainage, several of the former mesonephric arteries metamorphose into permanent form. Among them are the adrenal, the diaphragmatic, the renal, and the gonadal arteries. Branches from any or all of these arteries may be retained. Since the gland is growing principally from the ventral surface at this time, the mesonephric arteries are pushed dorsally. The branches that formerly supplied the gland still function,—supplying blood principally from the dorsolateral surface. Gradually, as the major portion of the blood is drawn off from many of the arteries to supply other organs and structures, one vessel gains ascendancy and becomes the adrenal artery while branches of the others remain in an auxiliary role.
This condition prevails in horizon xix. During the next three horizons, the capillaries begin to spread over the surface of the gland, sending branches through the capsule. In horizon xxiii, there is a rapid growth of the arterial vessels and their connections with the sinusoids beneath the capsule, producing a picture like the one shown in figure 27 (pl. 5). A retiform plexus of arterial vessels is formed in and superficial to the capsule of the gland. A few of these vessels eventually differentiate into arteries, but most of them remain capillaries or arterioles as growth and development progress. Vi/hether or not there are arteries passing through the substance of the cortex to anastomose directly with the central veins could not be determined from available material.
This account presents the general relation of the arterial supply, sinusoid circulation, and venous return throughout life, with minor variations due to changes in configuration.
Discussion
Before discussing some of the controversial points in the development of the human adrenal gland, a short resume of the findings during the present investigation is in order.
The earliest evidence of the adrenal primordium is found in horizon xv, when type C-I cells enter the mesenchyme between the aorta and the mesonephros from the level of the 6th to the 12th thoracic segments. During horizon xvi, type C-II cells enter the ventral surface of the primordium from the capsule of the mesonephric glomeruli. A nidus of these cells is formed in the primordium just ventral to each segmental meso nephric artery, from which type C-II cells migrate into the adrenal periphery during horizon xviii to become the capsule in horizon xxii. In horizon xvi, cell type C-III makes its appearance in the primordium by migrating from the celomic epithelium together with type C-I cells.
The dorsal portion of the whole primordium is disorganized in horizon xvii by the invasion of sympathetic nerves and the “parasympathetic” cells of ZLlCl{€I'kandl (1912), while the band of C-11 cells and the celomic epithelium remain intact.
During horizon xviii, the gland becomes reorganized. The C-I and C—III type cells continue to enter the primordium as before; some of these cells situated in the periphery of the primordium begin to multiply, as is evidenced by mitotic figures. The disruption by the sympathetic elements in the previous horizon (xvii) causes a change in the blood supply and drainage in the central portion of the gland. Sinusoids have begun to form and divide the primordium into thick cords. Type C-II cells from the actively proliferating nidus spread around the periphery of the primordium. In horizon xix, the C-11 cells move to the peritoneal surface, and migration of C-1 and G111 cells from the celomic epithelium ceases. By horizon xxii, the type C-II cells have changed to resemble fibrocytes.
During horizon xxiii, the pattern of the arterial blood supply is established; the glomerular zone containing types C-1 and C-III cells is formed, and type C-II cells enter the cortical cords, for the first time, along with the other two types. From this stage on, the changes in the gland are mostly morphological. The source and distribution of the three cell types remain essentially the same throughout life.
Adrenal Cortex
Early in this century, anatomists (Wiesel, 1902; Soulié, 1903; Poll, 1906) described the origin, mode of formation, and perpetuation of the adrenal cortex. Kern (I911) adequately considered the postnatal involution. Soulié (1903) is responsible for the concept of the cortex developing from little buds of celomic epithelium, which resulted from a similar condition found in his researches on lower vertebrates and transferred to higher classes, and which still persists in the literature (Felix, I912; Iordan and Kindred, 1948). From his descriptions, he also may have mistaken the primary tubule formation in the gonad in the human as the source of the epithelioid cells in the adrenal. It is now generally agreed that no buds are found in mammalian primordia (Keene and Hewer, 1927; Uotila, I940).
The cellular composition of the fetal cortex is the same as that of the postnatal or permanent cortex. Having served its function, the fetal cortex degenerates after birth and is replaced by the adult type of cortex, which derives its cells from the glomerular zone and the capsule in the same manner as did the fasciculoreticular zone in the fetus. Until recently, there has not been sufficient evidence concerning the chemistry of the fetal cortex to warrant discussion of its possible functions. After this paper had been outlined, Benirschke, Bloch, and Hertig (1936) and Bloch, Benirschke, and Rosemberg (1956) published a substantial body of evidence which indicates that the early fetal adrenal cortex synthesizes primarily weak androgenic steroids and a sodiumretaining factor similar to aldosterone. C19 steroid synthesis is considered to occur in the fetal zone with decreasing activity as fetal age advances; according to these authors, C-_-1 steroidogenesis is performed by the definitive cortex. They advance the hypothesis that the fetal and reticular zones are sites of adrenal androgen production, whereas the fascicular and glomerular zones. respectively, are the sites of glucocorticoid and mineralocorticoid synthesis. By evidence deduced in the present investigation, the “fetal cortex” is composed of cells originating in the glomerular zone and capsule. No reference is made by these authors to cell types in which lipids are demonstrated. The location and time of appearance of the demonstrable steroids are possibly dependent upon the functional activity of the gland initiated by the pituitary or hypothalamus.
The term “fetal cortex” was introduced into the literature by Keene and Hewer (1927) as descriptive of a structure formed by cells from the celomic epithelium before the stage corresponding to Streeter’s horizon xvii (1951). According to them, the “fetal cortex” is segregated from the celomic epithelium by a band of connective-tissue cells, and from then on is self-perpetuating. Politzer (1938) and Uotila (1940) express the same opinion. Figure 17 (pl. 3) illustrates the condition described by all these authors. In most areas in embryo no. 6758, from which the illustration was taken, as well as other embryos in horizons xvii and xviii, the continuity between the celomic epithelium and the central portion of the gland is retained.
Keene and Hewer (1927), on the basis of four embryos, of 5, I0, 12, and 18 mm., respectively, in the period of the primordium development, postulated a fetal cortex and a true cortex. The 5-mm. embryo showed no adrenal primordium. The Io-mm. embryo was “unfortunately cut obliquely,” so that the relations were distorted. Their description is accurate for this stage. In the 12-mm. embryo, “the cells of the celomic epithelium immediately on either side of the root of the mesentery are no longer columnar and are proliferating, forming a cap of cells 3 or 4 layers thick over the developing gland (better seen in an 18-mm. embryo); some of these cells tend to push posterolaterally between the mesonephros and the suprarenal mass.” This is a fairly accurate description of embryo no. 6742, 11 mm., as shown in figure 4; figure 3, however, shows that the apparent layers of cells posterior to the celomic epithelium are derived from Bowman’s capsule.
Uotila (1940) followed the lead of Keene and I-Iewer, utilizing a larger number of embryos. His statement that the gland is separated from the celomic epithelium at both poles in embryos between 9 and I0 mm. in length is confirmed. Sagittal sections, however, show three-fourths to two-thirds of the length of the gland to be in contact with and receiving cells from the celomic epithelium until the embryo has attained a length of 19 to 20 mm. Uotila also says, “In an embryo of 9.4 mm. a thin layer of mesenchymal cells definitely intervenes between the glandular epithelium and the mesothelium [celomic epithelium so that separation of the gland has been completed.”
In horizon xviii, after reorganization of the cortex of the gland, following its disruption by the sympathetic nerves, cell cords from the celomic epithelium continue to enter the gland. The cords contain both C-1 and C-Ill types of cells, just as they did previous to the neural invasion. It is only in horizon xix, when the C-11 cells move to the surface of the gland, that cells cease to enter the gland from the celomic epithelium. Figures 1] to 25 (pls. 1-5) and figure 36 (pl. 8) illustrate the life history of the type C-II cells from their first differentiation in Bowman’s capsule through their transformation into cells resembling fibroblasts in the capsule and final conversion to cells in the cords of the cortex. During all this time, the nuclei have the characteristics of that in figure 39 (pl. 8), even though the shape varies, except in the capsule, after horizon xx when they are very compact. During horizon xviii, when the migration of type C-II cells from the nidus is at its height, some of the cells enter the primordium of the gonad. These are the cells of which Gillman (19.18) speaks as “an accretion from the mesonephric mesenchyme” that helps form the interstitial cells of the testes. The three types of cells illustrated in figures 38 to 40 (pl. 8) are to be found in the cortex at all times after horizon xxiii in all fetal and postnatal glands. The types are not species specific.
The development of the adrenal is a continuing process from the first differentiation of cells in the celomic epithelium until the adult gland is formed. No interruption of the continuity could be found in the material studied; only hyperplasia and rapid disintegration make the central part of the fetal gland appear different.
For many years, it was thought that the regressive changes in the early postnatal period were limited to primates. Deane (1955) observed in the guinea pig :1 25 per cent decline in volume immediately after birth. A similar decline has been found in the hamster (Crowder, unpublished). Development of the adult type of cortex begins at 4 to 6 weeks postnatally, and is complete by the age of 10 to II years, at which time the fascicular zone assumes the relative proportion of the gland found in the adult.
When cells are lost from the cords, by disintegration or by karyopycnosis, they are replaced by others (cell niigration) from the glomerular zone and capsule, as is suggested by the changes in the cytology of all three types of cells from the periphery.
Whereas Greep and Deane (19497, 19495»), on the basis of their researches, have no firm conviction concerning the concept of cell migration, they do state (194917) that “it is an inescapable conclusion that cells which ultimately come to lie in the inner zone arose from the glomerulosa cells,” to which we add “and capsule cells.”
Reese and Moon (1938) had previously found that ACTH prevented degeneration if given before and after hypophysectomy. These data suggest, in support of embryological findings, that the inner zones are dependent upon cells from the glomerular zone and capsule for their continued existence.
Jones (1948), by means of mitotic counts, found that “Mitoses occur all through the zona glomerulosa with the majority towards the inner border.” Schaberg (1955 , working with adrenal fragments in wrro, found that fragments from the peripheral area reproduced both capsule and glomerulosa with occasional fasciculate-zone cells. He doubts cell migration, but states: “ACTH added to the culture media showed an increase in the number of cells with a vacuolar cytoplasm suggesting an increase in the number of cells which differentiate from glomerulosa into fasciculate cells.”
Two modes of disintegration of the cortical cells are of interest and pose a problem for further investigation. The one: internal histologic differentiation, swelling, and disintegration (p. 201) are found in the fasciculoreticular zone of the fetus and the fascicular and outer reticular zones of the adult gland. The other: hyalinization. shrinking of the cytoplasm, pycnosis of the nucleus, and phagocytosis of the cells are found during the first postnatal month (Kern, 1911, and personal observations) and in the cells of the innermost cords of the reticular zone. It is suggested, subject to further investigation, that the two phenomena have some relation to secretion or the lack of it. In the fetal gland, all the cells in the fasciculoreticular zone disintegrate by the first method, few intact cells reaching the center of the gland, whereas in the adult gland most of the cells in the fascicular and outer reticular zones disintegrate in the same manner. A few of the cells fail to change their internal structure during their passage through the fascicular zone, retaining their integrity in the attenuated cords of the adult reticular zone, where they are removed by the second mode of disintegration.
The question arises: Are these modes of degeneration in any way associated with secretion? It is not in the province of this discussion to consider that question.
Adrenal Medulla
Wiesel (1902), Poll (1906), and Zuckerkandl (1912) have described the development of the neural elements and the chromaflin Cells in the fetal adrenal from to weeks to term. V-Viesel and Zuckerkandl described and named the cells which become the chromaffin cells. \-Viesel called them “sympathetic formative cells,” and Zuckerkandl “parasympathetic cells," a term now generally used. None of the above authors or any others of whom the present author is cognizant describe the embryonic or postnatal phases of development. The development during the embryonic period was described under “Development of the Adrenal Medulla” (pp. 2o2— 203) and illustrated by figure 15 (pl. 2) and figures I6 and 17 (pl. 3).
The author has been unable to find in the literature a description of the movement of the medullary elements to the central portion of the gland and the formation of a definitive medulla during the early postnatal period. Kern (1911) touches upon it, but only in passing.
Other authors (Elliott, 1913; Hollinshead, I936; Swinyard, 1937; Young, 1939) have shown by section of the nerve roots and the splanchnic nerves that the integrity of the nerve supply to the medulla is dependent upon the continuity of spinal roots from thoracic-6 through thoracic-12 or lumbar-1.
It is desirable to think of the medullary neural elements as an integral part of the great abdominal plexus from their first appearance through the fetal and early postnatal periods, since they are both derived from the same sources, pass through the same developmental stages, and have the same cellular characteristics during that time. Examination of sections caudally from figure 14 (pl. 2; embryo no. 792) and Figure 17 (pl- 33 €mbfY0 no. 6758) demonstrates the continuity of the invading sympathetic nerves. Embryo no. 9226, from which figure 28 (pl. 5) was taken, also shows that neural elements in the abdominal perivertebral plexuses are in all respects identical with those in the adrenal. Throughout the fetal period, the neural elements of the adrenal are distributed in a retiform manner among the cortical cells.
Section of the last eight thoracic and first two lumbar nerve roots caused almost complete degeneration of nerves entering the adrenal (Svinyard, 1937), whereas section of the splanchnic nerves (Young, 1939) caused only partial degeneration. The inference is that both the adrenal and abdominal sympathetic plexuses are dependent upon the same nerves.
The sympathetic nerve development has periods of growth with intervening periods of quiescence. The cortical cells in the adrenal appear to have a relatively stable progressive rate of increase. \Vhen the neural elements are actively increasing, they seem to crowd the cortical cords, pushing them aside. During their ltill in growth, the cortical elements fill in, and the gland expands around the nerve tracts. The two elements appear to be in active competition for the same area; phylogenetically, this could be possible. In lower vertebrates, e.g. some of the elasmobranclis, the two components are completely separate entities (Hartman and Brownell, 1949). Hartman and Brownell state that the interrenal and chromallin tissues are usually associated all the way from mere contigtiity to a mingling in the higher forms, and chromallin cells continue to be found in the sympathetic ganglia. In /Irmm, the interrenal tissue has become an organized structure and takes up some of the area occupied by chromallin tissue in lower forms. In reptiles and birds, the cortical substance occupies most of the area between the mesonephros and aorta during the embryonic stage and becomes thoroughly intermeshed with the chromallin cells and nerves. In both classes, the same relation of interrenal and chromafiin tissues continues in the adult. In Orm}/ior/zync/ms, inedullary processes extend into the cortical tissue, sometimes reaching the periphery (I-Iartmari and Brownell, 1949). A similar condition obtains in the opossum, Didelp/ii: virginiczrza. The hilum is very large, and the medulla is a loose network of interlacing strands of nerves and chromaffin cells with interdigitatiiig cords of cortical cells (Crowder, unpublished). In placental mammals, there is a postnatal degeneration of the cortex subsequent to which the definitive medulla is formed, conforming more or less to the shape of the gland.
In the human, after the postnatal degeneration of the fetal cortex, the support afiorded by the cords of cortical cells and the reticulum of the intervening sinusoids is lost, and the medullary tissue gravitates to the central veins. At First, the strands are loosely intertwined with degenerating cortical cells and other detritus in the interstices. By 6 weeks, it has become fairly well organized into a rather compact, highly vascularized structure. VVhen the reticular zone begins to form, at about 12 to 18 months, cords of cortical cells are again seen interspersed among the groups of chromallin cells. Complete adult configuration is attained at about I0 years.
Blood Supply
The earliest organized blood supply to the primordium is a branch from each of the segmental mesonephric arteries, which is given off at the lateral, posterior border of the primordium. Each branch breaks up into a globular arborization, which collects at the anteromedial surface and is carried laterally again in a capillary vein that rtins parallel to the celomic epithelium, just posterior to the layer of type C-II cells, and empties into the postcarclinal vein. The blood supply and return is entirely segmental at this stage.
The pattern is disorganized by the neural invasion in horizons xvi and xvii, to be replaced by the more perinaneiit form. During horizons xviii and xix, the cortex of the gland is reorganized, and the gland as a whole begins to assume a pyramidal shape in which the three central veins take form.
The author interprets the change in form of the adrenal as follows: The active intrusion of the metanephros, beginning during horizon xix, into the area which the adrenal would occupy if it retained its cigarshaped form, causes it to assume a conical shape with a concave base. As growth continues, it is molded by the neighboring structures into a pyramid having a triangular base. Further development produces a lip overhanging the medial surface of the kidney. The morphogenetic forces involved are, of course, obscure.
The arterial blood supply is from the periphery of the gland after horizon xxii. The pressure, therefore, would be approximately equal from all sides, and the blood would accumulate in the center of the gland.
The following quotation from Minot at al. (1912) succinctly sums up the mechanism of the developing blood vessels in the adrenal: “The cause of the early appearance of vessels in a multiple capillary form is to be found in the view that this represents the fundamental method of vascular growth, and that larger vessels only come into existence secondarily when the number of capillaries induces an increased supply of blood. Such an event leads to enlargement of certain fortuitously situated capillaries into arteries and veins. The larger vessels are to be considered in the light of servants of the capillaries, for which they are but the delivering and draining pipes. Consequently, the cause for rich vascularity of a tissue cannot be sought in its possession of larger vessels, but rather in the influences which have brought about a more abundant growth of capillaries in it.” In the adrenal, this influence is the prodigious growth and increase in the number of cells.
From a pyramidal exterior, three main channels to the center of the gland will be formed. The most fortuitously situated branch of the subcardinal veins must then dilate to drain the blood from this central area and thus become the adrenal vein. The tributary veins, as such, do not form until the embryo is 50 to 60 mm. in length, and then only because the shape and mass of the trilobate gland are such that one vein of exit from the three tributary veins is the most efficient.
After the three tributary veins are formed, the line of least resistance for the forming cords of cells to travel, against the pressure of the blood and the adjacent cords, is toward the channels; thus, an approximately trilobate form evolves, with the cords tending toward these veins. Gersh and Grollman (1941) imply, in their discussion of the vascular pattern of the adrenal in the mouse and rat, that the cords of cells conform to the capillary pattern. The converse appears to be true in the present material, for the cords are the dominant feature, and the capillaries conform to the ever-changing cord pattern. There is no change from this blood supply throughout life.
Summary and Conclusions
- In the human, the adrenal cortex derives its cell components from (a) celomic epithelium and (b) the glomerular capsules of the mesonephros during the embryonic period.
- Three distinct types of cells, defined on the basis of nuclear size and structure, are so derived: two from the celomic epithelium, and the other from the mesonepbros.
- The first evidence of the adrenal primordium is demonstrated in horizon xiv early in the 4th week (3 daystz); it consists of type C-I cells.
- Type C-II cells begin differentiation in horizon xv, early in the 5th week. Two groups are found in the medial wall of Bowman’s capsule. The dorsal group forms a nidus in the dorsolateral portion of the primordium beneath each mesonephric artery, from which cells are derived to form the adrenal capsule. At the height of migration from the ventral group, the entire medial wall of each Bowman’s capsule forms new cells which migrate medially, forming a layer 4 to 6 cells thick across the ventral surface of the primordium just dorsal to the celomic epithelium. This layer of cells (C-II) does not interfere with cells of the other types entering the primordium from the celomic epithelium.
- Type C-III cells begin to enter the primordium from the celomic epithelium together with type C-I cells during horizon xvi, early in the 5th week.
- Migration of cells of type C-II from the nidi begins in horizon xvii, about the middle of the 5th week, and by the beginning of the 6th, horizon xxi, a definite capsule is formed around the primordium except where sympathetic nerves are entering the gland.
- When the capsule is formed, cells of types C-1 and C-III, having the same characteristics as those previously seen in the celomic epithelium, are found in the primordium just central to it. They form a layer 10 to 12 cells deep. Mitotic figures are frequent.
- The celomic epithelium has ceased to function as a source of cortical cells by horizon xxi; the cells just beneath the capsule assume that function.
- The invasion of the adrenal primordium by sympathetic neural elements begins about the middle of the 4th week and ends by the first of the 5th week. At this time, activation of the sympathetic ganglia from dorsal-6 to lumbar—1 results in the abdominal sympathetic plexuses, of which adrenal medullary nerves are a part. Growth is relatively slow, keeping pace with body growth from horizon xvii through horizon xxiii. At 7 weeks, a second increase in rate of growth begins, which lasts about 2 weeks; it results in an enormous development of nerve fibers and primitive chromaffin cells.
- Chromaffin cells have begun to differentiate morphologically by 8 weeks. They continue to do so until after birth.
- The so-called “fetal cortex” has the same cellular components as the adult cortex. The difference in appearance in the fetal gland is due only to the increased rate of maturation of the cortical cells in the central portion.
- The fact that mitoses are found in the glomerular and subglomerular regions at all times, but rarely in the other zones, speaks for the continuous production of new cells in the glomerular zone and their migration into the deeper zones.
- The postnatal reticular zone is composed of cells that retained their integrity during their passage through the middle zone and continue to travel within the framework of the cords. The supporting structures of the cords are the walls of the sinusoids. The connective tissue within the sinusoidal walls is continuous from the arteriolar capillaries in the capsule to the central veins. The attenuated cords lack the support of neighboring cords found in the fascicular zone, and, as a result, are forced into an irregular pattern to permit the length to adjust to the perpendicular space available.
- The neural elements and chromaflin cells are found throughout the cortex during fetal life in a retiform pattern in all respects similar to that observed in the adrenals of adult birds. Only after the massive degeneration of the cortex during the 1st postnatal week, when they lose the support of the cortical cords, do they gravitate to the central veins, together with the reticulum of the sinusoids. After the 4th postnatal week, most of the neural tissue and chromallin cells in the adrenal are clustered around the tributary veins. The medulla becomes adult in appearance between 12 and 18 months. In the adult medulla, there are cells among the chromaflin cells, similar in all respects to the paraganglion or primitive chromaflin cells.
Acknowledgments
I wish to thank Dr. George W. Corner, former Director of the Department of Embryology of the Carnegie Institution of Washington, for making the Carnegie Collection available to me and for the many other eourtesies extended to me. Without the thoughtful encouragement, patience, and inspirational criticism of Dr. George VV. Bartelmez, this work could never have come to fruition.
The credit for the photographs used as illustrations is due Mr. Richard D. Grill. The unstinting help given by Miss Harriet L. Caspari, in locating the histologic preparations and making data available, and by Miss D. G. Smith and Mrs. Emilie A. Littleton, in preparing the manuscript, deserve my grateful thanks.
Literature Cited
BENIRSCHKE, K., E. BLOCH, and A. T. I-I1-21:110. 1956. Concerning the function of the fetal zone of the human adrenal gland. Endocrinol., vol. 58, pp. 598-625.
BENNER, M. C. 1940. Studies on the involution of the fetal cortex of the adrenal glands. Amer. Iour. Path0l., vol. 16, pp. 787-798 B1.oe11, E., K. BI-ZNIRSCHKE, and E. ROSENBERG. 1956. Cm steroids, 17a-hydroxycorticosterone and a sodium retaining factor in human fetal adrenal glands. Endocrinol., vol. 58, pp. 626633.
D1aa.\'1=., H. W. 1955. I-Iistophysiology of the adrenal cortex in fetal and prepuberal rats, with comments on the carbonyl reactivity of the cortical lipid droplets. Report, 13111 M. and R. Pediatric Research Conference: Adrenal function in children, pp. 17-26.
E1.1.1o1'r, T. R. 1913. The innervation of the adrenal glands. Iour. Physiol., vol. 46, pp. 285-290.
Felix W. The development of the urinogenital organs. In Keibel F. and Mall FP. Manual of Human Embryology II. (1912) J. B. Lippincott Company, Philadelphia. pp 752-979.
Gsasn, 1., and A. GROLLMAN. 1941. The vascular pattern of the adrenal gland of the mouse and rat and its physiological response to changes in glandular activity. Carnegie Inst. W'ash. Pub. 525, Contrib. to Embi-yol., vol. 29, pp. 111-125.
GILLMAN, I. 1948. The development of the gonads in man, with a consideration of the role of fetal endocrines and the histogenesis of ovarian tumors. Carnegie Inst. “lash. Pub. 575, Contrib. to Embryol., vol. 32, pp. 81-131.
GREEP, R. O., and H. W’. DEANE. 194912. The cytology and cytochemistry of the adrenal cortex. Ann. N. Y. Acad. Sci., vol. 50. 1313- 596615 19495. Histological, cytochemical and physiological observations on the regeneration of the rat’s adrenal gland following enucleation. Endocrinol., vol. 45, pp. 42-56.
HARTMAN, F. A., and K. A. B1<ow:<1—:1.1.. 1949. The adrenal gland, pp. 23-33. Philadelphia.
HOLLINSHEAD, W. H. 1936. The innervation of the adrenal glands. Iour. Comp. Neurol., vol. 64, pp. 449-467.
HUNTER, O. I., I12. 1954. Personal communication.
IoNEs, I. C. 1948. Variation in the mouse adrenal cortex with special reference to the zona reticularis and to brown degeneration, together with a discussion of the “cell migration" theory. Quart. Iour. Microscop. Sci., vol. 89, pp. 53-74.
IORDAN, H. E., and I. E. K1.\'n1t£n. 1948. Textbook of embryology. 5th ed., pp. 220-224. New York.
KEENE, M. F. L., and E. H1:w1=.a. 1927. Observations on the development of the human suprarenal gland. Iour. Anat., vol. 61, pp. 302-324.
KERN, H. 1911. Ueber den Umhau der Nebenniere im extrauterinen Lehen. Deut. med. VVochschr., vol. 37, pp. 971-974.
LANMAN, I. T. 1953. Fetal zone of the adrenal gland; its developmental course, comparative anatomy, and possible physiologic functions. Medicine, vol. 32, pp. 389-430.
L1=.w1s, R. \V., and A. M. PAPPENHEIMER. 1916. A study of the involutional changes which occur in the adrenal cortex during infancy. Iour. Med. Research, Bost., vol. 34, pp. 81-93. (Also quoted in part by Benner, aide supra.)
MCCLURE, C. F. VV., and E. G. BUTLER. 1925. The development of the vena cava inferior in man. Amer. Iour. Anat., vol. 35. pp- 331-383 Mmor, C. S., H. M. Evans, I. TANDLER, and F. R. SABIN. 1912. The development of the blood, the vascular system, and the spleen. In Keibcl and Mall (eds.), Manual of human cmbryology, vol. 2, pp. 581-582. Philadelphia.
Paamzs, A. S. 1945. The adrenal-gonad relationship. Physiol. Revs., vol. 25, pp. 203-254.
Po1.1Tz1~:R, G. 1938. Uebcr die Friihentwicklung der Nebennierenrinde beim Menschen. Ztschr. f. Anat. u. Entwicklungsgesch., vol. 106, pp. 40-48.
PoL1., H. 1906. Die vergleichende Entwicklungsgeschichte der Nehennierensysteme der Wirbeltiere. In 0. I-Iertwig, Handbucl1 der vergleichenden und experimentellen Entwickelungslehre der Wirbeltiere, Bd. 3, Teil 1, pp. 443-603. Iena.
R1=.1:s1~:, I. D., and H. D. Moos‘. 1938. The Golgi apparatus of the cells of the adrenal cortex after hypophysectomy and on the administration of adrenocorticotropic hormone. Anat. Rec., vol. 70, pp. 543-556.
SC.-\.\tMON, R. Ii. 1925-1926. The prenatal growth and natal involution of the human suprarenal gland. Proc. Soc. Exper. Biol. and Med, vol. 23, pp. 806-811.
SCH.-\BERG, A. 1955. Regeneration of the adrenal cortex in vitro. Anat. Rec., vol. 122, pp. 205-221.
SoU1.11'=., A. H. 1903. Recherches sur le développement des capsules surrénales chez les vertébrés supérieurs. Iour. de l’anat. ct dc lc physiol., vol. 39, pp. 492-533.
STREETER, G. L. 1951. Developmental horizons in human embryos. Age groups xi to xxiii. Carnegie Inst. \Vash., Embryology Reprint Volume II. 196 pp.
SWl.\"1'.-\RI), C. A. 1937. The innervation of the suprarenal glands. Anat. Rec., vol. 68, pp. 417-429.
THOMAS, O. L. 1948. The spheroid system of sympathetic neurons with special reference to the problem of neurosecretion. Quart. Iour. Microscop. Sci., vol. 89, pp. 333 350 Plates 1 to 8 are untouched photomicrographs of sections of embryos in the Carnegie Collection illustrating relations and cell
UDTlL;\, U. U. 1940. Early embryological development of the fetal and permanent adrenal cortex in man. Anat. Rec., vol. 76, pp. 183-203.
\='1:L1c.-m, A. 1950. Les phases dc Févolution ontogénétique de la cortico—surréna|e. Bull. histol. appl. et tech. microscop., vol. 27, pp. 79-87. (Abstract in Excerpta Metl., 1951, vol. 5, sect. 1, p. 171.)
W1F,s1=.1., I. 190.1. Beitriige zur Anatomic und Entxt-'ickelu11g der mensehlichen Nebenniere. Anat. I-Iefte, vol. 19, pp. 481- 22.
You-.\'o, I. Z. 1939. Partial degeneration of nerve supply of atlrenal. Study in autonomic innerx-;11i0n. Iour. Anat., vol. 73, pp. 540-550.
ZUCKERKAXDL, E. 1912. The development of the chromaffin organs and of the suprarenal bodies. In Keibel and Mall (eds.), Manual of human embryology, vol. 2, pp. 157-179. Philadelphia.
types of adrenal primordium. Ages are estimated ovulation ages, following Streeter (I951).
Fig. 8. Transverse section of eml)r_vo from beginning of 5th week; midgastric level. Aorta with origin of a mesonephric artery, upper left. Postcardinal vein, upper right. Beneath it, mesonephros with genital ridge on its peritoneal surface. Arrow indicates a transient vessel. Between it and mesonephros, enlarged cells define adrenal primordium. The adjacent celomic epithelium can be distinguished from other mesench}-‘me. No. 3385, 1.;-3-I. Hm‘i’/..oIt xv. X150. 8.3 mm.
Fig. 9. From a section taken about 150 microns caudal to that of figure is’. Primordium differentiated from surrounding mesench_wne by its crowded type C-I cells (see p. 197) with large nuclei and more ahundant cytoplasm. Arrow indicates a mitosis. R. B. (3., red hlo¢_'u‘l eorpuscles marking capillary plexus on medial surface of primordium. No. 3385, 15-I-5. X 500.
PLATE 1
Fig. to. Area enclosed by rectangle in figure 9. All cells of type C-l. Cytoplasm abundant, and nuclei large with relatively little chromatin and an eccentric nucleolus. Arrow indicates nucleus showing highly refractile spheroid body (see lig. 33, pl. 8). Zeiss 3.0 mm. apochromat, ap. I40. ><'. moo.
Fig. II. From a more advanced embr_\-'0, slunvitig priuiordium of adrenal protruding into celom. Mesonephros with Malpighian bodies, tuhules. and Vt-'olllian duct (W. 1).). Increase in size of ganglion of sympathetic chain (I3) due to cell division and ingrowth of nerve lihers. Two groups of cells in l£tm.'|uan's capsule indicated by arrows mark a significant differentiation (see fig. 2: fig. I2, pl. 2“. N0. 721, 14-3-5. Horizon xv. X I50. 4. aorta: 5, celmnic ca\'it_v: (1. mesogastrium: H. stomach: 9. genital ridge: to, adrenal groove: II, postcardinal vein.
Plates
Plate 1

Plate 2

Plate 3

Cite this page: Hill, M.A. (2026, Mayıs 13) Embryology Paper - The development of the adrenal gland in man (1957). Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_The_development_of_the_adrenal_gland_in_man_(1957)
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G