Paper - On the fate of the posterior cardinal veins and their relation to the development of the vena cava and azygos in the embryo pig (1915)
| Embryology - 26 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Sabin FR. On the fate of the posterior cardinal veins and their relation to the development of the vena cava and azygos in the embryo pig. (1915) Pub. No. 223 Contrib. Embryol., Carnegie Inst. Wash. 3(7): 5-32. PDF
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
On the fate of the posterior cardinal veins and their relation to the development of the vena cava and azygos in the embryo pig

{kind=link}
Carnegie Institution of Washington Publication No. 223 (7 plates) 1915
Introducton
During a study of the lymphatic system in the pig, it became evident that a much more exact knowledge of the details of the development of the prevertebral veins was essential in order to trace the origin of the thoracic duct. A knowledge of the neighboring veins is more necessary for the study of the thoracic duct than for any of the superficial lymphatic trunks, since the latter can be so much more readily injected. Besides their relation to the lymphatic problem, the azygos veins have proven to be of great interest on the one hand, in connection with their relation to the fate of the posterior cardinal veins, and on the other, in connection with the larger subject of the origin of the vena cava and the primitive veins of the body-wall.
In the course of this study it has been found that the posterior cardinal veins, which develop in the border of the Wolffian body, disappear entirely with that organ; and that the azygos veins are new prevertebral veins which develop in connection with the vertebra; and subsequently drain the blood from the spinal and intercostal veins as the posterior cardinal veins disappear. It has been shown that the inferior vena cava is not as complex in its development as has been thought. It develops primarily to drain the Wolffian bodies. It has a ventral segment and a dorsal, post-renal segment. The ventral segment comes from two primitive veins, namely, the omphalo-mesenteric and the mesial cardinal (subcardinal) veins. The dorsal segment comes from a prevertebral plexus. This prevertebral plexus is extensive and from it the following veins develop: the azygos and hemiazygos systems, the post-renal segment of the inferior vena cava, and the ascending lumbar veins. This study has been made possible through the use of two methods, first, the injection of the vascular system of embryos, and secondly, the clearing of total specimens, which we owe to Spalteholz.
Technique
The methods of injecting embryos have developed as follows: The first extensive description of the methods of injecting embryonic blood-vessels was given by Popoff in 1894, in a study of the blood-vessels of the yolk-sac of the chick. He did not make a study of the blood-vessels within the embryo. Popoff brought out the fact that notwithstanding the many studies on the blood-vessels of the yolk-sac, it had been impossible to make out the richness of the vascular net without injections. The technique which he developed is as follows: In general he used line glass canulse to puncture the vessel. The camihr were made by drawing out soft glass tubing of a diameter of 0.5 mm., taking rare that the tube should narrow rapidly to a point. To this canula was attached a rubber tube ending in a small mouthpiece. The pressure was made by blowing into the tube and the fluid used was an aqueous Prussian blue. The place of injection varied according to the stage of development and the area desired; the most difficult stages were the early ones before t he marginal sinus was fully developed; from this time up to the fourth day the marginal sinus was used, and Popoff noted that the injection mass might run with or against the blood-stream from the heart, according to the point of injection; he found also that part of the bloodvessels of the yolk-sac might be injected by the action of the heart driving the fluid through the aorta. In chicks, after the fourth day of incubation, Popoff injected directly into the aorta by means of a hypodermic syringe.
The history of the different methods of injecting embryos as they have developed in this laboratory may be of interest. The work was begun by Dr. Mall and has been continued by his pupils. At first the work was a direct development from the methods of making injections of the vascular system in adult animals, and this has been summarized by Dr. Flint (1900) in his article on the adrenal. After injecting the blood-vessels of the adrenal in the adult, Flint applied a similar technique to the vessels in fetal stages, namely, he tied a canula into the aorta and injected Prussian blue by means of a gravity flask. For younger stages, he tied a canula into the apex of the heart, and finally in still younger stages into the umbilical artery or vein. By this last procedure he succeeded in injecting embryos as small as 2.5 to 3 cm. in length.
The next step in the injection of the embryos came in my study of the lymphatic vessels of the skin, which were filled with Prussian blue by means of a fine hypodermic needle. Inasmuch as lymphatic vessels can seldom be seen, good results are only obtained by knowing the level of the lymphatic plexus in order to puncture the vessels. Extensive injections of the lymphatics in stages before the valves develop can best be made by considering the lines of growth, that is to say, by finding the position of larger trunks, which are the primary vessels of an area and which drain an extensive zone. In these studies it became necessary to get the relation of the lymphatics to the blood-vessels, and I found that by puncturing the liver directly with a syringe and filling it with fluid, an injection of the main arteries could be made by means of the heart -beat. Thus was emphasized the necessity of having fresh embryos with the heart still beating. During the same work i 1*K>2) I repeated Budge's injection of the extra-embryonal ccelom and the tissue space connected with it. To make these injections I used a glass canula connected by means of a rubber tube to a pressure fla.sk of a very low column of mercury. This is easily arranged by putting a little mercury into a U-tube, one end of which is attached through a rubber tube to the canula and the other to a rubber bulb which can be compressed by a screw-clamp. These injections were made with India ink, which flows more easily than Prussian blue, since it has a finer granule. This manipulation was watched under a compound microscope, a procedure which was first emphasized by W. G. MacC'allum. Since then all injections of the young embryos have been made with the aid of the binocular microscope.
In 1905 Dr. Mall published a study of the development of the blood-vessels of the brain made from a series of embryos in which the injections were made either through the aorta or by a direct puncture of the liver. In this paper Dr. Mall analyzed completely the method of injecting the blood-vessels of the embryo through the liver, and showed that by this method either the main arteries or the main veins could be injected at will. To fill the arteries he injected a little ink into the liver and got their injection by means of the heart-beat. On the other hand, he obtained the main veins in a dead embryo by an overdistention of the liver. In these injections India ink was used.
Finally complete capillary injections were obtained by H. M. Evans. He found that by injecting India ink into the umbilical arteries with a hypodermic syringe, the needle being pointed toward the embryo, the entire capillary bed of the embryo could be filled. By the aid of this method Evans was able to demonstrate the validity of Thoma's theory of the growth of blood-vessels by showing that tho main arteries of the body could be traced to their antecedent plexuses.
In general embryos of medium size can be injected best with a hypodermic syringe. For these injections the chorionic sac should be opened immediately before the injection is made; until the arnnion and chorion have fused the amnion should be left intact. To make the injections it is essential to see the vessels clearly and to use a fine, sharp needle. The needles should be the finest obtainable, No. 28. Either the artery or the vein may he punctured, the more complete capillary injections always being made through the artery. For embryos from 7 to 15 mm. long it is not necessary to tie off any of tho vessels of the membranes; above this stage it is practically impossible to obtain complete injections without previously tying off the group of vessels of the membranes which are not used for the injection. The vein which accompanies the artery injected gives a sufficient vent for the fluid. The specimen should be kept moist with amniotic fluid. If extravasations occur in the skin vessels, they are readily seen at the time of injection and in general they are sure to occur if the embryo is not perfectly fresh; that is, perfect injections are never obtained unless the embryo is still warm, the heart beating, and there are no previous extravasations of blood due to rough handling. If any ink escapes on the surface of the skin, it must be washed off in running water before fixation, if the specimen is to be cleared in toto.
In the study of complete injections made with India ink, all of the vessels can be analyzed in the early stages; but as soon as the vascular system becomes complex, as in embryos of about 15 mm. in length, it becomes practically impossible to distinguish all the arteries from the veins. This can readily be seen, for example, in an injection such as the one shown in figure 10, plate 4. In these stages one can not determine with certainty given vessels without obtaining injections that will bring out just these arteries, or these arteries with their arterial capillaries or again just these veins. This precision of injection can be obtained by the use of silver nitrate as the injecting fluid. For embryos I use a solution of 0.25 per cent of silver nitrate in distilled water. The silver solution does not run readily. By varying the amount of fluid injected and the point of injection one can obtain either a partial injection of the entire embryo or a partial injection of any organ. The silver solution kills the endothelium and immediately whitens it, so that it is possible to estimate approximately the extent of the injection and to stop it at any desired point. The flow of the silver solution stops as soon as the pressure is taken off, that is to say, the flow of the solution is due entirely to artificial pressure and is not aided at all by the heart-beat nor by the constriction of the blood-vessels. For general arterial injections it is best to inject with a hypodermic syringe into the umbilical artery. For an embryo 20 mm. in length I use about 1 c.c. of the fluid. To fill the arterial capillaries it is only necessary to use more of the solution. For venous injections, since an injection of silver into the umbilical vein usually fills only the veins of the liver, I use a glass canula and inject directly into the primary head vein. If the embryo is placed in a strong light under the binocular microscope, the vein can practically always be made out, especially just behind the ear, by the presence of a little blood in it. An example of such an injection is shown in figure .">, plate _'. As soon as the injection is complete, the embryo should be placed in a bowl of distilled water and exposed to direct sunlight or to the table arc light to reduce the silver nitrate.
In the study of the vascular system, the Spalteholz method for clearing entire specimens, described in his article published in 1914, is invaluable. In general the essentials of the method are, first, fixation in formalin; second, a thorough bleaching of t he tissues with hydrogen peroxide to remove the hemaglobin and other pigments; third, dehydration; and fourth, clearing the specimens in an oil which has the same index of refraction as the tissues. As applied to embryonic tissues, the method, developed under the direction of Professor Spalteholz, to whom I am very much indebted, is as follows: The specimens which have been injected with India ink are fixed for 24 to 48 hours in 5 and 10 per cent formalin. Commercial formalin is slightly acid, which is an advantage for the India-ink injections, since the ink diffuses in an alkaline solution. Specimens which have been injected with silver nitrate are ruined by fixation in formalin, because the silver salt is changed to a white precipitate which obscures the vessels. If injections of bone are desired, the formalin may be made slightly alkaline and the diffusion of the ink prevented as much as possible by tying off all the vessels before fixation. For large fetuses, which are to be cleared in toto, Dr. P. G. Shipley has found that the subsequent bleaching is made easier by washing the specimen in running water before fixation, thus removing much of the hemaglobin. After fixation, the specimens are washed in running tap-water from 12 to 24 hours, followed by distilled water to remove the formalin. The bleaching is done in hydrogen peroxide. Spalteholz adds a few drops of ammonia to precipitate the barium salts. This is not necessary with bariumfree peroxide. For adult tissue, Spalteholz uses undiluted peroxide; for the embryonic tissues about 2 to 3 per cent is the best strength. The small embryos with ink injections take about 20 minutes to bleach ; for the silver specimens, 2 to 3 minutes suffice and they must be watched constantly and the bleaching stopped before the silver is affected. Following the bleaching, the specimen must be washed thoroughly in running water and in distilled water. The dehydration may be begun with 50 per cent alcohol and the percentage increased successively by five points or less. After two changes of a good grade of absolute alcohol, the specimens are passed through two changes of benzene into the synthetic oil of wintergreen. The small amount of benzene which is carried over evaporates quickly, and the few bubbles which develop in the bleaching process can be removed with needles. The
Oil of wintergreen should be entirely colorless, but both the specimens and the oil will gradually become brown with age. This is especially true of the silver-nitrate specimens, but they will keep for six months or a year in oil. They can be returned to alcohol for storage and recleared when desired, or they may be made permanent in balsam. The advantage of keeping the total specimens in oil rather than in balsam is that they can be dissected. On the other hand, they are much more permanent in the balsam. The oil of wintergreen makes the tissues tough, so that it is possible to obtain minute dissections of the injected specimens.
The Spalteholz method as applied to embryos can be very much simplified by changing the fixative. For mammalian embryos the best fixative is Carney's mixture. This is absolute alcohol 60 parts, chloroform 30 parts, and glacial acetic acid 10 parts. In this mixture the acid is sufficiently strong to bleach the hemaglobin so that the peroxide is unnecessary. The penetrating power of the fixative is very great, which is of importance, since no injected specimen can be cut into until it is thoroughly fixed. The relations of the tissues are well maintained and the swelling due to the acetic acid tends to counteract the shrinkage that always takes place in the oil of wintergreen. The fixative does not affect any of the injection fluids. The process after fixation in the Carnoy's mixture is simple; the specimens remain in the fixative from 2 to 12 hours and are then placed direct ly into 70 per cent alcohol, dehydrated in graded alcohols, and cleared as before. The specimens can then be studied in toto, or dissected or embedded in paraffin and sectioned. They should be embedded through a mixture of the oil of wintergreen and paraffin. They do not become brittle in the oil, so that they may be sectioned after staying in the oil for many weeks. The shrinkage in the oil, however, seems to increase on long standing.
The advantages of the fixation in Carnov's mixture are that the .specimens arc even clearer than after bleaching with peroxide, there are no bubbles formed to damage the tissues, the time of the procedure is shortened, and the fixation is much better should it be desired to section the specimens after studying the vessels in whole embryos. Specimens which are strongly pigmented, however, must be bleached with hydrogen peroxide before they can be cleared.
General Analysis of the Primitive Venous System
It has been the universal opinion that the primary veins of the embryo, as distinct from the veins of the embryonic membranes, are two. namely, the anterior and the posterior cardinal. In recent years the idea has developed that the umbilical vein is in part a vein of the membranes and in part a true vein of the body-wall, so that in this double relation it differs from the omphalo-mesenteric veins. It was shown by Miss Helen Smith, in 1909, that in the embryo pig, up to 10 mm. in length, all of the veins of the body-wall ventral to the myotomes drain into the umbilical veins. Moreover, these veins in the body-wall are non-segmental and have no accompanying arteries. In 1909 and 1912 H. M. Evans demonstrated that the primitive limb-bud veins drain not only into the posterior cardinal vein, but into the umbilical vein as well, and thus it is fully established that the umbilical vein is in part one of the primitive veins of the body-wall.
It has been shown that in the chick the first vein of the embryo is the duct of Cuvier, which develops as a direct cross-connection between the aorta and the vitelline veins and that the anterior cardinal, the posterior cardinal, and the umbilical veins develop as a further extension of the vascular arch between the aorta and the venous end of the heart. (Evans, 1909; Sabin, 1915.) Moreover, in the pig a mesial cardinal vein, the so-called subcardinal vein of F. T. Lewis, develops from the lateral branches of the aorta at the same time and in the same manner as the posterior cardinal vein. On account of its relation to the Wolffian body and to the posterior cardinal vein, the name mesial cardinal vein seems to me better than subcardinal. It is therefore necessary to recast the general statement in regard to the primitive veins of the mammalian embryo. The primary venous arch of the embryo is the duct of Cuvier, which connects the aorta with the vitelline veins and makes the first circle for blood within the embryo. Then, in succession, four veins form which may be grouped as the primary venous system of the embryo. First, an anterior cardinal vein by which the capillaries to the brain connect with the duct of Cuvier and establish a circulation for the head; second and third, a posterior and mesial cardinal vein from lateral branches of the aorta to the duct of Cuvier, developing in relation to the Wolffian bodies; fourth, the umbilical veins as an arch between the aorta and umbilical arteries, and the duct of Cuvier. It is clear that the anterior, posterior, and mesial cardinal veins make one system and may be considered as a primary venous trunk connecting with the heart through the duct of Cuvier and definitely accompanying the aorta, while the umbilical vein is in part a vein of the membranes and in part a vein of the body-wall, and is more remotely an accompanying vein to the aorta; that is to say, it forms an arch which is more like the wide arch of the early circiilat ion of the embryonic membranes.
The study of the development of the main veins of the body of the embryo consists. then, in tracing how four veins the umbilical, the anterior, posterior, and mesial cardinal veins develop, or disappear, or are transformed.
Since the work of Goette in 1875, quoted from Hochstetter ( IS9H), it has been known that the development of the vena cava in the liver was determined by a secondary connection of the cardinal svstein with the sinusoids of the liver. In 1901 02 this process was clearly explained by F. T. Lewis, who showed that the connection of the general cardinal system with the liver was not made by the posterior cardinal veins, but by the subcardinal or mesial cardinal vein of the Wolfnan body. He showed that this connecting vein joined the liver through the caval right mesentery. Since the sinusoids of the liver, with which the mesial cardinal vein connects, drain directly into the large remnant of the omphalomesenteric vein, the vena hepatica communis of Hochstetter, the vena cava within the liver is made from two elements, namely, the omphalo-mesenteric veins and certain hepatic sinusoids. Below the liver the vena cava is formed primarily from the mesial cardinal veins with their great anastomosis across the midline.
I can now show that the posterior cardinal vein disappears with the Wolffian body. It does not persist either as a part of the azygos system or as a part of the vena cava, but certain new veins (namely, the thoraco-epigastric and other veins of the body- wall, together with the azygos system, the ascending lumbar veins, and the lower segment of the inferior vena cava) develop to take over the drainage of the spinal cord and the body-wall. The azygos veins are not transformed posterior cardinal veins, but are new prevertebral veins which have their main and permanent connection with the heart through the duct of Cuvier. They grow large because, as the posterior cardinal veins are disappearing, they take over the spinal and intercostal veins. Moreover, the inferior vena cava itself, while it may be considered as a substitute for the cardinal system of veins, is not even in part a transformed posterior cardinal vein. Of the entire cardinal system in the mammal, the anterior cardinal veins persist as the internal jugular veins, the duct of Cuvier persists cither on one or both sides according to the form, but the posterior cardinal veins disappear.
The Veins of the Wolffian Body
General Survey of the Veins
The study of the posterior cardinal veins in this paper will begin with an early stage of the embryo before the tail begins to curve (fig. 1, pi. 1). In the head, only the midbrain bend has taken place. It is a stage measuring from 8 to 9 mm., but is considerably younger than embryos measuring the same after the tail has curved. This stage is about Keibel's figure 12 in the Normaltafeln, Das Schwein; it is before the posterior limb-bud appears. As seen in the figure, the heart has a definite ventricle and atrium, the brain has three vesicles, and there are three gill clefts. A study of still earlier stages shows that both the posterior and the mesial cardinal veins in the pig develop from lateral branches of the aorta and that the two veins are longitudinal anastomoses between these lateral branches, which in turn correspond to the developing tubules of the Wolffian bodies. The two longitudinal vessels are connected by branches determined by the pattern of the nephritic tubules. The posterior and mesial cardinal veins unite in a common stem which joins the duct of Cuvier. (Sabin, 1915.)
It has long been known that the posterior cardinal veins become incorporated early into the Wolffian bodies, and I wish here to emphasize the fact that the mesial cardinal vein (subcardinal) is primarily a vein of the Wolfrian body, so that the study of the cardinal system for the posterior part of the embryo is a study of the circulation of the Wolffian body.
This subject has been extensively studied, notably by Rathke in 1830, and more recent 1 y by Hochstetter, Minot, J. B. MacCallum, F. T. Lewis, Zumstein, and Davis. A study of the veins of the Wolffian body of the pig must be especially based on the work of Minot in 1899, in which the main points are brought out. Injections show that the posterior cardinal vein forms along the edge of the Wolffian body, extends the full length of the dorsal border of the organ, and is continued into the caudal vein at the lower pnle.
As has just been stated, there are at first hut two veins in the Wolrllan body, a posterior and a mesial cardinal, and these form a common stem, the direct, continuation of t Inposterior cardinal which connects with the duct of Cuvier. This condition is found in embryo pigs measuring 6 to 7 mm., when the mid-brain curve is the only curve of the cephalic end of the embryo, and the tail is not curved at all. The next stage in the development of the veins, as seen in figure 1, is characterized by three longitudinal veins, showing still the most important primitive relation of the veins, namely, that the mesial cardinal vein drains into the posterior cardinal. Figure 1 is from an injection with silver nitrate made into the anterior cardinal vein of a pig embryo 9 mm. long. It is clear that, even at this early stage, the mesial cardinal vein, though not as long as the posterior cardinal vein, is distinctly wider. The posterior cardinal vein runs the full length of the Wolffian body and is continued into the caudal vein. The Wolffian body is represented as transparent and the mesial vein is seen through it, lying along the ventro-mesial border of the organ. The mesial cardinal vein is connected by many large superficial mesial vessels with the posterior cardinal. These can be seen as distinct vessels only in the posterior part of figure 1, for in the anterior part they are obscured by the extensive capillary injection within the Wolffian body itself. The mesial cardinal vein curves dorsalward to join the posterior cardinal vein at a point somewhat caudal to the cephalic pole of the Wolffian body.
The third longitudinal vein, the ventral or ventro-lateral, begins at the anterior pole of the organ, where it joins the posterior cardinal vein. At the stage of figure 1 it is the smallest of the three long veins. In its course it follows the path of the Wolffian duct lying just mesial to it. It is represented in the figure as if cut off, but in the specimen it breaks up into capillaries a short distance below the point at which it is shown cut off. It is clear that this ventro-lateral vein is connected by transverse veins with both the posterior and the mesial cardinal veins.
As the tail of the embryo begins to curve and the limb buds develop, certain very important changes take place in the veins of the Wolffian body. These are shown in figure 2, plate 1, from an injection of silver nitrate into the veins of an embryo measuring 8 mm. in its longest diameter, and in figure 3, from a similar injection in an embryo measuring 1 1 mm. The further development of the ventral vein, to the caudal pole of the organ, where it also connects with the mesial cardinal veins, establishes the three longitudinal veins of the Wolffian body. In general these three main veins lie just beneath the capsule, and the posterior cardinal vein has as its primary position the dorso-median angle of the Wolffian body along the edge of the body-cavity, while the mesial vein lies medial to the nephritic arteries and the lateral vein lies just mesial to the Wolffian duct.
The position of these three veins can be recognized in section in figure 6, plate 3, which is from an embryo pig 9.5 mm. long, about the stage of figure 2. In general, such a survey of the vessels of an organ as is given in figure 2 is exceedingly valuable in studying the vascular system by means of serial sections in fact, by an easy method and without the chances of error, it gives the results of reconstruction. For example, if imaginary sections are taken through figure 2, it is clear that a longitudinal vein in one section must appear as a part of a transverse vein at the next level.
The section of figure 6 is through the lower half of the Wolffian body just below the level of the ventral vein as seen in figure 2. It shows the posterior cardinal vein in its primitive position in the dorso-median angle of the Wolffian body and at the edge of the bodycavity. The mesial vein is in the characteristic position mesial to a nephritic artery. The section shows two large transverse veins, one connecting the mesial and posterior cardinal veins and the other along the ventral surface of the organ. The position of the ventral vein is indicated in the legend, but, it is only in a little older stage that it extends so far caudalward (fig. 3). By the time the embryo is 11 mm. long the ventral vein extends the full length of the organ and joins the mesial vein. It must, however, be pointed out that it is only for a short time that the conditions of three complete longitudinal veins (extending throughout the organ as shown in figure 3) remain. By the time the embryo is 12 or 13 mm. long the marked development of the mesial cardinal system as a part of the vena cava has caused a breaking of the longitudinal veins, which remain as the main veins only for the anterior third of the organ.
The Posterior Cardinal Vein
Each of these three veins must now be considered in detail. The dorsal vein, which is the original posterior cardinal vein, lies at first along the dorso-median border of the Wolffian body and receives all the spinal veins. In view of its later history, however, it, is noteworthy that there are variations from this primary position in embryos as early as the 8 mm. stage. For example, in figure 2 the dorsal vein is deflected to the lateral surface for a short distance in the lower half of the organ and then returns to the dorso-medial border. In fact, though the dorsal vein is complete in this specimen, its caudal part was injected through the medial vein. Opposite the area in which the dorsal vein does not lie in the dorso-median angle the spinal veins drain into certain transverse veins. Another more common variation than that shown in figure 2 is that the main dorsal trunk is deflected to the lateral surface for the lower quarter of the organ. These variations explain why it is not always possible to find the dorsal vein exactly in the dorso-median angle of the Wolffian body throughout, even in stages as early as those measuring 8 to 10 mm. In fact, there is a marked tendency for the dorsal vein to be deflected from the dorso-median angle, so that in some specimens measuring 10 mm. the dorsal vein is so much deflected that it receives only the most anterior of the spinal veins of the area corresponding to the Wolffian body. This deflection of the posterior cardinal vein around to the lateral surface of the cephalic part of the organ is found in practically all specimens of embryo pigs about 12 to 13 mm. long. Its final position obliquely across the dorso-lateral surface of the organ is shown in figure 4, plate 1, from an embryo 23 mm. long. The process by which the posterior cardinal vein becomes limited to the cephalic end of the Wolffian body, as seen in figures 4 and .">. is involved in a study of the mesial cardinal vein.
The Mesial Cardinal Vein
The mesial cardinal vein is by far the most important of the veins of the Wolffian body. By the time the embryo is 18 mm. long, it is already the largest vein of the organ. Referring to figure 2, the fluid injected into the anterior cardinal vein ran in three directions into the liver, into the posterior, and into the ventral cardinal vein, while the most extensive injection of the Wolffian body was through the medial cardinal vein by means of its anastomosis with the posterior cardinal vein. The mesial cardinal vein lies obliquely across the mesial surface of the Wolffian body, joining the posterior cardinal about one-third the length of the Wolffian body from its anterior pole. At this stage the most important point in the development of the mesial cardinal vein has been established, namely, that it has formed the connection through the right caval mesentery with the posterior lobe of the liver. The position of this connection is shown in dotted lines in figure 2, the lines being drawn from an injection of another specimen from the same litter of pigs. The development of this connection between the right mesial cardinal vein and the liver is well known through the work of Minot, F. T. Lewis, and Davis. The present work serves to emphasize more strongly the position of Minot, that the vena cava develops primarily to drain the Wolffian both'. The mesial cardinal vein becomes the chief vein of the Wolff inn body and thus the vena cava forms to drain that organ. It is true that, even as early as the stage represented in figure 1, the mesial cardinal vein has tiny tributaries in the mesentery and subsequently the territory of direct drainage into the vena cava is increased through taking over the common trunk of the caudal vein and the primitive fibular vein, but primarily it drains practically only the Wolffian body.
The two mesial cardinal veins anastomose freely across the mid-line ventral to the aorta. At the stage of figure 2, in embryos 8 mm. long, there are two large anastomoses, the anterior one being just caudal to the junction of the mesial and posterior cardinal veins. Subsequently there may be five or six anastomoses, which finally fuse to make the median mesonephritic vein of Minot. This abundant anastomosis accompanies the unilateral development of the vena cava in the right caval mesentery.
The Anterior Mesial Vein of the Wolffian Body
Just at the anterior pole of the Wolffian body there is a short mesial vein lying between the dorsal and the ventral veins (fig. 2). This short vein does not connect with the main mesial or subcardinal vein, but rather with the posterior cardinal vein. It drains the mesial surface of the anterior pole of the organ and is present and constant on both sides. The vein is important because it is constant and remains for a considerable time (fig. 4, plate 1, and fig. 10, plate 4) and hence must be distinguished from the developing azygos vein, which will be brought out later. Its fundamental difference from the mesial cardinal vein is that it retains its connection with the posterior cardinal system and never joins the vena cava, whereas a comparison of figures 2 and 3 shows that the circulation through the mesial cardinal vein is gradually thrown entirely through the liver.
The Ventral Vein of the Wolffian Body
The third longitudinal vein of the Wolffian body, the ventral vein, eventually extends the full length of the organ (fig. 3) and joins the two other veins at the caudal pole, while it joins the posterior cardinal vein at the anterior pole of the organ. In fact, the posterior cardinal vein might be said to bifurcate at the anterior pole of the organ into a dorsal and a ventral branch. This is well known and was figured by Minot. The ventral vein extends along a ridge on the ventral surface of the Wolffian body, in which lies the Wolffian ductIn a general way this ridge marks a boundary between a mesial glomerular zone and a lateral tubular zone, as seen from the ventral aspect (fig. 4); the vein lies just mesial to the Wolffian duct (fig. 6). In early stages the ridge extends along the ventral border of the organ, but as the tubules become convoluted and so take up more space it becomes a curved line on the ventral surface (fig. 4).
The Transverse Veins of the Wolffian Body and the Vena Cava
Having described the longitudinal veins of the Wolffian body, it is now necessary to take up the transverse superficial veins, which are both characteristic and important . These veins have not yet been adequately emphasized, so far as I am aware. The longitudinal veins are connected by two sets of transverse veins, the lateral and the mesial. Both are present from the very beginning of the formation of the posterior and mesial cardinal veins from the lateral branches of the aorta and have from the first a characteristic pattern. The lateral transverse veins connecting the dorsal and ventral veins are small, regular, anil in the beginning alternate with the straight transverse tubules of the organ, so that there are as many transverse veins as there are tubules. An injection of the tubules with silver nitrate gives the negative pattern to that of the transverse veins shown in figure 3. These transverse veins receive many vessels from the depth of the organ (fig. 6) and have tiny anastomoses with each other.
The transverse veins of the mesial surface (fig. 2) connect the mesial vein with the other two. They are always larger than the lateral and are few in number. The importance of the transverse veins is that they develop into the largest tributaries of the vena cava as long as the Wolffian bodies persist. This transition will be readily followed by comparing figures 2, 3, 4, and 5, from embryos 8,11, 22, and 23 mm. long. By means of the marked development of the transverse veins the circulation of the caudal two-thirds of the organ is shifted over to the mesial vein (fig. 5, plate 2). Indeed, the entire relation of the vena cava is foreshadowed in figure 2, from an embryo pig 8 mm. long, in the large anastomosis of the mesial and posterior cardinal veins about the middle of the organ. Within the Wolffian body the lateral veins opposite the median mesonephritic vein become the chief roots of the vena cava, so that from the dorsal view the vena cava appears to bifurcate into these great transverse veins (fig. 14, plate 6). The median mesonephritic vein corresponds in the adult to the vena cava opposite the renal veins. Thus the posterior cardinal vein becomes more and more limited to the anterior pole of the organ (fig. 4), while in the caudal pole only remnants remain connecting the transverse veins (figs. 5 and 14). This change (by which the posterior and ventral veins become limited to the anterior pole of the organ) takes place while the embryo is developing from 12 to 13 mm. in length. The posterior cardinal vein, then, does not become a permanent part of the vena cava, but, disappears as the Wolffian body disappears. This point will not be wholly proved until the subject of the relation of the primitive fibular and caudal veins is taken up in detail. In general, in the stage represented by figure 3, the caudal vein is a continuation of the posterior cardinal vein and the blood from the tail and from the hind limb bud passes through the Wolffian body to the heart, either by way of the posterior cardinal vein or by way of the vena cava. At the stage of 12 to 13 mm. the changes of the circulation within the Wolffian body bring about the condition that almost all of the blood from the tail and hind limb bud passes through the veins of the Wolffian body, chiefly through remnants of the posterior cardinal veins in the caudal half of the organ, to the vena cava; while at the stage of 22 mm. (shown in fig. 5) most of the blood from the tail and hind limb bud passes around the Wolffian body to the vena cava and only a small part of it takes the earlier route through the veins of the Wolffian body. The process by which this shifting of the circulation around the Wolffian body takes place is bound up with the study of the prevertebral plexus of veins, which is taken up later, first in connection with the azygos veins and secondly in connection with the vena cava.
The importance of figure 5 is that it represents all of the permanent fundamental relations of the adult vena cava. The inferior vena cava thus consists primarily of a part within the liver derived from the omphalo-mesenteric vein and certain hepatic sinusoids, and of a part within the Wolffian body derived from the mesial cardinal veins and their tributaries, the transverse veins connected by remnants of the posterior cardinal veins. At first the vena cava drains the tail and hind legs only through the Wolffian body. Subsequently the blood from the caudal end of the embryo is shifted around the Wolffian body and that organ disappears. From this history the vena cava, inasmuch as it arises mainly from the omphalo-mesenteric and mesial cardinal veins, is to be regarded as one of the earliest veins of the embryo rather than as a late development. The final vena cava is made from three main sources: above and within the liver mainly from the omphalomesenteric veins; in the interrenal segment from the mesial cardinal veins; and below the kidneys from the prevertebral plexus. In other words, its ventral segment comes from the omphalo-mesenteric and mesial cardinal veins, while its dorsal part mines from the prevertebral plexus.
The relations of the mesial cardinal vein of the left side and of the vena, cava on the right side to the veins of the esophagus is an interesting one. In studying sections of injected embryos measuring 7.5 to 9.5 mm. long, it is clear that on the right side the vena cava (as seen in figure 2) passes forward a short distance in the caval mesentery and then curves ventralward toward the liver. Just at the point where it curves ventral ward a branch continues forward in the mesentery in the same dorsal position beyond the anterior end of the Wolman bodies and finally anastomoses with the esophageal vessels just below the bifurcation of the trachea. On the left side a corresponding branch, which is much shorter, extends a short distance in the mesentery from the level of the anastomosis of the two mesial cardinal veins, and also anastomoses with the esophageal veins.
The General Circulation of the Wolffian Bodies
At this point it may be well to take up the entire circulation of the WolHian bodies in the stages represented by figures 1 to 5, namely, in pigs measuring 7 to 23 mm. in length. In his article on the veins of the Wolffian bodies, Minot (p. 268) says: "The character of the circulation within the Wolffian bodies is not quite clear, for although the circulation of the blood channels between the tubules with the veins is easily determined, the pathways of the arterial flow are obscure." This point, is entirely cleared up by the silvernitrate injections. The mesonephritic arteries entering the glomeruli are well known. There are 16 pairs of ventro-lateral branches from the aorta, not entirely symmetrical, which enter groups of glomeruli arranged in series along the hilum of the organ. The silver specimens make plain that each glomerulus has two or three efferent arteries emerging from the side or opposite pole of the glomerulus, as can be seen in section in figure (i. These efferent arteries were shown by John B. MacCallum in 1901-02. The vessels are at first capillaries, but by the time the embryo is 20 to 25 mm. long they are distinct arterioles, with a wall thicker than the walls of the veins of the organ. The silvered specimens show that these efferent arteries leave the glomerulus near the point of origin of the Wolman tubules, so that the afferent artery enters the mesial pole of each glomerulus, while its efferent artery and its tubule leave the lateral surface. The efferent arteries enter the characteristic plexus around the tubules which in later stages become a complete cast of vessels, as it were, around the tubules. In embryos 9 mm. long (as is shown in figure 6) the efferent arteries are short, while at 15 mm. they may extend half way across the Wolffian body before breaking up into capillaries. All of the efferent arteries run transversely to the long axis of the organ. The silvered specimens show that the capillaries or sinusoids of the Wolffian body are definite vessels with a complete wall of endothelium, making an excessively complex plexus around the tubules.
The circulation of the blood of the Wolffian bodies is as follows: The blood enters the glomeruli through the afferent arteries at their mesial pole; it leaves the glomeruli through two or three efferent arteries from the lateral surface. These arteries open into a capillary plexus which surrounds the tubules and collects into veins which run transversely to the long axis of the organ. These capillary plexuses (as seen in figure 6) all run transversely, following the pattern of the tubules as they become convoluted, and drain into the transverse and longitudinal surface veins. As is indicated in figure 2, the transverse veins connecting the dorsal and mesial veins are larger than the lateral transverse veins in stages from 8 mm. on, so that there is an early tendency for the capillaries to drain toward the mesial cardinal svstem and thence into the vena cava.
From the stage of about 13 mm. onward, the blood of the anterior third of the organ is collected by three short trunks (a posterior cardinal vein, a ventral vein, and a short anterior mesial vein) and drained into the duct of Cuvier. All of the rest of the blood of the organ passes through transverse veins into the vena cava. The anastomosis of these two areas through the capillary bed is very complete. This stage of the circulation is shown in figures 4 and 5. The complete disappearance of the posterior cardinal veins, as the Wolftian bodies disappear, will be followed in connection with the development of the azygos system. The short trunk of the posterior cardinal vein in the cephalic part of the Wolffian body eventually lies obliquely across the dorsal surface of the organ and is only in its primitive position, just at the anterior pole (fig. 4). Its length is subject to great variation, even on the two sides of the same embryo (as seen in fig. 4). The ventro-lateral vein and the short mesial trunk are likewise limited to the anterior pole of the organ. The system of the vena cava is represented in this figure only by the transverse vein of the left Wolffian body. The relative position of the Wolffian body (in the two stages of 10 and 20 mm.) with reference to the duct of Cuvier is to be seen at a glance by comparing figures 3 and 4. The Wolffian body is gradually dragged caudalward, and hence the part of the posterior cardinal vein anterior to the organ itself is gradually lengthened. Subsequently the anterior pole of the Wolffian body is pushed far to the side by the developing kidney.
From the preceding description it will be seen that the circulation of the Wolffian body belongs to the primitive type that is, it is a circulation in which the veins never accompany the artery, but rather run as far as possible from it. The arteries are in the center of the organ and the veins on the periphery. In the early stages three longitudinal veins make up the cardinal system, draining into the duct of Cuvier. These three veins are very abundantly connected by transverse veins. As soon as the vena cava is established by the connection of the right mesial cardinal with the liver, a marked development of the transverse system takes place by which the circulation of about two-thirds of the organ is shifted from the duct of Cuvier to the vena cava. In this process the posterior cardinal vein eventually disappears.
Azygos and Hemiazygos Veins
General Survey
The azygos and hemiazygos veins are new longitudinal veins which develop in relation to these changes in the circulation of the Wolffian bodies and in connection with changes in the circulation of the body-walls. They are not transformed posterior cardinal veins. The facts following have been emphasized already: that the posterior cardinal vein drains primitively the spinal veins and the caudal veins; that there are two primitive veins of the Wolffian bodies, the posterior and the mesial cardinal ; and that the posterior cardinal system breaks up within the Wolffian body into a number of veins after the mesial cardinal establishes its connection with the liver. The vena cava, which then drains most of the Wolffian bodies and thus subsequently the renal veins and the iliacs, may be considered the substitute for the cardinal system of the Wolffian bodies. The anterior portion of the posterior cardinal system draining the anterior pole of the Wolffian bodies disappears entirely with the Wolffian bodies, and the corresponding spinal veins drain into the new azygos system. The remnants of the posterior cardinal system in the caudal pole of the organ likewise disappear, and the corresponding spinal veins drain into the vena cava and ascending lumbar veins. Confusion has arisen from the conception that the azygos veins and a part of the inferior vena cava were transformed posterior cardinal veins.
The prevailing view in regard to the origin of the azygos veins has developed through the early work of Rathke and especially through that of Hochstetter. Rathke's first idea of the azygos system (1X30, p. (17) seems to have been based on theoretical grounds; he said that in sheep the hemiazygos vein replaced the left inferior vena cava and that in the sheep and the pig the azygos vein did not develop. Subsequently (1X3X) he stated the theory that the azygos and hemiazygos veins were transformed posterior cardinal veins down to the sixth dorsal vertebra, and that below this point they were from longitudinal anastomoses between the intercostals. The work of Hochstetter forms the basis of the modern work on the venous system. His work on the azygos veins was done especially on guinea-pigs and his conclusions are that down to the tenth dorsal segment ( 1X93, p. 574), or, stated in general terms for mammals (1906, p. 145), through the thoracic cavity, the azygos and hemiazygos are transformed posterior cardinal veins, while below that level they are new veins. This new vein he described as formed in the following manner (1894, p. 488): the segmental veins give up their connection with the posterior cardinal vein and become collected into a longitudinal trunk which lies dorsal to the aorta. That is to say, the lower part of the azygos is a longitudinal anastomosis between the roots of the spinal veins. In this connection he noted that the segmental veins were originally ventral to the sympathetic system and subsequently became dorsal. This point will be thoroughly explained in the following pages, as well as the true relation of the spinal veins to the azygos system.
Zumstein was the first to give an entirely correct statement of the origin of the azygos veins (1897, p. 175). In studying the veins of an embryo guinea-pig of about 23 days, he says that near the cephalic end of the posterior cardinal veins two new veins appear, dorsal to the cardinal near the bodies of the vertebrae. They join with the cardinal near the duct of Cuvier and become the azygos and hemiazygos veins. In embryos a little larger he noted that both the cardinals and the more dorsal azygos veins were present, and that subsequently the cardinals became rudimentary. In his figure 9 he shows both the posterior cardinals and the azygos veins in the same specimen.
In the previous year Zumstein had published a study of the relations of the vena cava to the azygos and hemiazygos veins in human embryos and in the adult, in which he took the view that the azygos and hemiazygos veins were transformed posterior cardinal veins.
The following year Parker and Tozier made the next step in advance. In studying the development of the posterior cardinal veins in the pig, they noted that the cardinal veins, after becoming incorporated in the Wolffian bodies, become interrupted within the Wolffian bodies at the stage of 12 to 13 mm., and that the anterior portion of each posterior cardinal vein received blood not only from the Wolffian bodies but from the tissue between. This they found was due to a new outgrowth of vessels from the posterior cardinal veins near the anterior end of the mesonephros. These new accessory veins were the azygos and hemiazygos veins, which they described as having extremely delicate walls (capillaries) without a complete lumen, and as gradually progressing caudalward. The point of not having a complete lumen is, I think, due to a collapsing of the walls of the capillaries, since injections of a complete capillary network are readily made at the same stage. But the discovery of Parker and Tozier that the azygos and hemiazygos are new veins from a new subvertebral capillary plexus is the correct view, and is the main point, They found that these new vessels start from the posterior cardinal veins near the cephalic end of the mesonephros, but the permanent place of union of these new vessels with the cardinal system they thought to be at the level of the tenth rib, so that they finally agreed with Hochstetter that the thoracic part of the vessels came from the posterior cardinal veins and the rest from a new subvertebral plexus. This work of Parker and Tozier corrects the view of Rathke that the azygos veins are longitudinal anastomoses of the intercostals, and the view of Hoehstetter that they are longitudinal anastomoses of the spinal veins, but it is incorrect in regard to the relations of the new azygos veins to the posterior cardinals, for the permanent connections are not opposite the tenth rib, but at the duct of Cuvier on both sides. If both ducts of Cuvier are permanent, both right and left azygos veins open into the heart through them in the adult, as is the case in the pig. If, as in the human, the left duct of Cuvier disappears, the left azygos vein drains into the heart by means of anastomoses with the corresponding vein of the other side.
Since the work of Zumstein and of Parker and Tozier emphasizing the new prevertebral vessels as forerunners of the azygos veins, there have been two different views in regard to the relation of these new veins to the posterior cardinal veins. McClure stated in 1906 that in the marsupials the azygos vein was formed from the new vein plus the proximal end of the posterior cardinal vein. In 1912 Kampmeier expressed the view that, in the cat, the portion of the azygos vein between the duct of Cuvier and the \Yolffian body was made by a longitudinal fusing of the new vein with the posterior cardinal vein. The present study demonstrates that in the pig the posterior vein disappears entirely and does not enter into the formation of the azygos vein.
Circulation of a Typical Body-Wall Segment with Relation to the Azygos Veins
In order to follow the origin of the azygos veins it is necessary to consider the general vascular pattern of the body-wall in the stages that precede them. The azygos veins begin in the embryo pig in stages measuring from 13 to 14 mm. If we consider the development of a typical body-segment up to this stage, it will be remembered that Evans, in his chapter in the Manual of Human Embryology, Keibel-Mall, 1912, brings out the fact that the primitive circulation of a body-wall segment consists of a medial spinal artery, a capillary plexus in the pia, and a lateral spinal vein. In the development of the vascular system of the body -wall in the mammal there are two sets of arteries, a dorsal and a lateral. The dorsal arteries, which are intersegmental, pass to the cord, and establish a longitudinal anastomosis along its lateral surface. The lateral arteries arc not segmental, but also establish a longitudinal anastomosis in the Wolffian groove. This longitudinal vessel becomes connected with the posterior end of the heart and makes the primary venous system. The lateral arteries, except those going to the glomeruli of the Wolffian bodies, then disappear. When the dorsal segmental arteries connect by segmental branches with the cardinal vein the embryonic venous arch of a typical body-wall segment is established and this is the embryonic arch of Evans, dorsal segmental artery, capillary plexus of the pia, spinal vein to the cardinal vein. This arch can not be said to be primary in the sense of origin, since the dorsal segmental vessels do not give rise to the cardinal vein in the mammal. In other words, the first veins in the body-wall of the mammalian embryo are longitudinal chains from the lateral arteries and it is only when the dorsal or segmental arteries connect with this vein that there is a segmental vascular arch. The capillary circulation of the cord consists, as Evans has shown, of a primary capillary plexus along the dorso-lateral surface of the cord, and of a secondary ventral plexus along the surface of the cord in which the spinal arteries pass to the anterior spinal artery. The primitive dorsal segmental arteries fork into a ventral and a lateral branch just between each ganglion and the cord. Besides the two branches of the spinal artery, ventral and lateral, each ganglion receives one or more tiny arteries (fig. <>).
From the standpoint of the development of the azygos veins, the part of the vascular arch of a body-segment that is of most interest is the spinal vein. Moreover, the important point is the position of these veins. As can be seen in figure 6, the spinal veins lie wholly in the sagittal plane passing lateral to the spinal ganglia. The spinal veins collect the blood from an exceedingly abundant capillary plexus around each ganglion, and the vein in each case corresponds to the cephalic surface of the ganglion. This point is clear in figure 4, from a pig 7 mm. long, in my articles of 1913. Thus, from the edge of each spinal ganglion a short, stout vein passes ventral-ward to the posterior cardinal vein (fig. (>) in a plane corresponding to the edge of the body-cavity and distinctly farther lateralward than the posit ion of the future azygos system.
While the primitive spinal arch is being elaborated, the next step is being made namely, the second vascular arch of a typical body-segment is being formed. As can be seen in figure 6, a tuft of arteries from the root of the dorsal segmentals or the adjacent part of the aorta itself grows to a third longitudinal capillary plexus, which extends throughout the body and lies lateral to the myotomes. This capillary plexus, which is shown for the neck region in figures 4 and 5 of my articles of 1913, is very characteristic. It extends from the base of the skull to the tip of the tail. Thus the three sets of arteries give rise to longitudinal chains of capillaries: the dorsal segmental arteries to the pial capillaries, the lateral arteries to the cardinal system, and the third set of dorso-lateral arteries to the capillary plexus along the myotomes. These capillary plexuses are exceedingly significant. For example, the primary capillary plexus along the dorso-lateral surface of the cord lies along the line in which the nerve-cells are multiplying to form the spinal ganglia; secondly, the longitudinal chain from the lateral arteries develops in response to the tubules of the Wolffian body; and, thirdly, the superficial dorso-lateral plexus in response to the myotomes. Throughout the study of the evolution of the vascular system the pattern of the sheets of capillaries is each time determined by the development of the tissues. The lateral vascular arch is not at first a segmental one, for the vein is the umbilical vein. The relation of the capillary plexus along the myotomes to the umbilical vein is shown in figure 6. The lateral arch, then, consists of dorso-lateral arteries which lead through a longitudinal chain of capillaries, along the lateral surface of the myotomes, into veins which are branches of the umbilical vein. In this vascular pattern the umbilical vein is destined to be replaced first by the posterior cardinal vein and permanently by the thoraco-epigastric, the internal mammary, the transverse lumbar veins, and the azygos system.
Two other points of interest may be noted in figure 6, namely, that there are anastomoses between the capillary plexus lateral to the myotomes and the capillary plexus of the cord, and that there is a tiny vein from the myotome-plexus to the spinal vein, so that these two primary vascular arches are already connected at this stage.
To sum up the circulation of the body-wall at the stage of 10 mm., there are in general two vascular arches, a spinal and a lateral arch. The spinal arch is segmental and consists of spinal arteries which lead through three capillary plexuses (namely, the plexus of the lateral surface of the cord, of the ventral surface, and of each ganglion) into a series of segmental veins which connect with the cardinal vein of the Wolffian body. The lateral arch consists of dorso-lateral arteries which drain through a longitudinal capillary plexus lateral to the myotomes, into a series of non-segmental branches of the umbilical vein.
The next stage in the development of the veins of the body-wall is the beginning of the thoraco-epigastric vein, as has been shown by Miss Smith. It is a longitudinal trunk which develops in the capillary plexus of the body-wall and joins the primitive ulnar vein when the embryo is about 12 mm. long. The thoraco-epigastric vein thus precedes the azygos system in development, for the latter does not begin until the embryo is about 14 mm. long. The position of the thoraco-epigastric vein with reference to the body-wall is plain in figure 8, plate 3; figure 10, plate 4; and figure 13, plate 6. It lies just ventral to the capillary plexus of the body-wall and lateral to the ventral border of the myotomes. It is not in the thin lateral wall which contains the veins to the umbilical vein.
Origin and Development of the Azygos System and the Lower Segment of the Inferior Vena Cava from a Prevertebral Plexus
From this pattern of two vascular arches, a spinal and a lateral as a basis, the azygos veins must be followed. It will be noted in figure 6 that there are no vessels between the aorta and the ventral spinal plexus, that is, the area around the notochord is a non-vascular area; moreover, this remains a non-vascular area until the bodies of the vertebrae begin to differentiate. This differentiation begins in the neck region, and by the time the embryo is 1 1 mm. long there is an abundant prevertebral plexus in the neck. This is clearly shown in figure of my articles of 1913. In figure 7, plate 3, a dissection of an injected embryo 14 mm. long, it will be seen that the prevertebral plexus of the cervical region extends into the dorsal region. This figure represents a ventral view of an embryo from which the heart, left Wolffian body, and left posterior cardinal vein have been removed. The right Wolffian body is shown as solid, since the India-ink injection made it so black as to preclude any analysis of its veins.
The cervical and upper dorsal prevertebral plexuses join both thecardinal veins and their tributaries, the spinal veins, in many places. Between these two chains of capillaries are the bodies of the vertebra', so transparent in the specimen that the anterior spinal plexus shows faintly in the drawing. The double chain of capillaries, which is the anlage of the azygos system, shows in cross-section for about the same stage in figure 8, from an embryo 13.5 mm. long, the specimen being slightly further advanced than that of figure 7. This section passes through the upper half of the Wolffian bodies, where the organ is wide (see fig. 7), and shows within the Wolffian bodies only the transverse veins. On the right side of the figure the spinal artery joins the ventral spinal plexus; on the left, the section passes through the full length of a ventral spinal artery to the anterior spinal artery. On the left side the lateral spinal artery is clear. On neither side is the full course of a spinal vein shown, for the section passes through the intersegmental plexus; but on the right side a partial view of a spinal vein indicates their position. The point at which the spinal veins enter the Wolffian body is very clear on both sides, mesial to the edge of the body-cavity. Ventro-lateral to the bodies of the vertebra, along the spinal arteries, is a capillary plexus draining by many branches into the border of the Wolffian bodies, just lateral to the aorta. The section shows no indication of an azygos vein merely the antecedent capillary plexus. It is important to notice the numerous connections in every segment of this capillary plexus with the vessels of the Wolffian body.
This section represents well the point of the change in the body-wall circulation due to the development of the thoraco-epigastric vein; it shows a vein still draining the myotome-plexus into the umbilical, but the new thoraco-epigastric vein lies in the body- wall and makes a very important chain in the lateral vascular arch. The group of the lateral arteries to the myotome-plexus is very plain. The section shows that the thoraco-epigastric vein precedes the azygos system, for the former is a large vein, while the azygos is still a capillary plexus. In this cross-section it can be seen that the sub vertebral capillary plexus lies medial to the spinal vein. Its arteries are tiny branches of the spinal arteries which are best seen in the injections of silver nitrate. On the left side of this section three small veins connect this plexus with the border veins of the Wolffian body just lateral to the aorta. The sympathetic chain lies between the subvertebral or azygos plexus and the spinal veins.
As can be seen in figure 7, the azygos plexus at this stage, namely, at 14 mm., extends only three or four segments into the thoracic region. In this figure it is clear that from the fifth segment caudalward the spinal veins are in a continuous sheet of vessels distinctly lateral to the plexus which is the anlage of the azygos. In this continuous plexus of vessels which enters the Wolffiau body at its dorso-median angle, the spinal veins stand out only because they are a little larger than the rest. This lateral sheet of capillaries and veins is destined to develop very much and to become so important that it should be emphasized from the beginning (fig. 12). It indicates that the veins of the body-wall are far more numerous than the segmental veins. The subvertebral capillaries represent a t hird vascular arch for the body-wall, the first being the spinal, the second the lateral, and the third the subvertebral.
The first figure showing a definite azygos vein which develops out of this capillary plexus is seen in a dissection in figure 9, plate 4. The left Wolffian body, the entire left posterior cardinal vein, and hemiazygos vein are shown in place. The subvertebral plexus is much more highly developed, especially in the cervical region, and in the capillary plexus many definite veins are clearly seen. It is important to note that both the prevertebral cervical plexus and its continuation in the dorsal region have many more connections with the cardinal system than there are segments. The azygos system develops out of that part of the prevertebral system which is in the body region, and of the many connections with the cardinal vein that which is nearest the duct of Cuvier becomes the permanent one. This is true for both sides, and in the pig the duct of Cuvier is permanent on both sides. In the stage represented by figure 9 the duct of Cuvier is high up in the thorax, so that the vessels representing the accessory hemiazygos are a very short group connecting abundantly with the cervical plexus. In general the length of the azygos and of its corresponding superior intercostal is determined simply by the position of the duct of Cuvier. The lower end of the hemiazygos at this stage is opposite the third or fourth dorsal vertebra, where the vein breaks up into a subvertebral capillary plexus.
The azygos system which begins simply in response to the developing vertebrae becomes larger, because it establishes connections with the spinal veins and their new tributaries, intercostal veins. Moreover, this connection with the spinal veins takes place as the posterior cardinal system is becoming less important through being limited to the anterior part of the Wolffian body.
These new relations of the azygos system can be understood by comparing figure 13, plate 6", and figure 10, plate 4, one a dissection of a total specimen, the other a transverse section of approximately the same stage. It has been made clear that the azygos begins as a prevertebral plexus and that its marked increase in size is due to its secondary relations to the spinal and intercostal veins. By comparing figures 6 and 10 it will be seen that there has been a marked development of the veins of the body-wall as well as of the spinal veins. Figure 10 is a section of an injected embryo pig, 16 mm. long, cut transversely 250 M thick. The section passes through the anterior pole of the Wolffian body and shows that part of the organ which drains throughout its history into the posterior cardinal vein. It is well above the point of entrance of the vena cava into the liver (compare figure 2) and its veins do not drain into the vena cava. The posterior cardinal vein and the ventro-lateral vein are both shown. In the median position is the short median trunk, the anterior median vein shown in figure 2 for an earlier and in figure 4 for a later stage. In the series from which figure 10 is taken this vessel is short, measuring not more than 0.7 mm. It ends below by anastomosing with the esophageal vessels; above it joins the ventro-lateral vein and thence the posterior cardinal, figure 4. These short anterior median trunks are constant and can be found as long as the posterior cardinal veins persist. They lie in part ventro-lateral and in part directly lateral to the aorta. They anastomose with each other freely ventral to the aorta, and are connected with the esophageal veins, with the veins of the Wolffian body, and with the azygos and hemiazygos veins. I think them important because they account so readily for variations in the azygos veins, such as the one reported by E. R. Clark (1915).
Turning now to the azygos veins themselves at this stage, namely, 16 or 17 mm., the veins are gradually extending caudalward and for about 5 segments have taken over the spinal and intercostal veins (fig. 13). At the same time it will be noticed that two new longitudinal veins, namely, the internal mammary and the thoraco-epigastric veins, are hot li larger than the azygos vein, and thus carry much the largest part of the blood from the body-wall back to the heart. For the upper thoracic segments, however, the azygos vein now takes much of the blood that formerly passed through the Wolffian bodies, there being only small connections with that organ. Below the level of the fifth segment, as seen in figure 13, the spinal veins and the abundant plexus of the body-wall form a continuous sheet of vessels which enter the Wolffian body lateral to the line of the azygos vein.
Certain relations of the azygos system are only to be made out in sections. As seen in figure 10, the subvertebral plexus is much further developed. There is now a capillary plexus of fine capillaries, mesial to the spinal arteries, resting on the bodies of the vertebra?, while lateral to the spinal arteries is a coarser plexus in which lie the azygos veins. This lateral plexus drains not only the subvertebral plexus, but connects with the spinal veins and with the exceedingly complex plexus from the body-wall as well.
In figure 10 will be noted a space just between each azygos vein and the small vessels from the plexus which enter the Wolffian body just at the edge of the body-cavity in the position of the original spinal veins. In this space runs the sympathetic chain, and thus it is clear that by means of the subvertebral plexus the spinal veins have been shifted from a position lateral to the sympathetic to the azygos veins which are medial to the sympathetic.
From figures 10 and 13 one can readily see that the circulation of the body-wall has now become exceedingly complex as compared with the two simple arches of the stage of 10 mm. Moreover, the pattern is very definite and is repeated for segment after segment. The pattern of each segment is only to be analyzed by following the pattern of the capillaries with reference to the developing tissues, the ribs, the intercostal muscles, and the musculature of the back.
The shifting of the spinal veins to the azygos vein is most plainly shown in figure 11, from an injected embryo 20 mm. in length. Here there is a very definite azygos vein which is shown passing beneath the right posterior cardinal vein. By means of the plexus shown in figure 10, the second, third, and fourth spinal veins, with all the plexus between, have clearly shifted to the azygos vein. The fifth spinal vein still enters the Wolffian body in the more lateral line, while the sixth runs to the azygos vein. The azgyos vein ends in capillaries opposite the seventh segmental vein, which enters the Wolffian body directly. Thus it is clear that the azygos system receives at the stage of 20 mm. about 7 of the segmental veins.
The relations of the rest of the spinal veins at this stage are exceedingly interesting. It will be noted in figure 1 1 that the eighth and ninth join and run to the level of the tenth, where this common stem enters the Wolffian body in the primitive lateral line. This is simply an anomaly of this particular specimen and without especial significance. The position, however, of the tenth, eleventh, the combined stem of the twelfth and thirteenth, and the fourteenth is extremely interesting. From the primitive lateral position in which they entered the dorsal vein of the Wolffian body these vessels have shifted to a more medial position. Their relations can only be analyzed by means of sections. These spinal veins now drain through an excessively complex venous plexus, either directly into the median mesonephritic vein or vena cava, or into its direct tributaries. This is clear in figure 12, from an embryo 20 mm. long. It is this venous plexus which surrounds the cisterna chyli at a slightly later stage and makes it practically impossible to get a pure lymphatic injection Indirect puncture. Returning to figure 11, the fifteenth and sixteenth together and the seventeenth spinal vein still drain through the Wolffian body. Opposite the seventeenth segment is a transverse lumbar vein which drains into the Wolffian body. It is analogous to the thoraco-epigastric vein.
On the other hand, the common stem of the fibular and caudal veins, shown cut off in figure 11, no longer drains mainly into the veins of the Wolffian body as it did in the earlier stages (fig. 3), but has been shifted, by means of the sub vertebral plexus shown in figure 12, to the median mesonephritic vein or the vena cava. The history of this process is best studied in thick serial sections of injected embryos. In embryo pigs up to 19 mm. long the common stem of the primitive fibular and caudal veins still enters remnants of the posterior cardinal vein in the dorso-median angle of the caudal pole of the Wolffian bodies. These remnants of the posterior cardinal vein connect the transverse veins (figs. 5 and 14) and join the vena cava. The caudal vein, however, connects freely with the prevertebral plexus, as shown, for a level farther cerebralvvard in figure 12. When the embryo pig is about 20 mm. long, the permanent kidney begins and lies against the posterior cardinal vein in the Wolffian body and at that time the anastomoses of the caudal vein with the prevertebral plexus develop to such an extent that more of the blood of the caudal veins passes through the prevertebral plexus mesial to the permanent kidneys than through the more lateral primitive pathway through the caudal pole of the Wolffian body. This change in the circulation is shown in the diagram of figure 14, which represents the conditions found in embryo pigs measuring from 20 to 27 mm. In the prevertebral plexus (as seen in figure 12) symmetrical longitudinal trunks develop lateral to the aorta, which shunt the blood from the caudal pole of the Wolffian body to the median mesonephritic vein. The diagram of figure 14 is given to analyze the large vein which represents the bifurcation of the vena cava at this stage. In the mid-line is the anastomosis of the two mesial cardinal veins and this central vein leads on either side into three main dorsal, transverse veins of the Wolffian body. The number of these great transverse veins is not constant, but they drain all but the cephalic pole of the organ. These transverse veins are connected along the dotted line of the diagram by remnants of the posterior cardinal veins, which are marked by crosses. A remnant of the posterior cardinal vein is shown passing ventral to the kidney. These remnants of the posterior cardinal veins (as can be seen in the diagram, or better in the actual sections) lie within the Wolffian body, distinctly lateral to the sub vertebral plexus. They have the same position with reference to the Wolffian body as the posterior cardinal vein in figure 6, and when the Wolffian body disappears they disappear with it, as is indicated by the dotted line on figure 14. Thus the statement of Kerschner (1888, p. 813), referring to the lower segment of the vena cava, is entirely correct: "Bin medial und ventral von den Urnieren (den Niereiu gelagertes Gefass kann den Cardinal Vene nicht entsprechen."
In the stages represented by figure 14 it can be said that symmetrical vena? cava? empty into a large median vein which has remnants of the posterior cardinal vein along its sides; but when the Wolffian bodies disappear, only the median part of this great vein persists as a part of the vena cava; the more lateral part, consisting of the posterior cardinal veins and the transverse veins of the Wolffian bodies, disappears with that organ and hence the inferior segment of the inferior vena cava (which has been thought to be a persistent posterior cardinal vein) is in reality a part of the prevertebral plexus and is more medial in position than the posterior cardinal vein ever is.
The diagram of figure 14 is from an embryo 24 mm. long and shows that the lower segment of the inferior vena cava is symmetrical up to this stage. By the time the embryo is 28 mm. long a very extensive anastomosis across the mid-line dorsal to the bifurcation of the aorta allows a shifting of the blood of the left side to an enlarged trunk on the right side and thus the asymmetrical right trunk of the postrenal inferior vena cava and the ascending lumbar veins are formed.
The significance of these changes associated with the prevertebral plexus lies in the decreasing circulation of blood through the Wolffian bodies all along the line. In the thoracic region the development of the azygos system in the prevertebral plexus has resulted in a shunting of the blood of the spinal and intercostal veins away from the cardinal system and in the abdominal region a much more extensive prevertebral vascular plexus accomplishes the same purpose. In the abdominal region this prevertebral plexus is certainly of great physiological significance. In the first place, it drains practically entirely into the part of the Wolffian body which belongs to the vena cava, and hence it might be called a prevertebral caval plexus. Its beginning is shown in a sheet of vessels in figure 7, from an embryo 14 mm. long, and still better in figure 13, from an embryo 17 mm. long, in which it can be seen that the plexus of vessels coming from the body-wall is very much more extensive than the spinal or segmental veins.
Figure 12 may be taken as a typical section of the prevertebral plexus of the abdominal region. By comparison with figure 10, which gives the prevertebral plexus of the thoracic region, it becomes clear that the abdominal plexus is an extraordinarily complex one. It fills in the entire region between the aorta and the vertebrae on the one hand, and between the aorta and the Wolffian bodies on the other, so that it is not only dorsal to the aorta but lateral as well. In the thoracic region it is the more medial part of the plexus which is best developed (fig. 10), while in the abdominal region it is the more lateral part (fig. 12). That the lateral part of the plexus is more important throughout the abdominal region is clearly indicated in the beginning of its development (as seen in figure 13), in which the two sheets of veins distinctly lateral to the azygos system have already been described. In this plexus the azygos and hemiazygos veins have a characteristic position, directly dorso-lateral to the aorta (fig. 10), while the developing vena cava is farther lateral. From the whole history of the azygos system it is clear that there is a prevertebral plexus throughout the embryo. In the abdominal region this prevertebral plexus becomes excessively complex. It may well be divided into three longitudinal zones a zone cephalic to the median mesonephritic vein, a zone including the mesonephritic vein (fig. 12), and a zone caudal to the same vein. In the first zone, just cephalic to the median vein, the adrenal bodies develop in the area just lateral to the aorta. They, therefore, develop in an excessively vascular area. In the third abdominal zone, caudal to the median vein, the lower segment of the inferior vena cava develops also lateral to the aorta.
This history, then, enables one to complete the analysis of the inferior vena cava from the heart downward. The segment above the liver comes from the omphalo-mesenteric veins; within the liver from the omphalo-mesenteric veins and certain hepatic sinusoids; below the liver from the right mesial cardinal vein of the Wolffian body with its great median anastomosis; and below the kidneys from the prevertebral plexus. It thus becomes clear that the posterior cardinal vein, which early became incorporated in the Wolffian bodies, does not persist as a part of the vena cava. In fact, of the cardinal system below the duct of Cuvier, the only part that remains is the median anastomosis of the mesial cardinal veins making that part of the vena cava which is opposite- the renal veins in the adult; none of the posterior cardinal veins persists. Below the level of the renal veins the vena cava arises from the right prevertebral caval plexus. It connects freely with the veins of the Wolttian bodies as long as these organs persist. It thus becomes clear that the entire prevertehral plexus is an important structure. In the thoracic region the a/ygos system develops in the more medial part of the plexus, dorso-lateral to the aorta; in the lower abdominal region, the third segment of the inferior vena cava and the ascending lumbar veins develop from the more lateral part of the plexus, so that these veins must be considered as arising as longitudinal trunks of the same plexus. Thus a view of the prevertebral veins (such as is given in figure 495 of the Spalteholz Handatlas der Anatomic, Bd. II, Auflage 7, 1913) has this significance, that all of the prevertebral veins shown there, the azygos system, the ascending lumbar veins, and the inferior segment of the vena cava, have a common origin. The origin of the inferior vena cava, from the mesial cardinal veins and the prevertebral plexus, explains the well-known anastomoses of the vena cava in the adult.
No vein has a more extensive or a more interesting literature than the inferior vena cava. Our present ideas in regard to the vena cava may be said to begin with the work of Rathke, and the prevailing theory of the origin of the lower segment of the vein is the theory of Hochstetter. In 1830 Rathke discovered the fact that the lower segment of the inferior vena cava is double and symmetrical in the embryo. This was published in the Merckel's Archiv for 1830. In the same year he stated that the lower part of the inferior vena cava came from the right cardinal vein. His own words, as quoted by Kerschner, are: "Die Vene der rechten falschen Xiere ist die nachherige hintere (untere) Hohlvene, die der linken die nachherige Vena hemiazyga." This view of the inferior vena cava is the same as that of Hochstetter. Rathke, however, in his subsequent work, published in 1838, gave up the theory that the lower segment of the inferior vena cava came from the posterior cardinal vein, because he had specimens in which two vessels were present, namely, a vessel medial to the kidney, making the anlage of the vena cava, and a vessel lateral to the kidney or the posterior cardinal vein (see Kerschner, p. 812). This is the condition shown in the diagram of figure 14. Notwithstanding this observation of Rathke, the work of Hochstetter (1893) has been practically universally accepted. Hochstetter studied the development of the entire vena cava in the rabbit. He described the development of the vena cava in the liver and based the study of the relation of the cardinal system to the liver veins on the work of Goette. It has already been noted that F. T. Lewis has cleared up this part of the subject by showing that it is the mesial cardinal or subcardinal vein which makes the connection with the liver. Hochstetter then developed the theory that the lower segment of the inferior vena cava is a transformed posterior cardinal vein. As can be seen in his figures, especially in his figure 19, Taf. 22, 1893, Hoehstetter's view is merely a different interpretation of the same observations, for, in the same specimen, he shows both an inferior vena cava mesial to the kidney and a posterior cardinal vein lateral to the kidney.
In 1888 Kerschner analyzed the views of Rathke and Hochstetter and concluded that the lower segment of the inferior vena cava could not be a transformed posterior cardinal vein for two reasons: first, the vena cava has a position more mesial and more ventral than the posterior cardinal ever has; and secondly, at a certain stage both veins are present in the same specimen. This sums up the whole question, the facts being that the lower segment of the inferior vena cava is at first double and comes from the bilateral prevertebral plexus and that this new pathway takes the place of the posterior cardinal system which becomes incorporated in the Wolffian body and disappears with that organ. Subsequently the lower segment of the vena cava becomes unilateral. The fact that the lower segment of the vena cava comes from a prevertebral plexus has also been observed by Huntington and McClure (1907) in a study of the development of the veins in the cat. They have called the prevertebral plexus a supracardinal system of veins.
The entire point at issue in the development of the azygos system is the place of its permanent connection with the cardinal system. The belief has been that it lies opposite the tenth vertebra, but as a matter of fact it lies at the duct of Cuvier. The crucial point for distinguishing the azygos veins from the posterior cardinal veins lies in the more dorsal position of the azygos veins. It has been shown that the azygos plexus has a great number of connections with the cardinal system, anterior as well as posterior. At first the cardinal veins are enormous and the azygos veins tiny. At last there comes a stage in which there are two veins of equal size draining into the duct of Cuvier, a dorsal vein or the azygos, and a ventral vein or the posterior cardinal. This is clearly shown in figure 15, plate 6, from an embryo measuring 22 mm. The fact that the azygos vein is farther dorsal than any. portion of the posterior cardinal vein is the crucial point in distinguishing the two veins in sections. This is best shown in figure 5. Since the posterior cardinal vein becomes incorporated in the Wolffian body it necessarily lies as far ventral as that organ. Starting from the stage of from 21 to 22 mm., when the two veins are of the same size, the more dorsal vein (the azygos) increases in size, especially at the anterior end, and the more ventral vein begins to be dragged caudalward, becoming a tributary of the azygos vein. This is well shown in figure 4, from an embryo 23 mm. long. On the right side the dorsal vein (or the azygos) is the only one of the two which connects directly with the duct of Cuvier. Without a figure of the preceding stage in which the two veins are equal (fig. 15, plate (5) it would not be easy to prove from figure 4 alone that the posterior cardinal is really now a tributary of the azygos vein. The point, however, is readily proved from serial sections, because in them it is perfectly clear that it is the more dorsal of the two veins that finally joins the duct of Cuvier. In the specimen from which figure 4 is taken the azygos vein is a distinct vessel up to a point half-way between the two leaders v. A', and v. c. p., where the more dorsally placed azygos vein receives the posterior cardinal vein. The deep shadow on the vein obscures this point in the figure.
The right posterior cardinal vein then joins the azygos in figure 4 at the point where the azygos curves outward to join the duct of Cuvier. Comparing this with figure 15, where the two veins are equal and both enter the duct of Cuvier, it is plain that in the stage of figure 4 the posterior cardinal is being dragged backward, becoming thereby a branch of the azygos. On the left side of figure 4 the hemiazygos and posterior cardinal both join the duct of Cuvier, showing that the left side is not quite as far developed as the right.
The more dorsal position of the hemiazygos is especially clear in figure 4. The fundamental points of the azygos system are all shown in this figure. Both the right and the left azygos veins occupy a dorsal position and empty into the duct of Cuvier. The ladderlike pattern of the two azygos veins, which was noted by Hochstetter, is very clear. Draining into the azygos vein is a small descending vein which I shall call the right oblique vein. Its meaning and further development are clear in figures 17 and 18, plate 7. It drains the prevertebral tissues of the first four segments in the pig and is about as long as the accessory hemiazygos (fig. 17). It is usually larger than the hemiazygos and thus drains more than half of the prevertebral tissue. In figure 16 it is, however, slightly smaller than the hciniazygos. It is not the superior intercostal, which lies farther forward and drains into the azygos at the duct of Cuvier. It is a special branch of the azygos vein characteristic of the pig. Its interest is increased by the fact (hat Kampmeier (1912) thought that it disappeared and was replaced by the thoracic duct. The thoracic duct dues develop along the course of this vein and Kampmeier stated that beyond the stage of 'l'\ mm. (see lig. I > this vein disappeared. A glance at figure 17 (from a pig 28 mm. kmg) and again al figure IS (from a pig 40 mm. long) will show at once that it is on the contrary a developing vein.
The two figures of the stage measuring 2S mm. (namely figures 10 and 17) show a number of important points: first, the relative and marked development of the right oblique vein and the left descending azygos (accessory hemiazygosi ; secondly, the dragging of both right and left posterior cardinal veins along the azygos veins as the \Yolffian body comes to lie farther and farther caudalward. It is now much plainer that the posterior cardinals are becoming tributaries of the azygos system than it was in figure .">. Figure 10 is intended to show that the left descending azygos (accessory hemiazygos) is present alongside of the anterior cardinal to the duct of Cuvier and that it lies dorsal to the anterior cardinal.
The essential characteristics of the azygos veins are also shown in figure IS from a specimen measuring 40 mm. This particular injection shows a variation of the usual pattern in the pig, which is that the right and left veins are about equal, as in figure 17. In figure 18, however, the marked anastomosis of the two veins, which is pretty constant opposite the eighth to the tenth ribs (see fig. 17), has become very large, and below that point a single trunk represents both veins.
The development of the azygos system makes its nomenclature very simple: On either side the azygos opens into the duct of Cuvier. According to the position of the duct of Cuvier there is a short descending branch, the so-called superior intercostal and accessory hemiazygos, and a long ascending branch, the so-called azygos and hemiazygos. For the pig, where both ducts of Cuvier are permanent, the only nomenclature necessary is ascending and descending right and left azygos veins, together with the special branch, the right oblique vein.
Conclusions
This study has demonstrated that the posterior cardinal veins disappear with the Wolffian bodies. They are not transformed either into a part of the vena cava or into a part of the azygos veins. They develop along the edge of the Wolffian body and disappear with that organ. The vena cava is primarily a vein to drain the Wolffian bodies, and, being formed from the omphalo-mesenteric vein within the liver, and from the mesial cardinal vein below the liver, may be said to be foreshadowed in the primitive venous system of the embryo. Subsequently, as the Wolffian bodies disappear the lower segment of the vena cava develops from a prevertebral caval plexus by which the caudal veins and veins of tinlegs become shifted from the Wolffian bodies to the vena cava.
The azygos veins are new prevertebral veins, representing a vascular arch between the spinal arteries and the cardinal system of veins. They drain into the anterior and the posterior cardinal veins by very numerous connections. Of the many connections the ones which remain as the permanent ones are those opposite the ducts of Cuvier. and hence no part of the posterior cardinal vein is incorporated in the azygos system. The position of the duct of Cuvier in the adult determines the length of the ascending and descending branches. The more dorsal position of the azygos system throughout its development enables one to distinguish it from the cardinal system at every stage. The development of the azygos system enables one to simplify its nomenclature into right and left ascending and descending azygos veins.
Bibliography
Online Editor - Some references in this list require online addition (draft).
Clark ER. An anomaly of the thoracic duct with a bearing on the pig embryology of the lymphatic system. 8. Washington, 1915. (Carnegie Inst. Washington. Pub. No. 222.)(draft)
Clark EL. Injection and reconstruction of the jugular lymph sac in the chick. Anat. Record, Philadelphia, 1912, vi, 261-264. (draft)
Clark JG. The origin, development, and degeneration of the blood-vessels of the human ovary. Contrih. sc. nied. . pupils of W. H. Welch, Baltimore, 1900, 593-6711, 5 pi. Also: Johns Hopkins Hosp. Rep., Baltimore. 1900, ix, 5915-670, 5 pi. (draft)
Dandy WE. A human embryo with seven pairs of somites measuring about 2 mm in length. (1910) Amer. J Anat. 10: 85-109.
Davis DM. Studies on the chief veins in early pig embryos and the origin of the vena cava inferior. Amer. Journ. Anat., Philadelphia, 1910, x, 461472, 6 pi. (draft)
Evans HM. On the earliest blood-vessels in the anterior limb-buds of birds and their relation to the primary subclavian artery. (1909) Amer. J Anat. 9: 281-319.
Evans HM. On the development of the aortae, cardinal and umbilical veins, and the other blood vessels of vertebrate embryos from capillaries. (1909) Anat. Rec. 3: 498-518.
Keibel F. and Mall FP. Manual of Human Embryology II. (1912) J. B. Lippincott Company, Philadelphia.
Die Entwicklung des Blutgefasssystems. Handb. d. Entwcklngsgesch. d. Menseheu, (Keibel und Mall). Leipzig, 1911, II, 551-688. Also, trains!.: Man. Human Embryol. (Keibel and Mall), Philadelphia, 1912, II, 570-709.
Flint JM. The blood vessels, angiogenesis, organogenesis, reticulum, and histology of the adrenal. Contrib. sc. med. pupils of W. H. Welch, Baltimore, 1!)00, 153-228, 8 pi. Also: Johns Hopkins Hosp. Rep., Baltimore, 1900, ix. (draft)
GOETTE, A.: Die Entwicklungsgcschichte der Unke. 8. Leipzig, L. Voss, 1875.
GRAFE, E.: Beitrage zur Entwicklung der Urniere und ihrer Gefiisse beim Hiihnchen. Arch. f. mikr. Anat., Bonn, 1906, LXVII, 143-230, 5 pi.
GROSSER, O.: Die Elemente des Kopfvenensystems der Wirbeltiere. Anat. Anz. (Verhandl. d. anat. Gesellsch.), Jena, 1907, xxx, 179-192.
HOCHSTETTER, FH.: Ueber die Bildung der Hohlvpne bei den Saugethieren. Anat. Anz., Jena, 1887, n, 517-520.
Beitrage zur Entwicklungsgeschichte des Venen systems der Amnioten. Morphol. Jahrb., Leipzig, 18S7-8, xiii, 575-585, 1 pi.
Zur Morphologic der Vena Cava inferior. Anat. Anz., Jena, 1888, in, 867-872.
Ucber die hintere Hohlvene. Verhandl. d anal.. Gesellsch. (Wien), Jena, 1892, vi. 181.
Beitrage zur Entwicklungsgeschichte des Venen ~y>tems der Amnioten. u, Reptilien. Morphol. Jahrb., Leipzig, 1892-3, XIX, 428-501, 3 pi.
Beitrage zur Entwicklungsgeschichte des Venen systems der Amnioten, in, Sauger, Morphol. Jahrb., Leipzig, 1893, xx, 543-648, 3 pi.
Entwicklung des Venensystems der Wirbelthiere. Anat. Hefte, 2 Abt.. Wiesbaden, 1894, in, 460-489.
Die Entwicklung des Blutgefasssystems. Handb. d. vergleich. u. exper. Entwicklungslehre d. Wirbelthiere (Hertwig), Jena, 1903, HI, 2 Teil, 1-166.
HOFFMANN, C. K.: Zur Entwicklungsgeschichte des Venensystems bei den Selachiern. Morphol. Jahrb., Leipzig, 1893. xx, 289-304, 1 pi.
Huntington GS. The genetic principles of the development of the systemic lymphatic vessels in the mammalian embryo. Anat. Record, Philadelphia, 1910, iv, 399-424, 18 pi.(draft)
Huntington GS. and McClure CFW. Development of the postcava and tributaries in the domestic cat. Anat. Record. Philadelphia, 1906-7, I, 29. Also: Amer. Journ. Anat., Baltimore, 1906-7, vi, Suppl.,29.(draft)
JACOBSON. L, L. : Sur 1'anatomie et la physiologic d'un systeme veineux, particulier aux reptiles. Bull, de la Sue. philomat., Paris, 1812-13, in, 259-262.
Kampmeier OF. The development of the thoracic duct in the pig. Amer. Journ. Anat., Philadelphia, 1912, xin, 401-464, 5 pi. (draft)
KERSCHNER, L.: Zur Morphologic der Vena Cava inferior. Anat. Anz., Jena, 1888, in, 808, 943.
KNOWEU, H. McE.: A new and sensitive method of injecting the vessels of small embryos, etc., under the microscope. Anat. Record, Philadelphia, 1908. 11, 207-214.
Mall FP. On the development of the blood-vessels of the brain in the human embryo. (1904) Amer. J Anat. 4:1-18.
Lewis WH. The development of the arm in man. (1902) Amer. J Anat. 1(2): 145-184.
MacCallum JB. Notes on the Wolffian body of higher mammals. (1902) Amer. J Anat. 1(2): -259.
MACCALLUM, W. G.: Die Beziehung der Lymphgefassc ziim Bindegcwebe. Arch. f. Anal. u. Physiol., Anat. Abt., Leipzig. 1902.273-291. .l/.s.i. Ininxl.: Johns Hopkins Hosp. Bull., Baltimore, 1903, xiv, 1-9.
McClure CFW. A contribution to the anatomy and development of the venous system of didelphys marsupialis (L.) Part II. Development. (1906) Amer. J Anat. 5(1): 79-94.
McCLURE, C. F. W.: A contribution to the anatomy and development of the venous system of Diilili>lui* marsupialis (L). Amer. Journ. Anal., Baltimore, 1906, v, 163-226, 5 pi. The postcava of an adult Inili:iti chevrotain (Tragu lus meminna Erxleben). Anat. Anz., Jena, 1906, xxix, 375-377.
MILLER, A. M.: The development of the postcaval vein in birds. Amer. Journ. Anat., Baltimore, 1911 23, u, 283-298.
Minot CS. The veins of the Wolffian body in the pig. (1898) Proc. Bost. Soc. of Nat. Hist, 28: 265-274.
On a hitherto unrecognized form of blood circulation without capillaries in the organs of vertebrata. Proc. Boston Soc. Nat. Hist., 1900, xxix, 185-215.
PARKER, G. H., and C. H. TOZIER: The thoracic derivatives in the posterior cardinal veins of swine. Bull. Mus. Compar. Zool. Harvard College, Cambridge, 1897-98, xxxi, No. 6. 133-144.
POPOFF, DEMETRIUS: Die Dottersack-Gefässe des Huhnes. 8. Wiesbaden, C. W. Kreidel's Verlag, 1894.
RAUL, C. : Ueber die Entwicklung des Veni'iisyslcms der Selachier. Festschr. z. 70, Gcburtstage Leuckarts, 8. Leipzig, W. Engelmann, 1892.
RATHKE, H. : Ueber die friiheste Form und die Kntwirkluim de Venensystems und der Lungen beim Schafe. Arch, f . Anal. u. Physiol. (Meckel's), Leipzig, 1830, v, 63-73.
Abhandlungen zur Bildungs- und Entwicklungs Geschiehte des Menschen und der Tiere. Erster Theil. 4. Leipzig, F. C. W. Vogel, 1832.
Der Bau und die Entwicklung des Venensystems der Wirbelthiere. Bericht ii. d. Natunvissen-cli Seminar d. Univ. Konigsberg, 1S3X. in. 168-170.
SMUV, F. R.: On the origin of the duct of Cuvier and the cardinal veins. Anat. Record, Philadelphia, 1915, ix, 115-117.
Der Ursprung und die Entwickelung des Lymph gefasssyslem. Ergebn. d. Anat. u. Entwcklngsgesch., Wiesbaden, 1913, xxi, 1-98. Also trmisl.: The origin and development of the lymphatic system. Johns Hopkins Hosp. Rep., Monographs, New Series, No. 5, Baltimore, 1913, 1-94.
Smith HW. On the development of the superficial veins of the body wall in the pig. (1909) Amer. J Anat. 9: 439-462.
SPALTEHOLZ, W.: Ueber das Durchsichtigmachen von menschlichen vmd tierschen Praparaten. 8 Leipzig, S. Hirzel, 1911. Second edition, 1914.
TILNEY, F. : The development of the axial veins and lymphatics in Tragulus meminna Erxleben. Amer. Journ. Anat., Philadelphia, 1912 13, XIH, 193-221, 13 pi.
Zi'MsTEiN, J.: Zur Entwicklung des Venensystems bei dem Meerschweinchen. Anat. Hefte, 1 Abt., Wiesbaden, 1*97, viii, 165-190, 10 pi. Zur Anatomic und Entwicklung des \Yurn-ystems des Menschen. Anat. Hefte, 1 Abt., Wiesbaden, 1895, 1895-6, vi, 571-608, 2 pi.
Explanation of Plates
- Pig posterior cardinal veins
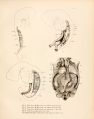
plate 1
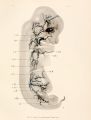
plate 2
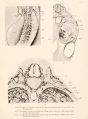
plate 3
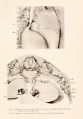
plate 4
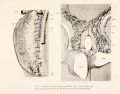
plate 5
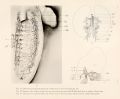
plate 6
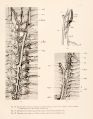
plate 7
Plate 1

Fig. 1. Embryo pig 9 mm long (about the stage of fig. 12 in Kcibel's Xonnaltafcln, Das Schweini, in which tinveins of the right Wolffian body have been injected with silver nit rale. This is a stage before the tail has curved and hence the specimen is younger than the one of fig. 2, which measures less. Cleared by the Spalteholz Method. Xll.
v. c., v. caudalis; v. c. M., v. cardinalis mesialis at the point at which it curves dorsalward to join the v. cardinalis posterior; v. c. p., v. cardinalis posterior; v. T. L., v. transversa lateralis of the Wolffian body; v. T. M., v. transvevsa mesialis of the WolfIian body; v. v., v. ventralis of the Wolffian body.
Fig. 2. Dissection of an embryo pig 8 mm long, in which the veins have been injected with silver nitrate through the anterior cardinal vein. The left Wolffian body has been dissected away so that the mesial surface of the right Wolffian body is exposed. Cleared by the Spalteholz Method. Xll.
v. c. P., v. cardinalis posterior; v. c. M., v. cardinalis mesialis at the point of the anastomosis of the veins of the two sides, which is also the point where the v. cardinalis mesialis dextra curves ventralwanl to the liver; v. M. A., v. mesialis anterior of the Wolffian body; v. OM., oniphalo-mcsenterica ; v. s., v. spinalis; v. T. M., v. transversa mesialis of the Wolffian body; v. u., v. umbilicalis; v. v., v. ventralis of the Wolffian body.
Fig. 3. Embryo pig 11 mm long, in which the veins have been injected with silver nitrate, showing the left Wolffian body and through it the right mesial cardinal vein. Cleared by the Spalteholz Method. Xll.
v. c. M.. v. cardinalis mesialis; v. c. p., v. cardinalis posterior; v. F. P., v. fibularis primitiva; v. T. M., v. transversa mesialis belonging to the left Wolffian body; v. T. L., v. transversa lateralis of the Wolffian body; v. u., v. umbilicalis; v. v., v. ventralis of the Wolffian body.
Fig. 4. Dissection of an embryo pig 23 mm long, in which the veins have been injected with silver nit rate. Cleared by Spalteholz Method. X10.
D. c., ductus Cuvier; v. A., v. azygos; v. A'., v. azygos above the 1 point where it is joined by the v. cardinalis posterior; v. c. P., v. cardinalis posterior; v. c. v'., v. cardinalis posterior within the Wolffian body; v. I., anastomosis which is the forerunner of the v. innominata; v. M. A., v. mesialis anterior of the Wolffian body; v. M. I., v. mammaria interna; v. TE., v. thoracoepigastrica; v. T. L., v. transversa lateralis making the mainroot of the vena cava within the Wolffian body; v. u. p., v. ulnaris primitiva; v. v., v. ventralis of the Wolffian body; w. B. i., Wolffian body, inner gloinerular zone.
Plate 2

Fig 5. Embryo pig 22 mm. long, in which the veins have been injected with silver nitrate, and the entire embryo cleared by the Spalteholz Method.
Xll. v. A., v. azygos; v. B., common stem of the v. fibularis primitiva and the v. caudalis which empties into the median mesonephritic vein and which becomes the lower segment of the vena cava inferior; v. c., v. caudalis; v. c. A., v. cardinalis anterior or v. jugularis interna; v. c. P., v. cardinalis posterior; v. c. i., v. cava inferior; v. F. p., v. fibularis primitiva; v. J. E., v. jugularis externa; v. L. T., v. lumbahs transversa; v. M. i., v. mammaria interna; v. T. L., v. transversa lateralis of the Wolffian body making the main roots of the vena cava; v. TE., v. thoraco-epigastrica; v. u. p., v. ulnaris primitiva; w. B., Wolffian body.
Plate 3

Fig. 6. Transverse section of an embryo pig 9.5 mm long in which the entire vascular system has been injected with India-ink through the umbilical artery. The section is unstained and is 250 M thick. It passes through the lower half of the Wolffian body and is to be compared with fig. 2. X36.
A., aorta; o. s., position of a ganglion spinale; v. c. P., v. cardinalis posterior; v. c. M., v. cardinalis mesialis; v. T. M., v. transversa mesialis of the Wolffian body, connecting the v. cardinalis posterior with the v. cardinalis mesialis; v. T. v., v. transversa ventralis of the Wolffian body, the leader being at the position of the v. ventralis at a little later stage; v. s., v. spinalis; v. u., v. umbilicalis; w. D., Wolffian duct.
Fig. 7. Dissection of an embryo pig 14 mm long in which the vascular system has been injected with India ink through the umbilical artery. The left Wolffian body has been removed, the right one is in place. X20.
A. s., a. spinalis; D. c., ductus Cuvier; P. P., prevertebral plexus of veins from the body-wall and from the spinal cord entering the Wolffian body in a sagittal plane lateral to the v. hemiazygos, the lower part of which plexus becomes a prevertebral caval plexus; v. A., v. azygos in the form of its antecedent capillary plexus; v. M. I., v. mammaria interna; v. j. E., v. jugularis externa; v. TE., v. thoracoepigastrica.
Fig. 8. Transverse section of an embryo pig 13.5 mm long in which the vascular system has been injected with India-ink through the umbilical artery. The section passes through the cephalic half of the Wolffian body and is to show the prevertebral capillary plexus in which the v. azygos and the v. hemiazygos develop. The section is unstained and is 250 M thick. It is about the stage of the specimen in fig. 7. X40.
A., aorta; A. s., a. spinalis; P. v. A., plexus in which the vena azygos develops; v. u ., vein of the body-wall; v. TE., v. thoraco-epigastrica; v. s., v. spinalis; v. T., v. transversa of the Wolffian body; x., plexus of capillaries lateral to the myotome.
Plate 4

Fig. 9. Dissection of an embryo pig 16 mm. long in which the vascular system has been injected with India ink through the umbilical artery. The specimen shows the prevertebral plexus of the cervical and upper dorsal region with the developing v. hemiazygos. X1S.
D. c., ductus Cuvier; v. HA., v. hemiazygos; v. J. K., v. jugularis externa; v. TE., v. thoraco-epigastrica.
Fig. 10. Transverse section of an embryo pig 16 mm. long in which the vascular system has been injected with India ink through the umbilical artery. The section passes through the cephalic pole of the Wolffian body and is to show the relation of the v. azygos to the prevertebral plexus. The section is unstained and is 2. r iO /u thick. It is about the stage of fig. 13. X34.
A., aorta; v. c. p., v. cardinalis posterior within the Wolffian body; v. HA., v. hemiazygos; v. M. A., v. mesialis anterior of the Wolffian body; v. M. I., v. mammaria interna; v. TE., v. thoraco-epigastrica; v. v., v. ventralis of the Wolrnan body; w. B., Wolffian body.
Plate 5

Fig. 11. Dissection of an embryo pig 20 mm. long in which the vascular system has been injected with India ink through the umbilical vein. All of the viscera have been removed. X20.
A., aorta; v. A., v. azygos; v. B., common stem of the v. fibularis primitiva and the v. caudalis which becomes the lower segment of the vena cava; v. o. p., v. cardinalis posterior; v. L. T., v. lumbalis transversa; v. s. 2, v. spinalis of the second segment; v. TE., v. thoraco-epigastrica; v. u. P., v. ulnaris primitiva.
Fig. 12. Transverse section of an embryo pig 20 mm. long in which the vascular system has been injected with India ink through the umbilical artery. The section is to show the prevertebral plexus and passes through the median mesonephritic vein. The section is unstained and is 250 n thick. X77.
A., aorta; A. s., a. spinalis, which is now a branch of a single dorsal segmental artery; p. P., prevertebral plexus at the position of the future azygos vein, the limit of the development of the v. azygos for this stage being shown in fig. 11; p. p'., prevertebral plexus at the point in which the third segment of the inferior vena cava and the corresponding ascending lumbar vein develop at the level below the median mesonephritic vein; p. p. v., prevertebral plexus of small veins resting on the vertebra; v. M. M., v. mediana mesonephritica, the anastomosis across the mid-line of the embryo of the two vv. cardinalis mesiales; v. T. M., v. transversa mesialis of the Wolffian body.
Plate 6

Fig. 13. Dissection of an embryo pig 17 mm. long in which the vascular system has been injected with India ink through the umbilical artery. X21.
A., aorta; D. c., ductus Cuvier; p. p., prevertebral plexus which receives veins from the body-wall and from the spinal cord and drains into the Wolffian body in a sagittal plane lateral to the hemiazygos vein; v. HA., v. hemiazygos; v. M. I., v. mammaria interna; v. TE., v. thoraco-epigastrica.
Fig. 14. Diagram from the embryo pig measuring 24 mm. long to show the relations of the symmetrical segments of the inferior vena cava to the median mesonephritic vein and to the remnants of the posterior cardinal vein in stages measuring from 20 to 27 mm. long. The parts of the large median vein marked with a cross are from the posterior cardinal vein. The dotted lines separate the part of the median vein which persists from that which disappears with the Wolffian body.
A., aorta; K., kidney; v. c., v. caudalis; v.c.i., vena cava inferior; v.c. i. L., vena cava inferior, lower segment; v. F. p., v. fibularis primitiva; v. M. M., v. mediana mesonephritica; V.T..V. transversa of the Wolffian body.
Fig. 15. Dissection of an embryo pig 20 mm. long, a little younger than the specimen of figs. 4 and ."i. The specimen has been injected with silver nitrate through the veins and is to show the stage at which the v. azygos and the v. cardinalis posterior are equal in size and both empty into the duct of Cuvier. XS.
D. c., ductus Cuvier; v. A., v. azygos; v. c. A., v. cardinalis anterior or v. jugularis interna ; v. c. I., vena cava inferior; v. c. P., v. cardinalis posterior; v. j. E., v. jugularis externa; v. TE., v. thoraco-epigastrica; w. B., Wolffian body.
Plate 7

Fig. 16. Injection of the veins in an embryo pig 28 mm. long to show the relation of the v. hemiazygos accessoria to the caudal part of the v. cardinalis sinistra which still persists at this stage. X17.
D. c., ductus Cuvier; v. A., v. azygos; v. c. A., v. cardinalis anterior; v. c. P., v. cardinalis posterior; v. HA., v. hemiazygos; v. HA. A., v. hemiazygos accessoria; v. o., oblique vein which is a branch of the v. azygos; \v u ., Wolffian body.
Fig. 17. Dissection of an embryo pig is mm. long in which the vascular system has been injected with India ink through the umbilical artery. The specimen shows the azygos and hemiazygos veins and their relation tn the disappearing vv. cardinales posteriores. X20.
v. A., v. azygos; v. c., common stem of the v. ulnaris primitiva and the v. thoraco-epigastrica; v. c. A., v. cardinalis anterior; v. c. P., v. cardinalis posterior; v. HA. A., v. hemiazygos accessoria; v. o., oblique vein which is a branch of the v. azygos; w. B., Wolffian body.
Fig. 18. Dissection of an embryo pig 40 mm. long in which the vascular system has been injected with India ink through the umbilical artery. View of the v. azygos and v. hemiazygos. X26.
v. A., v. azygos; v. HA., v. hemiazygos; v. HA. A., v. hemiazygos accessoria; v. o., oblique vein which is a branch of the v. azygos.
Cite this page: Hill, M.A. (2024, April 26) Embryology Paper - On the fate of the posterior cardinal veins and their relation to the development of the vena cava and azygos in the embryo pig (1915). Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_On_the_fate_of_the_posterior_cardinal_veins_and_their_relation_to_the_development_of_the_vena_cava_and_azygos_in_the_embryo_pig_(1915)
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G