Category:Carnegie Embryo 109: Difference between revisions
mNo edit summary |
mNo edit summary |
||
| (22 intermediate revisions by the same user not shown) | |||
| Line 1: | Line 1: | ||
This | [[File:Sabin1909 fig04.jpg|thumb|alt=Carnegie Collection Embryo No.109|Carnegie Collection Embryo No. 109]] | ||
This {{Embryology}} category shows pages and images that relate to the Carnegie Collection Embryo No. {{CE109}}. This embryo was classified as [[Carnegie stage 18]] occurring during [[Week 7]]. | |||
:'''Links:''' [[Carnegie stage 18]] | [[:File:Keibel Mall 2 488.jpg|Keibel Mall Fig 488]] | [[:File:Keibel Mall 2 489.jpg|Keibel Mall Fig 489]] | [[:File:Keibel Mall 2 491.jpg|Keibel Mall Fig 491]] | [[Carnegie Collection]] | |||
{{Carnegie stage 18 links}} | |||
{| class="wikitable" | |||
|- | |||
! Serial No. !! Size (mm) !! Grade !! Fixative !! Embedding Medium !! Plane !! Thinness (µm) !! Stain !! Semi. ducts !! P.-M. duct (mm) !! Year !! Notes | |||
|- | |||
| {{CE109}} || E., 12.0* Ch.,30 || Poor || Alc. || P || {{Transverse}} || 20 || Al. coch. || 1 || 0.4 || 1897 || {{Tubal}}. Least—advanced third | |||
|} | |||
{{Carnegie Collection stage 18 table}} | |||
===References=== | |||
{{Ref-BardeenLewis1901}} | |||
{{Ref-Lewis1902}} | |||
{{Ref-Bardeen1905a}} | |||
{{Ref-Sabin1909}} | |||
{{Ref-Streeter1906ear}} | |||
{{Ref-WhiteheadWaddell1911}} | |||
{{Ref-Lisser1911}} | |||
{{Ref-Mall1915}} | |||
==Limb== | |||
{{Ref-Lewis1902}} | |||
Embryo [[:Category:Carnegie Embryo 109|'''CIX''']] measures V. B. 10.5 mm. and X. B. 11 mm. in lengtli and is about five weeks old. There is a marked advance over the preceding stage. Cartilage has made its appearance both in the vertebrae and in portions of the arm skeleton. There is considerable difference in the character of the cartilage of the vertebra from that in the arm. The latter seems more advanced and lias more the appearance of true hyaline cartilage. It is possible that the cartilage appears first in the arm, though I have not been able to examine intervening stages to determine this with certainty. Other portions of the arm skeleton are in the precartilage and condensed tissue stages. Both cartilage and precartilage are surrounded in most places by a distinct perichondrium. This takes a very deep stain with the alum carmine. This perichondrium shades off into the condensed tissue of the carpus, which is like that composing the skeletal core in the preceding stage. This again shades into the even less differentiated tissue of the digits, which is at a.bont the same stage of development as the hand plate of the preceding stage, and it in turn shades off into the surrounding mesenchyma. | |||
The muscles in the arm region show very different degrees of development. Those derived from the muscle plate system are in advance of most of the others. The trapezius, levator scapulae and serratus magnus are about as far advanced as those from the muscle plate system, they show distinct muscle fibers and are for the most part quite sharply limited from the surrounding loose mesenchyma. In position they correspond with their premuscle masses of the preceding stage. The pectoral muscle is next in advance and the latissimus dorsi next. These two muscles grow from the humeral region towards their future attachments on the body wall. It is this portion which lies farthest from the humerus which seems to show the most advance in fibrillation and the sharpest limitation from the surrounding mesenchyma. At the humeral end these muscles gradually shade into a condensed mesenchyma, which fuses with neighboring muscle and skeletal elements. Both muscles correspond in position to their premuscle masses of the preceding stage. As in the preceding stage, embryo CLXIII, the trapezius and serratus premuscle masses were in advance of the pectoral and latissimus; in embryo CIX we find the same relation still continues. | |||
The remaining muscles of the arm apparently develop in situ from the premuscle sheath and undergo practically no migrations. They do not appear to be as far advanced as any of the above mentioned muscles. Of these muscles developing from, the arm premuscle sheath, the more proximal ones are more developed than the ones more distal. In the scapulo-humeral region most of the muscles show partial fibrillation, while those in the palm of the hand are in about the same condition as the proximal portion of the premuscle sheath in the preceding stage. | |||
The fibrillation, position and nerve supply have made it possible to determine the presence of most of the muscles of the arm. | |||
===The Skeletal, System=== | |||
====The Vertebral Column==== | |||
The intervertebral discs are composed of condensed mesenchyma, the cells having a concentric arrangement about the chorda. The vertebral bodies between the discs are each composed of two masses of cartilage, one on either side of the chorda. Tliey are surrounded by a perichondrium. Along the ventral surface of the vertebral coluuni is a layer of dense mesenchyma, which probably represents both perichondrium and the anterior common ligament. The neural processes, composed of condensed mesenchyma, are clearly defined. They are continuous with the discs and form a wide, shallow groove for the spinal cord. The transverse processes arise by two roots, one from the base of the neural process and the other from the disc. They are of condensed mesenchyma. | |||
The Ribs. - The ribs are more sharply defined than in CLXIII. They are of condensed tissue except for a small area near the head, which is of precartilage. They extend farther into the body Avail than in the preceding stage. | |||
'''Fig. 9.''' Skeleton of the arm region of embryo CIX, lateral view. X 12 diameters. | |||
====The Arm Skeleton==== | |||
The Scapula is composed of precartilage and has | |||
greatly altered in shape. It lies in the region of the lower four cervical | |||
and first one or two thoracic vertebrae. From the anterior border, which | |||
corresponds to the spine, springs the large curved acromion process. | |||
On the median surface at the junction of the humerus with the scapula | |||
arises the large hooked coracoid process. Eunning across the median | |||
surface of the scapula to the vertebral border is a slight ridge which | |||
separates the supraspinatus from the subscapularis muscles and corresponds to the future anterior border. The condensed tissue is thickened | |||
on the medial surface into a perichondrium, while on the lateral surface | |||
the precartilage shades off into the surrounding mesenchyma. | |||
====The Clavicle==== | |||
A rather poorly defined mass of condensed tissue continues from the tip of the acromion toward the tip of the first rib, extending for about one-third this distance. This mass represents the clavicle. From it a mass of ill-def]ned tissue extends to the coracoid process and represents the coraco-clavicular ligament. | |||
The Humerus is directly continuous with the scapula and root of the coracoid process. No signs of joint surfaces or cavity are present. Both ends of the shaft are enlarged and Ihe distal end shows both external and internal condyles. The core of the shaft is of hyaline cartilage; this is surrounded by very thick perichondrium, which shades ofE into the condensed tissue of each end in which is enclosed an area of precartilage. The distal end seems more advanced than the proximal. | |||
The Radius and Ulna are continuous with the distal end of the humerus, no indications of joint surfaces or cavities being present. | |||
'''Fig. 10.''' Outline of the arm region of embryo CIX, lateral view from Plate IV. Bardeen and Lewis, Vol. I, No. 1, this Journal. X 12 Diameters. | |||
There is more flexion at the elbow than in CLXIII. The forearm occupies a position about half way between pronation and supination. The core of each shaft is composed of hyaline cartilage. This is surrounded by a very thick perichondrium, which continues into the condensed tissue at either end of the bone, in which precartilage is enclosed. The hand-plate is continuous with the distal ends of the radius and ulna. It is composed of condensed mesenchyma. There are several centers of increased condensation which I believe must correspond to the carpal bones, namely, the scaphoid, lunar, pyramidal, trapezium, trapezoid, OS magnum and unciform. The scaphoid is in line with the radius and the lunar with the ulna, while the pyramidal is at the ulnar side of the carpus, and as the metacarpal V continues from it more than the uniform the whole hand has a peculiar bend toward the ulnar side. From the carpus five masses of condensed tissue project. They shade off into the surrounding mcsenchyma which fills the distal end of the arm. The condition of these finger masses corresponds to the condition of the hand-plate in CLXIII. There is not the sliglitest indication of segmentation into metacarpals and phalanges. The radial of the five projections probably consists of both trapezium and metacarpal I, which have not yet shown signs of separate centers of condensation. | |||
===The Muscular System=== | |||
The muscle plate system has become differentiated into several muscles, namely, the deep dorsal muscles, the intercostals, the abdominal muscles and the deep ventral neck muscles. | |||
The infrahyoid muscles correspond in position and nerve supply with the infrahyoid premuscle mass of the preceding stage. They extend nearly to the region where the median end of the clavicle will eventually extend. | |||
The trapezius muscle has extended posteriorly to the level of the fifth cervical vertebra. Its posterior end lies near to the lateral surface of the body and is connected to the tips of the neural processes as far posteriorly as the second thoracic vertebra by a considerable interval of fascia. As the muscle passes anteriorly it lies deeper and deeper from the surface, being separated from it by the platysma and facial muscles. Its ventral border is free from attachment to the scapula and clavicle. At the level of the second cervical vertebra it is joined by the stemomastoid muscle, which has ascended from the more ventral neck region. The nerve supply is as in the adult. | |||
The rhomboid mass lies in the region of the Y and A^I cervical vertebrae. It connects with the fascia passing to the dorsal tips of the neural processes but has no scapular attachment. A branch of the fifth cervical nerve supplies it. | |||
The levator scapula; and serratus anterior muscles form a continuous fibrillated mass, extending from the first cervical vertebra to the ninth rib. It occupies much the same position that its premuscle mass did in embryo CLXIII except that the posterior end now extends to the ninth rib. Digitations go to all the cervical transverse processes and to each of the anterior nine ribs. The anterior and posterior digitations are very slender and contain but few fibers. The thickest part of the- muscle lies in the scapular region. There is no scapular attachment. The ventral edge of the muscle lies at about the same level as the dorsal edge of the scapula but in a more median plane. Branches from the second to the seventh cervical nerves supply the muscle. The first three penetrate directly into the muscle. The last three form a trunk which runs along the lateral surface of the muscle as far as the fourth rib. | |||
The pedoralis major and minor "^ are united into a common muscle mass, which is well differentiated from the surrounding tissue. It forms a thick oval mass, which extends from the level of the second rib to the proximal portion of the humerus. The greater part of the muscle thus lies anterior to the first rib. As the mass bends towards the humerus it is attached also to the clavicle. So probably both sterno-costal and clavicular portions are present. The median side of the mass bulges towards the coracoid process and represents the minor. Most of the mass shows distinct fibrillation, but toward the humerus this passes into the condensed tissue which is not sharply outlined from the surrounding structures. The position of the pectoral muscle corresponds to the position of the pectoral premuscle mass in embryo CLXIII. Branches from the median side of the brachial plexus supply the pectoral. Two from the external cord contain fibers from the fifth, sixth and seventh cervical nerves. Two come from the inner cord. Within the muscle complicated anastomoses occur from which fibers spread out in all directions. | |||
The muscles thus far considered were fairly definite, and, as we have seen, come from quite definite premuscle masses. The remaining muscles of the arm are in process of differentiation from the arm premuscle sheath. The exact limits of the individual muscles are almost impossible to determine. | |||
The deltoid muscle extends from the acromion and clavicle and fascia over the infraspinatus to the humerus. It is very closely connected with the infraspinatus and only by the difference in the nerve supply can the two be separated. The position of the teres minor is also only indicated by its nerve and not by any line of separation between it and the infraspinatus or deltoid. The origin of part of the deltoid from the acromion and clavicle helps to distinguish some of its fibers, but a short distance from this origin no line of separation can be made between it and the infra- and supraspinatus muscles. Condensed tissue connects it with the triceps and pectoral muscles. The circumflex nerve supplies this muscle and also sends a branch to fibers which are closely associated with the infraspinatus and probably constitute the teres minor muscle. | |||
That portion of the infraspinatus which lies on the lateral surface of the scapula is fairly distinct except where the deltoid and teres minor muscles join it. The portion of tlie supraspinatus on the anterior onefourth of the median surface of the scapula is distinct, but after it passes the acromion it is inseparably' connected with the infraspinatus and deltoid and pectoral muscles. These muscles shade off into the 23roximal end of the humerus. The main portion of each of these muscles contains muscle fibers. The suprascapular nerve supplies the supra- and infraspinatus muscles. | |||
23 Lewis, Observations on the pectoralis major muscle in man, Johns Hopkins TTosp. Bui., Vol. XII, 1901. | |||
The subscapularis muscle arises from the posterior one-half of the median surface of the scapula and passes beneath the coracoid process to the humerus. The circumflex nerve separates a portion of it from the teres major muscle, but the scapular portions of the two are closely imited, as is also the long head of the triceps. A branch from the circumflex and another from the posterior cord of the brachial plexus supply the subscapularis. | |||
The teres major and latissimus dorsi muscles are closely associated at "their humeral end. The latissimus dorsi' lies in the lateral thoracic region, extending posteriorly as far as the fourth rib. It has no attachments to the ribs or vertebral column. The two muscles are inserted together into the proximal portion of the humerus. The teres major arises from the axillary border of the scapula near its posterior angle. The common portion of the latissimus and teres passes close to the posterior cord of the brachial plexus, from which a large branch is given off that runs into the latissimus and has a brush-like ending near the posterior limit of the muscle. A smaller branch of the posterior cord is given off to the teres major. | |||
The triceps muscle extends along the posterior and lateral surfaces of the humerus, extending from the scapula to the ulna. Indications of the three heads are present. The portion of the muscle lying near the insertion of the latissimus dorsi and the infraspinatus muscles is not sharply defined from them. The musculo-spiral nerve passes through the muscle and gives branches to it. | |||
The biceps and coracohracJnaUs muscles lie along the median side of the humerus, extending from the coracoid process to the radius. The two heads of the biceps are quite closely united nearly to their origins, Avhich are but a short distance apart. The portion of the coracoid process from which the long head arises must ultimately become a portion of the head of scapula. The attachment of the coracobrachialis to the humerus is by condensed tissue, as is the distal end of the biceps to the radius. The distal end of the biceps blends with the brachialis and the flexor mass. The musculo-cutaneous pierces this group and gives off branches to it. | |||
The brachialis muscle is closely attached to the distal one-half of the hmnerns over the anterior and median surfaces. It is also closely attached to the overlying biceps muscle and it is impossible to determine just the line between the two or between it and the brachioradialis muscle. It is also impossible to determine the exact line between the muscle and the underlying perichondrium. It is closely associated with the triceps on one side and the deltoid on the other. The main portion of the muscle is fibrillated and is inserted into the ulna by condensed tissue, which is closely associated with the, flexor mass of the forearm. The musculo-cutaneous nerve gives off a large branch which has a brush-like endins; within the muscle. | |||
'''Fig. 11.''' Outline of the arm region of embryo CIX, median view, from Plate V. Bardeen and Lewis, Vol. I, No. 1, this Journal. X 15 diameters. | |||
The flexor muscle mass of the forearm forms a thick layer over the median surface of the ulna, radius, carpus and proximal end of the metacarpus. It is with considerable difficulty that I have separated this mass into two layers. The superficial layer is smaller in extent and lies in the proximal region of the forearm. It is connected with the radial portion of the forearm by a condensed tissue mass and distally fuses with the deep layer to become continuous with the condensed tissue of the digits. The median nerve passes through the proximal portion and then comes to lie between the two layers. From its position and relation to the median nerve I believe this to be the layer from which the flexor carpi radialis, flexor sublimis digitorum, pronator teres and palniaris long-us muscles differentiate. Branches from the median nerve supply this layer. Both layers arise partly from the inner condyle of the humerus, and are continuous more or less with the muscles of the upper arm. The deep layer is closely attached to the perichondrium of the forearm and hand. It is wider in extent than the superficial and shows indications of separations into muscles. Tlie portion for the flexor carpi nlnaris shows most advance. The extension into the hand probably constitutes the portion from which the interossei and lumbrical muscles and flexor tendons develop. It is continuous with the condensed tissue of the digits. The portion on the forearm forms the flexor profundus digitorum, flexor pollicis longus, flexor carpi ulnaris and pronator quadratus muscles. Both the ulnar and median nerves supply the deep layer. | |||
The extensor mass of the forearm is farther advanced than the flexor. It can be differentiated into three groups of muscles which accord well with the adult groups. The first group, the largest and most superficial, extends from the lateral condyle to the proximal ends of the digits, where it blends with the condensed mesenchyma. It is a thin layer and spreads out over the ulnar two-thirds of the forearm and is quite closely applied to the perichondrium and cpndensed mesenchyma of the skeletal structures beneath. A portion of it overlaps the second and third groups. It is the still undifferentiated extensor communis digitorum, extensor carpi ulnaris, and extensor minimi digiti. It is supplied by branches of the posterior interosseus nerve. | |||
The second group occupies the proximal portion of the radial side of the forearm. It arises in connection with the first group from the external condyle and adjoining portion of the humerus. The muscle mass passes distally along the radius and soon divides into two parts between which the radial nerve passes. The radial part fuses with the condensed tissue of the distal end of the radius. It is the brachioradialis muscle. The second part passes beneath the third group and fuses with the condensed mesenchyma at the proximal ends of the second and third digits. It is the extensor carpi radialis longior et hrevior muscle. Branches of the musculospiral nerve supply this second group. | |||
The third group arises beneath the first from the ulna and radius. Its fibers pass toward the radial side of the forearm, passing from beneath the first group and over the second group, and finally end in the condensed tissue of the first and second digits. The portion to the second digit is closely fused with the portion of the first group which goes to this digit. This group is quite closely applied to the underlying skeletal condensed tissue. The third group represents the abductor pollicis longus, extensor pollicis brevis, extensor pollicis longus and extensor indicis proprius. Branches of the musculospiral nerve supply this group. | |||
The supinator I believe must arise in connection with the third group, judging from its position and the direction of its fibers. | |||
The muscle fibers of the extensor groups do not extend as far distally as do those of the flexor mass. | |||
===The Nerves=== | |||
The enormous size of the lower cervical nerves attracts the attention at once. In the plates and figures they are given in their true proportion to the other structures. The main portion of the brachial plexus has but a very slight posterior inclination. | |||
A branch from the V cervical supplies the rhomboid muscle mass. | |||
The V, VI, VII and VIII cervical and I thoraeic nerves unite to form the brachial plexus. The IV cervical does not connect with the plexus. The main portion of the plexus forms a continuous sheet in which indications of the three cords can be seen. The V and VI unite before joining the others and from this union is given off the -suprascapular. It leaves the trunk at right angles and has the appearance of having its proximal end dragged distally to\\'ard the arm by the main portion of the plexus. The VIII and I thoracic unite before joining the plexus. The continuous sheet formed by these five nerves soon splits into a lateral (dorsal) and median (ventral) division. The lateral corresponds to the posterior cord and from it arise the circumflex, subscapular and musculospiral nerves. These nerves take the normal course found in adult and supply the same muscles as in adult. Cutaneous branches are also given ofl^. The median sheet of the plexus quickly divides into several bundles. The anterior one corresponds to the distal end of the external cord. From it are given off the musculo-cutaneous, two branches to the pectoral mass, and one head of the median nerve. The posterior division corresponds to the distal end of the inner cord. From it arise branches to the pectoral mass, the inner head of the median, the ulnar and internal cutaneous nerves. The distal end of the median splits into a peculiar fan-like arrangement of its branches. Both median and ulnar give branches to the deep flexor mass and anastomose within the mass. | |||
I have attempted to trace the origin of the fibers in the main nerves of the arm. The results are given in the following table: | |||
Cervical. Thoracic. | |||
Suprascapular V, VI ? | |||
Subscapularis V, VI, VII | |||
Long thoracic VII, VIII I | |||
Anterior thoracics V, VI, VII, VIII I | |||
Musculo-cutaneous V, VI, VII ? | |||
Median V, VI, VII, VIII I | |||
Circumflex V, VI, VII | |||
Musculospiral V, VI, VII, VIII I | |||
Ulnar VI?, VII, VIII I | |||
==Larynx== | |||
{{Ref-Lisser1911}} | |||
Carnegie Embryo {{CE109}} 10.5 mm. Transverse sections, 20 micron | |||
(Embryo {{CE109}} measures V. B. 10.5 mm and N. B. 11 mm in length and is about 5 weeks old.) | |||
===The Cartilages=== | |||
The cartilages of the larynx, or better what become the cartilages of the larynx, are at this stage but very imperfectly formed, as is to be expected, since they reveal themselves for the first time in a 10.5 mm. human embryo. Consequently their appearance is introduced by condensations of mesenchyma, very well termed "pre cartilage." No true cartilage formation, whatsoever, has occurred at this time in any of the laryngeal cartilages, nor yet in the hyoid bone and styloid process. I have investigated the sections of 8, 8.5, 9, and 10 mm. embryos for earlier traces of these structures, but have been consistently unsuccessful, so that I place the first recognizable stage of the larynx skeleton (if one may be permitted to use such an expression) at 10.5 mm. in man. Of course, it is well known that all the structures of the larynx have their ultimate foundation in the gill arches, and I refer those interested in this very early stage of the subject to Frazaer's interesting studies ; but I would be skeptical of finding, wdth the present means of study at our disposal, actual structural status, as regards separate cartilage and muscle masses, earlier* than 10.5 mm. in man. | |||
====The cricoid-cartilage==== | |||
{Figs. 1-2.) Contrary to expectations, the cricoid cartilage does not appear to develop from two lateral portions of condensed mesenchyma, separate and independent of each other, which later grow together ventrally and dorsally about the lumen of the larynx; nor as one might be led to anticipate, form a large area posterior or dorsal to the larnyx lumen (this portion ater becoming more prominent by far than the anterior arcus). But at this stage, there is a predominance of condensed mesenchyma about the ventral portion or arcus, which fades of laterally, and then becomes more emphasized again, by greater compactness and deeper stain, dorsal or posterior to the lumen, but not so extensively nor so well marked as ventrally. This is true provided one considers the deeper staining, more strikingly isolated portions of condensed mesenchyma, as the anlage. So that it seems reasonable to assume that the cricoid cartilage originates from an anlage primarily ventral in the position of what later becomes the anterior arcus, and also, though less prominently from a posterior portion, of itself perhaps, originating from two slightly separated posteriorly lateral portions. The lateral portions then, develop by a welding of the anterior and posterior portions lateralward. At this stage then, the ventral portion of the cricoid cartilage is appreciably advanced in development over the rest of this structure. It does seem difficult to reconcile this with the appearance, for instance, in the 20 mm. stage, when the ventral portion still persists as condensed mesenchyma, while the lateral parts have undergone considerable chondrification. | |||
====The arytenoid cartilages==== | |||
It is doubtful whether these structures can be determined at this early period in development; probably not. There are two faint, indefinite masses which suggest beginning condensation, but as there is a possibility of these being a part of the superior portion of the cricoid, no positive assertions can be made as to their independence. | |||
====Thyreoid cartilage==== | |||
This cartilage like the cricoid, makes its initial appearance at this stage. I have looked in vain for its rudiments in earlier human embryos. There has been quite a lengthy discussion, during most of the nineteenth century, in which many have participated, as to whether the thryeoid cartilage develops from two lateral anlages which grow around and fuse ventrally, or whether there is in addition to these lateral anlages, a third one a pars intermedia. Nicolas, in his excellent resume of the subject treats this very fully. My observations inclined to the former view, as in all the stages studied, the lateral portions depict a decidedly more advanced stage of development; yet, I must add that I have found no stage, where the lateral halves alone were present, and in which there was no indication whatsoever of a ventral condensation. Fig. 2 shows the thjTCoid cartilage as it exists in a 10.5 mm. embryo. It will be noticed that there is no interruption in the continuity between the two lateral portions via the ventral part ; but it will also be seen that the lateral parts exhibit a denser condensation. There is no suggestion of an inferior or superior cornu. The thryeoid cartilage merely looks like a horse shoe mass of condensed mesenchyma. | |||
====The hyoid hone and styloid process==== | |||
These bodies are easily made out existing as pure condensed mesenchyma. .The styloid process is more advanced than the greater cornu of the hyoid at this time. | |||
The epiglottis. This cartilage can be discerned, much flattened in comparison with the adult type. It shows as a beginning area of condensation. | |||
===The Musculature=== | |||
The eight muscles are first visible at this period of development, not all as individual muscle masses however, although it is true that one or two of the muscles can be clearly differentiated from the rest. It is curious to note that the laryngeal musculature shows more advanced differentiation than the pharyngeal constrictors, as shown by clearer outline and more extensive fibrillation. The intrinsic oesophageal musculature however is farther developed. The sphincter formation, to which attention has been called by so many writers, can be recognized at this stage, and in a few sections there is some tendency to continuity between the fibres of the outer pharyngeal ring and the inner laryngeal one. Probably too much stress has been laid on this structure, perhaps by reason of the fact that an analogous muscle has been found in lower animals. At any rate, the muscles of the larynx differentiate themselves much earlier than has been previously believed. For instance, the crico artyaenoideus posterior (fig. 1) is unmistakably isolated at this stage, well defined of good size, and abundant fibrillation ; nor does it include any other laryngeal muscle. There is also a muscle mass with some fibrillation, not so large, but plainly evident, which apparently includes the crico aryteanoideus lateralis and thyreo arytaenoideus (fig. 2), principally the former. It is placed on the lateral surface of the cricoid cartilage. The inter-arytaenoideus; if it exists at all, does not appear as one muscle. In the position where one would expect to find it, there is continuity of the larynx and pharynx lumina. But just at the point where these join there are muscle fibres on either side, but which do not unite. These may later bridge across to form the m, interarytaenoideus, but of that point, I am by no means certain. No trace was found of the m. aryepiglotticus or the m. thyreoepiglotticus. The m. cricothyroideus » is fairly well developed, though not nearly so far advanced in form and size as the m. cricoary taenoideus posterior. It is the only one that shows any tendency to relation with the pharyngeal musculature, and Frazaer considers them to have the same origin. In general the musculature of the larynx is rather better defined than the cartilages at this | |||
period. Strazza in 1888, completed the only really valuable work done on the development of the human laryngeal musculature. Nothing of importance has been added since his paper. He thinks that the laryngeal, tongue and pharyngeal musculature develop out of one and the same muscle mass, which in the early embryo develops from an isolated 'muscle island,' which exists independently of the muscle plates. And that the premuscle tissue of the tongue and larynx is a continuous one, the latter merely lying inside the former. In the region of the epiglottis and larynx, he thinks, is also contained the premuscles masses, though he cannot differentiate them at all in his youngest embryo (12 to 13 mm). He associates the simultaneous development of the tongue and larynx musculature from the same source, with the fact of the union in speech between the muscles of the tongue and larynx. This is an attractive theory, but my observations cannot substantiate his statement. There is no indication that the larynx muscles develop from the myotomes, on the contrary, they apparently arise from the ventral visceral mesenchyme which continues up into the floor of the mouth. Bvt in this 10.5 mm. embryo in which even certain larynx muscles can he isolated, there is no association with the tongue musculature, and hut little with the pharyngeal set. In earlier stages the cells which are to form the premuscle masses cannot be distinguished by our present methods from other cells of the condensed mesenchyme of this region. | |||
===The Nerves=== | |||
The n. laryngeus superior can be traced to the vicinity of the greater cornu of the hyoid, and the wing of the thyreoid, but I could not follow either the motor branch to its inervation of the crico thyreoideus muscle, or the sensory portion, within the larynx. Bits of tissue, were seen that might be nerve tissue, but I cannot be certain of this branch of the vagus any further than to its proximity to the hyoid and thyreoid. The nerve Recurrens, later the nerve laryngeus inferior (fig. 1) is better developed and can be | |||
followed clearly to its innervation of the crico artyaenoideus posterior ; but I cannot trace it to the other muscles, nor to any anastomosis with the n. laryngeus superior. | |||
==Normal Embryos Over 9 mm Long== | |||
[[Book_-_Contributions_to_Embryology_Carnegie_Institution_No.1-3#Normal_Embryos_Over_9_mm_Long|Normal Embryos Over 9 mm Long]] | |||
{{Ref-Mall1915}} | |||
A more advanced stage is seen in a similar specimen (No. {{CE109}}) which contains an embryo 10.5 mm. long. This also shows a considerable amount of blood between the villi, but most of it seems to be fresh. A portion of the clot is stratified, but the rest does not show any marked signs of coagulation. Part of the stratified clot has within it a group of leucocytes. The chorion appears to be normal, and where it comes in contact with the tube wall its trophoblast is extremely active. It forms a wall which prevents further hemorrhage from the vessels of the tube into the intervillous spaces. Various stages of this process are shown in the figures of this specimen. In plate 1, figure 2, the trophoblast within the sinuses is shown. Most of the endothelial lining of the veins has been destroyed, but part still remains, as shown in the figure. An earlier stage of this process of destruction of the wall of the blood vessels is shown in plate 1, figure 1. Here are found all stages of trophoblastic development, from the tips of the villi to the free cells within the blood vessels. A somewhat more advanced stage is shown in plate 1, figure 3. The wall of the venous sinus has been completely destroyed and in its place are strands of trophoblast. Between the lumen and the villi is a pronounced vacuolated syncytium partly filled with blood. This specimen illustrates beautifully the dam thrown up by the trophoblast to prevent hemorrhage from the open veins. The blood between the villi is well encircled with trophoblastic cells; at some points the leucocytes have invaded it. A large clot, which is about 15 mm. long and 2 to 5 mm. thick, separates in part the chorion and the villi from the tube wall. On the chorionic side it is covered with an incomplete layer of trophoblastic cells, as shown in plate 4, figure 3. These seem to be invading the blood clot. Individual cells are often found far from their main strand, as shown in this figure. Undoubtedly we have here a clot of blood which the trophoblast is attempting to destroy. There is no indication of fresh hemorrhages in any of the sections. The trophoblast is extensive and vacuolated, forming a wall which protects nearly all of the intervillous spaces. | |||
{{Carnegie numbered embryo links}} | {{Carnegie numbered embryo links}} | ||
{{Footer}} | |||
[[Category:Carnegie Stage 18]] | [[Category:Carnegie Stage 18]] | ||
[[Category:Week | [[Category:Week 7]][[Category:1800's]][[Category:Ectopic Pregnancy]] | ||
[[Category:Transverse]] | |||
Latest revision as of 09:46, 20 October 2020

This Embryology category shows pages and images that relate to the Carnegie Collection Embryo No. 109. This embryo was classified as Carnegie stage 18 occurring during Week 7.
- Links: Carnegie stage 18 | Keibel Mall Fig 488 | Keibel Mall Fig 489 | Keibel Mall Fig 491 | Carnegie Collection
| Serial No. | Size (mm) | Grade | Fixative | Embedding Medium | Plane | Thinness (µm) | Stain | Semi. ducts | P.-M. duct (mm) | Year | Notes |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 109 | E., 12.0* Ch.,30 | Poor | Alc. | P | Transverse | 20 | Al. coch. | 1 | 0.4 | 1897 | Tubal. Least—advanced third |
| Carnegie Collection Embryos - Stage 18 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Serial No. | Size (mm) | Grade | Fixative | Embedding Medium | Plane | Thinness (µm) | Stain | Semi. ducts | P.-M. duct (mm) | Year | Notes |
| 109 | E., 12.0* Ch.,30 | Poor | Alc. | P | Transverse | 20 | Al. coch. | 1 | 0.4 | 1897 | Tubal Least—advanced third |
| 144 | E., 16.0* Ch, 40x30x30 | Good | Formalin | P | Sagittal | 40 | Al. eoch. | 3 | 0.85 | 1899 | Most—advanced third |
| 175 | E., 13.0 Ch, 30x25x25 | Poor | Alc. | P | Transverse | 20 | Al. coch. | 2 | 0.6 | 1900 | Tubal Partly macerated |
| 296 | E., 17.0 | Poor | Ale. | P | Coronal | 20 | Various | 3 | 0.85 | 1905 | Most—advanced third |
| 317 | E., 16.0 | Good | Formalin | P | Coronal | 20 | (Stain - Haematoxylin Eosin) or. G. | 2 | 0.7 | 1905 | Middle third |
| 351 | E.,14.0* | Good | Formalin | P | Coronal | 250 | Slightly carmine— | 2 | 038 | 1904 | Injected (Berlin blue) |
| 406 | E., 16.0 Ch., 40x40x40 | Good | Formalin | P | Sagittal | 20 | (Stain - Haematoxylin Eosin) | 3 | 0.7 | 1907 | Operative. Most—advanced third |
| 423 | E., 15.2 | Good | Formol—Zenker | P | Transverse | 50 | Carmine | 3 | 0.85 | 1904 | |
| 424 | E., 172 | Good | Formalin | P | Transverse | 50 | Carmine | 3 | 10 | 1904 | Double infection. Advanced |
| 492 | E, 16.8 Ch, 40 x 40 | Exc. | Zenker | P | Coronal | 40 | Al. coch. v | 3 | 0.7 | 1911 | Injected (India ink) |
| 511 | E., 160* Ch., 3?* 32x32 | Good | Ale. | P | Sagittal | 40 | Al. coch. | 3 | 1.1 | 1911 | Head injured. Most advanced in group |
| 670 | E, 12.5 | Poor | Ale. | P | Sagittal | 50 | (Stain - Haematoxylin Eosin) | 3 | 10 | 1913 | Tubal Advanced |
| 719 | E, 15.0 Ch, 50x50x50 | Good | Formalin | P | Trans | 40 | Al. coch. | 2 | 0.6 | 1913 | Median in group |
| 733 | E., Ch., 4Sx40x2S | ISO Poor | Formalin | P | Sagittal | 50 | Al. coch. | 2 | 0.6 | 1913 | Median in group |
| 841 | E. 15.0 Ch., 18 x 16x9 | Good | Formalin | P | Coronal & Trans, 20 | 10 | (Stain - Haematoxylin Eosin), carmine | 2 | 0.32 | 1914 | Operative. Head cut separately |
| 899 | E, 160* Ch. 50 x 18 x IS | Good | Bouin | P | Sagittal | S0 | Al. coch. | 3 | 0,65 | 1914 | Tubal Head injured |
| 991 | E. l?.0 | Good | Formalin | P | Sag | so | R, V, Gieson | 3 | 0.9 | 1914 | Advanced |
| 1909 | E., 14.6 | Good | Formalin | P | Coronal | 20 | Al. coch,or. G. | 1 | 0.3 | 1917 | Less advanced |
| 2673 | E.,15.5 | Good | Formalin | P | Transverse | 40 | Al. coch. | 2 | 0.52 | 1919 | Median in group |
| 4430 | E., 14.0 Ch, 51 x40x21 | Exc. | Corros. acetic | P | Transverse | 15 | Al. coch,or. G. | 3 | 0.9 | 1923 | Most—advanced third |
| 5542B | E., 16.0 Ch, 37x32x25 | Good | Formalin | P | Transverse | 40 | Al. coch. | 2 | 0.7 | 1927 | Other twin abnormal |
| 5747 | E, 15.2 Ch, 32x27x25 | Poor | Alc.—formol | P | Sagittal | 25 | Al. coch. | 2 | 0.25 | 1928 | Least—advanced or middle third |
| 5935A | E, 13.5 Ch, 40x30x30 | Good | Formalin | P | Coronal | 40 | Al. coch. | 1 | 0.38 | 1929 | Other twin stunted |
| 6522 | E, 13.2* | Good | Corros. acetic | C—P | Coronal | 10 | Al. coch. | 3 | 0.8 | 7 | Middle or most—advanced third |
| 6524 | E, 11.7* | Exc. | Corros. acetic | C—P | Transverse | 10 | Al. coch. | 1 | 0.4 | ? | Least—advanced third |
| 6525 | E, 13.8* | Exc. | Corros. acetic | C—P | Sagittal | 8 | Al. coch. | 2 | 0.42 | ? | Weak staining |
| 6527 | E, 14.4* | Exc. | Corros. acetic | C—P | Transverse | 15 | Al. coch. | 2 | 0.67 | ? | Mechanical damage |
| 6528 | E, 13.4* | Exc. | Corros. acetic | C—P | Coronal | 8 | Al. coch. | 1 | 0.33 | ? | Least—advanced third |
| 6529 | E, 15.6* | Good | Corros. acetic | C—P | Coronal | 10 | Al. coch. | 2 | 0.4 .5 | Middle third | |
| 6533 | E, 12.5* | Good | Corros. acetic | C—P | Sagittal | 6, 8, 10 | Al. coch. | 2 | 0.45 | ? | Middle third |
| 6551 | E, 18.0 | Poor | Formalin | p | Coronal | 40 | (Stain - Haematoxylin Eosin) | 3 | 0.8 | 1932 | Tubal |
| 7707 | E, 14.5 Ch ,37x32 | Exc. | Bouin | C—P | Transverse | 10 | (Stain - Haematoxylin Eosin), phlox. | 2 | 0.54 | 1939 | Operative. Middle third |
| 8097 | E, 15.5 Ch, 37x25x21 | Good | Formalin | C—P | Transverse | 10 | (Stain - Haematoxylin Eosin) | 1 | 0.19 | 1942 | Least advanced in group |
| 8172 | E, 16.5 | Exc. | Bouin | C—P | Transverse | 20 | (Stain - Haematoxylin Eosin) | 3 | 0.58 | 1943 | Operative. Very advanced |
| 8235 | E, 14.0* | Good | Bouin | C—P Sagittal | 10 | (Stain - Haematoxylin Eosin) | Mallory | 2 | 0.25 | 1944 | Tubal |
| 8355 | E, 15.0 Ch, 23 | Exc. | Formalin | C—P | Coronal | 10 | Azan | 1946 | Tubal. Duplicated spinal cord caudally | ||
| 8812 | E, 12_9 | Exc | Formalin | C—P | Transverse | 10 | (Stain - Haematoxylin Eosin) | 1950 | Rubella. Medical abortion. Midbrain punctured | ||
| 8945 | E, 13.9 | Good | Zenker | p | Transverse | 8 | Borax, carm. | 1952 | Univ. Chicago No. H 1254 | ||
| 9107 | E, 17.0 Ch, 38x28x22 | Good | Bouin | p | Transverse | 15 | Borax, carm. | 1918 | Univ. Chicago No. H 516 | ||
| 9247 | E, 15.0 | Exc. | Bouin | C—P | Sagittal | 8 | Azan | 1954 | Tubal | ||
Abbreviations
| |||||||||||
References
Bardeen CR. and Lewis WH. The development of the limbs, body-wall and back. (1901) Amer. J Anat. 1: 1-36.
Lewis WH. The development of the arm in man. (1902) Amer. J Anat. 1(2): 145-184.
Bardeen CR. Development of the thoracic vertebrae in man. (1905) Amer. J Anat. 4: 163-174.
Sabin FR. The lymphatic system in human embryos, with a consideration of the morphology of the system as a whole. (1909) Amer. J Anat. 9(1): 43–91.
Streeter GL. On the development of the membranous labyrinth and the acoustic and facial nerves in the human embryo. (1906) Amer. J Anat. 6:139-165.
Whitehead RH. and Waddell JA. The early development of the mammalian sternum (1911) Amer. J Anat. 12: 89-106.
Lisser H. Studies on the development of the human larynx. (1911) Amer. J Anat. 12: 27-66.
Mall FP. On the fate of the human embryo in tubal pregnancy. (1915) Contrib. Embryol., Carnegie Inst. Wash. Publ. 221, 1: 1-104.
Limb
Lewis WH. The development of the arm in man. (1902) Amer. J Anat. 1(2): 145-184.
Embryo CIX measures V. B. 10.5 mm. and X. B. 11 mm. in lengtli and is about five weeks old. There is a marked advance over the preceding stage. Cartilage has made its appearance both in the vertebrae and in portions of the arm skeleton. There is considerable difference in the character of the cartilage of the vertebra from that in the arm. The latter seems more advanced and lias more the appearance of true hyaline cartilage. It is possible that the cartilage appears first in the arm, though I have not been able to examine intervening stages to determine this with certainty. Other portions of the arm skeleton are in the precartilage and condensed tissue stages. Both cartilage and precartilage are surrounded in most places by a distinct perichondrium. This takes a very deep stain with the alum carmine. This perichondrium shades off into the condensed tissue of the carpus, which is like that composing the skeletal core in the preceding stage. This again shades into the even less differentiated tissue of the digits, which is at a.bont the same stage of development as the hand plate of the preceding stage, and it in turn shades off into the surrounding mesenchyma.
The muscles in the arm region show very different degrees of development. Those derived from the muscle plate system are in advance of most of the others. The trapezius, levator scapulae and serratus magnus are about as far advanced as those from the muscle plate system, they show distinct muscle fibers and are for the most part quite sharply limited from the surrounding loose mesenchyma. In position they correspond with their premuscle masses of the preceding stage. The pectoral muscle is next in advance and the latissimus dorsi next. These two muscles grow from the humeral region towards their future attachments on the body wall. It is this portion which lies farthest from the humerus which seems to show the most advance in fibrillation and the sharpest limitation from the surrounding mesenchyma. At the humeral end these muscles gradually shade into a condensed mesenchyma, which fuses with neighboring muscle and skeletal elements. Both muscles correspond in position to their premuscle masses of the preceding stage. As in the preceding stage, embryo CLXIII, the trapezius and serratus premuscle masses were in advance of the pectoral and latissimus; in embryo CIX we find the same relation still continues.
The remaining muscles of the arm apparently develop in situ from the premuscle sheath and undergo practically no migrations. They do not appear to be as far advanced as any of the above mentioned muscles. Of these muscles developing from, the arm premuscle sheath, the more proximal ones are more developed than the ones more distal. In the scapulo-humeral region most of the muscles show partial fibrillation, while those in the palm of the hand are in about the same condition as the proximal portion of the premuscle sheath in the preceding stage.
The fibrillation, position and nerve supply have made it possible to determine the presence of most of the muscles of the arm.
The Skeletal, System
The Vertebral Column
The intervertebral discs are composed of condensed mesenchyma, the cells having a concentric arrangement about the chorda. The vertebral bodies between the discs are each composed of two masses of cartilage, one on either side of the chorda. Tliey are surrounded by a perichondrium. Along the ventral surface of the vertebral coluuni is a layer of dense mesenchyma, which probably represents both perichondrium and the anterior common ligament. The neural processes, composed of condensed mesenchyma, are clearly defined. They are continuous with the discs and form a wide, shallow groove for the spinal cord. The transverse processes arise by two roots, one from the base of the neural process and the other from the disc. They are of condensed mesenchyma.
The Ribs. - The ribs are more sharply defined than in CLXIII. They are of condensed tissue except for a small area near the head, which is of precartilage. They extend farther into the body Avail than in the preceding stage.
Fig. 9. Skeleton of the arm region of embryo CIX, lateral view. X 12 diameters.
The Arm Skeleton
The Scapula is composed of precartilage and has greatly altered in shape. It lies in the region of the lower four cervical and first one or two thoracic vertebrae. From the anterior border, which corresponds to the spine, springs the large curved acromion process. On the median surface at the junction of the humerus with the scapula arises the large hooked coracoid process. Eunning across the median surface of the scapula to the vertebral border is a slight ridge which separates the supraspinatus from the subscapularis muscles and corresponds to the future anterior border. The condensed tissue is thickened on the medial surface into a perichondrium, while on the lateral surface the precartilage shades off into the surrounding mesenchyma.
The Clavicle
A rather poorly defined mass of condensed tissue continues from the tip of the acromion toward the tip of the first rib, extending for about one-third this distance. This mass represents the clavicle. From it a mass of ill-def]ned tissue extends to the coracoid process and represents the coraco-clavicular ligament.
The Humerus is directly continuous with the scapula and root of the coracoid process. No signs of joint surfaces or cavity are present. Both ends of the shaft are enlarged and Ihe distal end shows both external and internal condyles. The core of the shaft is of hyaline cartilage; this is surrounded by very thick perichondrium, which shades ofE into the condensed tissue of each end in which is enclosed an area of precartilage. The distal end seems more advanced than the proximal.
The Radius and Ulna are continuous with the distal end of the humerus, no indications of joint surfaces or cavities being present.
Fig. 10. Outline of the arm region of embryo CIX, lateral view from Plate IV. Bardeen and Lewis, Vol. I, No. 1, this Journal. X 12 Diameters.
There is more flexion at the elbow than in CLXIII. The forearm occupies a position about half way between pronation and supination. The core of each shaft is composed of hyaline cartilage. This is surrounded by a very thick perichondrium, which continues into the condensed tissue at either end of the bone, in which precartilage is enclosed. The hand-plate is continuous with the distal ends of the radius and ulna. It is composed of condensed mesenchyma. There are several centers of increased condensation which I believe must correspond to the carpal bones, namely, the scaphoid, lunar, pyramidal, trapezium, trapezoid, OS magnum and unciform. The scaphoid is in line with the radius and the lunar with the ulna, while the pyramidal is at the ulnar side of the carpus, and as the metacarpal V continues from it more than the uniform the whole hand has a peculiar bend toward the ulnar side. From the carpus five masses of condensed tissue project. They shade off into the surrounding mcsenchyma which fills the distal end of the arm. The condition of these finger masses corresponds to the condition of the hand-plate in CLXIII. There is not the sliglitest indication of segmentation into metacarpals and phalanges. The radial of the five projections probably consists of both trapezium and metacarpal I, which have not yet shown signs of separate centers of condensation.
The Muscular System
The muscle plate system has become differentiated into several muscles, namely, the deep dorsal muscles, the intercostals, the abdominal muscles and the deep ventral neck muscles.
The infrahyoid muscles correspond in position and nerve supply with the infrahyoid premuscle mass of the preceding stage. They extend nearly to the region where the median end of the clavicle will eventually extend.
The trapezius muscle has extended posteriorly to the level of the fifth cervical vertebra. Its posterior end lies near to the lateral surface of the body and is connected to the tips of the neural processes as far posteriorly as the second thoracic vertebra by a considerable interval of fascia. As the muscle passes anteriorly it lies deeper and deeper from the surface, being separated from it by the platysma and facial muscles. Its ventral border is free from attachment to the scapula and clavicle. At the level of the second cervical vertebra it is joined by the stemomastoid muscle, which has ascended from the more ventral neck region. The nerve supply is as in the adult.
The rhomboid mass lies in the region of the Y and A^I cervical vertebrae. It connects with the fascia passing to the dorsal tips of the neural processes but has no scapular attachment. A branch of the fifth cervical nerve supplies it.
The levator scapula; and serratus anterior muscles form a continuous fibrillated mass, extending from the first cervical vertebra to the ninth rib. It occupies much the same position that its premuscle mass did in embryo CLXIII except that the posterior end now extends to the ninth rib. Digitations go to all the cervical transverse processes and to each of the anterior nine ribs. The anterior and posterior digitations are very slender and contain but few fibers. The thickest part of the- muscle lies in the scapular region. There is no scapular attachment. The ventral edge of the muscle lies at about the same level as the dorsal edge of the scapula but in a more median plane. Branches from the second to the seventh cervical nerves supply the muscle. The first three penetrate directly into the muscle. The last three form a trunk which runs along the lateral surface of the muscle as far as the fourth rib.
The pedoralis major and minor "^ are united into a common muscle mass, which is well differentiated from the surrounding tissue. It forms a thick oval mass, which extends from the level of the second rib to the proximal portion of the humerus. The greater part of the muscle thus lies anterior to the first rib. As the mass bends towards the humerus it is attached also to the clavicle. So probably both sterno-costal and clavicular portions are present. The median side of the mass bulges towards the coracoid process and represents the minor. Most of the mass shows distinct fibrillation, but toward the humerus this passes into the condensed tissue which is not sharply outlined from the surrounding structures. The position of the pectoral muscle corresponds to the position of the pectoral premuscle mass in embryo CLXIII. Branches from the median side of the brachial plexus supply the pectoral. Two from the external cord contain fibers from the fifth, sixth and seventh cervical nerves. Two come from the inner cord. Within the muscle complicated anastomoses occur from which fibers spread out in all directions.
The muscles thus far considered were fairly definite, and, as we have seen, come from quite definite premuscle masses. The remaining muscles of the arm are in process of differentiation from the arm premuscle sheath. The exact limits of the individual muscles are almost impossible to determine.
The deltoid muscle extends from the acromion and clavicle and fascia over the infraspinatus to the humerus. It is very closely connected with the infraspinatus and only by the difference in the nerve supply can the two be separated. The position of the teres minor is also only indicated by its nerve and not by any line of separation between it and the infraspinatus or deltoid. The origin of part of the deltoid from the acromion and clavicle helps to distinguish some of its fibers, but a short distance from this origin no line of separation can be made between it and the infra- and supraspinatus muscles. Condensed tissue connects it with the triceps and pectoral muscles. The circumflex nerve supplies this muscle and also sends a branch to fibers which are closely associated with the infraspinatus and probably constitute the teres minor muscle.
That portion of the infraspinatus which lies on the lateral surface of the scapula is fairly distinct except where the deltoid and teres minor muscles join it. The portion of tlie supraspinatus on the anterior onefourth of the median surface of the scapula is distinct, but after it passes the acromion it is inseparably' connected with the infraspinatus and deltoid and pectoral muscles. These muscles shade off into the 23roximal end of the humerus. The main portion of each of these muscles contains muscle fibers. The suprascapular nerve supplies the supra- and infraspinatus muscles.
23 Lewis, Observations on the pectoralis major muscle in man, Johns Hopkins TTosp. Bui., Vol. XII, 1901.
The subscapularis muscle arises from the posterior one-half of the median surface of the scapula and passes beneath the coracoid process to the humerus. The circumflex nerve separates a portion of it from the teres major muscle, but the scapular portions of the two are closely imited, as is also the long head of the triceps. A branch from the circumflex and another from the posterior cord of the brachial plexus supply the subscapularis.
The teres major and latissimus dorsi muscles are closely associated at "their humeral end. The latissimus dorsi' lies in the lateral thoracic region, extending posteriorly as far as the fourth rib. It has no attachments to the ribs or vertebral column. The two muscles are inserted together into the proximal portion of the humerus. The teres major arises from the axillary border of the scapula near its posterior angle. The common portion of the latissimus and teres passes close to the posterior cord of the brachial plexus, from which a large branch is given off that runs into the latissimus and has a brush-like ending near the posterior limit of the muscle. A smaller branch of the posterior cord is given off to the teres major.
The triceps muscle extends along the posterior and lateral surfaces of the humerus, extending from the scapula to the ulna. Indications of the three heads are present. The portion of the muscle lying near the insertion of the latissimus dorsi and the infraspinatus muscles is not sharply defined from them. The musculo-spiral nerve passes through the muscle and gives branches to it.
The biceps and coracohracJnaUs muscles lie along the median side of the humerus, extending from the coracoid process to the radius. The two heads of the biceps are quite closely united nearly to their origins, Avhich are but a short distance apart. The portion of the coracoid process from which the long head arises must ultimately become a portion of the head of scapula. The attachment of the coracobrachialis to the humerus is by condensed tissue, as is the distal end of the biceps to the radius. The distal end of the biceps blends with the brachialis and the flexor mass. The musculo-cutaneous pierces this group and gives off branches to it.
The brachialis muscle is closely attached to the distal one-half of the hmnerns over the anterior and median surfaces. It is also closely attached to the overlying biceps muscle and it is impossible to determine just the line between the two or between it and the brachioradialis muscle. It is also impossible to determine the exact line between the muscle and the underlying perichondrium. It is closely associated with the triceps on one side and the deltoid on the other. The main portion of the muscle is fibrillated and is inserted into the ulna by condensed tissue, which is closely associated with the, flexor mass of the forearm. The musculo-cutaneous nerve gives off a large branch which has a brush-like endins; within the muscle.
Fig. 11. Outline of the arm region of embryo CIX, median view, from Plate V. Bardeen and Lewis, Vol. I, No. 1, this Journal. X 15 diameters.
The flexor muscle mass of the forearm forms a thick layer over the median surface of the ulna, radius, carpus and proximal end of the metacarpus. It is with considerable difficulty that I have separated this mass into two layers. The superficial layer is smaller in extent and lies in the proximal region of the forearm. It is connected with the radial portion of the forearm by a condensed tissue mass and distally fuses with the deep layer to become continuous with the condensed tissue of the digits. The median nerve passes through the proximal portion and then comes to lie between the two layers. From its position and relation to the median nerve I believe this to be the layer from which the flexor carpi radialis, flexor sublimis digitorum, pronator teres and palniaris long-us muscles differentiate. Branches from the median nerve supply this layer. Both layers arise partly from the inner condyle of the humerus, and are continuous more or less with the muscles of the upper arm. The deep layer is closely attached to the perichondrium of the forearm and hand. It is wider in extent than the superficial and shows indications of separations into muscles. Tlie portion for the flexor carpi nlnaris shows most advance. The extension into the hand probably constitutes the portion from which the interossei and lumbrical muscles and flexor tendons develop. It is continuous with the condensed tissue of the digits. The portion on the forearm forms the flexor profundus digitorum, flexor pollicis longus, flexor carpi ulnaris and pronator quadratus muscles. Both the ulnar and median nerves supply the deep layer.
The extensor mass of the forearm is farther advanced than the flexor. It can be differentiated into three groups of muscles which accord well with the adult groups. The first group, the largest and most superficial, extends from the lateral condyle to the proximal ends of the digits, where it blends with the condensed mesenchyma. It is a thin layer and spreads out over the ulnar two-thirds of the forearm and is quite closely applied to the perichondrium and cpndensed mesenchyma of the skeletal structures beneath. A portion of it overlaps the second and third groups. It is the still undifferentiated extensor communis digitorum, extensor carpi ulnaris, and extensor minimi digiti. It is supplied by branches of the posterior interosseus nerve.
The second group occupies the proximal portion of the radial side of the forearm. It arises in connection with the first group from the external condyle and adjoining portion of the humerus. The muscle mass passes distally along the radius and soon divides into two parts between which the radial nerve passes. The radial part fuses with the condensed tissue of the distal end of the radius. It is the brachioradialis muscle. The second part passes beneath the third group and fuses with the condensed mesenchyma at the proximal ends of the second and third digits. It is the extensor carpi radialis longior et hrevior muscle. Branches of the musculospiral nerve supply this second group.
The third group arises beneath the first from the ulna and radius. Its fibers pass toward the radial side of the forearm, passing from beneath the first group and over the second group, and finally end in the condensed tissue of the first and second digits. The portion to the second digit is closely fused with the portion of the first group which goes to this digit. This group is quite closely applied to the underlying skeletal condensed tissue. The third group represents the abductor pollicis longus, extensor pollicis brevis, extensor pollicis longus and extensor indicis proprius. Branches of the musculospiral nerve supply this group.
The supinator I believe must arise in connection with the third group, judging from its position and the direction of its fibers.
The muscle fibers of the extensor groups do not extend as far distally as do those of the flexor mass.
The Nerves
The enormous size of the lower cervical nerves attracts the attention at once. In the plates and figures they are given in their true proportion to the other structures. The main portion of the brachial plexus has but a very slight posterior inclination.
A branch from the V cervical supplies the rhomboid muscle mass.
The V, VI, VII and VIII cervical and I thoraeic nerves unite to form the brachial plexus. The IV cervical does not connect with the plexus. The main portion of the plexus forms a continuous sheet in which indications of the three cords can be seen. The V and VI unite before joining the others and from this union is given off the -suprascapular. It leaves the trunk at right angles and has the appearance of having its proximal end dragged distally to\\'ard the arm by the main portion of the plexus. The VIII and I thoracic unite before joining the plexus. The continuous sheet formed by these five nerves soon splits into a lateral (dorsal) and median (ventral) division. The lateral corresponds to the posterior cord and from it arise the circumflex, subscapular and musculospiral nerves. These nerves take the normal course found in adult and supply the same muscles as in adult. Cutaneous branches are also given ofl^. The median sheet of the plexus quickly divides into several bundles. The anterior one corresponds to the distal end of the external cord. From it are given off the musculo-cutaneous, two branches to the pectoral mass, and one head of the median nerve. The posterior division corresponds to the distal end of the inner cord. From it arise branches to the pectoral mass, the inner head of the median, the ulnar and internal cutaneous nerves. The distal end of the median splits into a peculiar fan-like arrangement of its branches. Both median and ulnar give branches to the deep flexor mass and anastomose within the mass.
I have attempted to trace the origin of the fibers in the main nerves of the arm. The results are given in the following table:
Cervical. Thoracic.
Suprascapular V, VI ?
Subscapularis V, VI, VII
Long thoracic VII, VIII I
Anterior thoracics V, VI, VII, VIII I
Musculo-cutaneous V, VI, VII ?
Median V, VI, VII, VIII I
Circumflex V, VI, VII
Musculospiral V, VI, VII, VIII I
Ulnar VI?, VII, VIII I
Larynx
Lisser H. Studies on the development of the human larynx. (1911) Amer. J Anat. 12: 27-66.
Carnegie Embryo 109 10.5 mm. Transverse sections, 20 micron
(Embryo 109 measures V. B. 10.5 mm and N. B. 11 mm in length and is about 5 weeks old.)
The Cartilages
The cartilages of the larynx, or better what become the cartilages of the larynx, are at this stage but very imperfectly formed, as is to be expected, since they reveal themselves for the first time in a 10.5 mm. human embryo. Consequently their appearance is introduced by condensations of mesenchyma, very well termed "pre cartilage." No true cartilage formation, whatsoever, has occurred at this time in any of the laryngeal cartilages, nor yet in the hyoid bone and styloid process. I have investigated the sections of 8, 8.5, 9, and 10 mm. embryos for earlier traces of these structures, but have been consistently unsuccessful, so that I place the first recognizable stage of the larynx skeleton (if one may be permitted to use such an expression) at 10.5 mm. in man. Of course, it is well known that all the structures of the larynx have their ultimate foundation in the gill arches, and I refer those interested in this very early stage of the subject to Frazaer's interesting studies ; but I would be skeptical of finding, wdth the present means of study at our disposal, actual structural status, as regards separate cartilage and muscle masses, earlier* than 10.5 mm. in man.
The cricoid-cartilage
{Figs. 1-2.) Contrary to expectations, the cricoid cartilage does not appear to develop from two lateral portions of condensed mesenchyma, separate and independent of each other, which later grow together ventrally and dorsally about the lumen of the larynx; nor as one might be led to anticipate, form a large area posterior or dorsal to the larnyx lumen (this portion ater becoming more prominent by far than the anterior arcus). But at this stage, there is a predominance of condensed mesenchyma about the ventral portion or arcus, which fades of laterally, and then becomes more emphasized again, by greater compactness and deeper stain, dorsal or posterior to the lumen, but not so extensively nor so well marked as ventrally. This is true provided one considers the deeper staining, more strikingly isolated portions of condensed mesenchyma, as the anlage. So that it seems reasonable to assume that the cricoid cartilage originates from an anlage primarily ventral in the position of what later becomes the anterior arcus, and also, though less prominently from a posterior portion, of itself perhaps, originating from two slightly separated posteriorly lateral portions. The lateral portions then, develop by a welding of the anterior and posterior portions lateralward. At this stage then, the ventral portion of the cricoid cartilage is appreciably advanced in development over the rest of this structure. It does seem difficult to reconcile this with the appearance, for instance, in the 20 mm. stage, when the ventral portion still persists as condensed mesenchyma, while the lateral parts have undergone considerable chondrification.
The arytenoid cartilages
It is doubtful whether these structures can be determined at this early period in development; probably not. There are two faint, indefinite masses which suggest beginning condensation, but as there is a possibility of these being a part of the superior portion of the cricoid, no positive assertions can be made as to their independence.
Thyreoid cartilage
This cartilage like the cricoid, makes its initial appearance at this stage. I have looked in vain for its rudiments in earlier human embryos. There has been quite a lengthy discussion, during most of the nineteenth century, in which many have participated, as to whether the thryeoid cartilage develops from two lateral anlages which grow around and fuse ventrally, or whether there is in addition to these lateral anlages, a third one a pars intermedia. Nicolas, in his excellent resume of the subject treats this very fully. My observations inclined to the former view, as in all the stages studied, the lateral portions depict a decidedly more advanced stage of development; yet, I must add that I have found no stage, where the lateral halves alone were present, and in which there was no indication whatsoever of a ventral condensation. Fig. 2 shows the thjTCoid cartilage as it exists in a 10.5 mm. embryo. It will be noticed that there is no interruption in the continuity between the two lateral portions via the ventral part ; but it will also be seen that the lateral parts exhibit a denser condensation. There is no suggestion of an inferior or superior cornu. The thryeoid cartilage merely looks like a horse shoe mass of condensed mesenchyma.
The hyoid hone and styloid process
These bodies are easily made out existing as pure condensed mesenchyma. .The styloid process is more advanced than the greater cornu of the hyoid at this time.
The epiglottis. This cartilage can be discerned, much flattened in comparison with the adult type. It shows as a beginning area of condensation.
The Musculature
The eight muscles are first visible at this period of development, not all as individual muscle masses however, although it is true that one or two of the muscles can be clearly differentiated from the rest. It is curious to note that the laryngeal musculature shows more advanced differentiation than the pharyngeal constrictors, as shown by clearer outline and more extensive fibrillation. The intrinsic oesophageal musculature however is farther developed. The sphincter formation, to which attention has been called by so many writers, can be recognized at this stage, and in a few sections there is some tendency to continuity between the fibres of the outer pharyngeal ring and the inner laryngeal one. Probably too much stress has been laid on this structure, perhaps by reason of the fact that an analogous muscle has been found in lower animals. At any rate, the muscles of the larynx differentiate themselves much earlier than has been previously believed. For instance, the crico artyaenoideus posterior (fig. 1) is unmistakably isolated at this stage, well defined of good size, and abundant fibrillation ; nor does it include any other laryngeal muscle. There is also a muscle mass with some fibrillation, not so large, but plainly evident, which apparently includes the crico aryteanoideus lateralis and thyreo arytaenoideus (fig. 2), principally the former. It is placed on the lateral surface of the cricoid cartilage. The inter-arytaenoideus; if it exists at all, does not appear as one muscle. In the position where one would expect to find it, there is continuity of the larynx and pharynx lumina. But just at the point where these join there are muscle fibres on either side, but which do not unite. These may later bridge across to form the m, interarytaenoideus, but of that point, I am by no means certain. No trace was found of the m. aryepiglotticus or the m. thyreoepiglotticus. The m. cricothyroideus » is fairly well developed, though not nearly so far advanced in form and size as the m. cricoary taenoideus posterior. It is the only one that shows any tendency to relation with the pharyngeal musculature, and Frazaer considers them to have the same origin. In general the musculature of the larynx is rather better defined than the cartilages at this period. Strazza in 1888, completed the only really valuable work done on the development of the human laryngeal musculature. Nothing of importance has been added since his paper. He thinks that the laryngeal, tongue and pharyngeal musculature develop out of one and the same muscle mass, which in the early embryo develops from an isolated 'muscle island,' which exists independently of the muscle plates. And that the premuscle tissue of the tongue and larynx is a continuous one, the latter merely lying inside the former. In the region of the epiglottis and larynx, he thinks, is also contained the premuscles masses, though he cannot differentiate them at all in his youngest embryo (12 to 13 mm). He associates the simultaneous development of the tongue and larynx musculature from the same source, with the fact of the union in speech between the muscles of the tongue and larynx. This is an attractive theory, but my observations cannot substantiate his statement. There is no indication that the larynx muscles develop from the myotomes, on the contrary, they apparently arise from the ventral visceral mesenchyme which continues up into the floor of the mouth. Bvt in this 10.5 mm. embryo in which even certain larynx muscles can he isolated, there is no association with the tongue musculature, and hut little with the pharyngeal set. In earlier stages the cells which are to form the premuscle masses cannot be distinguished by our present methods from other cells of the condensed mesenchyme of this region.
The Nerves
The n. laryngeus superior can be traced to the vicinity of the greater cornu of the hyoid, and the wing of the thyreoid, but I could not follow either the motor branch to its inervation of the crico thyreoideus muscle, or the sensory portion, within the larynx. Bits of tissue, were seen that might be nerve tissue, but I cannot be certain of this branch of the vagus any further than to its proximity to the hyoid and thyreoid. The nerve Recurrens, later the nerve laryngeus inferior (fig. 1) is better developed and can be followed clearly to its innervation of the crico artyaenoideus posterior ; but I cannot trace it to the other muscles, nor to any anastomosis with the n. laryngeus superior.
Normal Embryos Over 9 mm Long
Mall FP. On the fate of the human embryo in tubal pregnancy. (1915) Contrib. Embryol., Carnegie Inst. Wash. Publ. 221, 1: 1-104.
A more advanced stage is seen in a similar specimen (No. 109) which contains an embryo 10.5 mm. long. This also shows a considerable amount of blood between the villi, but most of it seems to be fresh. A portion of the clot is stratified, but the rest does not show any marked signs of coagulation. Part of the stratified clot has within it a group of leucocytes. The chorion appears to be normal, and where it comes in contact with the tube wall its trophoblast is extremely active. It forms a wall which prevents further hemorrhage from the vessels of the tube into the intervillous spaces. Various stages of this process are shown in the figures of this specimen. In plate 1, figure 2, the trophoblast within the sinuses is shown. Most of the endothelial lining of the veins has been destroyed, but part still remains, as shown in the figure. An earlier stage of this process of destruction of the wall of the blood vessels is shown in plate 1, figure 1. Here are found all stages of trophoblastic development, from the tips of the villi to the free cells within the blood vessels. A somewhat more advanced stage is shown in plate 1, figure 3. The wall of the venous sinus has been completely destroyed and in its place are strands of trophoblast. Between the lumen and the villi is a pronounced vacuolated syncytium partly filled with blood. This specimen illustrates beautifully the dam thrown up by the trophoblast to prevent hemorrhage from the open veins. The blood between the villi is well encircled with trophoblastic cells; at some points the leucocytes have invaded it. A large clot, which is about 15 mm. long and 2 to 5 mm. thick, separates in part the chorion and the villi from the tube wall. On the chorionic side it is covered with an incomplete layer of trophoblastic cells, as shown in plate 4, figure 3. These seem to be invading the blood clot. Individual cells are often found far from their main strand, as shown in this figure. Undoubtedly we have here a clot of blood which the trophoblast is attempting to destroy. There is no indication of fresh hemorrhages in any of the sections. The trophoblast is extensive and vacuolated, forming a wall which protects nearly all of the intervillous spaces.
| Carnegie Collection - Carnegie Embryos Sorted by Stage | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Week: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| Carnegie stage: | 1 2 3 4 | 5 6 | 7 8 9 | 10 11 12 13 | 14 15 | 16 17 | 18 19 | 20 21 22 23 |
Cite this page: Hill, M.A. (2024, June 27) Embryology Carnegie Embryo 109. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Category:Carnegie_Embryo_109
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G
Pages in category 'Carnegie Embryo 109'
The following 7 pages are in this category, out of 7 total.
P
- Paper - Development and variation of the nerves and the musculature of the inferior extremity and of the neighboring regions of the trunk in man
- Paper - On the development of the membranous labyrinth and the acoustic and facial nerves in the human embryo
- Paper - Studies of the development of the human skeleton (1905)
- Paper - The development of the arm in man (1902)
- Paper - The development of the limbs, body-wall and back
- Paper - The development of the limbs, body-wall and back (1901)
- Paper - The early development of the mammalian sternum
Media in category 'Carnegie Embryo 109'
The following 13 files are in this category, out of 13 total.
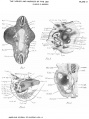 Bardeen1906-plate02.jpg 1,719 × 2,302; 512 KB
Bardeen1906-plate02.jpg 1,719 × 2,302; 512 KB
 Bardeen1906-plate31.jpg 1,571 × 2,330; 257 KB
Bardeen1906-plate31.jpg 1,571 × 2,330; 257 KB
 Keibel Mall 2 488.jpg 1,200 × 576; 317 KB
Keibel Mall 2 488.jpg 1,200 × 576; 317 KB
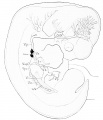 Keibel Mall 2 489.jpg 855 × 1,000; 97 KB
Keibel Mall 2 489.jpg 855 × 1,000; 97 KB
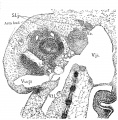 Keibel Mall 2 491.jpg 987 × 1,000; 294 KB
Keibel Mall 2 491.jpg 987 × 1,000; 294 KB
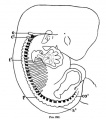 Keibel Mall 232.jpg 450 × 511; 35 KB
Keibel Mall 232.jpg 450 × 511; 35 KB
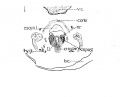 Lisser1911 fig01.jpg 1,119 × 900; 92 KB
Lisser1911 fig01.jpg 1,119 × 900; 92 KB
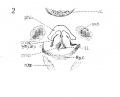 Lisser1911 fig02.jpg 1,119 × 900; 94 KB
Lisser1911 fig02.jpg 1,119 × 900; 94 KB
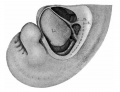 Mall1906-fig21.jpg 1,000 × 763; 72 KB
Mall1906-fig21.jpg 1,000 × 763; 72 KB
 Mall1906-fig25.jpg 1,165 × 887; 153 KB
Mall1906-fig25.jpg 1,165 × 887; 153 KB
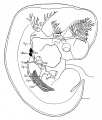 Sabin1909 fig04.jpg 769 × 902; 97 KB
Sabin1909 fig04.jpg 769 × 902; 97 KB
 Streeter1906 fig03.jpg 1,472 × 1,943; 285 KB
Streeter1906 fig03.jpg 1,472 × 1,943; 285 KB
 Streeter1906 plate01.jpg 2,708 × 1,786; 701 KB
Streeter1906 plate01.jpg 2,708 × 1,786; 701 KB












{kind=link}