Paper - A Human Embryo of Twenty-seven Pairs of Somites, Embedded in Decidua
| Embryology - 18 Apr 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Waterston D. A human embryo of twenty-seven pairs of somites, embedded in decidua. (1914) J Anat Physiol., 49(1): 90-118 PMID 17233016

| Historic Disclaimer - information about historic embryology pages |
|---|
 |
A Human Embryo of Twenty-seven Pairs of Somites, Embedded in Decidua
King's College, London.

History
IN the Journal of Obstetrics and Gynecology of the British Empire of February 1910, a communication was published from Dr (now Professor) Watson, of Toronto University, and Mr Henry Wade, on "The Histological Changes associated with an Early Abortion, with special reference to the Vessels of the Decidua."
That communication contained an account of a decidual cast of the uterus which the authors had examined, and which was found to contain, embedded in its wall, a young human embryo.
The observations of these authors were restricted to an examination of the decidua, and chiefly to the pathological changes which it showed. After completing their examination, they were so kind as to hand to me the sections which they had prepared, in order that the structure of the embryo and its annexa might be worked out. I wish to express to these two gentlemen my most cordial thanks for this opportunity of examining a most important and valuable specimen.
The full history of the specimen is given in the publication mentioned above, and it is necessary to repeat here only those facts which relate to the age and condition of the embryo.
They are as follows:-
- "In December 1907 the patient had gone beyond her usual menstrual period by ten days,and then began to bleed slightly. On the supposition that she was pregnant, she was instructed to keep carefully anything that came away, and next day she brought to us a piece of tissue, a complete decidual cast of the uterus, on one wall of which was visible, as a small pea-like elevation, theorem.
- "The uterine decidual cast is a piece of membrane triangular in shape, measuring 3-9 cms. along the base, and 45 cms. alone each wall. The external surface is shaggy in appearance, while on opening the sac the internal surface presents a more or less smooth surface, slightly pitted, and thrown into smooth elevations. Towards the apex of the cast,corresponding to the position of the os internum, is a small pea-like elevation, smooth on the surface and surrounded by a ridge of the mucous membrane rather bigger than the other ridges present. This pea-like elevation represents the ovum covered with decidua, and measures 8 mm. in diameter. The specimen was hardened in fornmalin and embedded in paraffin, and a series of 1800 serial sections were made through the ovum and adjacent parts of the decidua.
- "In the sectioning, we were fortunate in cutting the embryo almost transversely, and the result has been that the sections show the relations of the different parts of the embryo and its membranes with almost diagrammatic clearness."
Examination of the sections showed that a well-preserved human embryo had been obtained at an early stage of development. theinterest and value of the specimen were enhanced by the fact that the whole chorionic vesicle was intact within a complete decidual capsule, and it therefore seemed probable that an examination would prove of value not only as regards the embryo itself, but also in throwing light upon anearly stage of embryonic implantation and placental formation.
In the present paper I propose to give for the most part a descriptive account of the embryo and its organs and membranes, indicating briefly the bearing of some of the facts elicited upon the present state of our knowledge.
Methods
The sections had been stained some with hbenatoxylin and eosin, some by iron alumlhiematoxylin, and some with alcoholic eosin and methyl blue. The histolooy of the embryo and its membranes was, on the whole, good. Mitotic figures were visible in many of the embryonic tissues. Portions of the neural tube showed some shrinkage, but the other tissues, including the heart, blood, and gastro-pulmonary tissues, were in good preservation.
The deciduae showed pathological changes involving mainly the vessels, but the embryo itself appeared to be quite unaffected by organic pathological changes.
In order to work out the external form of the embryo and the general relation of the various parts a wax-plate reconstruction was made, at a magnification of 50 diameters, of the embryo with its yolk-sac, the amnion, and the adjacent zone of the chorion and decidua.
This degree of enlargement did not bring out al the details of the internal structure of the embryo, and subsequent reconstructions were made at a magnification of 200 diameters, both plastic and graphic, from which details of structure were worked out The sections had not been cut with " ritzer" lines for reconstruction, and it was necessary to use the decidua as a guiding structure, together with the photographs and measurements which had been taken of the nodule before it was sectioned.
Comparison of the model with the photographs and the measurements showed that an accurate reconstruction had been obtained.
Thus the measurement-taken directly from the specimen before it was sectioned-of the transverse diameter of the nodule which projected into the uterine cavity was 8mm. In there construction at 50 diameters the corresponding measurement in the model was almost precisely 40 cms, and the degree of correspondence reached in this dimension leads one to believe that the measurements of the embryo, etc., may be relied upon as being exact.
General Arrangement of Parts
The arrangement and something of the structure of the parts of the specimen are seen in fig. 1, which is taken from a section which cuts across the embryo, yolk-sac, body stalk, as well as the chorionic wall and decidua, and in fig. 2, which is taken from a wax-plate reconstruction. The chorionic vesicle is seen lying in the implantation cavity in the decidua. The cavity of the vesicle contains an irregular reticulum, the "rmCatgmiaculee" of Eternod, 1 and towards one end lies the embryo, the amnion, yolk-sac, and the body stalk.

The thin layer of decidua which separates the implantation cavity from the cavity of the uterus is the decidua capsular is; the layer on the opposite side which has separated from the uterine wall is the decidua basal is; while around the equator of the implantation cavity, at the meeting of these two portions, is the decidua marginalise.
The decidua forms a complete capsule for the chorionic vesicle, and it did not show anywhere any signs of the perforation aperture.
The decidua capsularis is thin, measuring only*2mm. in its transverse diameter, while the decidua basalisis much thicker, measuring about 1mm. in average thickness.
The free surface of the former, directed towards the cavity of the uterus, is quite smooth, while the surface of the decidua basalis, where it had separated from the uterine wall, is rough and shaggy.
The decidua showed the histological features characteristic of decimal tissue, which do not need tone described at length.
The layer next to the implantation cavity showed a finely reticulated or laminated appearance, and stained deeply with eosin. It consisted of a fine network of fibres resembling fibrin, lying parallel to the free surface, and in the meshes of the network there were large numbers of red blood cels.
To this surface the ends of some of the vili adhered, though for the greater part the vili terminated freely, and they did not anywhere perforate this laminated layer of fibrin. The fibrin stria was most distinct and thickest in the regions where the vili were most numerous, and the appearances certainly corroborate the view that the "fibrin zone" is produced by the necrotic action of the syncytial layer of the chorionic wall upon the adjacent uterine tissue1 rather than as a result of syncytial degeneration. The intervillous spaces contained red blood cels, in some places in large numbers.
Chorionic Vesicle

The chorionic vesicle (the "ovum" of some writers) is slightly smaller than the "implantation cavity" in which it lies, and closely conforms to it in shape, measuring 8-5 mm. in itslong axis, 7-7 mm. in width, and 4 mm. in depth (see Table I.). These measurements include the vili.
The external surface is partially covered with vili, which are very numerous on the aspect towards the decidua basalis and marginalis, but few and scanty on the aspect towards the decidua capsular is.
The vili were reconstructed on a portion of the wall at the junction of the decidua basalis and marginalis, where they were very numerous; an area of 6A1 square mm. was found to have 30 to40 viliof varying length, each of these measuring as much as 0.9 of a millimetre in length.
The Eternod ovum (Vuill) in a fresh condition measured, with its vili, 100x82x6mm. Its vili had a length of 12 to 2 mm, their diameter being 3 to*8 m., and it was expelled six days from the omitted period.
Structure of the Chorionic Wall and Villi

The chorionic wall and vili showed the structure characteristic of this stage of development. The most external layer consisted of a multi-nucleated layer of protoplasm, distinctly syncytial in character. At the tips of many of the vili this layer was increased in thickness, and formed a sort of knob, composed of a mass of syncytial tissue.
There was, in many places, a deeperlayer of cels, the Langhans layer; but, except in a few places, the distinction between the syncytial and the Langhans layer was not clear. Underneath the cellular layer the chorion wall was composed of a loose reticular mesenchyme tissue with elongated spindle-shaped nuclei. This layer was thicker than the epithelial wall,and its deeper surface was connected with a loose, fibrinous-like reticulum which occupied the interior of the chorionic vesicle and passed from the wall to the surface of the yolk-sac and of the amnion (fig. 1 and fig. 2). This reticulum cannot be said to be organised tissue, and has much greater resemblance to a precipitation deposit. An area by the side of the vitello-intestinal junction in which this reticulum was absent represents a clear portion of the extra-embryonic coelom. The vili consisted of a core of loose mesenchyme tissue, in which blood spaces were present, and a covering cellular layer, similar to that of the chorion, but more distinctly two-layered.
Amnion and Body Stalk
The same figure (fig. 1) shows also the stage of development of the amnion and the body stalk.
The former was a thin membrane, consisting of an inner layer of flattened epithelial cels and an outer mesenchymal layer. It invested only the dorsal aspect of the body stalk, leaving its ventral surface free in the chorionic cavity. In the ventral direction, the amniotic cavity was only very slightly longer than the embryo (see fig. 3).
- 1Nitabuch, R., Beitrdgez ur Kenntnissdermensc lichen Placenta, Disert., Bern, 1887.
In the decidua basalis were many enlarged uterine glands, which, in some cases, opened into the intervillous space. There were also numerous dilated spaces containing blood.
The body stalk came offfrom the right side of the embryo and extended from the lower part of the heart region to the tail bend. It consisted of a loose tissue in which lay the large and tortuous umbilical vessels.
External Form of the Embryo

The external form of the embryo is seen in fig. 3. That illustration shows that it is coiled in a slight spiral, the head lying to the right and the tail to the left. i There .is a well-marked vertex bend as the anterior portion of the bead is bent down towards the heart, but there is no distinct nape bend, and the dorsal aspect shows merely a uniform curvature and passes into a short roundedtailprocess.
The lateral surface of the head shows a prominence overlying the otic vesicle, and in front of that there is another elevated area which marks the position of the developing trigeminal nerve. The anterior and posterior neuropores are closed.
Behind the mouth orifice there are on each side three well-marked elevations corresponding to the three anterior bronchial arches, and behind the first and second of these there is a distinct cleft-like external pocket (fig. 4).
The groove behind the third arch is shallow, and in one or two sections the most dorsal part of this groove shows in its floor two slight elevations, which are the rudiments of the fourth and fifth arches (fig. 4). Behind the third groove is an elevation which is due to the anterior cardinal vein.
The heart forms a distinct prominence on the ventral aspect of the trunk, and the contours of its principal chambers are evident on the surface. The body stalk passes to the right side, and the vitello-intestinal connexion towards the left, and the yolk-sac itself was bent to the left side and lay on the left side of the tail at its lower part.
The limb-buds have not appeared, but on the caudal portion of the trunk the Wolffian ridges were distinct. There were twenty-seven pairsof somites, and the dimensions of the embryo and yolk-sac are shown in the accompanying table.
Table Showing Embryo Dimensions Comparison
Table showing dimensions in millimetres of the specimen described here, and of our comparison witih an earlier and a later specimen.
| Table Embryo Dimensions Comparison | |||
|---|---|---|---|
| Dimensions in millimetres of the specimen described here, and of our comparison witih an earlier and a later specimen. | |||
| Feature | Etertod - Vuill | His - Embryo M | 2W.I. |
| Implantation cavity | 9 x 8 x 4.1 | ||
| Chorionic vesicle | 1 x 8.2 x 6 | 7.5 to 8 | 8.5 x 7.7 x 4 |
| Embryo length (max.) | 1.3 | 2.6 | 2.9 |
| Embryo transverse | 0.9 | ||
| Yolk-sac length | 2.6 | 2.6 | |
| Yolk-sac dorso-ventral | 1.7 | 2.0 | |
| Yolk-sac transverse | 1.6 | ||
In its size and in the stage of development this embryo comes into the known series of human embryos at a stage not well represented.
It is slightly older than the embryo of Robert Meyer, Meyer 300 (N.T. vi.), which was described and figured by Thompson in this Journal,[1] and whose blood-vessels have been worked out and described by Felix.[2]
Another embryo which bears perhaps an even closer resemblance is embryo Wolff of the Berlin Anatom.-biolog. Institute (Professor 0. Hertwig), N.T. fig. vii.5, which was obtained in a decidual cast of the uterine mucosa. The profile outline of my specimen is relatively longer in the long axis and shorter in the dorso-ventral diameter than that specimen, which measures 3.7 mm. in maximum length, and has 23 pairs of somites and 3 pharyngeal pockets.
Beyond the brief description of that embryo in the Normaltafeln, I have not been able to discover any complete account of the internal structure, and none appears to have been published.
Other embryos which resemble the present one are No. IX. of the Normaltafeln, belonging to the collection of Professor Hammar of Upsala, the greatest length of which was 3-4 mm. after fixation, and which had 28 pairs of somites; and another embryo of 3 mm. length described by Broman and figured in the text of the Normaltafeln (text-fig. 10), which has a distinct resemblance to the present specimen in regard to several details.
Age of the Embryo
The history of the decidual cast being passed 10 days after a missed menstrual period refers, I understand, to the anticipated commencement of the period, and therefore presumably some 34 days from the end of the last menstruation.
According to our present knowledge, the age of the embryo would therefore be estimated as from 16 to 19 days; but obviously this is an underestimate, and fertilization must have occurred somewhat earlier in the intermenstrual period, and the age of the embryo is more probably nearer 30 days.
The stage of development can be estimated by reference to the stage of development of the heart rather than to the length of the embryo, and on that criterion the present specimen is slightly older than the embryo Meyer 300.
Internal Structure
Alimentary Canal (fig. 5).-The bucco-pharyngeal membrane had ruptured, but the remains of it were present around the margins of the buccal orifice. That orifice was bounded by the forebrain anteriorly and the mandibular arches on each side and below.
From the anterior portion of the buccal depression Rathke's pocket projected towards the ventral aspect of the forebrain in.the form of a blind recess, triangular on horizontal section, the apex of which passed towards but did not reach the forebrain (fig. 6).
The anterior end of the notochord terminated in the tissues forming the dorsal wall of this pocket (fig. 5).
There was no distinct indication of the presence of any projection of the roof of the pharyngeal cavity behind Rathke's pocket, and it may therefore be stated that Seessel's pocket was not present.
Pharynx and Alimentary Canal
The three anterior bronchial arches were present, and projected into the interior of the cavity of the pharynx, and the arches of the opposite sides met in the floor of the cavity. In that region behind the first arch there was the tubular growth of the thyroglossal duct, which extended downwards and terminated blindly on the ventral aspect of the conus arteriosus. Near its termination a large round mass of cells lay on the ventral aspect of the duct, the cells in the interior being loosely packed together, while the peripheral ones formed a continuous wall. This mass of cells appeared to be connected with the thyroglossal duct, and not with any other structure (fig. 4).
Three internal pharyngeal pouches were present in the form of narrow slit-like clefts. Behind the third cleft the cavity dilated transversely in the form of a wide flange-like lateral expansion. This appears to be the rudiment from which the fourth and fifth internal pharyngeal pouches are derived later.
Caudal to this dilatation the lumen contracted and became altered in shape. It was flattened from side to side, and the lining epithelium of the dorsal and ventral walls become different, that on the ventral aspect being long and columnar, while that of the dorsal half remained short and rather flat (fig. 12).
Still lower the tube became constricted about its middle,and shortly afterwards divided into a short ventral median lung-bud and oesophagus.
The median dorsal pharyngeal wall behind the level of the third pouch was found to be still in connexion with the notochord. At that level there was a small median cleft-like recess lined with epithelial cells, which extended for a short distance dorsally in the posterior part of the roof of the pharynx. The notochord was found to be connected with the cells lining the caudal part of this recess, and it was absent along the whole length of the recess. At this level apparently the notochord is late in being folded off from the endothelium of the vitello-intestinal cavity. This observation is in harmony with, and to some extent explains, the results obtained by HuberI in his examination of the development of the pharyngeal bursa, for this region coincides with an area of the pharyngeal wall in which, in embryos of 5-8 mm. length, the notochord is found in close contact with the pharyngeal endoderm, representing the seat of development of the pharyngeal bursa. It also confirms Meyer's [3] observations on the occasional persistence of a connexion between the chorda and the pharyngeal entoderm.

Fig. 6. Section of head end of embryo. |

Fig. 7. Section at level of liver bud. |

Fig. 8. Section cranial to vitello-intestinal junction. |

Fig. 9. Section |

Fig. 10. Section |

Fig. 11. Section |
A short distance caudal to the origin of the lung-bud the lumen of the foregut showed a slight dorso-ventral dilatation, marking the position of the stomach, and a short distance beyond this level the vitello-intestinal junction was found. The liver-bud was present as a tubular diverticulum arising from the the proximal portion of a groove on the ventral or cephalic aspect of the stalk of connexion. The hollow tubular pocket passed for a short distance in the substance of the septum transversum, in close relation to the vessels of that septum, and these vessels for the greater part lay dorsally to it. There was no trabecular formation as yet, nor was there any indication of a pancreatic diverticulum (fig. 7).
The hindgut formed a tubular and for the most part cylindrical ento- dermal tube, and terminated in the dilatation of the cloaca (fig. 5).
Over a considerable extent the mesodermal wall of the hindgut showed a trefoil appearance on section,produced by a surface depression on the ventral and on each lateral aspect near the root of the mesentery. This appearance was due rather to the prominence of vessels in the inter- vening areas than to any constriction of the wall.
The allantois arose from the ventral aspect of the hindgut at the upper part of the cloaca, but could not be traced for any distance into the body stalk. The lumen of the distal part of the cloaca ventrally is separated from the surface merely by its layer of lining entodermal cells and by the investing ectoderm, the cloacal membrane.
Its lumen is compressed from side to side, but is of some size in a dorso-ventral direction (fig. 9). The terminal parts of the Wolffian ducts end blindly on each side of it.
Respiratory System

(fig.12). The stage of development of the respiratory system shows the lung- bud as a median, short, somewhat cylindrical outgrowth from the lower part of the pharynx. It resembles the condition figured and described in an embryo 4 25 mm. vertex breech length (R. Meyer, No. 399), the principal difference being that in my specimen the median unpaired lung-bud is somewhat longer than in that specimen (fig.12).
It arose from the ventral aspect of the floor of the pharynx, the lumen taking origin in a long narrow groove which occupied the floor of the pharynx immediately above the level at which the lung-bud becomes free.
It lay ventral and parallel to the upper part of the Esophagus, and the upper part was cylindrical in outline. Lower down it became widened in the transverse axis, and terminated in a knob-like enlargement, flattened dorso-ventrally, the lateral expansions forming the only indication of the origin of the paired lung-buds from the median unpaired original outgrowth.
There were no pulmonary arteries, but two pulmonary veins arose from a small plexus about the lower part of the respiratory outgrowth and passed independently of the other venous channels of the body into the atrium of the heart just on the atrial side of the sinu-atrial junction.
Vascular System
Arteries

The arterial vessels are shown in figure 13.
From the heart a short conus arteriosus passed towards the floor of the pharynx, and divided into right and left branches, from which the bronchial arch vessels of the two sides arose.A rudimentary and incomplete first arch entered the tissue of the first arch, but did not reach the dorsal end of that arch.
In both the second and third arches a short wide vessel was present, and passed dorsally to join the head dorsal aorta.
This dorsal stem was prolonged forwards towards the root of the optic vesicle, and there divided. The posterior branch passed dorsally in the concavity of the cephalic flexure and extended backwards to the midbrain. Short vessels passed from it to the neural tube, and in the region of the neurorneres these branches were arranged segmentally, one vessel running in each of the surface grooves which separated the neuromere segments from one another.
The grooves between the 4th, 5th, and 6th segments were occupied by two short vessels which appeared to be branches of the dorsal aorta, one arising on the cranial and the other on the caudal side of the junction of the third arterial arch. These vessels did not communicate with one another. This stage of development of the cephalic and cerebral blood- vessels forms a stage intermediate between the one existing in the embryo of 23 soinites described by Felix, and that shown in Ingall's reconstruction of the head vessels in an embryo of 49 mm. long, both of which are figured in the Keibel-Mall Manuaclt, vol.i.pp.602 and 606. In the former the first and second arches were complete, but there were no prolongation forwards dorsally to the forebrain, nor any branches to the mid and hindbrains. In the latter there were four complete arches, whose dorsal ends are united, and there was also a cerebral blood-supply from a primitive internal carotid and vertebral artery.
The dorsal aortae united opposite the sixth somite,and passed thence almost to the end of the neural tube. The branches which arose from the aortae were somewhat irregular in their arrangement.
This may possibly be due to some vessels having been omitted or overlooked through their not containing any blood-cells,but against this is the fact that the vessels were everywhere packed full of blood-cells, which rendered their recognition easy, even though the lumen of some of the branches was large enough only to contain a single blood-cell. Segmental dorsal offshoots were found in the region of the 5th, 6th, 7th,and 8th somites,but a connexion between them and the aorta could not be followed out. There were also occasional lateral branches passing to the region of the Wolffian body, arranged somewhat irregularly.
There was a definite series of vessels passing to the intestinal tube. These vessels can be arranged in groups, consisting of a set arising from the aorta in the region of the 5th, 6th, 7th, and 8th somites, which went to the region of the gastric enlargement, while behind them the vitelline arteries arose by a series of rootlets three in number, which united to form a single vessel on each side.
Still further back, a series of ventral branches passed to the region of the hindgut, while in the region of the tail bend the aorta terminated in the manner described by Evans in a plexus of capillary-like vessels (the -ventral arch system of Felix), from the lower of which the umbilical arteries arose by multiple roots, while from the dorsal aspect of the plexus a series of dorsal segmental arteries passed towards the back, and occupied the intervals between the somites of this region.
Veins

(fig. 14). In the region of the hindbrain, an extensive capillary network gave origin to a vessel of some size, the vena capitis medialis, lying between the otic vesicle and the brain. This vessel continued downwards, received irregular tributaries, and passed onwards in the segmental area as the anterior cardinal vein.
The duct of Cuvier was forced in the lateral body wall at the level of the 3rd somite, and passed to the upper margin of the septum transversum. The posterior cardinal vein could be traced as far back as to the lower part of the 9th somite, or, in other words, as far as the upper end of the mesonephricduct. Below that level there was some indication of a few capillary-like vessels in the position of the subcardinal vein.
The right and left umbilical veins passed at the junction of body wall and amnion and turned medially to join the upper lateral part of the septum transversum, and to the right and left extremities of the sinus venous respectively. The vitelline veins were of very large size.
In the lower part of the septum transversum they fused with one another, forming a single large vessel lying on the ventral aspect of the intestine, and above that level they separated and terminated, close to one another, in the transverse part of the sinus venosus.
Nervous System

Central Nervous System
There was no histological differentiation of the wall of the neural tube by which its different segments could be separated from one another, nor were there any definite constrictions limiting the different parts of the cerebral region.
It is, however, clear that the midbrain occupied the region of the cephalic bend, and it passed anteriorly into the forebrain, with which are associated the optic vesicles, and posteriorly inta the hindbrain, which has certain structural characters by which it can be identified. The hind- brain was somewhat dilated, and its neural wall was incomplete dorsally in part.
The hindbrain merged in turn into the spinal portion of the neural tube.
The hindbrain showed transverse segmentation (fig. 15) in the form of bilateral and symmetrical constrictions of the external surface, represented by elevations on the inner aspect, these elevations separating from one another the rhombic grooves. In the intervals between the surface constrictions the wall was thickened, and thus the internal segmentation was less obvious than that of the external surface. Seven such segments were present, extending from the region of the cephalic flexure backwards; and certain of these segments had a distinct relationship especially to the origin of the trigeminal nerve, to the acoustic nerve, and to the otic vesicle, as is indicated in the figure.
The relationship of the surface constrictions to the blood-vessels has been pointedout,but thesegmentation isnot due to these vessels, since the actual thickness of the wall is different in the segments and in the intervals.
The number and arrangement of the segments correspond closely to those figured and described by Thompson in the 2 - 5 mm. embryo already referred to, and it need only be added here that the trigeminal nerve was associated with the 2nd anterior segment, the acoustic nerve with the 4th, while the otic-vesicle lay opposite the 5th segment.
Spinal Cord
The structure of the spinal cord showed no features requiring special description, since it was typical of this stage. There was no indication of the outgrowth of spinal nerves.
Neutral Crest
The neural crest extended from the tail region as far forwards as to the last neuromere of the hindbrain in an unbroken band.
In section it had the form of a triangular narrow lamina, lying between the dorsal portion of the neural tube and the mesial border of the somites. This lamina showed a slight amount of segmentation alternating with the mesodermic somites. As is seen in the figure, it did not extend as far forward's as to the otic vesicle.
Organs of Special Senses
The special-sense organs which had made their appearance were the olfactory, the optic, and the auditory.
Olfactory Organ
An early stage of the development of the olfactory organ was represented by a thickening of the surface of the epithelium immediately ventral to the root of the optic vesicles. This thickened epithelium occupied a shallow spherical dimple on the surface of the side of the head, but no nerve connexion between it and the neural tube was present (fig. 6).
Optic Vesicles
The position, shape, and form of the optic vesicles are shown in fig. 5 and fig. 6. They arose from the ventro-lateral aspect of the fore-brain, and formed two large hollow vesicles, which projected dorso-laterally and lay almost in contact with the surface of the forebrain, and gradually separated from it. There was no definite stalk of connexion, but their base was constricted near the point of the attachment. The cavity of the fore-brain was in free communication by a large aperture on each side with the cavity of each of the vesicles (fig. 4).
There was no indication of the formation of the lens, but the lateral surface of each of the vesicles was indented, and a slight groove ran from this hollow towards the region of attachment to the brain, constituting an early stage of the choroidal fissure.
Otic Vesicles
These two vesicles are shown in section in fig. 15. They formed on each side an oval hollow structure, whose communication with the surface of the head is closed, a narrow stalk passing from the dorsal part of the vesicle towards the surface, but not opening upon it.
Cranial Nerves
Trigeminal
The position of the trigeminal rudiment is indicated by an elevation on the lateral surface of the head, a short distance rostral to the elevation produced by the otic vesicle. The area was irregularly oval in shape, and was due to an area of marked proliferation in the epithelium and subjacent mesenchymal tissue. The general characters of this area are shown in fig. 15.
Sections dorsal to this level show that the dorsal aspect of this area is connected by a column of deeply staining cels with the surface of the 2nd neuromere.
fig. 5 gives a diagrammatic representation of the surface area and its connexion,and brings out the fact that there is a definite placode-like formation in connexion with the early development of the trigeminal nerve.
The proliferative area and the cellular strand appear to be similar to theso-called "epibranchial pro ganglion" and the "pro nerve," described and figured by GiglioIos in a human embryo of the seventeenth day.
Acustico-facial Nerve
This nerve complex was present as a cellular band passing from the rostral border of the otic vesicle to the 4th neuromere of the hindbrain, and it is shown in fig. 5 and fig. 15.
Vagus and Glossopharyngeal
The rudiments of these nerves were not so fully developed as the two former. The only traces found consisted of a thin cellular strand connected with the hindbrain behind the 5th neuromere, and passing towards the surface.
This strand resembled a portion of the neural crest, but was situated above the cephalic end of that structure.
Excretory System
The excretory system consisted of the Wolffian duct and of a series of mesonephric tubules at different stages of development( figs.8-11).
No pronephric tubules were present.
The Wolffian ducts began blindly at the level of the 9th. somite, and passed towards the cloaca, where they terminated blindly, between the 25th and 26th segments. The first tubule lay at the level of the 10th somite, which may be taken to be the 6th cervical segment, and thus the condition corresponds with the observations of Felix, who has not found mesonephric tubules further forwards than this level. The tubules were arranged irregularly, e.g. not segmentally, and were eighteen in number (fig. 14). The anterior ones communicated with the duct, while the more posterior ones were fre, and had not apparently established their connexion with it.
Heart

A separate model was made, at a magnification of 200 diameters, of the heart, pericardiumn, and the adjacent segment of the trunk, from which the three figures appended have been drawn (figs. 17-19). I have elsewhere[4] given a short account of some of the features of the heart in this embryo; but as Thompson's account of the heart in the R. Meyer embryo 300 is the only description of the heart at this stage, and as different interpretations have been put upon the appearances shown in his models, I have thought it advisable to give a more detailed account ofitnow.
It may be stated at once that in practically al its details the present specimenc corroborates Thompson's account, and omparison with his figures shows the degree of resemblance between the two specimens.
The heart consists of sinus venosus, single atrium, atrial canal, and of an additional ventricular chamber, conical in shape,with two limbs; the proximal of these limbs is connected on the left with the atrial canal and forms quite clearly a ventricle, while the distal chamber is more tubular in formn and leads cephalic to the short conus arteriosus.
This is the bulbus limb of the ventricular loop, and perhaps that term may be retained for descriptive purposes, without however committing us to the theory that this limb is to be taken to be the same as the bulbus cords of the reptilian heart. The angle at the junction of these two limbs constitutes the apex of the ventricular portion.
A long oblique groove separates the two limbs from one another, and it may be termed the bulbo-ventricular groove. The heart described by Thompson shows what I take to be a similar condition of the ventricular portion of the heart, the principal difference being that the bulbo-ventricular groove in his specimen runs almost horizontally, and that therefore the "ventricular loop" is caudal in position and the "bulbus" loop cephalic to it.
There is evidence that the bulbo-ventricular groove ontogenetically becomes more and more oblique, and is eventually almost vertical in direction, and I take it that this difference between the two specimens means that the former one is at a slightly earlier stage than the one described here. The conus arteriosus is a short stem which soon divides into branches, as hasbeendescribedearlier.
Sinus venosus

The sinu-atrial junction is placed on the right side of the heart, and the terminal portion of the sinus venosus is in continuity with the atrium. The remainder of the sinus venous is placed in the upper part of the septum transversum, and is seen in fig. 19 as it passes in the septum across to the leftside. The right end received the right duct of Cuvier and the right umbilical vein,and the left end the corresponding vessels of the left side, while the right and left vitelline vessels opened into the crosspiece. There was no indication of the left venous valve at this stage.
Atrium

This constituted the dorsal and cephalic portion of the heart, and projected on each side, but mainly to the left.
Its ventral wall was grooved by the bulbo-ventricular groove, and the infolding of the wall produced thus constitutes a rudiment of the septum primum of the atrium, most distinct on the ventral wall of that chamber and passing onto the floor and the roof. The myocardial and the endothelial walls were in apposition with one another in these two chambers for the greater part, but became separated as the atrial canal is approached. Between these walls in the other chambers of the heart is seen the delicate reticulum which has been shown to occur in other specimens about this stage[5] (see Atrial Canal). The position and size of the endothelial channel are shown in fig. 16 and fig. 18 , which show that there is a very narrow endothelial tube, separated from the myocardial wall by a wide delicate reticulum. The ventricular limb of the ventricle had myocardial wall of moderate thickness, which showed trabeculae on its inner surface. The bulbo-ventricular groove caused a slight infolding of the wall by which the two limbs are separated as by a partial septum on the ventral aspect, and the ridge constituted a partial bulbo-ventricular septum.
The sinu-atrial junction was a narrow, vertical, slit-like orifice, and there was an infolding of the junction of the two chambers on the right of the orifice, forming a right venous valve.
The ascending or " bulbus " limb passed first headwaids and then horizontally to the left.
At the junction of the two parts was a marked dilatation of the myoepicardial wall.
The myoepicardial and endothelial walls were widely separated from one another in the whole length of the "bulbus"'limb. The interval between the two is shown in fig. 16 as a space, which, however, is occupied by a delicate reticulum, as is seen in the section reproduced here (fig. 16).
The significance and the homologies of the bulbus limb cannot be discussed without reference to older specimens, and I do not propose to go into that question here.
Coelom


The intra-embryonic and the extra-embryonic divisions of the coelom communicated with one another freely by the opening on each side of the vitello-intestinal connexion, between it and the body wall (fig.1). The characters of the latter division have been described with the chorionic vesicle.
The intra-embryonic coelom may be conveniently described under the divisions which it presents into pericardial sac, peritoneal sac, and the peritoneo-pericardial communications, which connect the two former.
The dorsal wall of these portions of the body cavity is shown in fig. 20. There was a median continuous mesentery, and the extent of its attachment is shown in that figure. The cephalic portion constitutes the dorsal mesocardium, and below the level of the heart the mesentery was attached to the dorsal surface of the septum transversum, and thus formed the ventral enteric mesentery. In using the term "mesentery" at this stage, it must be pointed out that there is no proper mesentery, for the alimentary canal lies embedded in the trunk of the embryo (fig.9), and this mesenteric structure is merely a connexion between the dorsal wall of the embryo on the one hand and the heart and septum transversum ventrally.
The body wall on each side contain the ducts of Cuvier, which passed to the septum transversum, and the level of these two vessels gives the level of the upper margin of the septum transversum, whose cephalic surface constitutes the floor of the pericardial cavity.
The dorsal wall of the coelom showed a rounded bulging projection on each side of the mesenteric root. There can be no doubt that these bulgings represent the mesodermal elements of the lungs, which are already being formed as a thickening of mesenchymal tissue, which islater penetrated by the growing hypoblastic tubes, but the epithelial lung-bud does not descend. as yet to their level.
The pericardio-peritoneal communication is even at this stage being narrowed by these thickenings, which lay at a lower level than the upper margin of the septum transversum.
The figure (20) also shows the narrow orifices of the pneumato-enteric recesses.
These diverticula from the intra-emnbryonic coelom were present on both sides,as they were in the 3mm. embryo in which they have been found by Broman.[6] From each of the slit-like openings a pocket passed in a cephalic direction, tunnelling the body wall, and lying by the side of the alimentary canal.
Their lining membrane consisted of thickened coelomic epithelium. No special significance attaches to this fact of its being thicker here than else- where, as the coelomic epithelium tends to become thickened in regions where two surfaces are in contact.
Caudal to the vitello-intestinal junction, the peritoneal coelom presented no features of special significance, and its general character is shown in the sections figured through that region of the embryo.
Septum Transversum
The septum transversum at this stage lay very obliquely, in a dorso-ventral and cephalo-caudal axis.
I have included a figure (19) which shows its cephalic surface, forming the floor of the pericardium, and its lateral continuity, with the body wall. This cephalic surface is flat; the caudal surface, on the other hand, presents on each side of the mesial plane a large dorsal swelling, produced by the vitelline veins. The shape resembles that of the liver, the prominent veins being apparently the forerunners of the subsequent right and left lobes.
Summary of Characters of the Embryo Described Above
- Designation. - 2W.I.
- Dimensions.-
- Chorionic vesicle. . .8-5x7-7x4mm.
- Length: Amniotic sac 3mm. Embryo 2.9 (max.) x 1.2 dorso-ventral x 9 mm. (transverse at heart). Yolk-sac 26 x 2 (dorso-ventral) x 1.6 (transverse) mm.
- Age. - Period elapsed since beginning of last menstruation to commencement of abortion, 42 days. Period elapsed since due date of lapsed menstruation, 10 days.
- Body Form. - General uniform dorsal curvature and cephalic flexure. Slight spiral twisting. Blunted tail projection. Body stalk passes to the right side. Yolk-sac lies to the left.
- Mesodermal Somites.-Twenty-seven pairs.
- Chorda. - Almost completely formed, except in dorsal pharyngeal region, where it is not yet folded of. Terminates in dorsal wall of Rathke's pocket.
- Nervous System. - Neural tube completely closed. Forebrain,midbrain hindbrain, the latter with seven neuromeres. Neural crest. Cranial nerves, trigeminal, anda custico-facial. Vago-glossopharyngeal rudimentary.
- Eye.- Primary optic vesicles separated from surface ectoderm by mesenchyme. No lens plate.
- Ear.- Otic vesicles embedded in mesenchyme near surface of hindbrain and unconnected with the surface ectoderm.
- Nose.- Olfactory area present, as a rounded dimple, with thickened epithelium.
- Hypophysis.- Dorsal pocket from buccal cavity.
- Mouth.- Bucco-pharyngeal membrane ruptured.
- Alimentary Tract.- Intestine connected with yolk-sac by very small channel. Liver-bud present; no pancreatic bud.
- Pharyngeal Pockets,etc.-Three in number. Patent thyroglossal duct.Lung-bud, median ventral offshoot from lower end of pharynx.
- Urogenital System. - Wolffian duct present, but not connected with cloaca. Seventeen pairs of tubules. No pronephric structures.
- Heart and Vessels. - Sinus venous, single atrium, atrial canal, and ventricle with a left descending ventricular limb and right ascending "bulbus limb," leading to short conus arteriosus. Sinu-atrial junction with right venous valve. Continuous myo-epicardial wall. Endothelial tube widely separated from myo-epicardial wall in the "bulbus" limb, ventricular limb, and atrial canal. Incomplete first and complete second and third pharyngeal arterial arches and dorsal aortae. Anterior cerebral artery. Dorsal segmental arteries and ventral segmental vessels to intestine, arranged irregularly.
- Amnion.- Ectodermal and mesodermal layers. Invests dorsal aspect of body stalk.
- Allantois. - Traced for a short distance from intestine.
- Remarks.- Obtained in a decidual cast, passed 10 days after a missed menstrual period. Formalin fixation. Cut in parafin. Histology on the whole is good. Neural tube somewhat shrunken. Mitotic figures visible.
References
Literature .- WADE AND WATSON, Jour. of Obstetrics and Gynecol. Brit. Empire, Feb. 1910.
Images
Carnegie Staging Comparison: A 27 somite stage embryo would be similar to a Carnegie stage 12 (26 - 30 days), caudal neuropore closes, Somite Number 21-29.
- 27 Somite Paper: Fig 1 | Fig 2 | Fig 3 | Fig 4 | Fig 5 | Fig 6 | Fig 7 | Fig 8 | Fig 9 | Fig 10 | Fig 11 | Fig 12 | Fig 13 | Fig 14 | Fig 15 | Fig 16 | Fig 17 | Fig 18 | Fig 19 | Fig 20 | Carnegie stage 12
| Historic Disclaimer - information about historic embryology pages |
|---|
|
- Historic Paper Links: 13-14 Somites | 22 Somites | 23 Somites | 25 Somites | 27 Somites | Mall Human Embryo Collection | Embryology History | Carnegie stage 11 | Carnegie stage 12 | Journal of Anatomy | Embryonic Development | Category:Historic Embryology
Reference
Waterston D. A human embryo of twenty-seven pairs of somites, embedded in decidua. (1914) J Anat Physiol., 49(1): 90-118 PMID 17233016
Cite this page: Hill, M.A. (2026, April 18) Embryology Paper - A Human Embryo of Twenty-seven Pairs of Somites, Embedded in Decidua. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_A_Human_Embryo_of_Twenty-seven_Pairs_of_Somites,_Embedded_in_Decidua
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G
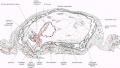
Fig. 1
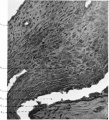
Fig. 2
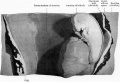
Fig. 3

Fig. 4
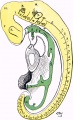
Fig. 5
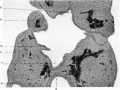
Fig. 6. Section of head end of embryo.
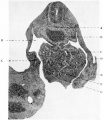
Fig. 7
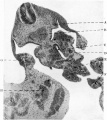
Fig. 8
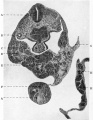
Fig. 9
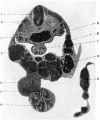
Fig. 10
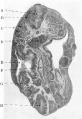
Fig. 11
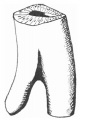
Fig. 12
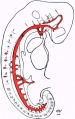
Fig. 13. Linear profile reconstruction of arteries.
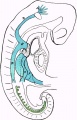
Fig. 14. Linear reconstruction of veins and of excretory system.
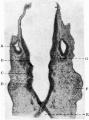
Fig. 15
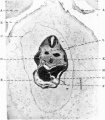
Fig. 16
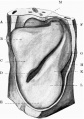
Fig. 17
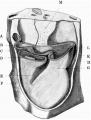
Fig. 18
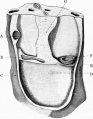
Fig. 19
Fig. 20

Animation
| Historic Disclaimer - information about historic embryology pages |
|---|
|
The information below was not part of the above original historic article.
Animation
Animation was generated from the original Figure 5, 13 and 14.

- Carnegie Stages: 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | About Stages | Timeline
Carnegie Stage Table
Weeks shown in the table below are embryonic post ovulation age, for clinical Gestational Age (GA) measured from last menstrual period, add 2 weeks.
(not to scale) |
||||
 |
fertilized oocyte, zygote, pronuclei | |||
 |
morula cell division with reduction in cytoplasmic volume, blastocyst formation of inner and outer cell mass | |||
 |
loss of zona pellucida, free blastocyst | |||
| attaching blastocyst | ||||
(week 2) |
 |
implantation | ||
 |
extraembryonic mesoderm, primitive streak, gastrulation | |||
| gastrulation, notochordal process | ||||
| primitive pit, notochordal canal | ||||
 |
Somitogenesis Somite Number 1 - 3 neural folds, cardiac primordium, head fold | |||
| Somite Number 4 - 12 neural fold fuses | ||||
| Somite Number 13 - 20 rostral neuropore closes | ||||
| Somite Number 21 - 29 caudal neuropore closes | ||||
| Somite Number 30 leg buds, lens placode, pharyngeal arches | ||||
| lens pit, optic cup | ||||
| lens vesicle, nasal pit, hand plate | ||||
| nasal pits moved ventrally, auricular hillocks, foot plate | ||||
| finger rays | ||||
| ossification commences | ||||
| straightening of trunk | ||||
| upper limbs longer and bent at elbow | ||||
| hands and feet turned inward | ||||
| eyelids, external ears | ||||
| rounded head, body and limbs | ||||
The embryos shown in the table are from the Kyoto and Carnegie collection and other sources.
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2026, April 18) Embryology Paper - A Human Embryo of Twenty-seven Pairs of Somites, Embedded in Decidua. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_A_Human_Embryo_of_Twenty-seven_Pairs_of_Somites,_Embedded_in_Decidua
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G