Introduction
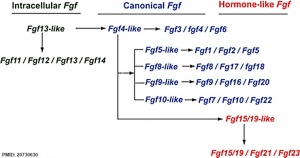
Fgf gene family evolution
[1]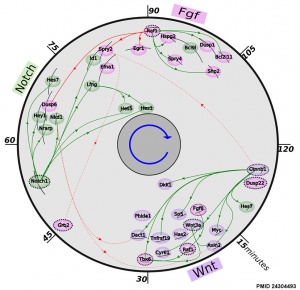
Mouse somitogenesis genes
[2]Fibroblast Growth Factors (FGF) were originally identified by their ability to stimulate fibroblast cell proliferation[3][4] but have since been identified as having a role in a growth of a number of different tissues development and differentiation and continue to have a role in the adult.
The first two identified factors were originally given the simple nomenclature of "acidic" or "basic". We now know there to be at least 22 different human FGFs (Fgf1–Fgf23). These protein growth factors are bound by 4 different cell membrane receptors (FGFR1-4). FGFRs belong to the tyrosine kinase receptor family.
The mammalian Fgf family can be divided into the intracellular Fgf11/12/13/14 subfamily (iFGFs), the endocrine hormone-like Fgf15/21/23 subfamily (hFGFs), and the paracrine canonical Fgf subfamilies, including Fgf1/2/5, Fgf3/4/6, Fgf7/10/22, Fgf8/17/18, and Fgf9/16/20.
Human FGF Family
| Table - Human Fibroblast Growth Factor Family FGF
|
Approved
Symbol
|
Approved Name
|
Previous
Symbols
|
Synonyms
|
Chromosome
|
| FGF1 |
fibroblast growth factor 1 |
FGFA |
"AFGF, ECGF, ECGFA, ECGFB, HBGF1, ECGF-beta, FGF-alpha, GLIO703" |
5q31.3
|
| FGF2 |
fibroblast growth factor 2 |
FGFB |
|
4q28.1
|
| FGF4 |
fibroblast growth factor 4 |
HSTF1 |
"K-FGF, HBGF-4, HST, HST-1, KFGF" |
11q13.3
|
| FGF5 |
fibroblast growth factor 5 |
|
|
4q21.21
|
| FGF6 |
fibroblast growth factor 6 |
|
|
12p13.32
|
| FGF7 |
fibroblast growth factor 7 |
|
KGF |
15q21.2
|
| FGF8 |
fibroblast growth factor 8 |
|
AIGF |
10q24.32
|
| FGF9 |
fibroblast growth factor 9 |
|
|
13q12.11
|
| Links: Developmental Signals - Fibroblast Growth Factor | OMIM Fgf1 | HGNC | Bmp Family | Fgf Family | Sox Family | Tbx Family
|
| Human FGF Family
|
| Table - Human Fibroblast Growth Factor Family FGF
|
Approved
Symbol
|
Approved Name
|
Previous
Symbols
|
Synonyms
|
Chromosome
|
| FGF1 |
fibroblast growth factor 1 |
FGFA |
"AFGF, ECGF, ECGFA, ECGFB, HBGF1, ECGF-beta, FGF-alpha, GLIO703" |
5q31.3
|
| FGF2 |
fibroblast growth factor 2 |
FGFB |
|
4q28.1
|
| FGF4 |
fibroblast growth factor 4 |
HSTF1 |
"K-FGF, HBGF-4, HST, HST-1, KFGF" |
11q13.3
|
| FGF5 |
fibroblast growth factor 5 |
|
|
4q21.21
|
| FGF6 |
fibroblast growth factor 6 |
|
|
12p13.32
|
| FGF7 |
fibroblast growth factor 7 |
|
KGF |
15q21.2
|
| FGF8 |
fibroblast growth factor 8 |
|
AIGF |
10q24.32
|
| FGF9 |
fibroblast growth factor 9 |
|
|
13q12.11
|
| Links: Developmental Signals - Fibroblast Growth Factor | OMIM Fgf1 | HGNC | Bmp Family | Fgf Family | Sox Family | Tbx Family
|
|
Protein Properties
Human FGF
- ~150–300 amino acids
- have a conserved ~120-residue core with ~30–60% identity
Some Recent Findings
- Ovotesticular Disorders of Sex Development (DSD) in FGF9 Mouse Models of Human Synostosis Syndromes[5] "In mice, male sex determination depends on FGF9 signalling via FGFR2c in the bipotential gonads to maintain expression of the key testis gene SOX9. In humans however, while FGFR2 mutations have been linked to 46,XY disorders of sex development (DSD), the role of FGF9 is unresolved. The only reported pathogenic mutations in human FGF9; FGF9S99N and FGF9R62G, are dominant, and result in craniosynostosis (fusion of cranial sutures) or multiple synostoses (fusion of limb joints). Whether these synostosis-causing FGF9 mutations impact upon gonadal development and DSD etiology has not been explored. We therefore examined embryonic gonads in the well-characterised Fgf9 missense mouse mutants; Fgf9S99N and Fgf9N143T, which phenocopy the skeletal defects of FGF9S99N and FGF9R62G variants respectively. XY Fgf9S99N/S99N and XY Fgf9N143T/N143T fetal mouse gonads showed severely disorganised testis cords and partial XY sex reversal at E12.5 days postcoitum (dpc), suggesting loss of FGF9 function. By E15.5 dpc, testis development in both mutants had partly recovered. Mitotic analysis in vivo and in vitro suggested that the testicular phenotypes in these mutants arise in part through reduced proliferation of the gonadal supporting cells. These data raise the possibility that human FGF9 mutations causative for dominant skeletal conditions can also lead to loss of FGF9 function in the developing testis, at least in mice. Our data suggest that in humans, testis development is largely tolerant of deleterious FGF9 mutations which lead to skeletal defects, thus offering an explanation as to why XY DSDs are rare in patients with pathogenic FGF9 variants."
- The Warburg Effect and lactate signaling augment Fgf-MAPK to promote sensory-neural development in the otic vesicle[6] "Recent studies indicate that many developing tissues modify glycolysis to favor lactate synthesis, but how this promotes development is unclear. Using forward and reverse genetics in zebrafish, we show that disrupting the glycolytic gene phosphoglycerate kinase-1 (pgk1) impairs Fgf-dependent development of hair cells and neurons in the otic vesicle and other neurons in the CNS/PNS. Fgf-MAPK signaling underperforms in pgk1-/- mutants even when Fgf is transiently overexpressed. Wild-type embryos treated with drugs that block synthesis or secretion of lactate mimic the pgk1-/- phenotype, whereas pgk1-/- mutants are rescued by treatment with exogenous lactate. Lactate treatment of wild-type embryos elevates expression of Etv5b/Erm even when Fgf signaling is blocked. However, lactate's ability to stimulate neurogenesis is reversed by blocking MAPK. Thus, lactate raises basal levels of MAPK and Etv5b (a critical effector of the Fgf pathway), rendering cells more responsive to dynamic changes in Fgf signaling required by many developing tissues."
- ERK Activity Dynamics during zebrafish Embryonic Development[7] "During vertebrate development, extracellular signal-regulated kinase (ERK) is activated by growth factors such as fibroblast growth factor (FGF), and it regulates the formation of tissues/organs including eyes, brains, somites, limbs, and inner ears. However, an experimental system to monitor ERK activity dynamics in the entire body of the vertebrate embryo is lacking. We recently studied ERK activity dynamics in the pre-somitic mesoderm of living zebrafish embryos injected with mRNAs encoding a Förster resonance energy transfer (FRET)-based ERK biosensor. ... A spatiotemporal map of ERK activity in the entire body during zebrafish embryogenesis was generated, and previously unidentified activation dynamics and ERK domains were identified."
- BMP and FGF signaling interact to pattern mesoderm by controlling basic helix-loop-helix transcription factor activity[8] "The mesodermal germ layer is patterned into mediolateral subtypes by signaling factors including BMP and FGF. How these pathways are integrated to induce specific mediolateral cell fates is not well understood. We used mesoderm derived from post-gastrulation neuromesodermal progenitors (NMPs), which undergo a binary mediolateral patterning decision, as a simplified model to understand how FGF acts together with BMP to impart mediolateral fate. Using zebrafish and mouse NMPs, we identify an evolutionarily conserved mechanism of BMP and FGF mediated mediolateral mesodermal patterning that occurs through modulation of basic helix-loop-helix (bHLH) transcription factor activity. BMP imparts lateral fate through induction of Id helix loop helix (HLH) proteins, which antagonize bHLH transcription factors, induced by FGF signaling, that specify medial fate. We extend our analysis of zebrafish development to show that bHLH activity is responsible for the mediolateral patterning of the entire mesodermal germ layer."
- A review of FGF signaling in palate development[9] "The fibroblast growth factors (FGFs) play a critical role during palatogenesis by mediating a variety of cellular responses. Extensive epidemiological and genetic studies over several decades in humans have revealed members of the FGF family function as candidate genes for syndromic and nonsyndromic cleft lip and cleft palate. The findings that FGFs signaling work delicately in the development of palate have been confirmed in mice carrying targeted mutations. Here we try to review recent progress toward a detailed understanding of FGF signaling including FGF7, FGF8, FGF9, FGF10, FGF18 and their receptors FGFR1, FGFR2 in palate development studies and discuss how they interact with other factors on the basis of animal studies regarding cleft palate." palate
|
| More recent papers
|
|
This table allows an automated computer search of the external PubMed database using the listed "Search term" text link.
- This search now requires a manual link as the original PubMed extension has been disabled.
- The displayed list of references do not reflect any editorial selection of material based on content or relevance.
- References also appear on this list based upon the date of the actual page viewing.
References listed on the rest of the content page and the associated discussion page (listed under the publication year sub-headings) do include some editorial selection based upon both relevance and availability.
More? References | Discussion Page | Journal Searches | 2019 References | 2020 References
Search term: Fibroblast Growth Factor | FGF | ERK | Fibroblast Growth Factor Receptor
|
| Older papers
|
| These papers originally appeared in the Some Recent Findings table, but as that list grew in length have now been shuffled down to this collapsible table.
See also the Discussion Page for other references listed by year and References on this current page.
- FGF8 morphogen gradients are differentially regulated by heparan sulphotransferases Hs2st and Hs6st1 in the developing brain[10] "Fibroblast growth factor (FGF) morphogen signalling through the evolutionarily ancient extracellular signalling-regulated kinase/mitogen activated protein kinase (ERK/MAPK) pathway recurs in many neural and non-neural developmental contexts, and understanding the mechanisms that regulate FGF/ERK function are correspondingly important. The glycosaminoglycan heparan sulphate (HS) binds to FGFs and exists in an enormous number of differentially sulphated forms produced by the action of HS modifying enzymes, and so has the potential to present an extremely large amount of information in FGF/ERK signalling. ...We discover that two different HS modifying enzymes, Hs2st and Hs6st1, indeed differentially modulate the properties of emerging Fgf8 protein concentration gradients and the Erk signalling output in response to Fgf8 in living tissue in ex vivo cultures. Both Hs2st and Hs6st1 are required for stable Fgf8 gradients to form as rapidly as they do in wild-type tissue while only Hs6st1 has a significant effect on suppressing the levels of Fgf8 protein in the gradient compared to wild type. Next we show that Hs2st and Hs6st1 act to antagonise and agonise the Erk signalling in response to Fgf8 protein, respectively, in ex vivo cultures of living tissue. Examination of endogenous Fgf8 protein and Erk signalling outputs in Hs2st-/- and Hs6st1-/- embryos suggests that our ex vivo findings have physiological relevance in vivo Our discovery identifies a new class of mechanism to tune Fgf8 function by regulated expression of Hs2st and Hs6st1 that is likely to have broader application to the >200 other signalling proteins that interact with HS and their function in neural development and disease."
- Review - The Multiple Roles of FGF Signaling in the Developing Spinal Cord[11] "During vertebrate embryonic development, the spinal cord is formed by the neural derivatives of a neuromesodermal population that is specified at early stages of development and which develops in concert with the caudal regression of the primitive streak. Several processes related to spinal cord specification and maturation are coupled to this caudal extension including neurogenesis, ventral patterning and neural crest specification and all of them seem to be crucially regulated by Fibroblast Growth Factor (FGF) signaling, which is prominently active in the neuromesodermal region and transiently in its derivatives. Here we review the role of FGF signaling in those processes, trying to separate its different functions and highlighting the interactions with other signaling pathways." spinal cord
- FGF8 coordinates tissue elongation and cell epithelialization during early renal tubulogenesis[12] "When a tubular structure forms during early embryogenesis, tubular elongation and lumen formation (epithelialization) proceed simultaneously in a spatiotemporally coordinated manner. We here demonstrate, using the Wolffian duct (WD) of early chicken embryos, that this coordination is regulated by the expression of FGF8, which shifts posteriorly during body axis elongation. FGF8 acts as a chemoattractant on the leader cells of the elongating WD and prevents them from epithelialization, whereas static ('rear') cells that receive progressively less FGF8 undergo epithelialization to form a lumen. Thus, FGF8 acts as a binary switch that distinguishes tubular elongation from lumen formation. The posteriorly shifting FGF8 is also known to regulate somite segmentation, suggesting that multiple types of tissue morphogenesis are coordinately regulated by macroscopic changes in body growth." chicken | renal
- Tissue-specific roles of Fgfr2 in development of the external genitalia[13] "Congenital anomalies frequently occur in organs that undergo tubulogenesis. Hypospadias is a urethral tube defect defined by mislocalized, oversized, or multiple openings of the penile urethra. Deletion of Fgfr2 or its ligand Fgf10 results in severe hypospadias in mice, in which the entire urethral plate is open along the ventral side of the penis. In the genital tubercle, the embryonic precursor of the penis and clitoris, Fgfr2 is expressed in two epithelial populations: the endodermally derived urethral epithelium and the ectodermally derived surface epithelium. Here, we investigate the tissue-specific roles of Fgfr2 in external genital development by generating conditional deletions of Fgfr2 in each of these cell types. These results demonstrate that urethral tubulogenesis, prepuce morphogenesis, and sexually dimorphic patterning of the lower urethra are controlled by discrete regions of Fgfr2 activity." genital
- The precise timeline of transcriptional regulation reveals causation in mouse somitogenesis network[2] "In vertebrate development, the segmental pattern of the body axis is established as somites, masses of mesoderm distributed along the two sides of the neural tube, are formed sequentially in the anterior-posterior axis. This mechanism depends on waves of gene expression associated with the Notch, Fgf and Wnt pathways."
- Prolonged FGF signaling is necessary for lung and liver induction in Xenopus[14] "FGF signaling plays numerous roles during organogenesis of the embryonic gut tube. Mouse explant studies suggest that different thresholds of FGF signaling from the cardiogenic mesoderm induce lung, liver, and pancreas lineages from the ventral foregut progenitor cells. The mechanisms that regulate FGF dose in vivo are unknown. Here we use Xenopus embryos to examine the hypothesis that a prolonged duration of FGF signaling from the mesoderm is required to induce foregut organs....These results suggest that the liver and lungs are specified at progressively later times in development requiring mesoderm contact for different lengths of time. Our data suggest that this is achieved at least in part through prolonged FGF signaling. In addition to providing a foundation for further mechanistic studies on foregut organogenesis using the experimental advantages of the Xenopus system, these data have implications for the directed differentiation of stem cells into foregut lineages."
- FGF-signaling gradient maintains symmetrical proliferative divisions of midbrain neuronal progenitors[15] "For the correct development of the central nervous system, the balance between self-renewing and differentiating divisions of the neuronal progenitors must be tightly regulated. To maintain their self-renewing identity, the progenitors need to retain both apical and basal interfaces. However, the identities of fate-determining signals which cells receive via these connections, and the exact mechanism of their action, are poorly understood. The conditional inactivation of Fibroblast growth factor (FGF) receptors 1 and 2 in the embryonic mouse midbrain-hindbrain area results in premature neuronal differentiation. Here, we aim to elucidate the connection between FGF-signaling and neuronal progenitor maintenance. Our results reveal that the loss of FGF-signaling leads to downregulation of Hes1 and upregulation of Ngn2, Dll1, and p57 in the ventricular zone (VZ) cells, and that this increased neurogenesis occurs cell-autonomously. Yet the cell-cycle progression, apico-basal-polarity, cell-cell connections, and the positioning of mitotic spindle in the mutant VZ appear unaltered. Interestingly, FGF8-protein is highly concentrated in the basal lamina. Thus, FGFs may act through basal processes of neuronal progenitors to maintain their progenitor status. Indeed, midbrain neuronal progenitors deprived in vitro of FGFs switched from symmetrical proliferative towards symmetrical neurogenic divisions. We suggest that FGF-signaling in the midbrain VZ is cell-autonomously required for the maintenance of symmetrical proliferative divisions via Hes1-mediated repression of neurogenic genes."
- Fgf8 morphogen gradient forms by a source-sink mechanism with freely diffusing molecules[16] "It is widely accepted that tissue differentiation and morphogenesis in multicellular organisms are regulated by tightly controlled concentration gradients of morphogens. How exactly these gradients are formed, however, remains unclear. Here we show that Fgf8 morphogen gradients in living zebrafish embryos are established and maintained by two essential factors: fast, free diffusion of single molecules away from the source through extracellular space, and a sink function of the receiving cells, regulated by receptor-mediated endocytosis. Evidence is provided by directly examining single molecules of Fgf8 in living tissue by fluorescence correlation spectroscopy, quantifying their local mobility and concentration with high precision. By changing the degree of uptake of Fgf8 into its target cells, we are able to alter the shape of the Fgf8 gradient. Our results demonstrate that a freely diffusing morphogen can set up concentration gradients in a complex multicellular tissue by a simple source-sink mechanism."
|
Ectoderm
neural crest cell migration during early chicken embryo development[17]
- "Fibroblast growth factor (FGF) signalling acts as one of modulators that control neural crest cell (NCC) migration, but how this is achieved is still unclear. In this study, we investigated the effects of FGF signalling on NCC migration by blocking this process. Constructs that were capable of inducing Sprouty2 (Spry2) or dominant-negative FGFR1 (Dn-FGFR1) expression were transfected into the cells making up the neural tubes. Our results revealed that blocking FGF signalling at stage HH10 (neurulation stage) could enhance NCC migration at both the cranial and trunk levels in the developing embryos. It was established that FGF-mediated NCC migration was not due to altering the expression of N-cadherin in the neural tube. Instead, we determined that cyclin D1 was overexpressed in the cranial and trunk levels when Sprouty2 was upregulated in the dorsal neural tube. These results imply that the cell cycle was a target of FGF signalling through which it regulates NCC migration at the neurulation stage."
Spinal Cord
Differentiation and localization of interneurons in the developing spinal cord depends on DOT1L expression[18]
"Genetic and epigenetic factors contribute to the development of the spinal cord. Failure in correct exertion of the developmental programs, including neurulation, neural tube closure and neurogenesis of the diverse spinal cord neuronal subtypes results in defects of variable severity. We here report on the histone methyltransferase Disruptor of Telomeric 1 Like (DOT1L), which mediates histone H3 lysine 79 (H3K79) methylation. Conditional inactivation of DOT1L using Wnt1-cre as driver (Dot1l-cKO) showed that DOT1L expression is essential for spinal cord neurogenesis and localization of diverse neuronal subtypes, similar to its function in the development of the cerebral cortex and cerebellum. Transcriptome analysis revealed that DOT1L deficiency favored differentiation over progenitor proliferation. Dot1l-cKO mainly decreased the numbers of dI1 interneurons expressing Lhx2. In contrast, Lhx9 expressing dI1 interneurons did not change in numbers but localized differently upon Dot1l-cKO. Similarly, loss of DOT1L affected localization but not generation of dI2, dI3, dI5, V0 and V1 interneurons. The resulting derailed interneuron patterns might be responsible for increased cell death, occurrence of which was restricted to the late developmental stage E18.5. Together our data indicate that DOT1L is essential for subtype-specific neurogenesis, migration and localization of dorsal and ventral interneurons in the developing spinal cord, in part by regulating transcriptional activation of Lhx2."
Endoderm
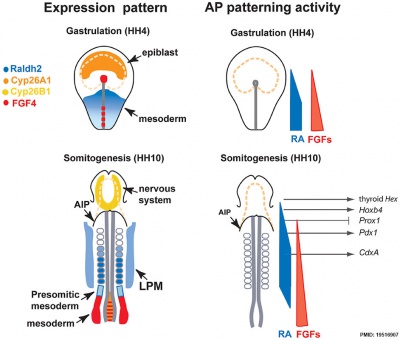
Chicken antero-posterior endoderm patterning[19]
- Links: endoderm | chicken
Mesoderm
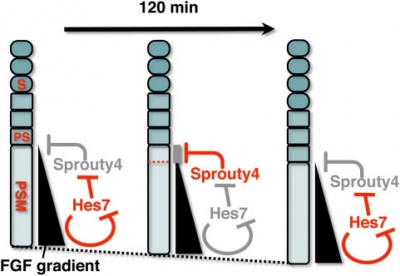
A Putative Model for the role of Sprouty4 as a mediator that links the mouse segmentation clock to the gradient of FGF signaling.[20]
The FGF signaling may be periodically inhibited by Sprouty4, by which temporal periodicity of Notch segmentation clock may be translated to spatial periodicity of the array of somites.
- In the PSM - FGF signaling establishes a posterior-to-anterior gradient, which is involved in the positioning of presumptive somite boundaries.
- Cyclic Sprouty4
- which is controlled by the Notch segmentation clock, the mechanism of which includes negative feedback loop of Hes7,
- may inhibit the FGF signaling possibly around the anterior border of the FGF signaling positive area
- where the FGF signaling is close to its threshold.
- S - somite
- PS - presumptive somite.
- Links: somitogenesis | Axial Skeleton Development | Notch | FGF
Bone
FGF9 has been shown to induce endochondral ossification in cranial mesenchyme in the mouse model.[21]
Respiration
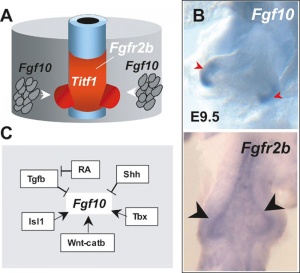
Mouse Respiratory Development Fibroblast Growth Factor Signaling
[22]Lung Buds
Fibroblast growth factor 10 (FGF10) expression in mesoderm required for initial lung buds, through FGFR2IIIb transmembrane tyrosine kinase receptor protein.
Branching
Fibroblast growth factor 10 (FGF10) and sonic hedgehog (SHH) form a feedback loop for branching
- mesenchyme produced FGF10 signals to the distal epithelium to upregulate SHH expression.
- SHH then feeds back to inhibit Fgf10 expression in the adjacent mesenchyme, dividing in two the Fgf10 expression domain.
- new FGF10 signaling domains serve as two chemoattractant sources, leading to bifurcation of the epithelial tip.
Loop process mediated through FGF-activated transcription factor genes Etv4 and Etv5.[23]
- Links: respiratory
Limb
FGF soaked beads (FGF-1, FGF-2 and FGF-4) are capable of inducing additional limbs in chicken embryos.[24] A later study[25] identified the endogenous signal as Fgf-8 from initially the intermediate mesoderm and then the prelimb field ectoderm for limb initiation and outgrowth, respectively.
- Links: limb] | chicken
Hearing
- fibroblast growth factor 1 - (Fgf-1) a growth factor released from cochlea sensory epithelium which stimulates spiral ganglion neurite branching.
- fibroblast growth factor 8 - (Fgf-8) a growth factor released by inner hair cells which regulates pillar cell number, position and rate of development.
- fibroblast growth factor receptor 3 - (Fgfr-3) a tyrosine kinase receptor with a role in the commitment, differentiation and position of pillar cells in the organ of corti
- Links: hearing
Palate
Fibroblast growth factors are required during palatogenesis and are candidate genes for syndromic and nonsyndromic cleft lip and cleft palate, see the recent review.[9]
FGF7, FGF8, FGF9, FGF10, FGF18 and their receptors FGFR1, FGFR2
- Links: palate | cleft palate
Abnormalities
- FGFR1 mutation has been associated with the relatively milder form of Pfeiffer syndrome type 1.
- FGFR2 and FGFR3 have been associated with the Apert, Crouzon and Pfeiffer syndromes.
FGF10
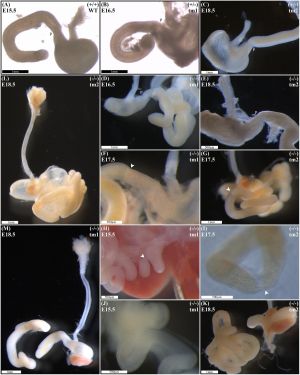
A recent mouse knock-out study has shown a single-gene deletion of Fgf10 can generate duodenal atresia in this animal model.[26]
Duodenal Atresia (Mouse Model)[26]
|
(A) Normal gastric, pyloric, and duodenal morphology was demonstrated by wild type embryos.
Null Fgf10 embryos universally demonstrated microgastria, but duodenal morphology varied according to presence and type of DA. Null embryos provided examples of: normal continuity and morphology of the duodenum for tm1 (D) and tm2 (E); type 1 DA, tm1 (F) and tm2 (G); type 2 DA, tm1 (H) and tm2 (I); and type 3 DA, tm1 (J) and tm2 (K).
(L) Type 2 DA demonstrating a “double bubble” with significantly dilated proximal duodenum; tm2.
(M) Type 3 DA demonstrating an intact esophagus, in the presence of tracheal atresia; tm1. Annotations denote genotype, scale bars, and location of pylorus, P. Arrows indicate location atresia in type 1 DA and type 2 DA examples.
|
References
- ↑ Itoh N. (2010). Hormone-like (endocrine) Fgfs: their evolutionary history and roles in development, metabolism, and disease. Cell Tissue Res. , 342, 1-11. PMID: 20730630 DOI.
- ↑ 2.0 2.1 Fongang B & Kudlicki A. (2013). The precise timeline of transcriptional regulation reveals causation in mouse somitogenesis network. BMC Dev. Biol. , 13, 42. PMID: 24304493 DOI.
- ↑ Gospodarowicz D. (1974). Localisation of a fibroblast growth factor and its effect alone and with hydrocortisone on 3T3 cell growth. Nature , 249, 123-7. PMID: 4364816 DOI.
- ↑ Gospodarowicz D & Moran JS. (1975). Mitogenic effect of fibroblast growth factor on early passage cultures of human and murine fibroblasts. J. Cell Biol. , 66, 451-7. PMID: 1170180 DOI.
- ↑ Bird AD, Croft BM, Harada M, Tang L, Zhao L, Ming Z, Bagheri-Fam S, Koopman P, Wang Z, Akita K & Harley VR. (2020). Ovotesticular disorders of sex development (DSD) in FGF9 mouse models of human synostosis syndromes. Hum. Mol. Genet. , , . PMID: 32452519 DOI.
- ↑ Kantarci H, Gou Y & Riley BB. (2020). The Warburg Effect and lactate signaling augment Fgf-MAPK to promote sensory-neural development in the otic vesicle. Elife , 9, . PMID: 32338604 DOI.
- ↑ Wong KL, Akiyama R, Bessho Y & Matsui T. (2018). ERK Activity Dynamics during Zebrafish Embryonic Development. Int J Mol Sci , 20, . PMID: 30597912 DOI.
- ↑ Row RH, Pegg A, Kinney B, Farr GH, Maves L, Lowell S, Wilson V & Martin BL. (2018). BMP and FGF signaling interact to pattern mesoderm by controlling basic helix-loop-helix transcription factor activity. Elife , 7, . PMID: 29877796 DOI.
- ↑ 9.0 9.1 Weng M, Chen Z, Xiao Q, Li R & Chen Z. (2018). A review of FGF signaling in palate development. Biomed. Pharmacother. , 103, 240-247. PMID: 29655165 DOI.
- ↑ Chan WK, Price DJ & Pratt T. (2017). FGF8 morphogen gradients are differentially regulated by heparan sulphotransferases Hs2st and Hs6st1 in the developing brain. Biol Open , 6, 1933-1942. PMID: 29158323 DOI.
- ↑ Diez Del Corral R & Morales AV. (2017). The Multiple Roles of FGF Signaling in the Developing Spinal Cord. Front Cell Dev Biol , 5, 58. PMID: 28626748 DOI.
- ↑ Atsuta Y & Takahashi Y. (2015). FGF8 coordinates tissue elongation and cell epithelialization during early kidney tubulogenesis. Development , 142, 2329-37. PMID: 26130757 DOI.
- ↑ Gredler ML, Seifert AW & Cohn MJ. (2015). Tissue-specific roles of Fgfr2 in development of the external genitalia. Development , 142, 2203-12. PMID: 26081573 DOI.
- ↑ Shifley ET, Kenny AP, Rankin SA & Zorn AM. (2012). Prolonged FGF signaling is necessary for lung and liver induction in Xenopus. BMC Dev. Biol. , 12, 27. PMID: 22988910 DOI.
- ↑ Lahti L, Saarimäki-Vire J, Rita H & Partanen J. (2011). FGF signaling gradient maintains symmetrical proliferative divisions of midbrain neuronal progenitors. Dev. Biol. , 349, 270-82. PMID: 21074523 DOI.
- ↑ Yu SR, Burkhardt M, Nowak M, Ries J, Petrásek Z, Scholpp S, Schwille P & Brand M. (2009). Fgf8 morphogen gradient forms by a source-sink mechanism with freely diffusing molecules. Nature , 461, 533-6. PMID: 19741606 DOI.
- ↑ Zhang XT, Wang G, Li Y, Chuai M, Lee KKH & Yang X. (2018). Role of FGF signalling in neural crest cell migration during early chick embryo development. Zygote , , 1-8. PMID: 30520400 DOI.
- ↑ Gray de Cristoforis A, Ferrari F, Clotman F & Vogel T. (2020). Differentiation and localization of interneurons in the developing spinal cord depends on DOT1L expression. Mol Brain , 13, 85. PMID: 32471461 DOI.
- ↑ Bayha E, Jørgensen MC, Serup P & Grapin-Botton A. (2009). Retinoic acid signaling organizes endodermal organ specification along the entire antero-posterior axis. PLoS ONE , 4, e5845. PMID: 19516907 DOI.
- ↑ Hayashi S, Shimoda T, Nakajima M, Tsukada Y, Sakumura Y, Dale JK, Maroto M, Kohno K, Matsui T & Bessho Y. (2009). Sprouty4, an FGF inhibitor, displays cyclic gene expression under the control of the notch segmentation clock in the mouse PSM. PLoS ONE , 4, e5603. PMID: 19440349 DOI.
- ↑ Govindarajan V & Overbeek PA. (2006). FGF9 can induce endochondral ossification in cranial mesenchyme. BMC Dev. Biol. , 6, 7. PMID: 16504022 DOI.
- ↑ Cardoso WV & Kotton DN. (2008). Specification and patterning of the respiratory system. , , . PMID: 20614584 DOI.
- ↑ Herriges JC, Verheyden JM, Zhang Z, Sui P, Zhang Y, Anderson MJ, Swing DA, Zhang Y, Lewandoski M & Sun X. (2015). FGF-Regulated ETV Transcription Factors Control FGF-SHH Feedback Loop in Lung Branching. Dev. Cell , 35, 322-32. PMID: 26555052 DOI.
- ↑ Cohn MJ, Izpisúa-Belmonte JC, Abud H, Heath JK & Tickle C. (1995). Fibroblast growth factors induce additional limb development from the flank of chick embryos. Cell , 80, 739-46. PMID: 7889567
- ↑ Vogel A, Rodriguez C & Izpisúa-Belmonte JC. (1996). Involvement of FGF-8 in initiation, outgrowth and patterning of the vertebrate limb. Development , 122, 1737-50. PMID: 8674413
- ↑ 26.0 26.1 Teague WJ, Jones MLM, Hawkey L, Smyth IM, Catubig A, King SK, Sarila G, Li R & Hutson JM. (2018). FGF10 and the Mystery of Duodenal Atresia in Humans. Front Genet , 9, 530. PMID: 30473704 DOI.
Reviews
Articles
Su N, Xu X, Li C, He Q, Zhao L, Li C, Chen S, Luo F, Yi L, Du X, Huang H, Deng C & Chen L. (2010). Generation of Fgfr3 conditional knockout mice. Int. J. Biol. Sci. , 6, 327-32. PMID: 20582225
Search Pubmed
Search Bookshelf Fibroblast Growth Factor
Search Pubmed Now: Fibroblast Growth Factor
Cite this page: Hill, M.A. (2026, July 16) Embryology Developmental Signals - Fibroblast Growth Factor. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Developmental_Signals_-_Fibroblast_Growth_Factor
- What Links Here?
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G






