Paper - Description of a Human Embryo of 13-14 Mesodermic Somites
| Embryology - 26 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Low A. Description of a human embryo of 13-14 mesodermic somites. (1908) J Anat Physiol. 42(3): 237-51. PMID 17232769 | PMC1289161
| Online Editor | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Modern Notes: | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Description of a Human Embryo of 13-14 Mesodermic Somites
Alexander Low
Senior Assistant to the Professor of Anatomy and Lecturer on Embryology, University of Aberdeen. (From the Anatomy Department, University of Aberdeen.)
- 13-14 Somite Paper: Plate 1 | Plate 2 | Plate 3 | Fig 1 | Fig 2 | Fig 3 | Fig 4 | Fig 5 | Fig 6 | Fig 7 | Fig 8 | Fig 9 | Fig 10 | Fig 11 | Fig 12 | Fig 13 | Fig 14 | Fig 15
Introduction
THE human embryo Pfannenstiel- III. was placed at the disposal of Professor Keibel, for publication in his Normentafel zur Entwickelungsgeschichte des Menschen, by Professor Pfannenstiel of Griefswald. In the Normentafel the embryo is No. 6 (fig. Vr. and Vv.) Professor Keibel kindly afforded me the opportunity of studying this embryo in the Anatomical Institute in Freiburg, and at his suggestion I undertook to reconstruct models of the embryo according to Born's method. The work of modelling was begun in the Anatomical Institute in Freiburg, and completed in the Anatomy Department, Aberdeen University.
The embryo Pfannenstiel III. was obtained at an operation, and as measured through the amnion had a length of 2.6 mm. The embryo was fixed in formalin-Muller's fluid, stained in paracarmine, and sectioned at a thickness of 10µm. Histologically the embryo is in an excellent state of preservation. A model of the whole embryo has been reconstructed at an enlargement of 100. A separate model of the nervous and alimentary systems has also been made, and three models of pharynx, heart with its blood-vessels, and septum transversum at an enlargement of 150.
External Form
(Plates I. and II.) The embryo is somewhat similar to the embryo von Bulle of Kollmann[1], which shows fourteen mesodermic somites and measured 25 mm. in length, but is not so far developed; indeed, its outer form more nearly approaches that of the embryo of eight paired somites and a length of 2.15 mm. described and modelled by Professor Eternod.[2] Like the Eternod embryo, it is spirally twisted in its long axis, and shows the same three back curvatures -a convex neck bend, a concave dorsal, and a convex rump. The dorsal flexure is slight, while the neck bend makes a right angle with the body, approaching in this respect the form of the embryo of twenty-three somites and a length of 2.15 mm. described by Professor Peter Thompson[3]. The medullary plate is still open at its head and caudal ends. The front of the head is directed down and slightly to the right over the bulging heart region,and thus is formed a well-developed stomodaeum. Three visceral clefts are visible-the first and second being deep, the third a shallow groove. The first and second visceral clefts lie dorsal to the rounded swelling caused by the commencing mandibular arch. The first cleft occupies a position more to the cranial end of the embryo. The second cleft is separated by a convex ridge from the first, and its caudal end extends to the convexity of the body wall caused by the heart. The third visceral cleft is caudal and somewhat more dorsal, and separated by a considerable interval from the other two clefts. Its ventral end leads into a depression of the body wall behind the pericardial region. The ear pit is represented by an oval depressed area of thickened epithelium just cranial to the dorsal end of the third visceral cleft. There is a large elongated umbilical opening through which the embryonic and extra-embryonic coelom are in communication. The yolk sac is constricted at its communication with the primitive intestine to form a distinct neck. The body stalk arises from the ventral aspect of the embryo and passes to the right, lying first in front and then to the right of the caudal end of the embryo, the tail being directed to the left. The body stalk is separated from the neck of the yolk sac by a narrow interval. The caudal end of the embryo is swollen out, and terminates in a blunt point; a trace of the primitive streak is prolonged from the medullary groove to the cloacal membrane. Along the line of attachment of the amnion to the lateral body wall on each side there is a ridge caused by the underlying umbilical vein; this ridge bounds the umbilical orifice laterally. The amnion extends on to the dorsal aspect of the body stalk, leaving its lateral and ventral aspects uncovered-a disposition similar to that in the embryo described by Professor Thompson.
Mesodermic Somites and Notochord
There are thirteen pairs of well-formed mesodermic somites, and in addition to these there is an anterior pair incompletely formed. Thismost anterior and rudimentary pair of somites is situated just at the level of the neck bend. The last pair of somites are well formed, and situated at the level of the commencement of the rump bend. As the ganglia are not developed, it is not possible to differentiate the regions to which the somites belong. Each somite has a uniformly thick wall three or four cells deep, enclosing a distinct cavity (myocoel). Towards the cavity many of the cells are undergoing mitotic division. The more cranial-placed somites are triangular on cross-section, while the more posterior ones are quadrangular (fig.1). The notochord extends from a point behind the buccopharyngeal membrane, to end in the caudal region just short of the hind-gut and between the cloaca and medullary plate, and not extending into the tail.

Fig. 1. - Right half of a transverse section of the embryo through the region of the eleventh mesodermic somite. Ec., ectoderm; XI.M.S., mesodermic somite; St., segmental tubule; Coe., coelom; d.Ao., dorsal aorta; Sp.c., medullary canal; En.,entoderm; Ch.,notochord.
The chorda lies in close relation with entoderm forming the roof of the intestine, and only toward its caudal termination is it quite separated of, and here lies in the mesoderm midway between the gut and the medullary plate. At its cranial commencement the chorda is at first not separated of, but appears more as a heaping up of entoderm cels in the roof of the pharynx; soon, however, it becomes more differentiated, as represented in fig.2. At the level of the first pair of mesodermic somites the chorda appears more as an evagination of the entoderm of the root of the gut, and this condition obtains almost to its caudal end, where it gets quite separated off from entoderm (fig.3). The cells of the chorda are large, oval, and clear, and contain small, distinctly staining nuclei; in some of the cells mitosis is seen. There is no trace of a cuticular membrane.

Fig. 2 and 3.
Vascular System



Cardinal Veins
The anterior cardinal veins, containing blood-cells, are present in the region of the first to the fifth mesodermic somites. A connection with the heart cannot be established from a careful examination of the sections. No trace of posterior cardinal veins is to be seen.
Vitelline Veins
Although inthe wall of the yolk sac are numerous blood-vessels containing blood cells, still the vitelline veins are difficult to trace. Their terminal parts can be defined where they lie in the septum transversum on the ventral aspect of the gut, and open into the sinus venous medial to the umbilical veins. Before they open into the sinus venosus, a transverse piece connects the two vitelline veins.
Umbilical Veins
The umbilical veins are large vessels, and lie in the body stalk at first dorsal to the umbilical arteries; here they communicate, and then run cranialwards in the body stalk to gain the lateral body wall. From the lateral body wall the cranial end of each vein bends abruptly, passing medially and ventrally each to join its own horn of the sinus venosus; here each vein lies embedded in the septum transversum and close to the floor of the parietal recess(fig.12).
Aortae
The aortic stem divides into a pair of aortic arch vessels, which pass on either side of the median thyroid anlage backwards in the first visceral arch, and each of these is continued caudally as a dorsal aorta. There are traces also of a second pair of aortic arch vessels. The dorsal aortas rest on the roof of the gut on either side of the notochord, and do not unite in any part of their course, remaining paired throughout and terminating on the sides of the cloaca. A short distance before the termination of the dorsal aortae the umbilical arteries are given off and pass on either side of the allantoic duct into the body stalk, where they unite, forming a single trunk, which again divides into two vessels.
The Heart
The heart is in a stage very similar to that represented by Born's model[4] of the heart of a rabbit embryo of 095 mm. length - model 1 (Ziegler). The heart as a whole is S-shaped, and shows sinus venosus, auricle, ventricle, and just at the base of the aortic stem a slight dilatation which may represent a bulbus cords.
The sinus venosus is crescentic in shape, with dilated ends, and is situated in the septum transversum in close relation with the ventral wall of the gut. Opening into its horns are the terminations of the large umbilical veins, and medial to these the vitelline veins. The sinus passes into the auricle without any very distinct constriction except on the left side, where there is a slight furrow between the two. The auricle lies dorsal to the ventricle. Its transverse diameter is the greater, and it is divisible into a smaller right part and a larger left by a distinct in-pushing- the dorsal mesentery being attached here. The left part of the auricle is separated from the ventricle by a slight groove.
The ventricle is U-shaped, with the left limb much the larger and passing dorsally and to the left to open into the left side of the auricle. The cranial end of the right limb becomes constricted, and then is succeeded by a somewhat dilated part, which may indicate the bulbus cordis which is so largely developed in the embryo described by Professor Thompson.
Alimentary System




The mouth is represented as a narrow transverse fissure between the forebrain and the body wall. In its roof is a slight funnel-shaped out-pushing - the commencement of the pocket of ectoderm to form the hypophysis. The mouth is separated from the pharynx by a complete buccopharyngeal membrane. The pharynx widens out rapidly behind this, and is flattened dorso-ventrally (fig.7).
There are four pairs of pharyngeal pouches, of which the third and fourth are small and pointed. The second pocket is so situated that it lies much more ventral than caudal to the first pocket. The first two pairs of pockets are elongated vertically, and come in direct contact with the external ectoderm, where it forms the bottoms of the first and second pairs of visceral clefts (fig.7). The third and fourth pockets do not come in contact with the ectoderm. In the floor of the pharynx in the region of the second pockets is present the median thyroid anlage as a relatively large, widely open, funnel-shaped cavity with its apex passing caudally between the aortic arches (fig.8). The lung anlage is not plainly indicated, but caudal to the median thyroid is a groove in the entoderm which may indicate the lung groove. As the fore-gut is traced towards the yolk sac, its lumen becomes narrowed transversely and elongated dorso-ventrally; and just before it opens out into the yolk sac, there is a liver bay with walls of thickened entoderm. There is no actual liver bud and no trace of liver trabeculae. The middle third of the alimentary tract is still in communication with the yolk sac. Thehind gut is narrowed at a point opposite the attachment of the body stalk, and then dilates to form a large cloaca. Dorsally the cloaca is expanded with convex walls, while ventrally it is narrowed so that its cavity forms a fissure.
Ventrally the entoderm of this anterior fissure-like part of cloaca is in direct contact with the ectoderm, forming an elongated cloacal membrane. Cranial to the cloacal membrane the cavity of the cloaca leads into the small lumen of the allantois. The allantoic duct passes into the body stalk between the two umbilical arteries, which eventually surround it by anastomosing across it. There is a very small post-anal gut projecting caudally between the posterior limit of the cloacal membrane and the tip of the notochord.
Excretory System
The excretory system is in a very early stage of development. Professor Felix of Zurich is to describe this in detail in a separate paper on the development of the excretory system in early human embryos. I may say that this eml)ryo shows rudimentary pronephric tubules in the region of the eighth, ninth, and tenth mesodermnic somnites, and segmental vesicles in connection with the eleventh, twelfth and thirteenth segments. There is no Wolffian duct.
Coelom and Septum Transversum
All parts of the coelom are in communication with each other. The pericardial coelom is very large, and extends cranially on either side of the median thyroid anlage. Ventrally the pericardial coelom causes a bulging of the body wall, the ventricle lying in close apposition with it; while dorsally it is prolonged round into the parietal recesses of His[5].
Figs. 11 and 12, fromn a model, show the relation of the umbilical vein and sinus venosus to the floor of the parietal recess, as also the two horn-like extensions of the septum transversum described by Professor Thompson[6]. On the right side there is an infolding of the body wall which produces a ridge running from the anterior part of the dorsal mesocardium to the septum transversum, and slightly constricting off the pericardial coelom from the parietal recess; this may be the "pulmonary ridge" of Mall[7]. Only the cranial end-of this ridge is present on the left side.


Fig. 11 Model showing septum transversum viewed from above.
Fig. 12 Model showing right half of septum removed and the right umbilical vein passing in to open into the sinus venous.
- Sp.c., medullary canal* dAo. dorsal aorta; P.R., parietal recess; Am., amnion; S.F., sinus venos s; S.74., septum transversum; Y.S., yolk sac; r.u.v., right
vitelline vein; F.G., fore-gut; I l., third somite.
Nervous System



The medullary plate is open at its head and tail ends almost to the same extent as in Eternod's embryo of eight mesodermic somites. In Kollmann's embryo of fourteen somites the medullary groove is closed behind, but open anteriorly. The brain shows very distinctly the three parts, prosencephalon, mesencephalon, and rhombencephalon, separated by grooves (Plate I.). There is no demarcation between the rhombencephalon and the spinal cord, but the junction seems to be at the neck bend at the level of the first pair of mesodermic somites. Just cranial to this point the two folds of the medullary groove have fused. The fore-brain shows elongated, gutter-like evaginations of its later walls which are the commencing optic vesicles. The walls of these vesicles are many cells thick, and are in close contact with the external ectoderm (fig.13). From the floor of the fore-brain is a slight out-pushing, the anlage of the infundibulum. The mid-brain is bounded in front and behind by grooves, and its transverse diameter is rather less than that of either fore-brain or hind-brain. The hind-brain forms by far the largest part of the whole brain, and passes without distinct demarcation into the spinal part of the medullary tube. The hind-brain is of interest in that it shows certain neuromeres very clearly; these neuromeres form folds involving the whole thickness of the lateral wall of the hind-brain, and are convex externally, with corresponding concavities internally (fig.14). The neuromeres are seven in number, as has already been noted by Broman[8] in a human embryo of 3 mm., and more recently by Thompson in the twenty-three somite embryo and Ingalls in an embryo of 4-9mm.[9]. The first neuromere is the smallest, and does not reach the ventral margin of the brain, while the second neuromere is the largest and best defined. The third neuromere is also prominent, but not so wide as the second, and its ventral end tapers off to a point between the ventral ends of the second and fourth neuromeres. The fourth neuromere is pointed dorsally and widens out below, between its ventral end and that of the second nenromere being a notch as described by Bremer[10] in a 4mm. embryo. The ganglion acustico-facialis is attached to the upper end of this neuromere, but largely lies in the furrow between the third and fourth neuromeres. The fifth, sixth, and seventh are not so clearly marked off.
The oval auditory field of thickened ectodermn lies over the dorsal end of the fifth neuromere, but also extends over the upper end of the fourth. The trigeminal ganglion is not developed. The ganglion acustico-facialisis large, and is attached to the lateral wall of the hind-brain in connection with the anterior margin of the fourth neuromere. The ganglion of the vagus is only being developed.
The flexures of the brain agree very closely with the flexures described by Professor Thompson as being present in his embryo of twenty-three somlites. There is a prominent flexure in the region between the mid-brain and hind-brain, the mid-brain and fore-brain being bent downward. The anterior extremity of the notochord is bent down in a corresponding manner. There is also a distinct flexure of the hind-brain at the notch between the ventral ends of the second and fourth neuromeres. rreneck flexure is just commencing, and forms an open angle (Plate I.).
In conclusion, I wish to express my thanks to Professor Wiedersheini for kindly granting me permission to work in his laboratory; more especially to Professor Keibel for the loan of the embryo, and much help and advice; and also to Professor Pfannenstiel to whom the embryo belongs. I also express my indebtedness to Professor Reid in connection with mly research work, and to the Carnegie Trustees for a grant toward the cost of embryological research and the illustration of this paper.
Explanation of Plates
- 13-14 Somite Paper: Plate 1 | Plate 2 | Plate 3 | Fig 1 | Fig 2 | Fig 3 | Fig 4 | Fig 5 | Fig 6 | Fig 7 | Fig 8 | Fig 9 | Fig 10 | Fig 11 | Fig 12 | Fig 13 | Fig 14 | Fig 15
Plate I

Model of the embryo, left side (enlarged 60 times).
Plate II

Model of the embryo. frontal view (enlarged 60 times).
Plate III

The same model in sagittal section, viewed from the left.
Al., allantoic duct; Am., anmnion; Ao., origin of first aortic arch; B.S., body stalk ; Ch., chorda dorsalis ; Chr., choriol ; Cl., cloaca; Cae., ewlom ; D.M., dorsal miesentery; F.B., fire-braii; F.G., fore-gut; G.af., ganglion acustico-facialis; H.G., biind-gut; Hy., hypophysis; AM., mouth ; M.B., mid-brain; M.G., mid-gut; Op.,Optic vesicle; Ot., otic pit; P.C., pericardial coelom; S.T., septum itransversum S.V., sinus venosus; Th., median thyroid ; L.U.A., left umbilical artery; L.U.V., left umbilical vein; R.U.V. right umbilical vein; Y.S., yolk sac; I. and III., first and third visceral clefts; I.-VII., first to seventh neuromeres.
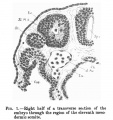
Fig. 1.

Fig. 2 and 3.
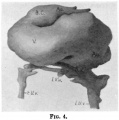
Fig. 4.
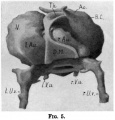
Fig. 5.
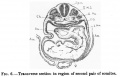
Fig. 6.
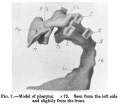
Fig. 7.
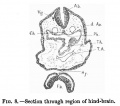
Fig. 8.
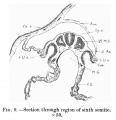
Fig. 9.
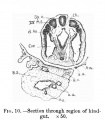
Fig. 10.
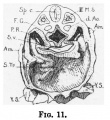
Fig. 11.
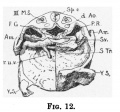
Fig. 12.
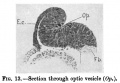
Fig. 13.
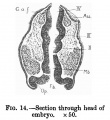
Fig. 14.
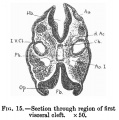
Fig. 15.
References
- ↑ KOLLMANN, J., " Die Ktrperform menschlicher normaler und pathologiselmer Embryonen," Arch.f. Anat. u.Phys., Suplemi.Bd., 1889.
- ↑ ETERNOD, A., "Surun ceuf humainde 1-3mm. avereimbryonde, 21mm.," Actes de la Societe helcetique des Sciences naturelles, Zurich, 1896.
- ↑ Thompson P. Description of a human embryo of twenty-three paired somites. (1907) J Anat Physiol, 41(3):159-71. PMID 17232726
- ↑ BORN, G., " Beitrige zur Entwicklungsgeschichte des Saugethierherzens," Arch.f.nikr. Anat., Bd. xi., 1889.
- ↑ His W. Anatomie Menschliche Embryonen I - Embryonen des ersten monats (Anatomy of human embryos - Embryos of the first month). (1880) Leipzig.
- ↑ Thompson P. A note on the development of the septum transversum and the liver. (1908) J Anat Physiol. 42(2): 170-5. PMID 17232762
- ↑ Mall FP. On the development of the human diaphragm. (1901) Johns Hopkins Hospital Bulletin 12: 158-171.
- ↑ BROMAN, J., "Beschreibung eines menschlichen Embryos von beinahe 3 mm. Lange, mit spezieller Bemerkung uber die bei demselben befindfichen Hirnfalten," Morph. Arbeiten, Bd. v., 1896.
- ↑ Ingalls NW. Description of a human embryo of 4.9 mm (Beschreibung eines menschlichen Embryos von 4.9 mm). (1907) Arch. f. mik. Anat., 70: 506-576.
- ↑ Bremer JL. Description of a 4-mm human embryo. (1906) Amer. J Anat. 5: 459-480.
| Historic Disclaimer - information about historic embryology pages |
|---|
|
- Carnegie Stages: 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | About Stages | Timeline
Cite this page: Hill, M.A. (2024, April 26) Embryology Paper - Description of a Human Embryo of 13-14 Mesodermic Somites. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_Description_of_a_Human_Embryo_of_13-14_Mesodermic_Somites
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G