Developmental Mechanisms
| Embryology - 25 Jul 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Introduction
These notes are intended to introduce a number of different mechanisms at the all levels that form structures within the embryo. These are general notes about these mechanisms giving only a few examples. For specific tissues and organs read the system notes. What is remarkable, given our biological diversity, is the strong evolutionary conservation of these developmental mechanisms. Researchers should still recognise that there still exist significant differences between "animal models" and human development.
| Factor Links: AMH | hCG | BMP | sonic hedgehog | bHLH | HOX | FGF | FOX | Hippo | LIM | Nanog | NGF | Nodal | Notch | PAX | retinoic acid | SIX | Slit2/Robo1 | SOX | TBX | TGF-beta | VEGF | WNT | Category:Molecular |
Some Recent Findings

|
| More recent papers |
|---|
 This table allows an automated computer search of the external PubMed database using the listed "Search term" text link.
More? References | Discussion Page | Journal Searches | 2019 References | 2020 References Search term: Developmental Mechanisms | Molecular Development | Axis Formation |
| Older papers |
|---|
| These papers originally appeared in the Some Recent Findings table, but as that list grew in length have now been shuffled down to this collapsible table.
See also the Discussion Page for other references listed by year and References on this current page.
|
Apoptosis
| This single term "apoptosis" describes the way in which the majority of cells die within our adult body are removed every day, "Programmed Cell Death". In development, apoptosis begins in the early blastocyst and is a developmental mechanism found throughout tissues in the embryo and fetus developmental stages. In addition to the many developmental roles this process is used in multicellular organisms to remove cells that are: aged, superfluous, infected, contain genetic errors or are transformed. | 
Neuron apoptosis or necrosis (EM).[6] |
Axes

All embryos occupy 3 D space and appear differently externally and internally within this space. This gives us 3 different axes which need to be determined: Head/Tail (rostro/caudal), Front/Back (anterior/posterior), and Left/Right. The left/right axis appears at first mirror symetrical, and looks very similar externally, but clearly many internal organs are not duplicated (heart, liver, brain etc).
Left/Right
The left/right axis appears at first mirror symetrical, and looks very similar externally, but clearly many internal organs are not duplicated (heart, liver, brain etc).

Embryo left-right asymmetry pathway[7] |
|
Cellular Mechanisms
These notes are intended to introduce a number of mechanisms at the cellular level that form structures within the embryo. These cellular dynamic and physical interactions form basic tissue structures of epithelia, connective tissues and features such as tubes.
Cell Migration
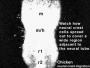
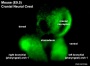
Neural Crest Migration
|
|
|

|
|

|
|
|
| Cranial Migration |
Germ Cell Migration
| Migration 1 | Migration 2 | Migration 3 |
Adhesion
- Adhesion EM Images: GIT epithelia EM1 | GIT epithelia EM2 | GIT epithelia EM3 | Desmosome EM
- Adhesion Cartoons: Tight junction | Adherens Junction | Desmosome | Gap Junction
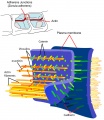
Adhesion Cartoons

Tight junction
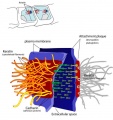
Adherens Junction
Desmosome
Gap Junction




Adhesion EM Images
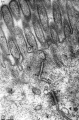
GIT epithelia EM1
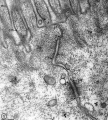
GIT epithelia EM2
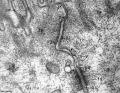
GIT epithelia EM3
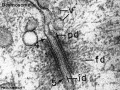
Desmosome EM




Differential positioning of adherens junctions is associated with initiation of epithelial folding
Nature. 2012 Mar 28. doi: 10.1038/nature10938.
Wang YC, Khan Z, Kaschube M, Wieschaus EF. Source 1] Department of Molecular Biology, Princeton University, Princeton, New Jersey 08544, USA [2] Howard Hughes Medical Institute, Princeton University, Princeton, New Jersey 08544, USA.
Abstract
- "During tissue morphogenesis, simple epithelial sheets undergo folding to form complex structures. The prevailing model underlying epithelial folding involves cell shape changes driven by myosin-dependent apical constriction. Here we describe an alternative mechanism that requires differential positioning of adherens junctions controlled by modulation of epithelial apical-basal polarity. Using live embryo imaging, we show that before the initiation of dorsal transverse folds during Drosophila gastrulation, adherens junctions shift basally in the initiating cells, but maintain their original subapical positioning in the neighbouring cells. Junctional positioning in the dorsal epithelium depends on the polarity proteins Bazooka and Par-1. In particular, the basal shift that occurs in the initiating cells is associated with a progressive decrease in Par-1 levels. We show that uniform reduction of the activity of Bazooka or Par-1 results in uniform apical or lateral positioning of junctions and in each case dorsal fold initiation is abolished. In addition, an increase in the Bazooka/Par-1 ratio causes formation of ectopic dorsal folds. The basal shift of junctions not only alters the apical shape of the initiating cells, but also forces the lateral membrane of the adjacent cells to bend towards the initiating cells, thereby facilitating tissue deformation. Our data thus establish a direct link between modification of epithelial polarity and initiation of epithelial folding."
Wang YC, Khan Z, Kaschube M & Wieschaus EF. (2012). Differential positioning of adherens junctions is associated with initiation of epithelial folding. Nature , 484, 390-3. PMID: 22456706 DOI.
Role of the gut endoderm in relaying left-right patterning in mice
PLoS Biol. 2012 Mar;10(3):e1001276. Epub 2012 Mar 6. Viotti M, Niu L, Shi SH, Hadjantonakis AK. Source Developmental Biology Program, Sloan-Kettering Institute, New York, New York, United States of America.
Abstract
- "Establishment of left-right (LR) asymmetry occurs after gastrulation commences and utilizes a conserved cascade of events. In the mouse, LR symmetry is broken at a midline structure, the node, and involves signal relay to the lateral plate, where it results in asymmetric organ morphogenesis. How information transmits from the node to the distantly situated lateral plate remains unclear. Noting that embryos lacking Sox17 exhibit defects in both gut endoderm formation and LR patterning, we investigated a potential connection between these two processes. We observed an endoderm-specific absence of the critical gap junction component, Connexin43 (Cx43), in Sox17 mutants. Iontophoretic dye injection experiments revealed planar gap junction coupling across the gut endoderm in wild-type but not Sox17 mutant embryos. They also revealed uncoupling of left and right sides of the gut endoderm in an isolated domain of gap junction intercellular communication at the midline, which in principle could function as a barrier to communication between the left and right sides of the embryo. The role for gap junction communication in LR patterning was confirmed by pharmacological inhibition, which molecularly recapitulated the mutant phenotype. Collectively, our data demonstrate that Cx43-mediated communication across gap junctions within the gut endoderm serves as a mechanism for information relay between node and lateral plate in a process that is critical for the establishment of LR asymmetry in mice."
Viotti M, Niu L, Shi SH & Hadjantonakis AK. (2012). Role of the gut endoderm in relaying left-right patterning in mice. PLoS Biol. , 10, e1001276. PMID: 22412348 DOI.
Molecular Mechanisms
This page is a link to many different resources related to Molecular Development. In current years we have turned from wanting to merely describe the events of embryogenesis, to a desire to understand the mechanisms of development. This has been a boon in allowing the use of many (easier) model systems such as the genetist's tool the fruitfly, and the worm, frog, chicken, zebrafish and mouse (see other embryos page).
A continuing theme also seems to be the reuse of signals at different times and places within the embryo, for diiferent jobs. This has given rise to the concept of "switches" which by themselves may contain no "information" but to activate other genes or switches. Finally, you can imagine that of our 20,000-25,000 protein-coding genes, a large number of these may only be expressed during development or if reused, have a completely different role in the mature animal.
Sex Determination
Eukaryotic organisms reproduce by generating haploid gamete cells, oocytes and spermatozoa, that in fertilization recombine to form the first diploid cell of the new organism. In normal human development, each gamete contains a single sex chromosome, the oocyte carries an X chromosome and the spermatozoa carries either an X or a Y chromosome. Initial sex determination is therefore determined by the spermatozoa and the specific sex chromosome it contains.
In development, the term sex determination used to apply more broadly to how the Y chromosome determined male development. It was thought that female sex determination was the "default mechanism", but we now know that there are also female specific mechanisms.
- Links: genital
Epigenetics
Epigenetics, as the name implies, is the inheritance mechanisms that lie outside the DNA sequence of our genome and genes. These molecular mechanisms include: DNA methylation, histone modification, and those related to the microRNA machinery.
- Links: epigenetics
Model Differences
While many mechanisms of development are strongly conserved, researchers should still recognise that there still exist significant differences between "animal models" and human development.
A few examples:
- respiratory development in human and mouse[8]
- X inactivation - ZFX on the X chromosome in humans escapes X inactivation while in mouse it is inactivated.[9]
References
- ↑ Waldner C, Roose M & Ryffel GU. (2009). Red fluorescent Xenopus laevis: a new tool for grafting analysis. BMC Dev. Biol. , 9, 37. PMID: 19549299 DOI.
- ↑ Ebisuya M & Briscoe J. (2018). What does time mean in development?. Development , 145, . PMID: 29945985 DOI.
- ↑ Inomata H. (2017). Scaling of pattern formations and morphogen gradients. Dev. Growth Differ. , 59, 41-51. PMID: 28097650 DOI.
- ↑ Nance J. (2014). Getting to know your neighbor: cell polarization in early embryos. J. Cell Biol. , 206, 823-32. PMID: 25267293 DOI.
- ↑ Harding MJ, McGraw HF & Nechiporuk A. (2014). The roles and regulation of multicellular rosette structures during morphogenesis. Development , 141, 2549-58. PMID: 24961796 DOI.
- ↑ Ueda H, Fujita R, Yoshida A, Matsunaga H & Ueda M. (2007). Identification of prothymosin-alpha1, the necrosis-apoptosis switch molecule in cortical neuronal cultures. J. Cell Biol. , 176, 853-62. PMID: 17353361 DOI.
- ↑ Norris DP. (2012). Cilia, calcium and the basis of left-right asymmetry. BMC Biol. , 10, 102. PMID: 23256866 DOI.
- ↑ Nikolić MZ, Sun D & Rawlins EL. (2018). Human lung development: recent progress and new challenges. Development , 145, . PMID: 30111617 DOI.
- ↑ Adler DA, Bressler SL, Chapman VM, Page DC & Disteche CM. (1991). Inactivation of the Zfx gene on the mouse X chromosome. Proc. Natl. Acad. Sci. U.S.A. , 88, 4592-5. PMID: 2052543
Journals
Reviews
Shilo BZ & Barkai N. (2017). Buffering Global Variability of Morphogen Gradients. Dev. Cell , 40, 429-438. PMID: 28292422 DOI.
Amourda C & Saunders TE. (2017). Gene expression boundary scaling and organ size regulation in the Drosophila embryo. Dev. Growth Differ. , 59, 21-32. PMID: 28093727 DOI.
Bryant SV & Gardiner DM. (2016). The relationship between growth and pattern formation. Regeneration (Oxf) , 3, 103-22. PMID: 27499882 DOI.
Search PubMed
Search PubMed: Developmental Mechanisms
External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2026, July 25) Embryology Developmental Mechanisms. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Developmental_Mechanisms
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G