Paper - A study of a 7 mm human embryo with special reference to its peculiar spirally twisted form, and its large aortic cell-clusters
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Jordan HE. A study of a 7 mm human embryo; with special reference to its peculiar spirally twisted form, and its large aortic cell-clusters (1918) Anat. Rec. 14(7): 479–492.
| Online Editor | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| This historic 1918 paper by Jordan describes a human embryo that on external appearance of this embryo suggests it is a Carnegie stage 14 occurs in week 5, 31 - 35 days, 5 - 7 mm.
|
| Historic Disclaimer - information about historic embryology pages |
|---|
 |

A study of a 7 mm human embryo with special reference to its peculiar spirally twisted form, and its large aortic cell-clusters
H. E. Jordan
Department of Anatomy, University of Virginia
This embryo, obtained after hysterectomy, was very kindly sent to me, with the chorionic vesicle intact, by one of my former students, Dr. Joseph S. Hume of Norfolk, Virginia. It had been placed in a 10 per cent formalin solution about an hour after the operation. The embryo would seem to merit a brief separate description by reason of its extreme spiral form (fig. 1), and its unusually numerous and large aortic cell-clusters (fig. 7). The study of this embryo supplies important data for the interpretation of these two characteristics, here greatly accentuated, present in small and variable degree in practically all young human embryos of a certain stage of development. The fact that we possess the requisite data also for a fairly close computation of its age,‘ adds to the importance of this specimen.
Chorionic vesicle
The complete ovum had an oval shape measuring approximately 20 mm. by 12 mm. With the exception of a small, approximately circular, equatorial area, it was thickly covered with villi. There was no macroscopic evidence of anypathologic or abnormal conditions. Under microscopic examination the chorionic villi, the amnion, the mucous tissue of the umbilical cord, and the trophoblast islands appear perfectly normal. There is no round cell infiltration in the membranes, nor are any leukocytes present among the trophoblast cells.
Yolk-sac
Due to my special interest in the histology of the human yolk-sac, I was very anxious to obtain and study the sac of this specimen. On opening the chorionic vesicle the embryo was immediately noted, but the umbilical vesicle could not be discerned. This was the more surprising since this vesicle, attached to its stalk, is generally quite conspicuous in its usual position just forward of the head. On exploring the exocelom a considerable, though judging from Mall’s description,11 not abnormally excessive, reticular magma (‘magma réticulé’) composed of delicate fibers was encountered; but the yolk—sac could not be disclosed. I noticed, furthermore, that the embryo had suffered a complete spiral twist (figs. 2 and 3) in its post- cardiac portion, that the umbilical cord was exceptionally short, and that the amnion was closely adherent to the chorion in a narrow annular zone about the point of attachment of the body- stalk to the chorion. Since the complete series of sections of the embryo, including the placental pole of the cord, the adjacent portion of the fetal placenta, and the umbilical celom, reveal no trace of the yolk—sac or stalk, I conclude that the sac had become caught between the amnion and the chorion and had suffered dissolution. If this conclusion is correct, we are led to the further conclusion that, since the sections show no clear indication of abnormality (figs. 4 to 6), the yolk-sac represents no essential factor in determining normal ontogeny at least after the third week of development. This calculation assumes that the consummation of the disintegration of the sac, under the conditions obtaining for this embryo, required about ten days.
Age of the Embryo
The embryo lay on its right side (fig. 2) with the umbilical cord passing over the left side. The longest measurement was along the nape-breech line, 7 mm. After transferring the specimen to alcohol "during the paraflin embedding-process it shrunk to about 5 mm. Using the greatest length, 7 mm., for determining the age by length by comparison with standard series of embryos (His) Keibel und Elze,9 Mall,10 Triepel17) it falls somewhere between 26 and 34 days. Mall11 computed the age of a 7 mm. embryo (No. 208) at 26 days. Triepel17 ascribes an age of 34 days to the same embryo. Our embryo corresponds most closely with Embryo A (7.5 mm.) of His’s collection, which he estimated to be from 27 to 30 days old. It corresponds closely in external appearance also with Embryo 112 of Keibel’s collection (Normentafeln, Keibel und Elze, 9 fig. 9, plate 6) the greatest length of which is given as only 5.3 mm., though it has thirty-six somites and four branchial arches, and is described as being slightly less developed than His’s Embryo A.
The data pertaining to our embryo (Hume) are: Last menstrual period began August 15; uterus removed October 3. The greatest possible age, therefore, allowing seven days for a period probably unfavorable for fertilization, could be only 42 days. Computing the age according to the conventional formula of His, on a basis of 28 days for the menstrual cycle, we arrive at only 21 days. To this result, however, should be added at least 7 days to allow for the passage of the segmenting ovum through oviduct and the resulting inhibition of the omitted menstruation, giving 28 days. The latter result harmonizes more closely with the revised seriations of Keibel und Elze” and of Triepel,17 and we may regard this embryo as entering upon its fifth week of development. It is interesting to note that the interval between the lapsed menstruation and operation is the same (21 days) as the corresponding interval in the case of the aborted embryo of 7 mm. of Mall’s collection.
Gross Morphology
A very striking feature about the gross form of the embryo is its extreme spiral twist, the main element of which is post-cardiac, chiefly pelvic; but the entire embryo is to some extent involved. Reckoning from the tip of forebrain to tip of tail, the embryo makes one complete spiral turn. The only other embryos described or illustrated, as far as I am aware, as spirally twisted to a similar degree, are the Fischel embryo of Hochstetter’s Collection (fig. 44, Keibel and Mall’s Embryology‘°), and the Keibel embryo No. 112 (fig. 9, plate 6, Keibel und Elze’s Normentafeln°). However, neither of these embryos is twisted to the same extreme degree. Moreover, the Fischel embryo (4.02 mm.), which most closely resembles our embryo with respect to the spiral twist, except that it has a sharper nuchal flexure, thus bringing the forebrain over the hind limb-bud, is certainly somewhat younger. Judging from the number of branchial arches (the fourth is vaguely discernible), the condition of the maxillary process, the character of the rhombencephalon, the number of segments (approximately 38), and the stage of development of the limb-buds, our embryo is more nearly of the stage of development of the Keibel embryo No. 112 (5.3 mm.). However, a comparison of my sections with those published by Keibel und Elze shows a thicker and more condensed myocardium, and a more advanced stage in the development of the liver and the pancreas, which conditions bring the actual age of our embryo nearer that of the Embryo A of His’s collection.

Fig. 1. View of embryo from right side, attached to chorion. It is sharply curved about the neck region, so that the tip of the forcbrain touches the umbilicus. It is also twisted through a complete spiral, the mid-dorsal line coming into view caudal to the right fore limb-bud, and the left hind limb-bud showing below the pelvic flexure. The rhombencephalon, the eye, and the right vena capitis lateralis are also conspicuous; and certain of the nuchal and pelvic myotomes may be seen. Approximately thirty-eight somites could be counted. The chorionic villi are seen beyond the reflected edges of the opened vesicle. This is the View obtained when the embryo was turned on the short umbilical cord so as to expose the side opposite to the one shown when the embryo was in its more natural position as in figure 2. In this position it could only be maintained by force. When detached from the chorionic vesicle, by cutting from the wall of the vesicle a square piece of tissue including the area of attachment of the umbilical cord, the embryo assumed the position shown in figure 3, when the right cephalic surface was turned uppermost. Photo. X3. All photographs were made by Mr. F. L. Foster.

Fig. 2. Left side-view of embryo. The left ventricle and the bulbs cordis are conspicuous. The anlages of the cerebral hemispheres, the eye, the rhombencephalon, three branchial arches, the umbilical cord, the hind limb-buds and the tail are also clearly outlined. The photograph was retouched so as to bring into sharper relief also the fourth branchial arch, the left fore limb-bud and the otocyst with the short ductus endolymphaticus. Magnification X 8. The lines a—a’, b—b’, and c—c’ indicate approximately the levels of sections in figures 4 to 6.

Fig. 3. View of right cephalic surface. The mass to the left consists of amnion above, and fused amnion and chorion below. The right fore limb-bud has become pushed forward by the spiral twisting and turned so as to bring its originally ventral border on a line parallel with the ventral surface of the pharynx. Just above the branchial arches may be seen through the translucent tissue the vena capitis lateralis. Photo X 8.

Fig. 4. Section at approximately level a—a’ in figures 2 and 3, just below the point of bifurcation of the trachea. This is the point where the single-cell ‘angioblasts’ (hemoblasts) and the smaller cell-clusters (‘blood-islands’) begin to appear ventrally in both aortic roots. The section shows also the base of the right fore limb-bud, the right brachial plexus, the esophagus, the left ventricle, the bubus cordis, the inferior vena cava, the left duct of Cuvier, the liver with the ductus venosus dorsally, and the telencephalon with the right olfactory placode. The celom contains considerable blood at the right. Photo X 15. (In comparing the photograph of this section with fig. 2 the top should be turned to the right; with fig. 3, to the left.) (In the process of paraffin embedding the specimen changed its shape somewhat, chiefly through aceentuation of the several flexures, so that it no longer corresponds exactly with the form shown in the photographs 1 to 3, in consequence of which the level of sections cannot be indicated with absolute precision in the latter, nor any longer quite accurately with straight lines. Previous to embedding the embryo had been stained in tote with Delafield’s hematoxylin. It was sectioned at 10 microns.)

Fig. 5. Section approximately at level b~b’, figs. 2 and 3, showing left forelimb-bud below. (To compare with fig‘. 2 the top of the figure should be turned to left; with fig. 3, to right.) Five spinal ganglia are shown; also the mesonephros, with the pest-cardinal vein dorsally. The section passes through the point where the yolk-stalk was attached to the primitive ileum (shown in transverse section as minute opening at extreme tip of mesentery). 1n the lower portion of the mesentcry are shown the vitelline arteries (superior mesenteric artery). This portion of the coelom contains considerable blood. The aorta is completely filled with blood. Within the umbilical cord, to left of mesentery, is shown the umbilical vein. Adjacent sections contain the largest of the aortic cell-clusters (fig. 7). Photo X 15.

Fig. 6. Section approximately at level c—c’, figs. 2 and 3, through the length of the umbilical cord, showing point of reflection of amnion onto cord, below at the left. (To compare with fig. 2, turn top of figure to left; with fig. 3, to the right). The celom contains much blood; the blood-cells are perfectly preserved and entirely normal. On either side of the umbilical cclom are shown the large umbilical veins. The ventral ramus of the aorta is the inferior mesenterie artery. Note the compressed notochord; it is as wide as in fig 5, Where the section passes very obliquely. Photo. X 15.

Fig. 7. Drawing of large aortic cell-cluster in relation to superior mesenteric artery, 200 microns cephalad of fig. 5. Note the small erythrocytes (microcytes) at right. Above, at the right, is shown a normal-ized erythrocyte; immediately below this cell is a binucleated hemoblast; to the left of the latter appears a hemoblast in ameboid activity. The drawing only shows the cells at one level; in the section the cluster appears more compact due to the cells of adjacent levels coming into view. The preceding sections of the ventral ramus show this vessel free of cells;.hence the cluster projects only along the caudal wall of the mouth of the superior mesenteric artery. X 1000. Drawing by H. C. Cox.
The inclusion by Keibel and Mall” of the Fischel embryo in a series purporting to be typical Warrants the conclusion that they regarded it as normal and representative for this stage of development. However, the fact that our embryo, of a clearly later stage of development, shows even a greater degree of spiral twisting, would seem to contradict the logical deduction that the peculiarity is limited to a certain restricted early stage of development. Nothing in my sections either of the embryo or the chorion, exclusive of the blood content of the celom, which may have resulted from operative trauma, suggests any pathologic condition. The red blood-cells are beautifully preserved and perfectly normal; there is much mitosis especially in the brain and spinal cord, and there is no tissue dissociation or round cell infiltration anywhere. But the body-stalk was abbreviated to such an extent that the amnion fused with the chorion at its placental terminal. Our embryo is accordingly characterized by an unusually short umbilical cord. I conclude that the spiral twist of the embryo is the necessary mechanical consequence of the shortened body-stalk (fig. 6), the result of an attempt to accommodate its lengthening form about a too restricted attachment to the chorion.
The question then a:rises whether such an embryo can develop normally. If the umbilical cord should have failed subsequently to lengthen considerably, the fetus probably would have become malformed caudally, or perhaps even suffered amputation. Among the anomalies described none seem to answer exactly to the requirements of such a process, nor have any to my knowledge been thus interpreted. But the incidental interference with the placental blood-supply in such cases might result in certain of the various moles and merosomatous monsters which have been described. In the region overlying the caudal limit of the umbilicus (fig. 6), the notochord seems to be flattened, as if by undue pressure (compare fig. 6 with fig. 5), while the spinal cord and other organs in this region appear to have a normal shape; but in consequence of the slight transverse obliquity of the sections it seems unsafe to make any very definite statement in this regard. Moreover, the lumen of the central canal is unduly constricted towards its caudal extremity by a close apposition of the lateral walls, which condition may indicate a slight abnormality. The rhombencephalic brain fragmented extensively in sectioning and mounting, but this fact does not necessarily indicate abnormality of this tissue, in view of the apparent integrity of the constituent cells.
Aortic cell-clusters
Aortic cell-clusters have a special importance at this time on account of their bearing on the much mooted question of the hemogenic capacity of endothelium. The embryo under consideration contains such clusters in unusual size and abundance for an embryo of this stage of development (fig. 7). Minot“ was the first~to describe aortic cell-clusters in human embryos, in stages of from 8 to 10 mm. length. But his only illustration (fig. 368, Keibel and Mall’s Embryology”) shows only four cells; nor does his description of these cell aggregations allude specifically to larger clusters; we may therefore infer that the clusters seen in these stages by Minot were only of the smaller variety. In a 5 mm. human embryo (Watt’s embryo) in my collection, previously described,4 no aortic cell-clusters occur, though occasional endothelial cells can be seen separating from the wall of the aorta and assuming hemoblast characteristics. Also in a 13 mm. human embryo (Crenshaw embryo) of my collection no aortic clusters, other than occasional groups of two or at most four cells, occur. The available evidence from human embryos indicates that the clusters of the present embryo are of quite exceptional size and abundance.
Minot” regarded the cells of these clusters as hemoblasts which had become aggregated along the ventral portion of the aorta near the points of origin of the larger ventral rami. The mechanical factors involved were presumably conceived to be the force of the blood stream moving towards the ventral branches of the aorta, and the adhesive properties of the cytoplasm of the involved hemoblasts.
Van der Stricht16 had previously reported comparable aortic cell-clusters in the bat, and Maximoww in the rabbit; and both regarded them as endothelial differentiation products. Emmel2 subsequently described similar clusters for pig embryos of from 6 to 15 mm., and interpreted them as endothelial derivatives resulting from the presence of toxic stimuli, conceived as having their source in the degenerating ventral segmental rami, and the included blood-cell ‘content, of the abdominal aorta. Jordan likewise described these clusters for pig embryos of from 10 to 12 mm.,‘ and in mongoose embryos of from 5 to 7 mm.,“ and reported them also in chick and turtle embryos.’ He only regards them as the expression of a normal hemogenic function-of young endothelium. Danchakoff1also had previously reported them in chick embryos; and Sabin“ now describes comparable structures in the living chick embryo of the second day grown in Locke’s solution.
Van der Stricht regards the cells of the clusters as leukocytes; Emmel’ regards. them as ‘macrophages ;’ Maximow,13 Minot,14 J ordan6,7,8 and Sabin15 as hemoblasts (erythroblasts). Our embryo offers a very favorable material for the demonstration of the endothelial origin of these clusters, and of their erythropoietic function.
The larger clusters are found along the ventral portion of the aorta from a point just behind the level of fusion of the dorsal aortic roots to the level of the inferior mesenteric artery (fig. 6), a distance of about 1.8 mm. They are larger and more crowded about the level of the superior mesenteric artery (figs. 5 and 7). Just forward of the level of the fusion of the dorsal aortic roots for a short distance, at about the level of bifurcation of the trachea (fig. 4), occasional cells may be seen rounding up singly and separating from the endothelium as hemoblasts. The number of larger clusters in the abdominal aorta is eight. Beside these there are also. five smaller clusters, and a number of groups of only several cells and occasional single cell ‘clusters.’ The largest groups are in close spatial relationship with the ‘three chief ventral rami, the celiac, superior mesenteric and inferior mesenteric arteries. In each of these cases some of the cells extend for a short distance into the ramus along one side (fig. 7 ). In contrast with the clusters generally in the pig and the mongoose embryos, where they have a more or less spheroidal shape, in this human embryo they have a generally flattened shape with scattering cells peripherally. The group in relation to the superior mesenteric is an exception, its shape being roughly oval (fig. 7).
Just forward of the level where the aorta divides into the umbilical arteries there occur also several encapsulated cell-clusters. Similar clusters have been described by Emmelz in the aorta of the pig embryo and by myself’-3 in the inferior vena cava of the 12-day loggerhead turtle embryo. The larger encapsulated cluster in our human embryo consists of a group of irregular mesenchyma-like cells interspersed among which are a few spheroidal hemoblasts and a few erythroblasts, the whole group being enveloped by an endothelioid membrane except at its proximal pole where the central mass is continuous with the underlying more condensed mesenchyma. The capsule of the cluster is continuous laterally with the endothelium of the aorta. One of the smaller clusters lies directly over an atrophied ventral ramus. In my former studies?’ 8 I interpreted these clusters as differentiating invaginated areas of ventral endothelium and periaortic mesenchyma. The two clusters here described would seem to confirm the accuracy of this interpretation. The cause of the invagination may be conceived to be the mechanical effect of tissue shrinkage following atrophy of a ventral ramus, thus pushing into the aortic lumen the endothelium and enveloping mesenchyma overlying the area of shrinkage.
Another interesting and instructive ‘cluster’ of this region consists of a single encapsulated cell (hemoblast) similar to those described for the mesenchyma of the mongoose’? (fig. 14) and the pig embryos, and there tentatively interpreted as originating from a binucleated hemoblast, one nucleus of which with its enveloping cytoplasm diflerentiated into a hemoblast, the other into an endothelial cell. The aortic ‘cluster’ here specified also adds plausibility to this interpretation; here a binucleated hemoblast differentiated from the endothelium, and then redifferentiated into an endothelial cell enclosing an erythroblast.
To return to the larger naked clusters; these shade at some point on the proximal pole into endothelium—like cells and then into the underlying more condensed mesenchyma. Since these clusters are relatively very large, flattened and more or less scattered, one can hardly apply to them exactly the same mode of origin outlined for the encapsulated clusters. These larger clusters are clearly difierentiation products of the endothelium, the latter replenished from the underlying hemopoietically active mesenchyma. The cells of the clusters include hemoblasts, and more peripherally a few erythroblasts and erythrocytes (fig. 7). More centrally, and mingled with the more regular hemoblasts are also many cells, in some cases including the majority, which are of‘ less regular shape and contain bean-shaped, bi-lobed, and irregularly lobed nuclei. Some can be seen in ameboid activity (fig. 7). . A few cells also may be seen in mitosis. I incline to interpret the lobed nuclei as stages in amitotic division. Indeed a complete series of stages of amitotic division of these cells can be arranged, and it is difficult to avoid the conclusion that the prevailing mode of division here is amitotic. This conclusion is strongly supported also by the following observation: Certain of the nuclei divide very unequally, the smaller moiety being represented by a mere spheroidal bud. Corresponding with this nuclear size-difference there occur peripherally to the cluster, scattered among normal sized erythrocytes, numerous much smaller erythrocytes with minute nuclei (microcytes) (fig. 7).
A point of prime importance in connection with this and other recent studies,6,7,8 is the close correspondence of my ideas regarding the hemogenic capacity of endothelium (based upon the study of sections of the yolk—sac of the pig embryo5 and the mongoose embryos and of certain intraembryonic blood-vessels including the aorta) and. those arrived at by Sabin” as a result of her studies of the living chick-embryo. Indeed her description of the manner in which certain of the endothelial cells lining the blood-vessels of the area vasculosa of the chick embryo of the second day project into the lumen, become filled with basophilic granules (hemoblasts) and then develop hemoglobin (erythroblasts) is substantially identical with my description for a comparable process in the yolk-sac of the pig embryo of 10 mm. and the mongoose embryo of 5 to 7 mm. The only obvious difference is that in the latter material the hemoblast generally separates from the endothelial wall; though occasionally here also it may become bi- or quadri-nucleated (‘blood—island’) before separating from the wall. And her further description of how such a ‘unicellular blood-island’ in the yolk-sac vessels (area vasculosa), vitelline veins and arteries, and the dorsal aorta divides "and the mass is increased also by the addition of other cells which dijfer entiate from the eradothelium in the neighborhood and creep along the wall to join the first cell’ (p. 202), forming ‘a yellow syncytial mass projecting into the lumen of the Vessel,’ from the surface of which ‘red cells break free from the mass and float away in the blood-plasma,’ corresponds essentially to my conception of the origin and fate of the aortic cell-clusters as formulated from my studies of sections of mammalian embryos. With the admission, then, on the part of former adherents to the ‘angioblast theory of His’ that the blood-vessels of the embryo differentiate from the intraembryonic mesenchyma and that blood-cells may differentiate from endothelium, little of importance remains of this tenacious hypothesis.
The new work of Sabin15 supplies valuable additional data also to the advocates of the monophyletic theory of blood-cell origin. Surely her conclusion that since ‘all of the bl'ood—cells of the chick of the second day of incubation can be seen to have hemoglobin in the living chick . . . . they cannot be considered as forerunners of White blood-cells’ (p. 204) has no meaning as an evidently intended argument in opposition to the monophyletic theory when read in juxtaposition with her preceding statement that the endothelial cell in the process of differentiation into a unicellular blood-island ‘becomes filled with basophilic granules and develops hemoglobin’ (p. 202). No one to my knowledge seriously proposes that leukocytes differentiate from erythroblasts. The monophyletic theory, as I understand it, is based precisely upon the fact of the initial presence of this basophilic cell (primary ‘lymphocyte’ of Maximow), from which first develop erythrocytes and subsequently, from a similar cell, granular leukocytes. In other Words the theory holds that the first leukocytes (‘lymphocytes’) precede in development the first erythrocytes, just as described for the chick by Sabin.
To return to our human embryo, one more point calls for further consideration: This embryo gives the combination of unusually large and numerous aortic cell-clusters (essentially blood-islands) and absence of a yolk—sac. This association may be simply fortuitous; but there may possibly exist a causal connection. It seems reasonable to suppose that when the yolksac end othelium was early incapacitated for hemogenic function due to the compression of the sac between the amnion and the chorion, resulting in disintegration, an extra hemopoietic burden was passed on -to the aorta. This portion of the early hemogenic tissue may have been stimulated to compensatory hemopoietic activity in lieu of that of the yolk-sac.

Fig 1
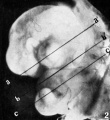
Fig 2

Fig 3

Fig 4

Fig 5

Fig 6
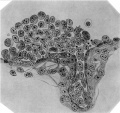
Fig 7

Fig 1-6

{kind=link}
Literature Cited
1. DANCHAKOFF, VERA 1908 Untersuchungen iiber die Entwicklung des Blutes und Bindegewebes bei den Vogelen. I. Die erste Entstehung der Blutzellen beim Hiihnerembryo, u.s.W. Anat. Hefte, Bd. 37.
2. EMMEL, V. E. 1916 The cell-clusters in the dorsal aorta of mammalian embryos. Amer. Jour. Anat., vol. 19, p. 401.
3. His, W. 1880 Anatomie Menschlichen Embryonen. Leipzig.
4. Jordan HE. Description of a 5 mm human embryo. (1909) Anat. Rec. 3:
5. JORDAN, H. E. 1916 The microscopic structure of the yolk-sac of the pig embryo, With special reference to the origin of the erythrocytes. Amer. Jour. Anat., vol. 19, p. 277.
6. JORDAN, H. E. 1916 Evidence of hemogenic capacity of endothelium. Anat. Rec., vol. 10, p. 417.
7. JORDAN, H. E. 1917 Aortic Cell Clusters in Vertebrate Embryos. Anat. Rec., vol. 11, p. 372; also Proc. Nat. Acad. Sci., Vol.3, p. 149.
8. JORDAN, H. E. 1917 Hemopoiesis in the Mongoose Embryo, With special reference to the activity of the endothelium, including that of the yolksac. Pub. 251 of the Carnegie Institution of Washington, p. 291.
9. KEIBEL, F. UND ELZE, C. 1908 Normentafeln zur Entwicklungsgeschichte der Wirbeltiere. Jena.
10. Keibel F. and Mall FP. Manual of Human Embryology I. (1910) J. B. Lippincott Company, Philadelphia.
11. Mall FP. A human embryo twenty-six days old. (1891) J Morphol. 5: 459-480.
12. MALL, F. P. 1916 The Human Magma Réticulé in Normal and in Pathological Development. Pub. No. 224, Carnegie Institution of Washington.
13. MAXIMOW, A. 1909 Untersuchungen fiber Blut und Bindegewebe. I. Die friihesten Entwicklungsstadien der B1ut- und Bindegewebszellen beim Saiigetierenembryo, u.s.W. Arch. f. mikr. Anat., Bd. 73.
14. MINOT, CHARLES S. 1912 Development of the blood. Chapter XVIII, part 1, pp. 523-524, Human Embryology, Keibel and Mall.
15. SABIN, F. R. 1917 Preliminary note on the differentiation of angioblasts and the method by which they produce blood-vessels, blood plasma and red blood-cells as seen in the living chick. Anat. Rec., vol. 13, p. 199.
16. STRICT, O. VAN DER. 1899 L’origine des primiéres cellules sanguines et des primiéres vaisseaux sanguine dans 1’aire vasculaire de chauvessouris. Bull. de l’Acad. Roy. de méd. de Belgique, T. 13, p. 4.
17. TRIEPEL, H. VON. 1914 Altersbestimmung bei menschlichen Embryonen. Anat. Anz., Bd. 46, S. 385.
Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - A study of a 7 mm human embryo with special reference to its peculiar spirally twisted form, and its large aortic cell-clusters. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_A_study_of_a_7_mm_human_embryo_with_special_reference_to_its_peculiar_spirally_twisted_form,_and_its_large_aortic_cell-clusters
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G