Book - Contributions to Embryology Carnegie Institution No.59
| Embryology - 28 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Jenkins GB. Relative weight and volume of the component parts of the brain of the human embryo at different stages of development. (1921) Contrib. Embryol., Carnegie Inst. Wash., 59: 5-54.
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
Relative Weight and Volume of the Component Parts of the Brain of the Human Embryo at Different Stages of Development
Jenkins, G.B. (1921). pp5-54, 12 text-figures, 1 chart.
Introduction

The following volumetric study of the developing encephalon in the human embryo was undertaken as a supplementary part of a program of investigations in the morphology and differentiation of the central nervous system with which I have been engaged for several years. The work was made possible through the kindness of Dr. Streeter who, in furtherance of its purpose, has placed at my disposal the collection of embryos of the Carnegie Laboratory of Embryology, together with models, notes, and such other related material as has been accumulated. Under these conditions it was possible to secure a sufficient number of developmental stages for the investigation, including the consideration of the relative rate of growth of the various parts of the encephalon during the whole period of intrauterine life. It is with pleasure that I take this opportunity to acknowledge my indebtedness for the hospitality, aid, and interest that have been extended to me on the part of the staff of this laboratory.
The question of growth of the animal body as a whole and of its various component parts has occupied the minds of many investigators over a long period of time, and yet in the mass of literature which has resulted from these studies comparatively Httle is to be found that deals with the volumetric development of the central nervous system in the human embryo. A study of the literature on the subject of weight, size, degree of development, etc., of the brain reveals the fact that practically all of the observations reported have dealt with postnatal material and the great majority with adult brains, secured in the main from various institutions, such as asylums for the insane, almshouses, and homes for the aged. The great questions seem to have been those concerning body size, the attempted correlations between brain size and intellectuality, and the controversy over the differences in brain size in the two sexes. Not a few reports frankly deal with grossly pathological material. Much additional work, of anthropological value mainly, has been done along the lines of craniometry, notably by West (1894), who made a number of observations upon school children of both sexes, recording the head measurements as a part of the general growth conditions, Lewandowsky (1910), and HrdUcka (1919), who treat of the more pertinent question of the correlation between skull size and cranial capacity.
Aside from these latter cases the bulk of the work reported may be roughly grouped into four divisions :
- (a) The study of brains of individuals of superior intellectual attainments (Spitzka, 1907; Marshall, 1892; Manouvrier, 1885).
- (b) The tabulation of the brain-weights and measurements obtained from large series of cases, including material of various races and ages and representing both sexes (Boyd, 1861; Donaldson, 1895; Ziehen, 1899, 1903; Vierordt, 1893).
- (c) Statistical studies in which these tables are used to determine the correlations existing between brain-size and age, brain-size and intellect, and the comparison of brain, body, and intellect in the two sexes at various ages and in different races, as has been so ably done by Pearl (1905a, 19056) who, in addition to the above material, used the tables reported by Bischoff (1880), G. Retzius (1900), Marchand (1902), Matiegka (1903), and Boyd-Marshall, all as reported by Donaldson (1895). The employment of such biometric methods gives results of great value to the investigator, since it enables him to make a great number of comparisons and to tabulate and express his results with a clarity and an exactness not possible by any other method.
- (d) This last class would embrace the relatively limited number of observations dealing with material sufficiently young to come within the scope of the present paper, and includes extracts from Major Boyd's (1861) monumental work on approximately 2,600 cases autopsied at Marleybone Workhouse and the Somerset Lunatic Asylum. These cases were grouped, according to age, into 18 periods, the first 3 of which are of value in this connection. They comprise 48 premature stillborn, 83 term still-born, and 90 new-born infants. The various measurements as given by Boyd were the total body -length from the vertex to the inner side of foot behind the ball of the great toe, and the following head measurements : circumference around the occipital protuberance to the space behind the eyebrows; transverse, from the opening of one ear, over the vertex to the opening of the other. Doubtless the method of subdividing the encephalon employed by Boyd was, as assumed by Donaldson, for the purpose of making the cerebrum one unit, the cerebellum one unit, and the medulla, pons, and midbrain one unit; though it is probable that a part, at least, of the mesencephalon was included with the cerebrum, since, upon removing the calvarium and dura, Boyd "removed the right and left hemispheres of the cerebrum in sUces to the tentorium." The cerebellum was isolated by cutting the crura close to the pons.
A closer study of the group of specimens tabulated as premature still-born, which are recorded as measuring 10 to 18 inches total length (CH) , or approximately 250 to 460 mm., shows that they would correspond to an age range from the end of the fifth to the end of the eighth month, according to Stratz's tables as reported by Martin (1914). According to Mall (1910), these two extremes would for the younger correspond to a sitting-height (CR) of 167 mm., age 20 weeks; and for the older a sitting-height of 310 mm., age 35 weeks. The weights of the brain parts in these two extremes would be in the smaller fetus 41.25 grams for the cerebrum, 1.98 grams for the cerebellum, and 1.32 grams for the medulla-pons ; and in the larger fetus 297 grams for the cerebrum, 24.75 grams for the cerebellum, and 1.65 for the medulla-pons. These figures must at best be accepted with reservations, since the tables from which they are taken are undoubtedly inaccurate in some particulars. For example, the weight of the encephalon in no case equals the added weights of the various parts of the same brain. These results are probably due to the use of crude apparatus and to a tendency to give results in round numbers.
Ziehen (1903) includes 9 embryos in a large series of cases reported. The age of 6 of these was 30 days and the recorded total brain-weights were respectively 0.98, 1.0, 1.05, 1.13, 1.15, and 1.15 grams; 3 were 29 days old and the total brain-weights were 1.29, 1.23, and 1.15 grams respectively. All of the material has been preserved in formalin.
Michaelis (1907), in a number of cases of children autopsied by him in Leipzig, reports 7 groups of embryos. Jackson (1909), in a study of prenatal growth, records observations made upon 43 specimens ranging from 6 mm. to full term. In these studies the measurements were by volume and not by weight, as in the other cases recorded. The volume was determined in some cases by the amount of wa'er displacement of the brain of the embrj^o modeled in wax by the Born method; in others by making enlarged drawings of the parts, measuring these drawings with a planimeter and calculating the volume by multiplying the areas by the thickness of the sections. His conclusions are that the brain, although subject to considerable variation, shows a fairly regular curve of growth. At the second month it forms slightly more than 20 per cent of the total body-volume, the average dropping to 12.8 per cent in still-born fetuses. In the hving-born, however, the average was about 14.6 per cent.
Dockeray (1915), working toward the same end which is the purpose of my own paper, and using similar methods, gives a complete volumetric study of the brain parts in a fetus 156 mm. in length, and I have incorporated his results in this paper.
Methods and Material
Two points in the technic employed in my own studies were determined upon because their worth had been proved, both in this laboratory and by other workers :
First, the use of the crown-rump or sitting-height as a standard for determining the size of the material used: Both Le Bon (1879) and Donaldson (1909) agree that body-length is a better criterion than body-weight from which to infer brainweight. The use of this method relieves us of the necessity of determining the leglength, an unsatisfactory procedure even in older fetuses.
Second, the method of preservation : All of the material used in this study had been preserved in formalin which alone in varying strengths, or in combination with other agents, is by far the most universally used preservative for animal tissues. Hrdlicka (1906) experimented with the effects of various preservatives upon the brains of both human and lower vertebrates and found formalin the most satisfactorJ^ Though confirming in part the work of others who claim that this agent causes an increase in volume in the tissues preserved in it, in his summary he states that the simple formalin solutions all show the same effects in all brains. These consist of a sharp initial rise in the weight of the specimen, reaching a maximum within less than a week, and a subsequent gradual, long-continued decrease. The rise is in inverse ratio to the strength of the formalin solution, the percentage of loss being apparently independent of the formaUn percentage.
Many workers have tested out various fixatives with more or less indifferent success, and while for special tissues and under favorable conditions many satisfactory methods have been developed, still for general purposes, where availability of the agent and the ease with which it can be prepared are the main desiderata, none have yet been discovered which are as satisfactory and consequently as widely used as is formalin in varying degrees of strength.
In order to secure the necessary data upon which to base conclusions, a great number of embryos were studied and such stages selected as it was thought would best show the developmental phases under consideration. Out of this number, 10 specimens were chosen, ranging in size from 4.3 mm. CR, estimated age 4.5 weeks, to a new-born infant of 367 mm. CR. All the material used is to be found in the Carnegie Embryological Collection. In table 1 will be found listed the embryos that were selected as being particularly suitable for the purposes of this study. They are arranged serially in the apparent order of their development. The catalogue numbers given are those under which the embryos are listed in the records of the laboratory. The measurements all signify the crown-rump length in millimeters. The ages in weeks were determined from the last known menstrual period and conform to the curve of age (based on length and weight) plotted by Streeter (1920). All of the specimens had been preserved in formalin, and the first 8, in addition to fixation, had been embedded, cut, and stained. The material was used as found, no attempt being made to account for any tissue changes dependent upon the technic. The volume is therefore that of the embedded specimen. The shrinkage resulting from the embedding process introduces a considerable error in connection with actual volume, but as the shrinkage of the component parts of a given brain is uniform, the error does not prevail in reference to the relative volume of the separate parts.
In each of these younger embryos the brain was projected, drawn, and modeled, according to the well-known method of Born. (See table 1 for magnifications used in each instance.) The resulting models could then be easily subdivided into the various parts decided upon and the weights and volumes ot these readily obtained. The two older specimens were of a sufficiently advanced stage of development to permit of dissection and weighing of the actual tissue.
Table 1. List of specimens the brains of which were studied.

The first question that arose was the selection of a method of subdivision which could be adapted to all stages and which would yet permit the greatest number of dissections and comparisons. Only simple determinations were possible in the early stages, additions being made as the encephalon increased in complexity. In seeking to establish definite landmarks by which to be guided in dehmiting the various parts, it was decided to base the initial procedure upon the early subdivision of the encephalon into the primary brain vesicles and then to add such later subdivisions as could be accurately determined by a study of the gross models and a microscopic study of the tissues. This plan permitted the development of a logical method of procedure.
Since in all stages it was possible to delimit the three early vesicles - the rhombencephalon, mesencephalon, and prosencephalon - the first point to determine was that of landmarks by which these vesicles could be separated from each other and the hindbrain from the spinal medulla. In the latter, of the three criteria most commonly used - the highest root fibers of the first spinal nerve, the lowest root fibers of the hypoglossal nerve, and the lowest crossing fibers of the pyramidal decussation the first proved to be the most satisfactory. Accordingly, the first section was made in a transverse plane immediately anterior or cranial to the highest rootlets of the suboccipital nerve. This section was the same in all cases, and the resulting anterior mass, the encephalon, was the portion used for these studies. The second section the separation of the hindbrain from the midbrain was made in the youngest specimen (4.3 mm. CR length) in the constriction immediately in front of the outward curve of the rhombic lip, the cerebellar rudiment. Later, when it was possible to discern the inferior coUiculi and the superficial transverse pontine fibers, the incision was started dorsally, just posterior to the colliculi, to end ventrally immediately anterior to these pontine fibers.
The mesencephalon is much more prominent proportionally in the younger subjects than in the older, and forms the arch of the cephalic flexure. It is wedgeshaped, the ventral surface being concave and shorter, the dorsal surface convex and longer, and delimited in front and behind by transverse surface grooves. The posterior incision was comparatively easy and was made, as described above, to remove the rhombencephalon. The line of demarcation between the mesencephalon and the prosencephalon was, however, more difficult to follow, since it was found upon studying the tissues that the thickened lateral plates of the mesencephalon project forward into the prosencephalon or, more properly speaking, are invaginated as a result of the caudal growth of the prosencephalon, so that the line of incision must extend anteromesially between these mesencephalic plates and the more prominent overlapping masses of the thalamencephalon. This lateral thickening of the mesencephalic plates, which is more pronounced in the basal parts ventral to the iter (the site of the future tegmental nucleus), is sufficiently well marked in these early stages to prevent error. Later, other landmarks make their appearance; dorsally, the epiphysis, the habenular nuclei, the posterior commissure, and the increasing prominence of the posterior extremities of the thalami; anteriorly, the pulvinars; posteriorly, the differentiation of the superior coUiculi; and ventrally, the developing mammillary bodies. In older subjects, therefore, this anteromesial incision must, on each side, extend between the mesencephalon and the thalami, to end just caudal to the epiphyseal attachment dorsally and the mammillary bodies ventrally.
Having divided the entire encephalon into the three primary vesicles, the succeeding steps consist in separating these structures into their more prominent constituent parts. In the 4 mm. embryo it was not possible to accurately remove the cerebellar rudiment. In the 16 mm. embryo, in the case of the hindbrain, the cerebellum could be removed from the stem. In the early stages this was accomplished by removing the prominent anterior rhombic lip on either side, which corresponds to the cerebellar rudiment, as shown in figure 2. This mass, at first a prominent roll or lip, becomes more and more sharply defined in the succeeding stages and is delimited from the mesencephalon by the transverse groove mentioned above. Owing to the difficulty of accurately separating the pons and the medulla, they were considered as a unit mass. The mesencephalon was considered as a single mass throughout the series. The fore-brain was then subdivided into the telencephalon and the diencephalon. In the early stage this division was accomplished by cutting along the thalamic margin, which extends as a slight groove along the dorsal border of the optic evagination and its later developing stalk connected with the diencephalon. In the older specimens the separation was effected by cutting through the internal capsule between the thalamus and the corpus striatum dorsally, the stria terminalis being used as a guide; ventrally the incision was made along the lateral margin of the optic tract, leaving this structure connected with the thalamencephalon. Thus the diencephalon comprises the optic tracts, the thalamus, the epithalamus, and the hypothalamus (except the hypophysis, which could not be considered in all cases). The diencephalon was not subdivided. The telencephalon in all stages was subdivided into the neopallium and the archipaUium. In the early stages the line of incision was made to include the shallow depression in the lumen of the ventral wall of the first vesicle, lateral to the lamina terminalis and anterior to the optic stalk. In the older specimens (16 mm. and upwards) this rudiment is larger and consequently more easily separated from the remainder of the vesicle. Here there is a considerable ventral prominence appearing lateral to the lamina terminalis anterior to the torus opticus. This comprises the olfactory bulb and tract. The latter extends along the lower or ventral margin of the depressed cortical area over the base of the corpus striatum. In older subjects (50 mm. and upwards) the anterior commissure, paraterminal body, fornix, and hippocampus could easily be made out by microscopic studj^ and as they became differentiated they were removed and studied.
The method employed in isolating these various structures in the older modeled embryos was essentially the same as that followed in dissecting the brain of the term fetus, which was as follows: The cord was separated from the medulla at the conventional level (in these specimens the bodies had been injected with a 10 per cent solution of formalin and the brain, after removal, had been kept in the same solution until studied) and the upper part of each cerebral hemisphere removed by a transverse incision made just above the level of the fibers of the corpus callosum. The occipital lobe on each side was then removed flush with the caudal margin of the splenium and the basal surfaces of the temporal lobes were pared away until the junction of the mesencephalon and diencephalon could be seen. The mesencephalon was then separated from the rhombencephalon by an incision passing just cranial to the transverse pontine fibers ventrally, and just caudal to the posterior collicuU dorsally. The mesencephalon was next removed by an incision passing anteromedially between the anterior coUiculi and the pulvinars, just caudal to the epiphyseal stalk dorsally and the corpora mamillaria ventrally. Next, the olfactory bulbs and tracts were freed and removed by cutting each tract at its junction with the base of the hemisphere. The two hemispheres were then separated by cutting through the median sagittal plane of the corpus callosum, the lamina terminalis, chiasma, and the basal structures, the incision passing between the mammillary protuberances. The resulting blocks were treated alike; the corpus callosum was lifted up and carefully dissected off from the fornix, exposing that structure, as well as the thalamus, stria terminalis, and the corpus striatum. The remainder of the cortex was removed anteriorly, with the head of the caudate nucleus as a guide. Laterally, the cortex of the insula was pared down to the lenticular nucleus and the temporal lobe removed, leaving the hippocampus, the fimbriae, and the fornix intact, Following the fornix around anteriorly, its anterior pillar and the mammillary and paraterminal bodies were removed together. The thalamencephalon was then separated from the corpus striatum by cutting through the internal capsule, following dorsally the line of the stria terminalis, and ventrally, cutting just lateral to the optic tract, leaving that structure connected with the diencephalon. The cerebellum was then separated from the remainder of the hindbrain by cutting through the peduncles flush with the hemispheres.
This method of subdivision gave 8 separate units for study: (1) the medullapons, (2) the cerebellum, (3) the mesencephalon, (4), the diencephalon, (5) the fornix and hippocampus, including the paraterminal and mammillary bodies, (6) the olfactory bulb and tract, (7) the telencephalon, and (8) the corpus striatum. In all of the specimens a regrouping of these units was made, giving us the following additional units: (1) the total hindbrain, which comprises both the cerebellum and the meduUa-pons ; (2) the total telencephalon which was subdivided into archipallium (including the olfactory apparatus) and the neopallium that is, the remainder of the telencephalon, minus the corpus striatum, which in the more advanced specimens was considered as a separate unit. In each instance the total weight and volume of the encephalon were determined; then those of each of the subdivisions were ascertained and the percentages calculated from the totals. The actual weight and volume could both be determined in the case of the dissected specimens, while for the modeled specimens the weight and volume of the entire model, or of a part, were first determined. The weights of these wax models were simply carried through, no attempt being made to ascertain the actual weight, since the relative values would have been the same. The volume of the model was obtained by determining the volume of a gram of the wax used in making the model and multiplying this by the weight of the part under consideration. The cubic centimeter vaUie of the wax being thus obtained in grams, it was a simple matter to calculate the various amounts, percentages, etc. The actual volume was obtained by dividing the model volume by the cube of the magnification used in the reconstruction.
In making this study the companion parts of the two sides have in every instance been grouped together, thus giving one unit for comparison instead of studying each of these paired structures separately. In all cases, too, the percentage weight has been chosen as the unit of comparison, since this method gives a stable basis, whatever may have been the magnification used in the reconstruction, and would apply equally well when the actual weights were considered, as in the two dissected specimens. Weight, rather than volume, has been selected for comparison for the same reason that model weights were considered instead of attempting to calculate actual weights, since it was thought that by reducing the method employed to the simplest possible working basis the percentage of error would likewise be reduced to a minimum. It is to be remembered, also, that these percentage weights are of only relative value, for the various parts of the encephalon present a steady and consistent growth and development throughout the period of gestation, and that the decrease in percentage weight, recorded for certain parts, as opposed to increased percentages for others, means merely that those parts which present an increase in weight grow so much more rapidly that they outstrip their more sluggish neighbors in relative bulk, as will be seen in a study of both models and tissues.
In every case, except the two oldest specimens, a careful check was kept upon all steps by a microscopic study of the tissues. This was found to be especially valuable in determining the landmarks to guide the incisions. However, while occasional reference may be made to the degree of differentiation, it is the intention at this time to deal entirely with the question of growth in bulk or mass, without reference to histologic detail.
Descriptions of Individual Specimens
No. 148, 4.3 mm., estimated menstrual age 4.5 weeks

This embryo (see table 3 and fig. 1), which has been the subject of a number of studies by various investigators, presents some quite unusual features and was selected for the reason that, for a young stage, it presents the various developmental features to such a degree as to render it especially valuable as a basis upon which to build a serial study of the various parts of the encephalon. At this stage the central nervous system is distinctly tubular in character and the anterior neuropore is closed . Anterior and dorsal to the eye-stalks, and separated from them by a distinct fold, can be seen the olfactory region or archipallium, and more dorsally the neopallium, consisting at this stage of two faintly outlined evaginations, one to either side of the mid-line. The entire prosencephalon comprises 31.3 per cent of the weight of the encephalon and can be quite easily subdivided into the telencephalon, weighing 7 per cent, and the diencephalon, weighing 24.3 per cent. The further subdivision of the telencephalon yielded an archipallium of 4.2 per cent and a neopaUium of 2.8 per cent. The cephalic flexure is well defined; its arch, rather narrow, is formed by the mesencephalon, which constitutes 14.3 per cent of the encephalon. The rhoml^encephalon at this stage is jaroportionately very large, comprising 54.4 per cent of the total weight of the encephalon.
A study of this vesicle shows that the lateral plates have a wide dorsal spread; the thin roof plate is closely applied to the overlying ectoderm and only a slight thickening is observed at the point of junction of the roof and lateral plate - the rhombic lip. The cerebellar rudiment is merely a thickened fold which stretches across the more anterior, cephalic portion of the dorsum or roof, showing a tendency to overlap the constricted caudal extremity of the mesencephalon. The cerebellum is not sufficiently differentiated at this stage to permit of its delimitation. The site of the pontine flexure is not indicated at this stage, though both the cervical and the cephalic flexures are well marked.
No. 43, 16 mm., estimated menstrual age 7 weeks
This specimen (see fig. 2) presents a much more advanced stage of development. The tubular character is less pronounced and the pontine flexure is well defined, serving to accentuate both the cervical and cephalic flexures. That portion of the rhombic lip constituting the cerebellar rudiment was sufficiently well differentiated to permit of its removal and study, 36.63 per cent of the total brain. Of this the cerebellum comprises 9.74 per cent and the medulla-pons 26.89 per cent, a slight increase for the former, a considerable decrease for the latter. The mesencephalon has increaised to 14.76 per cent, the total prosencephalon to 48 per cent, to which the diencephalon contributes 19 per cent - a slight shrinkage, and the telencephalon 28.84 per cent, a gain of 7 per cent over the preceding stage. A further study of the telencephalon shows an archipallium of 1.43 per cent, a tremendous drop as compared with 18.09 per cent at 16 mm. The neopallium has, on the other hand, increased to 27.41 per cent, a complete reversal of relative values.
The basal ganglia, both the thalami and the corpora striata, show a considerable increase in contributing 7..56 per cent of the total weight as oppoised to 38.11 per cent for the medulla-pons, the combined weights giving a total of 45.67 per cent for the rhombencephalon, a relative decrease from the preceding specimen. The mesencephalon has decreased to 12.52 per cent and the diencephalon to 20.38 per cent, whereas the telencephalon has increased to 21.43 per cent, three times its weight in the younger embryo. Of this the archipallium constitutes 18.09 per cent and the neopallium 3.34 per cent, making a total of 41.81 per cent for the prosencephalon, as opposed to 31.3 per cent in the 4.3 mm. stage.
No. 584a, 25 mm., estimated menstrual age 8 weeks
This embryo presents a still further increase in development, the rhombencephalon weighing bulk at this stage. The walls of the vesicles also show a corresponding increase in thickness, so it will be observed that, although there has been a marked increase in size and weight of the encephalon up to this stage of development, it consists in cell proliferation rather than in dedifferentiation and does not permit of much added study of individual parts.
No. 96, 50 mm., estimated age 11 weeks
| This embryo (figs. 3, 4, and 5) has advanced much farther in development and permits of a greater amount of subdivision of the model. The total brain-weight is 514.58 grams, of which the rhombencephalon comprises 12.77 per cent, the cerebellum contributing 3.79 per cent and the medulla-pons 8.96 per cent, a relatively considerable decrease in all three values. The mesencephalon has likewise decreased, weighing in this specimen but 7.09 per cent of the total, as compared with twice that amount in tlie 8 weeks' embryo. The total prosencephalon has nearly doubled its relative weight, being now 80.14 per cent, the diencejihalon drojiping to 12.77 per cent, and the telencephalon increasing to 67.37 per cent, of which the archipallium constitutes 3.52 per cent and the neopallium 51.18 per cent, a tremendous increase over all preceding stages. The corpora striata, which could be isolated in this specimen, weighed 12.67 per cent of the total brain.
The various parts of the archipallium could also be isolated, giving an olfactory bulb weighing 0.3 per cent, and the paraterminal body, fornLx, and hippocampus, which together weighed 3.22 per cent of the total. The pontine flexure is much less acute and the cerebellum, as yet rudimentary, is a narrow, beaklike structure (fig. 3). The mesencephalon shows a distinct caudal projection overhanging the median part of the roof of the cerebellar rudiment. The cerebral hemispheres project well back over the diencephalon, encroaching upon the mesencephalon. A curious feature, noted only at this age, is that the corpora striata, the diencephalon, and the rhombencephalon, are all of equal weight value. |
 Fig. 3. Mesial view of right hemisphere of embryo No. 96, crown-rump length 60 mm., showing subdivisions of archipallium. X 5. Fig. 4. Left view of model of brain-stem (same embryo as shown in fig. 3), showing its subdivisions. X 5 Fig. 5. Mesial view of hemisection of model shown in fig. 4, showing subdivisions of diencephalon. X 5. |
No. 1400-20, 69.5 mm., estimated age 12.5 weeks
This specimen still more closely approaches the full adult differentiation. The cerebral hemispheres have grown in all directions to a considerable degree, the total prosencephalon weighing 84.84 per cent of the whole. Of this the diencephalon contributes 11.47 per cent, a steady relative decrease, despite the increase in bulk of the developing thalami. The archipallium has relatively decreased, weighing but 2.87 per cent, of which the olfactory bulb contributes but 0.15 per cent and the paraterminal body, fornix, and hippocampus 2.72 per cent. The neopallium, on the contrary, weighs 60.62 per cent and the corpus striatum 8.88 per cent, making a total of 69 per cent of the entire cncephalon. The mesencephalon has dropped to 5.97 per cent and the total rhombencephalon to 10.19 per cent, of which the medulla-pons represents 6.23 per cent and the cerebellum 3.96 per cent, thus showing the continued and rapid increase of the prosencephalon, especially the neopallium, over the other parts.
No. 1400-22, 80 mm., estimated age 13.5 weeks
| A still more decided advance in bulk and differentiation is here observed (fig. 6). The cerebral hemispheres are considerably more extensive, the telencephalon totaling 80.56 per cent of the entire weight of the encephalon. Of this the neopallium constitutes 65.55 per cent and the corpus striatum 11.23 per cent, a total of 76.78 per cent. The archipallium weighs 3.78 per cent, the olfactory bulb contributing 0.35 per cent and the paraterminal body, fornix, and hippocampus 3.43 per cent; all of which agree substantially with the figures given for the corresponding parts of the preceding embryo. On the other hand, the neopallium has increased and the diencephalon has diminished to 8.04 per cent, the total prosencephalon having increased to 88.6 per cent. The mesencephalon presents a sligh decrease, weighing now 4.78 per cent. The total rhombencephalon has also lost relatively, weighing 6.62 per cent, of which the cerebellum comprises 2.89 per cent, the medulla-pons 3.73 per cent. |

No. 1400-2.5, 119 mm., estimated age 16 weeks
| The advance in general bulk is distributed in this embryo (figs. 7 and 8) as follows: The total prosencephalon constitutes 91.91 per cent of the brain weight, of which the diencephalon comprises 6.29 per cent - a decrease, the telencephalon having meanwhile increased to 85.62 per cent. To this the neopallium contributes 69.55 per cent, the corpus striatum 11. C4 per cent, and the archipallium 4.43 per cent. Of the latter the olfactory bulb shows a decreased weight of 0.28 per cent, and the paraterminal body, fornix, and hippocampus a weight of 4.15 per cent - a slight rise. The mesencephalon has fallen to 2.06 per cent.
The total rhombencephalon weighs 6.03 per cent, of which the cerebellum forms 2.74 per cent and the medulla-pons 3.29 per cent, all of which weights are practically identical with those of the corresponding parts in the 80 nmi. specimen, so that the telencephalic gain is balanced by the losses sustained by the diencephalon and the mesencephalon, the fractional increase in the rhinencephalon being likewise accounted for. |

No. 1400-28, 156 mm., estimated age 19 weeks
| This specimen (figs. 9, 10, 11, and 12) was studied and published by Dockeraj- (1915), and his findings have been rearranged and incorporated into this study. The total prosencephalon has attained the enormous weight-value of 93.69 per cent of the total brain weight, the diencephalon dropping to 4.81 per cent and the
telencephalon weighing 88.78 per cent. Of the latter the neopallium comprises 78.82 per cent and the corpus striatum 7.02 per cent, the former an increase, the latter a decrease as compared with llie preceding stage. The archipallium has declined to 2.34 per cent, the olfactory bulb weighing 0.14 per cent, the paraterminal body, fornix, and hippocampus 2.2 per cent, the lowest point yet reached by these parts. The mesencephalon has also decreased, constituting but 1.44 per cent of the encephalon. The total rhombencephalon weighs 4.97 per cent, sUghtly below that of the 119 mm. specimen, the loss being sustained by the meduUa-pons which now weighs 2.23 per cent, while the cerebellum weighs 2.74 per cent, the same weight-ratio as that in the preceding stage and virtually the same as that in the 80 mm. stage. This structure thus maintains a relative weight-level extending over a period of six weeks, which really means that there has been an acceleration of growth during this interval. The foregoing specimens were all modeled and the models separated into these various parts for study.
Fig. 9. Ventral view of a reconstruction of brain of embryo No. 1400-28, 156 mm. crown-rump length, showing manner in which it was subdivided. X 2. |
 Fig. 10. Mesial view of right half of telencephalon of embryo shown in fig. 9. X 2. Fig. 11. Mesial view of the subdivided archipallium of embryo shown in figs. 9 and 10. X 2. Fig. 12. Drawing of a hemisection of brain shown in figs. 9, 10, and 11, with subdivisions of basal ganglia outlined. (Section 6, slide 145.) X 2. |

Fetus 230 mm, estimated age 26 weeks
This stage is represented by a fetus of which only the brain was available for study. Since the specimen had reached a sufficiently advanced state of development to permit the removal and dissection of the brain, the actual weights and volumes can be given. In form and fissures this brain corresponds to a stage between the two shown by Ketzius (1890, plate 14, figures 3, 4, 5, and figures 6, 7, 8), and has therefore been placet! at 340 mm. total length, or 230 mm. crownrump length, and its age estimated as 20 weeks. These data were obtained from Dr. Streeter, who dissected and studied the spechnen.
The total prosencephalon weighed 93.32 per cent of the encephalon, a fractional loss as compared with the 156 mm. fetus. Of this the diencephalon constituted 3.7 per cent, less than the preceding one. The telencephalon, the total weight of which was 89.62 per cent, was subdivided into the neopallium, weighing 79.21 per cent, and the corpus striatum, weighing 8.33 per cent, both showing an increase. The archipallium weighed but 2.08 per cent, the lowest point yet reached. To this the olfactory bulb contributed 0.13 per cent, the paraterminal body, fornix, and hippocampus, 1.95 per cent. The mesencephalon weighed 1.42 per cent, practically the same as in the preceding .specimen. The total rhombencephalon weighed 5.26 per cent, a somewhat greater relative weight than at 19 weeks. This added amount was contributed by the cerebellum, which weighed 3.15 per cent, as comparetl with a medulla-pons weight of 2.11 per cent, which is only a small fraction less than in Dockeray's fetus, the last and largest of the modeled specmiens.
No. 2558, 367 mm., new-born white male
This specimen Template:CE2558 had been injected and preserved in 10 per cent formalin. The sitting height was 367 mm., body-weight 3,210 grams. The head measurements were as follows: length 130 mm., width 102 mm., circumference 381 mm., biaural arc 248 mm. The brain was removed by Dr. A. H. Schultz, of the Carnegie Embryological Laboratory, to whom I am indebted for the foregoing data. Judging from macroscopic appearance, both body and brain were normal and well developed. The encephalon weighed 505.61 grams; the actual volume, as determined by water displacement in a large graduated cylinder where the indices could be easily read, was 489.97 c. c. The cerebral hemispheres were very large, richly convoluted, and had thickened until the cavity of the lateral ventricle was reduced to a mere slit. The total prosencephalon weighed 467.12 grams, or 92.38 per cent of the whole brain, slightly less than that of the preceding. Of this the diencephalon contributed 2.03 per cent, a loss of about 30 per cent as compared with the 230 mm. fetus.
The telencephalon weighed 89.75 per cent of the whole, maintaining the same weight-value as the preceding specimen. Of this the neopallium weighed 424.3 grams, or 83.92 per cent, and the corpus striatum 24.3 grams, or 4.8 per cent, a consiilerable relative gain for the former. The archipallium has declined to 5.2 grams or 1.03 per cent, which is the lowest mark in the weight-curve for this complex. Of this the olfactory bulb contributed 0.2 per cent, and the paraterminal body, fornLx, and hippocampus, 0.83 per cent. The mesencephalon likewise reaches its lowest relative level at this stage, weighing about 0.64 per cent of the total weight of the encephalon. The rhombencephalon weighed 35.28 grams, or 6.98 per cent, the medulla-pons weighing 4.58 grams (0.91 per cent) and the cerebellum 30.7 grams (G.07 per cent). This represents for the former the lowest point in the series, and for the latter a marked gain over preceding figures. In this specimen the total brain constituted 15.75 per cent of the body-weight, a somewhat greater proportion than is usually reported. Jackson (1909) gives 14.6 per cent and Vierordt (1893) 12.29 per cent.
Brain Volume at Different Stages

Since we have the volume of the models of the brain and its separate parts for the different stages of development, it is possible to roughly calculate the actual volume by dividing the model volume by the cube of the magnification used for the reconstruction. It must be borne in mind, however, that since we are dealing with brains that have been embedded in paraffine or celoidin, the models represent the embedded brain and one must take into account a considerable shrinkage of the tissues. The shrinkage of a given brain is more or less uniform throughout and therefore, in itself, does not interfere with the determination of the relative volume of its different parts. As regards the actual volume, however, one must take into consideration the primary swelling of the brain when placed in a formalin solution and its subsequent shrinkage in the course of its preparation into serial sections. From our own experience and that of others we can estimate that the volume of the brain as embedded is from one-tenth to one-third less than the original, depending on the character of the tissues and their reaction to formalin, and on the dehydrating and embedding media. The extent of this shrinkage for the individual brains I have at present no way of accurately determining. In spite of the fact that my figures are only approximate (probably one tenth to one-third less than the original volume), the interest attached to the question of the volume of the fetal brain perhaps warrants my including the data given in table 2, which were obtained on the basis of the volume of the models. From these same figures one can readily calculate the volume of any individual part of the brain by multiplying the total volume by the percentage of the total brain formed by that particular part (table 3).
Table 3. Data on the relative weight and volume of the subdivisions of the brain at various stages of development.
With the exception of the two oldest stages, these data are based on wax-plate reconstructions. Where the structures are bilaterally symmetrical the data for the right and left are combined.

Summary
A study of the preceding data shows the rapid growth of the brain as a whole and the relatively enormous rate of growth of some of its component parts as compared with others; for, while all parts of the brain consistently increase in size, they do not all grow at the same rate. The telencephalon, for example, shows a relatively rapid increase throughout the entire series, greatly surpassing all other parts; the cerebellum shows a similar, though less marked increase during the latter half of gestation; while all of the other parts show a relative decrease in weight- values.
Starting with an embryo of 4.5 weeks, 4.3 mm. long, with an actual total brain volume of approximately 0.003 c.c, and in which it is possible to definitely outline little more than the three primary brain vesicles common to all vertebrates, one can follow the rapid growth and development until birth, when this complex and highly specialized organ attains a total actual volume of 490 c.c, a gain in less than 36 weeks of over 160,000 times the initial volume.
An analysis of the values for the different parts shows a steady upward curve for the prosencephalon, from 31.3 per cent in the early embryo to 92.38 per cent at term, a gain of nearly three times its initial bulk, despite the fact that all of its component parts, except the telencephalon, have shrunken considerably in weightvalues. The percentage of the total brain-weight formed at different stages by the five chief subdivisions of the brain is shown graphically in chart 1.
The most striking feature in this growth curve, as well as the most significant one, is to be found in the enormous proportional and actual increase in the size of the telencephalon, which gives the amount of cortical expanse necessary to provide for the control of all subsidiary parts of the nervous system, as well as being the seat of the higher psychic functions. Beginning with an initial weight of 7 per cent at 4.5 weeks, the telencephalon increases rapidly up to 13.5 weeks, when it constitutes 80.56 per cent of the entire encephalon. Then follows a period of more gradual growth up to the twenty-sLxth week, when the telencephalon attains its maximum relative weight (89.62 per cent) which it maintains until term. Assuming that the specific gravity of the brain tissue is very little over 1.0, the actual increase in weight is from approximately 0.0002 gram (as calculated from volume) at 4.5 weeks to 493.8 grams at birth.
Another noteworthy point in the growth rate of the cerebral hemisphere is that it attains its maximum relative weight before any folding occurs to increase the extent of its periperhal surface, showing that in the later weeks its development is one of complexity rather than of bulk, a fact which would appear to argue that these later changes consist in perfecting the development of already existing structural units, rather than in the acquisition of new ones.
These figures for the telencephalon are rendered even more striking when we study the growth conditions of its various subdivisions. It was possible in all cases to separate this structure into the neopallium and the archipallium. At 4.3 mm. the undivided neopallium weighed 2.8 per cent; at 25 mm. 27.4 per cent; at 50 mm. 51.18 per cent, the curve continuing to mount rapidly to a maximum of 83.92 per cent at term. At the stage of 50 mm. it was possible to isolate the corpus striatum which weighed 12.67 per cent. From this point its weight-curve declines steadily to a minimum of 4.8 per cent at term. The archipallium at 4.3 mm. weighed 4.2 per cent, twice the initial weight-value of the neopallium, reaching a maximum of 18 per cent at 7 weeks, from which point the weight-curve declined steadily to 1.03 percent in the new-born. At 11 weeks the archipallium could be subdivided into the olfactory bulb, which weighed 0.3 per cent, and the paraterminal body, fornix, and hippocampus, which together weighed 3.22 per cent. Both of these units show a consistent shrinkage in relative weight until birth, when the former weighed 0.2 per cent and the latter 0.83 per cent.

Chart 1. Diagrammatic representation of tho percentage weights of the brain parts at different stages of development.
The diencephalon could also be isolated in all cases, and in the 4.3 mm. embryo had a weight-value of 24.3 per cent, or more than three times that of the telencephalon. From this high initial percentage the curve declined gradually to 2.63 per cent at term, notwithstanding the considerable increase in the size of the thalami.
The mesencephalon, weighing in the youngest specimen 14.3 per cent, drops steadily to 0.64 per cent at term, declining from a structure of considerable size to a mere slender, connecting stem, which attains its chief differentiation through the development of secondary centers connected with the fiber systems passing through it.
The total rhombencephalon starts with a maximum weight of 54.4 per cent of the whole in the 4.3 mm. specimen, from which point its curve sinks rapidly until at 156 mm. it reaches a minimum of 4.97 per cent, then rises gradually to 5.2G per cent at 230 mm., and 6.98 per cent at term. This unusual curve is explained when we study the growth-rate of the two component units of the rhombencephalon, which behave in a widely different manner. The curve for the cerebellum begins in the 16 mm. embryo at 7.56 per cent, ascends to 9.74 per cent at 8 weeks, its maximum point during prenatal growth. From this level it falls to 2.74 per cent at 13.5 weeks, remaining there until the end of the nineteenth week when it rises rapidly, reaching 6 per cent at term, its actual weight being 30.7 grams. This reversal of the usually observed conditions shows the increasing importance of the cerebellum at these stages of development, its individual growth-rate becoming sufficiently marked to maintain an unvarying level during six weeks of changing values in other parts. It then gains a new impetus which is sustained throughout of the remainder gestation. The medulla-pons in the 16 mm. embryo comprises 38.11 per cent of the weight of the entire encephalon. The curve falls rapidly to 3.73 per cent at 13.5 weeks, then more gradually to its minimum of 0.91 per cent at term, the general weight-curve for this part being comparable only to that of the diencephalon, though the relative loss is much greater in the medulla-pons. It would seem probable that the early large size of the medulla-pons is due to its association with the development of the cranial nerves which are relatively very large at this time.
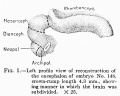
Fig 1
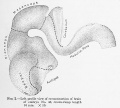
Fig 2
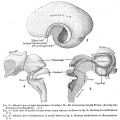
Fig 3-5
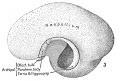
Fig 3
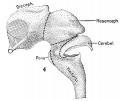
Fig 4
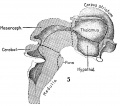
Fig 5
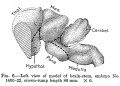
Fig 6
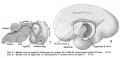
Fig 7-8
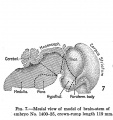
Fig 7
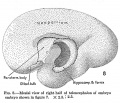
Fig 8
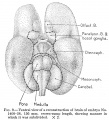
Fig 9
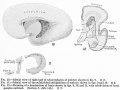
Fig 10-12
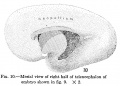
Fig 10

Fig 11
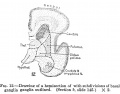
Fig 12

Table 1
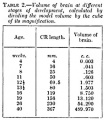
Table 2

Table 3

Chart 1








References
Bischoff, 1880. Das Hirngewieht dea Menschen. Bonn. Boyd, R., 1861. Tables of the weights of the human body and internal organs in the sane and insane of both sexes at various ages. Phil. Trans. Royal Soc. of London, vol. 151.
Dockeray, F. C, 1915. Volumetric determinations of the parts of the brain in a human fetus 150 mm. long (crown-rump). Anat. Roc, vol. 9.
Donaldson, H. H., 1895. The growth of the brain. London. ,
- 1908. A comparison of the albino rat with man in respect to the growth of the brain and spinal cord. Jour. Comp. Neur., vol. IS. , 1909. Relation of body-length to body-weight and to the weight of the brain and of the spinal cord in the albino rat. Ibid., vol. 19.
Hrdlicka, AleS, 1906. Brain and brain preservatives. Proe. U. S. Nat. Mus., vol. 30. , 1919. Anthropometry. Amer. Jour. Phys. Anthrop., vol. 2.
Jackson, C. M., 1909. On the prenatal growth of the various organs and parts. Amer. Jour. Anat., vol. 9.
Le Bon, G., 1879. Recherches anatomiques et mathferaatiques sur les lois des variations du volume de cerveau et sur leur relations avec I'intelligence. Rev. d'anthrop., Paris, vol. 2. Lewandowsky, 1910. Handbuch der Neurologic, vol. 1.
Mall FP. A list of normal human embryos which have been cut into serial sections. (1910) Anat. Rec. 4(10): 355-367.
Mall, F. P., 1910. Determination of the age of human embryos and fetuses. Manual of Human Embryology, Keibel & Mall, vol. 1. Manouveier, 1885. La quantity dans I'encaphalie. Paris.
Marchand, F., 1902. Ueber das Hirngewieht des Menschen. Abh. der math. phys. 01. d. K6nigl. Sachs. Gesellsch. d. Wiss., vol. 27. Marshall, John, 1892. The brain of the late George Grote, with comments and observations on the human brain and its parts generally. Jour. Anat. & Phys.. vol. 27.
Martin, 1914. Lehrbuch fur Anthropologic. Jena.
Matieoka, J., 190;j. The significance of the weight of the brain in man. Sitzungber. der Kon. koruischen Gesellsch. d. Wiss. Math. Natrwiss. Claaae Jahr., vol. 20.
Michaelis, P., 1907. Das Hirngewieht dea Kindea. Monatschr. f. Kinderh., vol. 6.
Pearl, Raymond, 1905. Some results of a study of variations and correlations in brain-weight. Jour. Comp. Neur., vol. 15. , 1905. Biometrical studies on man. Variation and correlation in brain weight. Biometrika, vol.4.
Retzius, G., 1896. Das Menschenhirn. Tafel 2. Stockholm. , 1900. Ueber das Gehirngewicht der Schweden. Biol. Untersuch., N. F., vol. 9.
Spitzka, E. A., 1907. A study of the brains of six eminent scientists and scholars belonging to the American Anthropometric Society. Amer. Phil. Soc, n. 3., vol. 21, part 3.
Streeter, G. L., 1920. Weight, sitting-height, headsize, foot-length, and menstrual age in the human embryo. Contritubions to Embryology, vol. 10, Carnegie Inst. Wash. Pub. 274.
Vjerordt, 1890. Das Massenwachsthum der Korperorgane des Menschen. Arch. f. Anat. u. Phys., Anat. Abth., Supp. Bd. , 1893. Daten und Tabellen, 3 Aufl., Jena.
West, G. M., 1894. Anthropometrische Untersuchungcn liber die Schulkinder in Worcester, Mass. Arcliiv f. Anthrop., vol. 22.
Ziehen, 1899. Macroskopisohe Anatomie des Gehirna. Bardeleben's Handbuch d. Anat., vol. 4. , 1903. Gehirngewichte. Monatschr. f. Psych, u. Neurol., vol. 13.
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2024, April 28) Embryology Book - Contributions to Embryology Carnegie Institution No.59. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Book_-_Contributions_to_Embryology_Carnegie_Institution_No.59
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G