Endocrine - Hypothalamus Development: Difference between revisions
mNo edit summary |
|||
| (104 intermediate revisions by 2 users not shown) | |||
| Line 1: | Line 1: | ||
{{Header}} | |||
==Introduction== | ==Introduction== | ||
[[File:Stage_22_image_055.jpg|thumb|Human embryonic hypothalamus ([[Carnegie_stage_22|Stage 22]])]] | [[File:Stage_22_image_055.jpg|thumb|Human embryonic hypothalamus ([[Carnegie_stage_22|Stage 22]])]] | ||
[[File:Adult diencephalon.jpg|Adult hypothalamus position]] | [[File:Adult diencephalon.jpg|thumb|Adult hypothalamus position]] | ||
The hypothalamus has a key role in regulating the nearby pituitary and peripheral endocrine organ functions. This neural region therefore is associated with hormonally related behaviour (anger and sexual activity), homeostatic regulation (blood pressure, heart rate, appetite, and temperature) and functions that relate to both (puberty, reproductive cycles, and lactation). | The {{hypothalamus}} has a key role in regulating the nearby pituitary and peripheral endocrine organ functions. This structure should though be considered as a {{neural}} region therefore is associated with hormonally related behaviour (anger and sexual activity), homeostatic regulation (blood pressure, heart rate, appetite, and temperature) and functions that relate to both (puberty, reproductive cycles, and lactation). | ||
In the early embryo, neuroectoderm of the forebrain (prosenecephalon) primary brain vesicle divides to form two secondary brain vesicles, telencephalon (endbrain, cortex) and diencephalon. | In the early embryo, neuroectoderm of the forebrain (prosenecephalon) primary brain vesicle divides to form two secondary brain vesicles, telencephalon (endbrain, cortex) and diencephalon. The historic description is that the diencephalon ventro-lateral wall, intermediate zone proliferation generates the primordial hypothalamus. A recent "Prosomeric model"{{#pmid:7939711|PMID7939711}}{{#pmid:25852489|PMID25852489}}, based on gene expression in the mouse, suggests an overall different origin of the hypothalamus arising from the secondary prosencephalon (telencephalon} and consists of an alar and basal region. The basal hypothalamus is further subdivided into tuberal and mamillary regions, each with distinct subregions. | ||
Hypothalamus development also occurs differentially in male and female embryos, described as part of neural "sexual dimorphism". | |||
In the adult, large neurosecretory cells of the supraoptic nucleus (SON) and the paraventricular nucleus (PVN) project to the {{neurohypophysis}} from the hypothalamus. Other key nuclei within the hypothalamus include the sexually dimorphic nucleus (SDN, intermediate nucleus, INAH-1), suprachiasmatic nucleus (SCN) and tuberal lateral nucleus (NTL). Anatomically surrounded by the anterior commissure and lamina terminalis anteriorly, the mammillary bodies and midbrain posteriorly, and thalamus superiorly. | |||
{{Endocrine Links}} | |||
[[:Category:Hypothalamus|Category:Hypothalamus]] | [[Neural System Development]] | [[Lecture - Ectoderm Development]] | [[Lecture - Neural Development]] | [[Lecture - Head Development]] | |||
==Some Recent Findings== | ==Some Recent Findings== | ||
[[File:Hypothalamus model 01.jpg|thumb|alt=Hypothalamus model|Hypothalamus model{{#pmid:25852489|PMID25852489}}]] | |||
[[File:Adult mouse brain - prosomeric model.jpg|thumb|alt=Adult mouse brain - prosomeric model|Adult mouse brain - prosomeric model{{#pmid:25852489|PMID25852489}}]] | |||
{| | {| | ||
|-bgcolor="F5FAFF" | |-bgcolor="F5FAFF" | ||
| | | | ||
* '''Embryonic gonadotropin-releasing hormone signaling is necessary for maturation of the male reproductive axis.''' | * '''Conserved Genoarchitecture of the Basal Hypothalamus in {{Zebrafish}} Embryos'''{{#pmid:32116574|PMID32116574}} "Analyses of genoarchitecture recently stimulated substantial revisions of anatomical models for the developing {{hypothalamus}} in mammalian and other vertebrate systems. The prosomeric model proposes the hypothalamus to be derived from the secondary prosencephalon, and to consist of alar and basal regions. The basal hypothalamus can further be subdivided into tuberal and mamillary regions, each with distinct subregions. ...Our comparison of gene expression patterns reveals that the genoarchitecture of the basal hypothalamus in {{zebrafish}} embryos 48 hours post fertilization is highly similar to {{mouse}} embryos at {{ME13.5}}. We found the tuberal hypothalamus in zebrafish embryos to be relatively large and to comprise previously ill-defined regions around the posterior hypothalamic recess. The mamillary hypothalamus is smaller and concentrates to rather medial areas in proximity to the anterior end of the neural tube floor plate. Within the basal hypothalamus we identified longitudinal and transverse tuberal and mamillary subregions topologically equivalent to those previously described in other vertebrates. However, the hypothalamic diencephalic boundary region and the posterior tuberculum still provide a challenge." | ||
* '''Development of the Basal Hypothalamus through Anisotropic Growth'''{{#pmid:31050853|PMID31050853}} "The adult hypothalamus is subdivided into distinct domains: pre-optic, anterior, tuberal and mammillary. Each domain harbours an array of neurons that act together to regulate homeostasis. The embryonic origins and development of hypothalamic neurons, however, remains enigmatic. Here we summarise recent studies in model organisms that challenge current views of hypothalamic development, which traditionally have attempted to map adult domains to correspondingly-located embryonic domains that expand isotropically. Instead, new studies indicate that hypothalamic neurons arise from progenitor cells that undergo anisotropic growth in different dimensions. Here we describe how a multipotent {{Shh}}/ {{Fgf}}10-expressing progenitor population gives rise to progenitors that grow anisotropically, expanding to a greater extent than other progenitors and giving rise to cells throughout the basal hypothalamus. Further, Shh/Fgf10+ive -derived progenitors grow sequentially in different directions from the multipotent Shh/ Fgf10 population: first, a subset displaced rostrally give rise to anterior-ventral/tuberal neuronal progenitors, then a subset displaced caudally give rise to mammillary neuronal progenitors; finally, a subset(s) displaced ventrally give rise to tuberal infundibular glial progenitors. As this occurs, stable populations of Shh+ive and Fgf10+ive progenitors form. We describe current understanding of the mechanisms that induce Shh/Fgf10+ive progenitors, and begin to direct their differentiation to anterior-ventral/tuberal neuronal progenitors, mammillary neuronal progenitors and tuberal infundibular progenitors. Together these studies suggest a new model for hypothalamic development that we term the Anisotropic growth model." | |||
* '''Anatomy, development, and plasticity of the neurosecretory hypothalamus in {{zebrafish}}'''{{#pmid:30109407|PMID30109407}} "The paraventricular nucleus (PVN) of the hypothalamus harbors diverse neurosecretory cells with critical physiological roles for the homeostasis. Decades of research in rodents have provided a large amount of information on the anatomy, development, and function of this important hypothalamic nucleus. However, since the hypothalamus lies deep within the brain in mammals and is difficult to access, many questions regarding development and plasticity of this nucleus still remain. In particular, how different environmental conditions, including stress exposure, shape the development of this important nucleus has been difficult to address in animals that develop in utero. To address these open questions, the transparent larval zebrafish with its rapid external development and excellent genetic toolbox offers exciting opportunities. In this review, we summarize recent information on the anatomy and development of the neurosecretory preoptic area (NPO), which represents a similar structure to the mammalian PVN in zebrafish. We will then review recent studies on the development of different cell types in the neurosecretory hypothalamus both in mouse and in fish. Lastly, we discuss stress-induced plasticity of the PVN mainly discussing the data obtained in rodents, but pointing out tools and approaches available in zebrafish for future studies. This review serves as a primer for the currently available information relevant for studying the development and plasticity of this important brain region using zebrafish." | |||
* '''Expression of progesterone receptor, estrogen receptors α and β, and kisspeptin in the hypothalamus during perinatal development of gonad-lacking steroidogenic factor-1 knockout mice'''{{#pmid:30776325|PMID30776325}} "Gonadal hormones contribute to brain sexual differentiation. We analyzed expression of {{progesterone}} receptor (PR), estrogen receptor-α (ERα), ERβ, and {{kisspeptin}}, in the preoptic area (POA) and/or the arcuate nucleus (ARC), in gonad-lacking steroidogenic factor-1 knockout (KO) mice during perinatal development. At postnatal-day (P) 0-P7, POA PR levels were higher in wild-type (WT) males compared with WT females, while those in KO males were lower than in WT males and similar to those in WT and KO females. At P14-P21, PR levels in all groups increased similarly. POA ERα levels were similar in all groups at embryonic-day (E) {{ME15.5}}-P14. Those in WT but not KO males reduced during postnatal development to be significantly lower compared with females at P21. POA ERβ levels were higher in WT males than in WT females, while those in KO males were lower than in WT males and similar to those in WT and KO females at P0-P21. POA kisspeptin expression was female-biased in WT mice, while levels in KO females were lower compared with WT females and similar to those in WT and KO males. ARC kisspeptin levels were equivalent among groups at {{ME15.5}}-P0. At P7-P21, ARC levels in WT but not KO males became lower compared with WT females. Diethylstilbestrol exposure during P0-P6 and P7-P13 increased POA PR and ERβ, and decreased POA ERα and ARC kisspeptin levels at P7 and/or P14 in both sexes of KO mice. These data further understanding of gonadal hormone action on neuronal marker expression during brain sexual development." | |||
|} | |||
{| class="wikitable mw-collapsible mw-collapsed" | |||
! More recent papers | |||
|- | |||
| [[File:Mark_Hill.jpg|90px|left]] {{Most_Recent_Refs}} | |||
Search term: [http://www.ncbi.nlm.nih.gov/pubmed/?term=Hypothalamus+Development ''Hypothalamus Development''] | [http://www.ncbi.nlm.nih.gov/pubmed/?term=Hypothalamus+Embryology ''Hypothalamus Embryology''] | [http://www.ncbi.nlm.nih.gov/pubmed/?term=Supraoptic+Nucleus ''Supraoptic Nucleus''] | [http://www.ncbi.nlm.nih.gov/pubmed/?term=Paraventricular+Nucleus ''Paraventricular Nucleus''] | [http://www.ncbi.nlm.nih.gov/pubmed/?term=Suprachiasmatic+Nucleus ''Suprachiasmatic Nucleus''] | [http://www.ncbi.nlm.nih.gov/pubmed/?term=Tuberal+Lateral+Nucleus ''Tuberal Lateral Nucleus''] | |||
|} | |||
{| class="wikitable mw-collapsible mw-collapsed" | |||
! Older papers | |||
|- | |||
| {{Older papers}} | |||
* '''Novel insights into the spatial and temporal complexity of hypothalamic organization through precision methods allowing nanoscale resolution'''{{#pmid:30027599|PMID30027599}} "The mammalian hypothalamus contains an astounding heterogeneity of neurons to achieve its role in coordinating central responses to virtually any environmental stressor over the life-span of an individual. ... Mechanistic data encompass cell identities, synaptic connectivity within and outside the hypothalamus to link vegetative and conscious levels of innate behaviours, and context- and circadian rhythm-dependent rules of synaptic neurophysiology to distinguish hypothalamic foci that either tune the body's metabolic set-point or specify behaviours. Consequently, novel insights emerge to explain the evolutionary advantages of non-laminar organization for neuroendocrine circuits coincidently using fast neurotransmitters and neuropeptides." | |||
* '''Regulation of feeding by somatostatin neurons in the tuberal nucleus'''{{#pmid:29976824|PMID29976824}} "The tuberal nucleus (TN) is a surprisingly understudied brain region. We found that somatostatin (SST) neurons in the TN, which is known to exhibit pathological or cytological changes in human neurodegenerative diseases, play a crucial role in regulating feeding in mice. GABAergic tuberal SST (TNSST) neurons were activated by hunger and by the hunger hormone, ghrelin. Activation of TNSST neurons promoted feeding, whereas inhibition reduced it via projections to the paraventricular nucleus and bed nucleus of the stria terminalis. Ablation of TNSST neurons reduced body weight gain and food intake. These findings reveal a previously unknown mechanism of feeding regulation that operates through orexigenic TNSST neurons, providing a new perspective for understanding appetite changes." | |||
* '''Development of the hypothalamus: conservation, modification and innovation'''{{#pmid:28465334|PMID28465334}} "The hypothalamus, which regulates fundamental aspects of physiological homeostasis and behavior, is a brain region that exhibits highly conserved anatomy across vertebrate species. Its development involves conserved basic mechanisms of induction and patterning, combined with a more plastic process of neuronal fate specification, to produce brain circuits that mediate physiology and behavior according to the needs of each species. Here, we review the factors involved in the induction, patterning and neuronal differentiation of the hypothalamus, highlighting recent evidence that illustrates how changes in Wnt/β-catenin signaling during development may lead to species-specific form and function of this important brain structure." | |||
* '''A new scenario of hypothalamic organization: rationale of new hypotheses introduced in the updated prosomeric model'''{{#pmid:25852489|PMID25852489}} "In this essay, we aim to explore in depth the new concept of the hypothalamus that was presented in the updated prosomeric model. Initial sections deal with the antecedents of prosomeric ideas represented by the extensive literature centered on the alternative columnar model of Herrick (1910), Kuhlenbeck (1973) and Swanson (1992, 2003); a detailed critique explores why the columnar model is not helpful in the search for causal developmental explanations. In contrast, the emerging prosomeric scenario visibly includes many possibilities to propose causal explanations of hypothalamic structure relative to both anteroposterior and dorsoventral patterning mechanisms, and insures the possibility to compare hypothalamic histogenesis with that of more caudal parts of the brain. Next the four major changes introduced in the organization of the hypothalamus on occasion of the updated model are presented, and our rationale for these changes is explored in detail. It is hoped that this example of morphological theoretical analysis may be useful for readers interested in brain models, or in understanding why models may need to change in the quest for higher consistency." (See also editorial{{#pmid:26157363|PMID26157363}}) | |||
* '''Neuropeptide Signaling Differentially Affects Phase Maintenance and Rhythm Generation in SCN and Extra-SCN Circadian Oscillators'''{{#pmid:21559484|PMID21559484}} "Circadian rhythms in physiology and behavior are coordinated by the brain's dominant circadian pacemaker located in the suprachiasmatic nuclei (SCN) of the hypothalamus. Vasoactive intestinal polypeptide (VIP) and its receptor, VPAC(2), play important roles in the functioning of the SCN pacemaker. ... Further, our data indicate a link between circadian rhythm strength and the ability of tissues to resist circadian phase resetting." | |||
* '''Embryonic gonadotropin-releasing hormone signaling is necessary for maturation of the male reproductive axis.'''{{#pmid:20805495|PMID20805495}} "Surprisingly, genetic ablation (in mice) of GnRHR expressing cells significantly increased the number of GnRH neurons in the anterior hypothalamus, suggesting an unexpected role of GnRH signaling in establishing the size of the GnRH neuronal population. | |||
|} | |||
==Hypothalamus Endocrine Axes== | |||
===HPG Axis=== | |||
'''H'''ypothalamus - '''P'''ituitary - '''G'''onad endocrine axis. | |||
{| | |||
| [[File:XXhpgaxis.jpg|200px]] | |||
| This cartoon shows the '''H'''ypothalamus - '''P'''ituitary - '''G'''onad (female) endocrine axis. | |||
The arrows indicate how each of these endocrine organs interact and regulate each others secretions through endocrine feedback mechanisms. | |||
|} | |||
<gallery> | |||
File:HPG female axis.jpg|Female | |||
File:HPG male axis.jpg|Male | |||
</gallery> | |||
===Other Endocrine Axes=== | |||
{| | |||
| [[File:HPA_axis.jpg|300px]] | |||
'''H'''ypothalamus - '''P'''ituitary - '''A'''drenal endocrine axis. | |||
| [[File:HPT axis.jpg|300px]] | |||
'''H'''ypothalamus - '''P'''ituitary - '''T'''hyroid endocrine axis. | |||
|} | |} | ||
==Development Overview== | |||
{| | {| | ||
|- | |- | ||
| Line 36: | Line 92: | ||
| [[File:Gray0654.jpg|200px]] | | [[File:Gray0654.jpg|200px]] | ||
|- | |- | ||
| Diencephalon region<br>([[Carnegie_stage_13|Stage 13]]) | | [[#Secondary Brain Vesicles - Stage 13|Diencephalon region]], shown by [[O#optic stalk|optic stalk]]<br>([[Carnegie_stage_13|Stage 13]]) | ||
| | | [[#Embryo Brain - Stage 22|Late embryonic hypothalamus]]<br>([[Carnegie_stage_22|Stage 22]]) | ||
| Early fetal human brain<br>([[Fetal Development|3 months]], from a model by [[Embryology_History_-_Wilhelm_His|Wilhelm His]]) | | [[#Early Fetal Brain - Week 10|Early fetal human brain]]<br>([[Fetal Development|3 months]], from a model by [[Embryology_History_-_Wilhelm_His|Wilhelm His]]) | ||
|} | |} | ||
* Neuroectoderm - prosenecephalon then diencephalon | |||
* Neuroectoderm - prosenecephalon then rostral diencephalon after induction by the underlying prechordal plate. | |||
** Sonic hedgehog (Shh) - initially expressed in prechordal plate, is essential for inductive process. | |||
* ventro-lateral wall intermediate zone proliferation | * ventro-lateral wall intermediate zone proliferation | ||
* Mamillary bodies - form pea-sized swellings ventral wall of hypothalamus | * Mamillary bodies - form pea-sized swellings ventral wall of hypothalamus | ||
=== | ===Week 5=== | ||
{| | |||
|- | Secondary Brain Vesicles - Stage 13 | ||
{| | |||
|+'''[[Carnegie stage 13 - serial sections|Stage 13]]''' ''serial labeled images'' | |||
| [[File:Stage 13 image 057.jpg|120px]] | |||
| [[File:Stage 13 image 058.jpg|120px]] | |||
| [[File:Stage 13 image 059.jpg|120px]] | |||
| [[File:Stage 13 image 060.jpg|120px]] | |||
| [[File:Stage 13 image 061.jpg|120px]] | |||
| [[File:Stage 13 image 062.jpg|120px]] | |||
| [[File:Stage 13 image 063.jpg|120px]] | |||
|- | |- | ||
| [[:File:Stage 13 image 057.jpg|B1L]] | |||
| | | [[:File:Stage 13 image 058.jpg|B2L]] | ||
| | | [[:File:Stage 13 image 059.jpg|B3L]] | ||
| [[:File:Stage 13 image 060.jpg|B4L]] | |||
| | | [[:File:Stage 13 image 061.jpg|B5L]] | ||
| [[:File:Stage 13 image 062.jpg|B6L]] | |||
| [[:File:Stage 13 image 063.jpg|B7L]] | |||
|} | |||
===Week 8=== | |||
Embryo Brain - Stage 22 | |||
{| | |||
|+'''[[Carnegie stage_22 - serial sections|Stage 22L]]''' ''serial labeled images'' | |||
| [[File:Stage_22 image 050.jpg|120px]] | |||
| [[File:Stage_22 image 051.jpg|120px]] | |||
| [[File:Stage_22 image 052.jpg|120px]] | |||
| [[File:Stage_22 image 053.jpg|120px]] | |||
| [[File:Stage_22 image 054.jpg|120px]] | |||
| [[File:Stage_22 image 055.jpg|120px]] | |||
| [[File:Stage_22 image 056.jpg|120px]] | |||
|- | |- | ||
| [[:File:Stage_22 image 050.jpg|A1L]] | |||
| | | [[:File:Stage_22 image 051.jpg|A2L]] | ||
| | | [[:File:Stage_22 image 052.jpg|A3L]] | ||
| [[:File:Stage_22 image 053.jpg|A4L]] | |||
| | | [[:File:Stage_22 image 054.jpg|A5L]] | ||
| [[:File:Stage_22 image 055.jpg|A6L]] | |||
| [[:File:Stage_22 image 056.jpg|A7L]] | |||
|- | |- | ||
| [[File:Stage_22 image 057.jpg|120px]] | |||
| | | [[File:Stage_22 image 058.jpg|120px]] | ||
| | | [[File:Stage_22 image 059.jpg|120px]] | ||
| [[File:Stage_22 image 060.jpg|120px]] | |||
| | | [[File:Stage_22 image 061.jpg|120px]] | ||
| [[File:Stage_22 image 062.jpg|120px]] | |||
| [[File:Stage_22 image 063.jpg|120px]] | |||
|- | |- | ||
| [[:File:Stage_22 image 057.jpg|B1L]] | |||
| | | [[:File:Stage_22 image 058.jpg|B2L]] | ||
| | | [[:File:Stage_22 image 059.jpg|B3L]] | ||
| [[:File:Stage_22 image 060.jpg|B4L]] | |||
| | | [[:File:Stage_22 image 061.jpg|B5L]] | ||
| [[:File:Stage_22 image 062.jpg|B6L]] | |||
| [[:File:Stage_22 image 063.jpg|B7L]] | |||
|} | |} | ||
===Week 10=== | |||
Early Fetal Brain | |||
{| | |||
|+'''[[Fetal_Development_-_10_Weeks|10 Week Fetus]]''' ''head images'' | |||
| [[File:Human-_fetal_week_10_head_A.jpg|200px]] | |||
| [[File:Human-_fetal_week_10_head_B.jpg|200px]] | |||
| [[File:Human-_fetal_week_10_head_C.jpg|200px]] | |||
| [[File:Human-_fetal_week_10_head_D.jpg|200px]] | |||
|} | |||
<gallery> | |||
File:Gray0654.jpg|Human Fetal Brain <br>(3 months) | |||
File:Gray0655.jpg|Human Fetal Brain <br>(4 months) | |||
</gallery> | |||
==Adult Hypothalamus Hormones== | |||
==Sexually Dimorphic Nucleus== | [[File:Adult_human_hypothalamus_02.jpg|600px]] | ||
Adult Human Hypothalamus{{#pmid:23682342|PMID23682342}} | |||
{{Adult Hypothalamus Hormones table}} | |||
'''Hormones''' - Thyrotrophin releasing hormone (TRH), Corticotrophin releasing hormone (CRH), Arginine vasopressin (AVP), Gonadotrophin releasing hormone (GnRH), Growth hormone releasing hormone (GHRH), Somatostatin, Prolactin relasing factor (PRF), Dopamine | |||
Growth hormone (GH) secretion from the pituitary is controlled in many different ways, including positive/negative regulation of synthesis and release by the hypothalamus. Hypothalamic GH releasing hormone (GHRH) activates and somatostatin suppresses growth hormone synthesis and release. | |||
==Hypothalamic Nuclei== | |||
===Supraoptic Nucleus=== | |||
(SON) Arginine vasopressin (AVP) and oxytocin (OT) synthesis and released in the posterior pituitary. These 2 hormones differ only in 2 amino acids in positions 3 and 8. | |||
'''Arginine vasopressin''' (AVP) peptide hormone (9aa, nonapeptide) | |||
* synthesized as inactive preprohormone | |||
* regulates blood volume and pressure acting on the kidney and heart | |||
* Increased osmolality, due to blood volume reduction, increases AVP secretion. | |||
'''Oxytocin''' (OT) peptide hormone (9aa, nonapeptide) | |||
* mainly produced in hypothalamic magnocellular neurons | |||
* regulates female reproductive pregnancy-associated changes (stimulation of milk ejection, uterine contractions) | |||
* acts on one type of OT receptor (OTR) | |||
* hormone is a non-glycosylated protein undergoes an initial cleavage by the convertase magnolysin (EC 3.4.24.62) to OT-Gly-Lys-Arg (OT-GKR). | |||
* additional processing produces other OT extended molecules: OT-Gly-Lys (OT-GK) and OT-Gly (OT-G, OT-X). | |||
* OT-G is converted by an α-amidating enzyme to C-amidated nonapeptide which is released into the circulation | |||
* may have a role in fetal heart growth{{#pmid:15316117|PMID15316117}} and in vascular tone, regrowth and remodeling.{{#pmid:10811917|PMID10811917}} | |||
{{Oxytocin table}} | |||
Abnormalities: diabetes insipidus (vasopressin deficiency), syndrome of inappropriate antidiuresis (vasopressin excess) | |||
:'''Links:''' [[Birth]] | [http://www.ncbi.nlm.nih.gov/books?term=Supraoptic%20Nucleus Search Bookshelf] | |||
===Paraventricular Nucleus=== | |||
[[File:Hypothalamus histology 001.jpg|thumb|Hypothalamus PVN and SCN]] | |||
[[File:Oxytocin receptor pathways.jpg|thumb|Oxytocin receptor pathways]] | |||
(PVN) Arginine vasopressin (AVP) and oxytocin synthesis and released in the posterior pituitary. These 2 hormones differ only in 2 amino acids in positions 3 and 8. | |||
* magnocellular neurons | |||
* parvocellular neurosecretory neurons project axons to the pituitary median eminence | |||
* interneurons and neurons also project centrally | |||
'''Arginine vasopressin''' (AVP) peptide hormone (9aa, nonapeptide) | |||
* synthesized as inactive preprohormone | |||
* regulates blood volume and pressure acting on the kidney and heart | |||
* Increased osmolality, due to blood volume reduction, increases AVP secretion. | |||
'''Oxytocin''' (OT) peptide hormone (9aa, nonapeptide) | |||
* mainly produced in hypothalamic magnocellular neurons | |||
* regulates female reproductive pregnancy-associated changes | |||
* central effect on social and other behaviors, release from magnocellular dendrites and axonal projections of parvocellular neurons | |||
* acts on one type of OT receptor (OTR) | |||
* hormone is a non-glycosylated protein undergoes an initial cleavage by the convertase magnolysin (EC 3.4.24.62) to OT-Gly-Lys-Arg (OT-GKR). | |||
* additional processing produces other OT extended molecules: OT-Gly-Lys (OT-GK) and OT-Gly (OT-G, OT-X). | |||
* OT-G is converted by an α-amidating enzyme to C-amidated nonapeptide which is released into the circulation | |||
* may have a role in fetal heart growth{{#pmid:15316117|PMID15316117}} | |||
:'''Links:''' [[Birth]] | [http://www.ncbi.nlm.nih.gov/books?term=Paraventricular%20Nucleus Search Bookshelf] | |||
===Sexually Dimorphic Nucleus=== | |||
(SDN, intermediate nucleus, INAH-1) | (SDN, intermediate nucleus, INAH-1) | ||
==Suprachiasmatic Nucleus== | * Twice as large in young male adults as in young females. | ||
* At birth 20% of the adult SDN cell number is present, from then until 2-4 years of age cell numbers increase equally rapidly in both sexes. | |||
* After this age cell numbers start to decrease in girls, creating the sex difference. | |||
The SCN is the clock of the brain and shows circadian and seasonal fluctuations in vasopressin-expressing cell numbers. (SDN and SCN text modified from: Swaab, 1995) | |||
:'''Links''': [http://www.ncbi.nlm.nih.gov/books?term=Sexually%20Dimorphic%20Nucleus Search Bookshelf] | |||
===Suprachiasmatic Nucleus=== | |||
[[File:Hypothalamus histology 001.jpg|thumb|Hypothalamus PVN and SCN]] | |||
(SCN) This nucleus is the clock of the brain and shows circadian and seasonal fluctuations in vasopressin-expressing cell numbers. | (SCN) This nucleus is the clock of the brain and shows circadian and seasonal fluctuations in vasopressin-expressing cell numbers. | ||
:'''Links''': [http://www.ncbi.nlm.nih.gov/books?term=Suprachiasmatic%20Nucleus Search Bookshelf] | |||
===Tuberal Hypothalamus=== | |||
The tuberal hypothalamus consists of the dorsomedial, ventromedial, arcuate nuclei, and parts of the lateral hypothalamic area. In mouse model, Neurog2 shown to regulate the temporal progression.<ref>[https://www.jneurosci.org/content/40/18/3549.abstract?etoc Neurog2 Acts as a Classical Proneural Gene in the Ventromedial Hypothalamus and Is Required for the Early Phase of Neurogenesis] Journal of Neuroscience 29 April 2020, 40 (18) 3549-3563; DOI: https://doi.org/10.1523/JNEUROSCI.2610-19.2020</ref> | |||
'''Mouse Neurogenesis Timeline''' | |||
{{ME9.5}} - begins lateral hypothalamic and arcuate, rostrocaudal progression | |||
{{ME10.5}} - dorsomedial and ventromedial neurons, outside-in progression | |||
{{ME11.5}} - peaking throughout the region | |||
:'''Links''': [http://www.ncbi.nlm.nih.gov/books?term=Tuberal%20Lateral%20Nucleus Search Bookshelf] | |||
(NTL) This nucleus is involved in feeding behavior and energy metabolism. | |||
==Tracts== | |||
* '''retino-hypothalamus tract''' - (RHT)is the main connection between the eye and the hypothalamus, acts as a direct afferent pathway from the retina going through the optic chiasma. | |||
==Molecular== | |||
{| | |||
! Hypothalamus prosomeric model{{#pmid:25852489|PMID25852489} | |||
|- | |||
| [[File:Hypothalamus model 01.jpg|600px|alt=Hypothalamus model]] | |||
| Apparent sources of patterning diffusible morphogens that may have effects on the {{hypothalamus}}. | |||
Anterior neural ridge (ANR; yellow), which releases {{FGF}}8 is in fact a part of the {{roof plate}} (dorsalizing influence), rather than a source of AP effects. | |||
in contrast, the retromamillary and mamillary {{floor plate}} (dark blue associated to RM and M) releases {{SHH}} (ventralizing influence; note Shh secondarily also is expressed throughout the basal plate, and is later downregulated at the Tu area). | |||
The acroterminal midline as a source of AP patterning effects. Recent observations show {{Fgf}}18 expression within the postulated alar acroterminal organizer (fuchsia-labeled) and {{Fgf}}8 and {{Fgf}}10 expression within the postulated basal acroterminal organizer (green-labeled). There also are bilateral spots of Fgf8 expression at the optic stalks (not shown). (text modified from original figure legend) | |||
|} | |||
Hypothalamus Development Gene Interaction Model{{#pmid:24360028|PMID24360028}} | |||
[[File:Hypothalamus gene interaction model.jpg|500px]] | |||
Transcription factor cascade ([https://www.omim.org/entry/603128 Sim1], [https://www.omim.org/entry/604529 Otp], [https://www.omim.org/entry/600892 Sim2], and [https://www.omim.org/entry/600494 Brn2]) regulate differentiation of the anterior hypothalamus neuroendocrine cells. | |||
:'''Links:''' {{SHH}} | {{FGF}} | {{Notch}} | {{prosomere}} | |||
== References == | == References == | ||
| Line 95: | Line 315: | ||
===Reviews=== | ===Reviews=== | ||
{{#pmid:30109407}} | |||
{{#pmid:28465334}} | |||
{{#pmid:25589270}} | |||
{{#pmid:24290347}} | |||
{{#pmid:20626426}} | |||
{{#pmid:12127306}} | |||
===Articles=== | ===Articles=== | ||
{{#pmid:20637232}} | |||
{{#pmid:20628204}} | |||
{{#pmid:15840737}} | |||
===Books=== | |||
Endocrinology - An Integrated Approach Stephen Nussey and Saffron Whitehead St. George's Hospital Medical School, London, UK Oxford: BIOS Scientific Publishers; 2001. ISBN-10: 1-85996-252-1 | |||
* [http://www.ncbi.nlm.nih.gov/books/NBK26/#A516 Hypothalamic control of adrenocortical steroid synthesis - CRH and vasopressin] | |||
* [http://www.ncbi.nlm.nih.gov/books/NBK27/#A1267 Anatomical and functional connections of the hypothalamo-pituitary axis] | |||
* [http://www.ncbi.nlm.nih.gov/books/NBK27/#A1297 Blood supply of the hypothalamo-pituitary axis] | |||
* [http://www.ncbi.nlm.nih.gov/books/NBK27/#A1457 Autonomic functions of the hypothalamus] | |||
===Historic=== | |||
{{#pmid:28265842}} | |||
===Search PubMed=== | ===Search PubMed=== | ||
| Line 107: | Line 349: | ||
==Additional Images== | ==Additional Images== | ||
<gallery> | |||
File:Mouse hypothalamus SHH expression 01.jpg|Mouse hypothalamus SHH expression | |||
File:Adult human hypothalamus 02.jpg | |||
File:Adult human hypothalamus 03.jpg | |||
File:Adult human hypothalamus 04.jpg | |||
</gallery> | |||
==Terms== | |||
* '''3ß-diol''' - 5alpha-androstane-3beta, 17beta-diol | |||
* '''3α-diol''' - 5alpha-androstane-3alpha, 17beta-diol | |||
* '''5-HT''' - serotonin | |||
* '''5αR''' - 5-alpha-reductase | |||
* '''ACTH''' - adrenocortcotropin releasing hormone | |||
* '''AR''' - androgen receptor | |||
* '''ARN''' - (AR) Arcuate nucleus | |||
* '''AVP''' - arginine vasopressin | |||
* '''BnST''' - bed nucleus of the stria terminalis | |||
* '''BSA''' - bovine serum albumin | |||
* '''CBP''' - CREB binding protein | |||
* '''CORT''' - corticosterone | |||
* '''CRE''' - cyclic adenosine monophosphate response element | |||
* '''CREB''' - CRE binding protein | |||
* '''CRF''' - corticotropin releasing factor | |||
* '''DES''' - diethylstilbesterol | |||
* '''DEX''' - dexamethasone | |||
* '''DHT''' - dihydrotestosterone | |||
* '''DMN''' - (DM) Dorsomedial hypothalamic nucleus | |||
* '''DPN''' - diarylproprionitrile | |||
* '''ER''' - estrogen receptor | |||
* '''ERE''' - estrogen response element | |||
* '''FSH''' - follicle stimulating hormone | |||
* '''FSL''' - flinders sensitive line | |||
* '''GABA''' - gamma-aminobutyric acid | |||
* '''GAS''' - general adaptation syndrome | |||
* '''GH''' - growth hormone | |||
* '''GLP-1''' - glucagon-like peptide 1 | |||
* '''GnRH''' - gonadotropin releasing hormone | |||
* '''GPER''' - G-protein coupled estrogen receptor | |||
* '''GR''' - glucocorticoid receptor | |||
* '''HPA''' - hypothalamo–pituitary–adrenal | |||
* '''HPG''' - hypothalamo–pituitary–gonadal | |||
* '''HRE''' - hormone respone element | |||
* '''HSD''' - hydroxysteroid dehydrogenase | |||
* '''ICV''' - intracerebroventricular | |||
* '''ir''' - immunoreactive | |||
* '''ISH''' - in situ hybridization | |||
* '''LH''' - luteinizing hormone | |||
* '''LHRH''' - luteinizing hormone releasing hormone | |||
* '''median eminence''' - part of the inferior boundary of the hypothalamus | |||
* '''MPOA''' - medial preoptic area | |||
* '''MR''' - mineralocorticoid receptor | |||
* '''NTS''' - nucleus of the solitary tract | |||
* '''PPT''' - propylpyrazoletriol | |||
* '''PR''' - progesterone receptor | |||
* '''PVN''' - (PVA, PVH) Paraventricular nucleus | |||
* '''RAR''' - retinoic acid receptors | |||
* '''SCN''' - (SC) Suprachiasmatic nucleus | |||
* '''SON''' - (SO) Supraoptic nucleus | |||
* '''SRC''' - steroid receptor coactivators | |||
* '''T''' - {{testosterone}} | |||
* '''THR''' - thyroid hormone receptors | |||
* '''TPH''' - tryptophan hydroxylase | |||
* '''TRH''' - thyrotropin releasing hormone | |||
* '''TSH''' - thyroid stimulating hormone | |||
* '''VMN''' - (VM) Ventromedial nucleus | |||
==External Links== | |||
{{External Links}} | |||
{{ | {{Glossary}} | ||
{{ | {{Footer}} | ||
[[Category:Endocrine]] [[Category:Hypothalamus]] | [[Category:Endocrine]] [[Category:Neural]][[Category:Hypothalamus]] | ||
Latest revision as of 09:50, 14 May 2020
| Embryology - 21 May 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Introduction


The hypothalamus has a key role in regulating the nearby pituitary and peripheral endocrine organ functions. This structure should though be considered as a neural region therefore is associated with hormonally related behaviour (anger and sexual activity), homeostatic regulation (blood pressure, heart rate, appetite, and temperature) and functions that relate to both (puberty, reproductive cycles, and lactation).
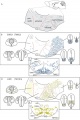
In the early embryo, neuroectoderm of the forebrain (prosenecephalon) primary brain vesicle divides to form two secondary brain vesicles, telencephalon (endbrain, cortex) and diencephalon. The historic description is that the diencephalon ventro-lateral wall, intermediate zone proliferation generates the primordial hypothalamus. A recent "Prosomeric model"[1][2], based on gene expression in the mouse, suggests an overall different origin of the hypothalamus arising from the secondary prosencephalon (telencephalon} and consists of an alar and basal region. The basal hypothalamus is further subdivided into tuberal and mamillary regions, each with distinct subregions.
Hypothalamus development also occurs differentially in male and female embryos, described as part of neural "sexual dimorphism".
In the adult, large neurosecretory cells of the supraoptic nucleus (SON) and the paraventricular nucleus (PVN) project to the neurohypophysis from the hypothalamus. Other key nuclei within the hypothalamus include the sexually dimorphic nucleus (SDN, intermediate nucleus, INAH-1), suprachiasmatic nucleus (SCN) and tuberal lateral nucleus (NTL). Anatomically surrounded by the anterior commissure and lamina terminalis anteriorly, the mammillary bodies and midbrain posteriorly, and thalamus superiorly.
Category:Hypothalamus | Neural System Development | Lecture - Ectoderm Development | Lecture - Neural Development | Lecture - Head Development
Some Recent Findings


|
| More recent papers |
|---|
 This table allows an automated computer search of the external PubMed database using the listed "Search term" text link.
More? References | Discussion Page | Journal Searches | 2019 References | 2020 References Search term: Hypothalamus Development | Hypothalamus Embryology | Supraoptic Nucleus | Paraventricular Nucleus | Suprachiasmatic Nucleus | Tuberal Lateral Nucleus |
| Older papers |
|---|
| These papers originally appeared in the Some Recent Findings table, but as that list grew in length have now been shuffled down to this collapsible table.
See also the Discussion Page for other references listed by year and References on this current page.
|
Hypothalamus Endocrine Axes
HPG Axis
Hypothalamus - Pituitary - Gonad endocrine axis.

|
This cartoon shows the Hypothalamus - Pituitary - Gonad (female) endocrine axis.
The arrows indicate how each of these endocrine organs interact and regulate each others secretions through endocrine feedback mechanisms. |
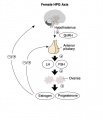
Female
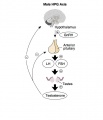
Male


Other Endocrine Axes

Hypothalamus - Pituitary - Adrenal endocrine axis. |

Hypothalamus - Pituitary - Thyroid endocrine axis. |
Development Overview

|
|

|
| Diencephalon region, shown by optic stalk (Stage 13) |
Late embryonic hypothalamus (Stage 22) |
Early fetal human brain (3 months, from a model by Wilhelm His) |
- Neuroectoderm - prosenecephalon then rostral diencephalon after induction by the underlying prechordal plate.
- Sonic hedgehog (Shh) - initially expressed in prechordal plate, is essential for inductive process.
- ventro-lateral wall intermediate zone proliferation
- Mamillary bodies - form pea-sized swellings ventral wall of hypothalamus
Week 5
Secondary Brain Vesicles - Stage 13

|

|

|

|
|

|

|
| B1L | B2L | B3L | B4L | B5L | B6L | B7L |
Week 8
Embryo Brain - Stage 22

|

|

|

|

|
|

|
| A1L | A2L | A3L | A4L | A5L | A6L | A7L |

|

|

|

|

|

|

|
| B1L | B2L | B3L | B4L | B5L | B6L | B7L |
Week 10
Early Fetal Brain

|

|

|

|
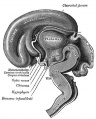
Human Fetal Brain
(3 months)
Human Fetal Brain
(4 months)

Adult Hypothalamus Hormones

Adult Human Hypothalamus[13]
| Secreted hormone | Abbreviation | Produced by | Effect |
|---|---|---|---|
| Thyrotropin-releasing hormone (Prolactin-releasing hormone) |
TRH, TRF, or PRH | Parvocellular neurosecretory neurons | thyroid-stimulating hormone (TSH) release from anterior pituitary (primarily) Stimulate prolactin release from anterior pituitary |
| Dopamine (Prolactin-inhibiting hormone) |
DA or PIH | Dopamine neurons of the arcuate nucleus | Inhibit prolactin release from anterior pituitary |
| Growth hormone-releasing hormone | GHRH | Neuroendocrine neurons of the Arcuate nucleus | Growth hormone (GH) release from anterior pituitary |
| Somatostatin (growth hormone-inhibiting hormone) |
SS, GHIH, or SRIF | Neuroendocrine cells of the Periventricular nucleus | Growth hormone (GH) release from anterior pituitary Inhibit Thyroid-stimulating hormone|thyroid-stimulating hormone (TSH) release from anterior pituitary |
| Gonadotropin-releasing hormone | GnRH or LHRH | Neuroendocrine cells of the Preoptic area | follicle-stimulating hormone (FSH) release from anterior pituitary Stimulate Luteinizing hormone|luteinizing hormone (LH) release from anterior pituitary |
| Corticotropin-releasing hormone | CRH or CRF | Parvocellular neurosecretory neurons | adrenocorticotropic hormone (ACTH) release from anterior pituitary |
| Oxytocin | Magnocellular neurosecretory cells | Lactation (letdown reflex) | |
| Vasopressin (antidiuretic hormone) |
ADH or AVP | Magnocellular neurosecretory neurons | Increases water permeability in the distal convoluted tubule and collecting duct of nephrons, thus promoting water reabsorption and increasing blood volume |
| Melanin-concentrating hormone | MCH | tuberal lateral nucleus (lateral hypothalamic area) | Neuropeptide with appetite stimulant (orexigenic) and sleep-promoting activities by projecting to a variety of brain areas. |
| Links: hypothalamus | |||
Hormones - Thyrotrophin releasing hormone (TRH), Corticotrophin releasing hormone (CRH), Arginine vasopressin (AVP), Gonadotrophin releasing hormone (GnRH), Growth hormone releasing hormone (GHRH), Somatostatin, Prolactin relasing factor (PRF), Dopamine
Growth hormone (GH) secretion from the pituitary is controlled in many different ways, including positive/negative regulation of synthesis and release by the hypothalamus. Hypothalamic GH releasing hormone (GHRH) activates and somatostatin suppresses growth hormone synthesis and release.
Hypothalamic Nuclei
Supraoptic Nucleus
(SON) Arginine vasopressin (AVP) and oxytocin (OT) synthesis and released in the posterior pituitary. These 2 hormones differ only in 2 amino acids in positions 3 and 8.
Arginine vasopressin (AVP) peptide hormone (9aa, nonapeptide)
- synthesized as inactive preprohormone
- regulates blood volume and pressure acting on the kidney and heart
- Increased osmolality, due to blood volume reduction, increases AVP secretion.
Oxytocin (OT) peptide hormone (9aa, nonapeptide)
- mainly produced in hypothalamic magnocellular neurons
- regulates female reproductive pregnancy-associated changes (stimulation of milk ejection, uterine contractions)
- acts on one type of OT receptor (OTR)
- hormone is a non-glycosylated protein undergoes an initial cleavage by the convertase magnolysin (EC 3.4.24.62) to OT-Gly-Lys-Arg (OT-GKR).
- additional processing produces other OT extended molecules: OT-Gly-Lys (OT-GK) and OT-Gly (OT-G, OT-X).
- OT-G is converted by an α-amidating enzyme to C-amidated nonapeptide which is released into the circulation
- may have a role in fetal heart growth[14] and in vascular tone, regrowth and remodeling.[15]
| About Oxytocin Hormone | |
|---|---|
| Structure | Function |
|
|
| Links: Birth | Hypothalamus | Endocrine | Mammary Gland | Milk | image - Oxytocin receptor pathways | |
Abnormalities: diabetes insipidus (vasopressin deficiency), syndrome of inappropriate antidiuresis (vasopressin excess)
- Links: Birth | Search Bookshelf
Paraventricular Nucleus


(PVN) Arginine vasopressin (AVP) and oxytocin synthesis and released in the posterior pituitary. These 2 hormones differ only in 2 amino acids in positions 3 and 8.
- magnocellular neurons
- parvocellular neurosecretory neurons project axons to the pituitary median eminence
- interneurons and neurons also project centrally
Arginine vasopressin (AVP) peptide hormone (9aa, nonapeptide)
- synthesized as inactive preprohormone
- regulates blood volume and pressure acting on the kidney and heart
- Increased osmolality, due to blood volume reduction, increases AVP secretion.
Oxytocin (OT) peptide hormone (9aa, nonapeptide)
- mainly produced in hypothalamic magnocellular neurons
- regulates female reproductive pregnancy-associated changes
- central effect on social and other behaviors, release from magnocellular dendrites and axonal projections of parvocellular neurons
- acts on one type of OT receptor (OTR)
- hormone is a non-glycosylated protein undergoes an initial cleavage by the convertase magnolysin (EC 3.4.24.62) to OT-Gly-Lys-Arg (OT-GKR).
- additional processing produces other OT extended molecules: OT-Gly-Lys (OT-GK) and OT-Gly (OT-G, OT-X).
- OT-G is converted by an α-amidating enzyme to C-amidated nonapeptide which is released into the circulation
- may have a role in fetal heart growth[14]
- Links: Birth | Search Bookshelf
Sexually Dimorphic Nucleus
(SDN, intermediate nucleus, INAH-1)
- Twice as large in young male adults as in young females.
- At birth 20% of the adult SDN cell number is present, from then until 2-4 years of age cell numbers increase equally rapidly in both sexes.
- After this age cell numbers start to decrease in girls, creating the sex difference.
The SCN is the clock of the brain and shows circadian and seasonal fluctuations in vasopressin-expressing cell numbers. (SDN and SCN text modified from: Swaab, 1995)
- Links: Search Bookshelf
Suprachiasmatic Nucleus
(SCN) This nucleus is the clock of the brain and shows circadian and seasonal fluctuations in vasopressin-expressing cell numbers.
- Links: Search Bookshelf
Tuberal Hypothalamus
The tuberal hypothalamus consists of the dorsomedial, ventromedial, arcuate nuclei, and parts of the lateral hypothalamic area. In mouse model, Neurog2 shown to regulate the temporal progression.[16]
Mouse Neurogenesis Timeline E9.5 - begins lateral hypothalamic and arcuate, rostrocaudal progression E10.5 - dorsomedial and ventromedial neurons, outside-in progression E11.5 - peaking throughout the region
- Links: Search Bookshelf
(NTL) This nucleus is involved in feeding behavior and energy metabolism.
Tracts
- retino-hypothalamus tract - (RHT)is the main connection between the eye and the hypothalamus, acts as a direct afferent pathway from the retina going through the optic chiasma.
Molecular
| PMID25852489} | |
|---|---|
|
|
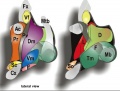
Apparent sources of patterning diffusible morphogens that may have effects on the hypothalamus.
Anterior neural ridge (ANR; yellow), which releases FGF8 is in fact a part of the Template:Roof plate (dorsalizing influence), rather than a source of AP effects. in contrast, the retromamillary and mamillary Template:Floor plate (dark blue associated to RM and M) releases SHH (ventralizing influence; note Shh secondarily also is expressed throughout the basal plate, and is later downregulated at the Tu area). The acroterminal midline as a source of AP patterning effects. Recent observations show Fgf18 expression within the postulated alar acroterminal organizer (fuchsia-labeled) and Fgf8 and Fgf10 expression within the postulated basal acroterminal organizer (green-labeled). There also are bilateral spots of Fgf8 expression at the optic stalks (not shown). (text modified from original figure legend) |
Hypothalamus Development Gene Interaction Model[17]

Transcription factor cascade (Sim1, Otp, Sim2, and Brn2) regulate differentiation of the anterior hypothalamus neuroendocrine cells.
References
- ↑ Rubenstein JL, Martinez S, Shimamura K & Puelles L. (1994). The embryonic vertebrate forebrain: the prosomeric model. Science , 266, 578-80. PMID: 7939711
- ↑ 2.0 2.1 2.2 2.3 Puelles L & Rubenstein JL. (2015). A new scenario of hypothalamic organization: rationale of new hypotheses introduced in the updated prosomeric model. Front Neuroanat , 9, 27. PMID: 25852489 DOI.
- ↑ Schredelseker T & Driever W. (2020). Conserved Genoarchitecture of the Basal Hypothalamus in Zebrafish Embryos. Front Neuroanat , 14, 3. PMID: 32116574 DOI.
- ↑ Fu T, Pearson C, Towers M & Placzek M. (2019). Development of the Basal Hypothalamus through Anisotropic Growth. J. Neuroendocrinol. , , e12727. PMID: 31050853 DOI.
- ↑ Nagpal J, Herget U, Choi MK & Ryu S. (2019). Anatomy, development, and plasticity of the neurosecretory hypothalamus in zebrafish. Cell Tissue Res. , 375, 5-22. PMID: 30109407 DOI.
- ↑ Ikeda Y, Kato-Inui T, Tagami A & Maekawa M. (2019). Expression of progesterone receptor, estrogen receptors α and β, and kisspeptin in the hypothalamus during perinatal development of gonad-lacking steroidogenic factor-1 knockout mice. Brain Res. , 1712, 167-179. PMID: 30776325 DOI.
- ↑ Alpár A & Harkany T. (2018). Novel insights into the spatial and temporal complexity of hypothalamic organization through precision methods allowing nanoscale resolution. J. Intern. Med. , , . PMID: 30027599 DOI.
- ↑ Luo SX, Huang J, Li Q, Mohammad H, Lee CY, Krishna K, Kok AM, Tan YL, Lim JY, Li H, Yeow LY, Sun J, He M, Grandjean J, Sajikumar S, Han W & Fu Y. (2018). Regulation of feeding by somatostatin neurons in the tuberal nucleus. Science , 361, 76-81. PMID: 29976824 DOI.
- ↑ Xie Y & Dorsky RI. (2017). Development of the hypothalamus: conservation, modification and innovation. Development , 144, 1588-1599. PMID: 28465334 DOI.
- ↑ Alvarez-Bolado G, Grinevich V & Puelles L. (2015). Editorial: Development of the hypothalamus. Front Neuroanat , 9, 83. PMID: 26157363 DOI.
- ↑ Hughes AT, Guilding C & Piggins HD. (2011). Neuropeptide signaling differentially affects phase maintenance and rhythm generation in SCN and extra-SCN circadian oscillators. PLoS ONE , 6, e18926. PMID: 21559484 DOI.
- ↑ Wen S, Ai W, Alim Z & Boehm U. (2010). Embryonic gonadotropin-releasing hormone signaling is necessary for maturation of the male reproductive axis. Proc. Natl. Acad. Sci. U.S.A. , 107, 16372-7. PMID: 20805495 DOI.
- ↑ Lemaire JJ, Nezzar H, Sakka L, Boirie Y, Fontaine D, Coste A, Coll G, Sontheimer A, Sarret C, Gabrillargues J & De Salles A. (2013). Maps of the adult human hypothalamus. Surg Neurol Int , 4, S156-63. PMID: 23682342 DOI.
- ↑ 14.0 14.1 Jankowski M, Danalache B, Wang D, Bhat P, Hajjar F, Marcinkiewicz M, Paquin J, McCann SM & Gutkowska J. (2004). Oxytocin in cardiac ontogeny. Proc. Natl. Acad. Sci. U.S.A. , 101, 13074-9. PMID: 15316117 DOI.
- ↑ Jankowski M, Wang D, Hajjar F, Mukaddam-Daher S, McCann SM & Gutkowska J. (2000). Oxytocin and its receptors are synthesized in the rat vasculature. Proc. Natl. Acad. Sci. U.S.A. , 97, 6207-11. PMID: 10811917 DOI.
- ↑ Neurog2 Acts as a Classical Proneural Gene in the Ventromedial Hypothalamus and Is Required for the Early Phase of Neurogenesis Journal of Neuroscience 29 April 2020, 40 (18) 3549-3563; DOI: https://doi.org/10.1523/JNEUROSCI.2610-19.2020
- ↑ Ratié L, Ware M, Barloy-Hubler F, Romé H, Gicquel I, Dubourg C, David V & Dupé V. (2013). Novel genes upregulated when NOTCH signalling is disrupted during hypothalamic development. Neural Dev , 8, 25. PMID: 24360028 DOI.
Reviews
Nagpal J, Herget U, Choi MK & Ryu S. (2019). Anatomy, development, and plasticity of the neurosecretory hypothalamus in zebrafish. Cell Tissue Res. , 375, 5-22. PMID: 30109407 DOI.
Xie Y & Dorsky RI. (2017). Development of the hypothalamus: conservation, modification and innovation. Development , 144, 1588-1599. PMID: 28465334 DOI.
Clarke IJ. (2015). Hypothalamus as an endocrine organ. Compr Physiol , 5, 217-53. PMID: 25589270 DOI.
Pearson CA & Placzek M. (2013). Development of the medial hypothalamus: forming a functional hypothalamic-neurohypophyseal interface. Curr. Top. Dev. Biol. , 106, 49-88. PMID: 24290347 DOI.
Viero C, Shibuya I, Kitamura N, Verkhratsky A, Fujihara H, Katoh A, Ueta Y, Zingg HH, Chvatal A, Sykova E & Dayanithi G. (2010). REVIEW: Oxytocin: Crossing the bridge between basic science and pharmacotherapy. CNS Neurosci Ther , 16, e138-56. PMID: 20626426 DOI.
Markakis EA. (2002). Development of the neuroendocrine hypothalamus. Front Neuroendocrinol , 23, 257-91. PMID: 12127306
Articles
Watts AG. (2011). Structure and function in the conceptual development of mammalian neuroendocrinology between 1920 and 1965. Brain Res Rev , 66, 174-204. PMID: 20637232 DOI.
Divall SA, Williams TR, Carver SE, Koch L, Brüning JC, Kahn CR, Wondisford F, Radovick S & Wolfe A. (2010). Divergent roles of growth factors in the GnRH regulation of puberty in mice. J. Clin. Invest. , 120, 2900-9. PMID: 20628204 DOI.
Alkemade A, Friesema EC, Unmehopa UA, Fabriek BO, Kuiper GG, Leonard JL, Wiersinga WM, Swaab DF, Visser TJ & Fliers E. (2005). Neuroanatomical pathways for thyroid hormone feedback in the human hypothalamus. J. Clin. Endocrinol. Metab. , 90, 4322-34. PMID: 15840737 DOI.
Books
Endocrinology - An Integrated Approach Stephen Nussey and Saffron Whitehead St. George's Hospital Medical School, London, UK Oxford: BIOS Scientific Publishers; 2001. ISBN-10: 1-85996-252-1
- Hypothalamic control of adrenocortical steroid synthesis - CRH and vasopressin
- Anatomical and functional connections of the hypothalamo-pituitary axis
- Blood supply of the hypothalamo-pituitary axis
- Autonomic functions of the hypothalamus
Historic
Castro-Dufourny I, Carrasco R, Prieto R & Pascual JM. (2017). Jean Camus and Gustave Roussy: pioneering French researchers on the endocrine functions of the hypothalamus. Pituitary , 20, 409-421. PMID: 28265842 DOI.
Search PubMed
Search Pubmed: hypothalamus development
Additional Images
Mouse hypothalamus SHH expression





Terms
- 3ß-diol - 5alpha-androstane-3beta, 17beta-diol
- 3α-diol - 5alpha-androstane-3alpha, 17beta-diol
- 5-HT - serotonin
- 5αR - 5-alpha-reductase
- ACTH - adrenocortcotropin releasing hormone
- AR - androgen receptor
- ARN - (AR) Arcuate nucleus
- AVP - arginine vasopressin
- BnST - bed nucleus of the stria terminalis
- BSA - bovine serum albumin
- CBP - CREB binding protein
- CORT - corticosterone
- CRE - cyclic adenosine monophosphate response element
- CREB - CRE binding protein
- CRF - corticotropin releasing factor
- DES - diethylstilbesterol
- DEX - dexamethasone
- DHT - dihydrotestosterone
- DMN - (DM) Dorsomedial hypothalamic nucleus
- DPN - diarylproprionitrile
- ER - estrogen receptor
- ERE - estrogen response element
- FSH - follicle stimulating hormone
- FSL - flinders sensitive line
- GABA - gamma-aminobutyric acid
- GAS - general adaptation syndrome
- GH - growth hormone
- GLP-1 - glucagon-like peptide 1
- GnRH - gonadotropin releasing hormone
- GPER - G-protein coupled estrogen receptor
- GR - glucocorticoid receptor
- HPA - hypothalamo–pituitary–adrenal
- HPG - hypothalamo–pituitary–gonadal
- HRE - hormone respone element
- HSD - hydroxysteroid dehydrogenase
- ICV - intracerebroventricular
- ir - immunoreactive
- ISH - in situ hybridization
- LH - luteinizing hormone
- LHRH - luteinizing hormone releasing hormone
- median eminence - part of the inferior boundary of the hypothalamus
- MPOA - medial preoptic area
- MR - mineralocorticoid receptor
- NTS - nucleus of the solitary tract
- PPT - propylpyrazoletriol
- PR - progesterone receptor
- PVN - (PVA, PVH) Paraventricular nucleus
- RAR - retinoic acid receptors
- SCN - (SC) Suprachiasmatic nucleus
- SON - (SO) Supraoptic nucleus
- SRC - steroid receptor coactivators
- T - testosterone
- THR - thyroid hormone receptors
- TPH - tryptophan hydroxylase
- TRH - thyrotropin releasing hormone
- TSH - thyroid stimulating hormone
- VMN - (VM) Ventromedial nucleus
External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2024, May 21) Embryology Endocrine - Hypothalamus Development. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Endocrine_-_Hypothalamus_Development
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G