Paper - 1879 The Morphology of the Vertebrate Olfactory Organ
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Marshall AM. The morphology of the vertebrate olfactory organ. (1879) Quarterly Journal of Microscopic Science. 19: 300–340.
| Online Editor Note - Neural Crest and About the Author | |||||||||
|---|---|---|---|---|---|---|---|---|---|

Arthur Milnes Marshall (1852–1893) Died in a mountaineering accident in 1893 at Scafell (Scaw-fell) a mountain in the English Lake District, part of the Southern Fells.
|
| Historic Disclaimer - information about historic embryology pages |
|---|
|
The Morphology of the Vertebrate Olfactory Organ

By A. Milnes Marshall, M.A., D.Sc, Fellow of St. John's College, Cambridge.
With Plates XIII and XIV.
An abstract of this paper was read before the Royal Society ou February 13th, Proc. Roy. Soc., No. 193, 18?9.
Quarterly Journal of Microscopic Science (1879) Vol. 19: 300–340.
Of the two parts into which the present paper is divided, the first deals with the development of the olfactory nerve in certain selected types of vertebrates ; the second with the development of the olfactory organ in the same types.
Since the value and interest of anatomical and embryological facts consist largely in their application to the solution of morphological problems, I have not hesitated to draw inferences freely from such facts as I have been able to bring to light, or to point out the conclusions to which these facts seem to me to lead. However, in order to separate facts from theories as sharply as possible, each part of the paper has been farther subdivided, those portions which are concerned with matters of direct observation being considered before those which are of a more theoretical or speculative nature.
I. The Development of the Olfactory Nerve
a. In the dogfish
For the opportunity of investigating the development of Elasmobranchs I am indebted to Mr. Balfour, who, on the completion of his monograph on Elasmobranch fishes, very kindly placed at my disposal the whole of his stock of uncut embryos. In addition to this I have had the great advantage of free access to the very complete series of preparations made by Mr. Balfour in the course of his investigations, and have availed myself of his permission to figure four specimens, illustrating stages of which I had not prepared satisfactory sections myself.
The greater number of the embryos thus placed in my hands were those of the Scyllmm canicula, some few of Prhtiurus ; but inasmuch as the two genera have yielded identical results so far as the subject in hand is concerned, I have made no attempt to distinguish between them either in my descriptions or figures. Some few of the specimens were hardened in picric acid, and afterwards stained with haematoxylin ; but all my best sections were from embryos hardened and stained in a quarter per cent, solution of chromic acid, to which a few drops of a weak solution of osmic acid had been added.
With regard to the earliest stages in the development of the olfactory nerve, I have, unfortunately, heen unable to make any satisfactory observations, for all the specimens younger than Balfour's stage k were in bad condition. The chief points I wished, if possible, to determine were — firstly, whether the neural ridge extends to the anterior end of the fore brain in Elasmobranchs, as I have already shown it to do in the chick[2] ; secondly, whether the olfactory nerve is developed from this ridge ; and, lastly, the exact date of appearance of the olfactory nerve. On all the.se points I have, owing to the unsatisfactory condition of my specimens, failed to obtain reliable evidence.

Plate XIV, fig. 19, represents a section through the head of a dogfish embryo at stage m of Balfour's nomenclature; the section is made in a plane transverse to the longitudinal axis of the head, and passes through the fore brain (. h.), the olfactory sacs (olf.), and the olfactory nerves (I).
This figure, which is taken from one of an excellent series of preparations in perfect histological preservation, illustrates several features of considerable interest —
- In the first place it will be noticed that the fore brain presents no trace whatever of a division into cerebral hemispheres ; in other words, that the olfactory nerves come into existence before the cerebral hemispheres, and are therefore connected at first with the forebrain, and not with the hemispheres. As confirmation of this point, I may repeat that fig. 19 is taken from an embryo at stage m, while Balfour has already shown, and my own observations are in complete accordance with his on this point, that until stage o there is no trace whatever of a division of the forebrain into cerebral hemispheres.[3]
- There is no trace of an olfactory lobe or vesicle. This is a point of considerable importance, and one on which I desire to lay stress. The figure shows that at stage m the olfactory nerves are solid, and present no trace of a central lobe or vesicle, either at their roots or at any part of their length.
- The olfactory nerve at stage m agrees closely in its general relations and in its histological characters with the other cranial nerves, either at the same or at slightly younger stages. Like these, it arises from the upper part of the sides of the brain, and takes a course downwards and outwards, at right angles to the longitudinal axis of the head. Histologically it consists of roundish or oval nucleated cells, with, as yet, very few nerve fibres, agreeing completely with corresponding stages of development of the other cranial nerves.

Fig. 20 is taken from a section through the same region as fig. 19, but from a dogfish embryo at the commencement of stage o. The magnifying power employed is the same in the two drawings, so that an exact comparison can be made between them. There is still no indication of a division into cerebral hemispheres; the forebrain, as in fig. 19, is still undivided. Though the embryo has grown considerably the olfactory nerve (i), though somewhat thicker, is no longer in fig. 20 than in fig. 19, a fact of some interest; its point of attachment to the brain has, however, shifted down somewhat towards the ventral side. The most important fact shown by fig. 20 is, however, the existence of the earliest rudiment of an olfactory lobe (ol. v.). This, as may be seen from the figure, is exceedingly small, and might indeed be easily overlooked ; it is a small shallow pit, formed almost entirely at the expense of the inner wall of the forebrain, and situated opposite the root of origin of the olfactory nerve.

In fig. 21, taken from one of Mr. Balfour's specimens, the same parts are shown at a stage intermediate between stages o and p. The olfactory vesicle (ol. v.) is seen to have grown very rapidly, and is now a conspicuous object. The olfactory nerve (i), on the other hand, has remained almost stationary as far as size is concerned ; it has, however, undergone considerable histological change ; the cells composing its proximal part or root of origin are more elongated and fusiform than before, while beyond this part the nerve presents a ganglionic expansion consisting mainly of roundish cells, similar to those which previously constituted the whole nerve, and which gives off, distally, bundles of nerve fibres distributed to the Schneiderian folds of the olfactory mucous membrane.

The condition of the olfactory nerve and lobe at stage q is shown in fig. 22, also taken from one of the specimens lent me by Mr. Balfour, who has described this stage as follows : — " The lateral ventricles are now separated by a median partition, and a slight external constriction marks the lobes of the two hemispheres ; these, however, are still united by nervous structures for the greater part of their extent. The olfactory lobes are formed of a distinct bulb and stalk, and containj as before^ prolongations of the lateral ventricles."[4] It will be noticed that, while in fig. 21 the olfactory lobe projects out at right angles to the brain, and the olfactory nerve arises from its extreme tip, in fig. 22 the olfactory lobe is bent downwards, so as to lie against the side of the cerebral hemisphere, and the olfactory nerve no longer arises from its apex, but slightly from its dorsal surface. From the condition here represented to that of the adult the changes are unimportant.
The earlier stages of the olfactory nerve I have not been able to work out satisfactorily, for reasons already mentioned. In fig. 15 the nerve is represented in longitudinal and vertical section at stage m. It is easily recognisable at stage L, and I have also succeeded in satisfying myself of its existence as far back as stage k.
Fig. 14 represents a transverse section through the anterior part of the head of an embryo at the commencement of stage K ; the section passes through the forebrain, and through both olfactory pits ; on the right side a small mass of cells (i) in contact with the bottom of the pit is stained rather more deeply than the surrounding mesoblast cells. From comparison with the condition of what is undoubtedly the olfactory nerve at slightly later stages, I consider it very probable that these cells form part of the olfactory nerve, but cannot, of course, speak with any certainty on this point. Apart from the insufficient material at my disposal, the inherent difficulties of the investigation are very great, for at these early stages the olfactory nerve consists entirely of cells, which differ but little from the surrounding mesoblast cells ; the nerve is also exceedingly short, owing to the close proximity of the olfactory pit to the brain, while a new difficulty is introduced by cranial flexure, which is increasing rapidly about this time, and so causes a constant shifting in the relations of the surrounding parts to one another.
My investigations, then, lead me to give the following account of the development of the olfactory nerve inElasmobranchs. The nerve arises at some period earlier than stage K ; it is at first connected with the upper part of the side of the forebrain ; between stages l and o its root shifts downwards to a certain extent towards the ventral surface ; the nerve itself is, from the earliest period at which it can be recognised, solid ; the earliest trace of an olfactory lobe appears at the commencement of stage o as a shallow depression of the inner wall of the forebrain opposite the root of the olfactory nerve ; this olfactory lobe grows very rapidly, and soon attains a large size, while the olfactory nerve remains almost stationary ; the nerve is at first connected with the apex of the olfactory lobe, but subsequently mounts somewhat on to its dorsal surface ; finally, the olfactory nerve, throughout its development, agrees closely in histological characters and in the changes which it undergoes with the other cranial nerves.
Balfour has given a somewhat different account of the development of the olfactory nerve. After noticing that the olfactory lobes first arise during the stage o, he says : — "From the peripheral end of each olfactory lobe a nerve, similar in its histological constitution to any other cranial nerve, makes its appearance ; this divides into a number of branches, one of which passes into the connective tissue between the two layers of epithelium in each Schneiderian fold. On the root of this nerve there is a large development of ganglionic cells. I have not definitely observed its origin, but have no reason to doubt that it is a direct outgrowth from the olfactory lobe, exactly similar in its mode of development to any other nerve of the body."^ A little further on he remarks : "Even the few preparations of which I have given figures appear to me to prove that . . . from the (olfactory) bulb a nerve grows out which has a centrifugal growth like other nerves of the body, and places the central olfactory lobe in communication with the peripheral olfactory sack."2
The differences between this account and my own are sufficiently obvious. According to Balfour, the olfactory lobe exists before the olfactory nerve, and the nerve is a " direct outgrowth from the olfactory lobe." A minor point of difference is that, according to Balfour, the connection between the olfactory nerve and the olfactory pit is not acquired till towards the end of stage o. I believe, however, that these differences are due to Balfour having overlooked the existence of the olfactory nerve during its early stages. The first stage at which he has described the olfactory nerve is that which I have represented in fig. 21,^ while the specimens I have figured (figs. 19 and 20) appear to me to prove indisputably the existence of the olfactory nerve at a much earlier period, and the connection between the olfactory nerve and olfactory pit appears to be acquired at least as early as stage K.
1 Op. cit., p. 178.
2 Op. cit., p. 183.
3 The section from which this figure is drawn is one of the same series, if not the identical specimen, as that described by Balfour, and figured by him in n. XA fig. 2.
b. In the chick
I propose to consider the chick next, partly because, having devoted more time to the embryology of the chick than of other vertebrates, I have a better and more complete series of preparations to refer to, and partly because I wish to direct particular attention to the very close correspondence that exists between the chick and the dogfish in the mode of development of the olfactory nerve.
Concerning the early stages of the olfactory nerve in the chick I have little or nothing to add to the account I have already given in this Journal.[5] The result of a careful re-examination of my former preparations, and the investigation of a considerable number of new specimens prepared since my former paper was published, has been to confirm my previous description on all points. Though I have again failed to trace satisfactorily the changes that occur between the thirtieth and fiftieth hours my further work has shown no reason for altering the view I have previously expressed, that the olfactory nerve is developed, like all the other cranial nerves (except the optic, the sixth, and (?) the fourth nerve), from the neural crest.[1] However, whether this be so or not is of comparatively little importance to the subject with which we are now concerned.
Plate XIII, fig. 10, represents part of a transverse section through the fore part of the head of a duck embryo towards the end of the fourth day. This figure, which is repeated with slight alterations from a former paper,[6] happens to show the points to which I wish to call attention rather better than any of my chick preparations, the specimen from which it was taken being in unusually good preservation. The section passes through the forebrain (f. b.), the olfactory pit [olf.), and the olfactory nerve (i). From it we learn (1) that the olfactory nerves exist prior to the cerebral hemispheres, of which latter there is in this specimen no trace whatever ; (2) that in this stage there is no indication whatever of an olfactory lobe ; (3) that the olfactory nerve is in its early stages connected with the upper or dorsal part of the side of the forebrain '^ (4) that the connection between the olfactory nerve and olfactory pit is very early acquired ; (5) that the olfactory nerve at this stage agrees closely in histological characters with the corresponding stages of the other cranial nerves, consisting almost entirely of roundish or oval nucleated cells with few or no nerve fibres.
This figure may be advantageously compared with fig. 19, which represents, as already described, a section through the same region in a dogfish embryo at stage m. The resemblance between these two figures is indeed very striking, and extends even to the minute histological details. I would lay great stress on this resemblance, and submit that this close correspondence, amounting almost to identity, in the condition of the olfactory nerve at similar stages in two vertebrates so widely separated as the chick and the dogfish, affords very strong evidence in favour of the correctness of my observations. Such differences as do exist are of very minor importance. Apart from the slight difference in general configuration, the most significant are the rather larger relative size of the olfactory nerve and pit in the dogfish, obviously correlated with their condition in the adult, and the fact that in the duck the attachment of the olfactory nerve is rather nearer to the summit of the forebrain than in the corresponding stage of the dogfish.
The appearance of the cerebral hemispheres towards the close of the third day in the chick causes considerable alteration in the position and relations of the olfactory nerves. The hemispheres are lateral outgrowths of the forebrain, and are from the first situated on the dorsal side of the roots of the olfactory nerves. They grow forwards and upwards with exceeding rapidity, and by so doing drive the olfactory nerves down to the base of the brain, and so cause these nerves to appear to arise from their under and anterior part ; a change which has proved a fruitful source of misconception as to the true nature and relations of the olfactory nerves, especially as these latter are usually not recognised until they have taken up this secondary position.
1 Though the nerve is in close contact with the brain, the actual connection between the two is not seen in the specimen figured ; it is clearly visible in one of the sections of the same series immediately adjacent, which however, does not show the whole length of the nerve, and is, therefore, less suitable for figuring.
The change to which I have just referred is well illustrated by fig. 11, a transverse section through the anterior part of the head of a chick embryo at the eightieth hour. The section shows the commencing cerebral hemispheres [c. h.) growing upwards and outwards from the forebrain; it also passes through the margins of the two olfactory pits {olf.), and on the left side through the root of the olfactory nerve (i) at its point of origin from the brain ; the figure shows very clearly the effect of the appearance of the cerebral hemispheres on the position of the olfactory nerves, and shows further how the secondary connection of these nerves with the hemispheres is acquired.
Fig. 12 represents a section from the same series as fig. 11, but taken a little further back, passing through the olfactory pits {olf.) at their deepest parts. On the right side the section passes through the distal portion of the olfactory nerve (i), which is seen to be in continuity with the bottom of the olfactory pit.
In figs. Hand 12 the olfactory nerve has the same histological character as in fig 10 ; it is, however, relatively, if not indeed absolutely, smaller than at the earlier period. The figures further show clearly that there is as yet no trace whatever of an olfactory lobe.
I have elsewhere^ given figures and description of the condition of the olfactory nerves at the ninety-third hour in the chick, at which date, excepting a general increase in size, their condition differs but little from that at the eightieth hour.
Figs. 7 and 8 represent longitudinal and vertical sections through the anterior part of the head of a chick embryo towards the end of the sixth day of incubation. As the olfactory nerve did not lie exactly in the plane of section it has been necessary to figure two sections, of which the more superficial one (fig. T) shows the greater part of the length and the peripheral distribution of the olfactory nerve ; while the second section (fig. 8), taken at a slightly deeper level, shows the root of origin of the nerve from the brain. The olfactory nerve, which is still short, presents a proximal ganglionic swelling at its point of origin from the hemisphere, seen best in fig. 8 ; along the greater part of its length the nerve consists of very elongated fusiform cells, with a few spherical ganglionic cells at intervals ; distally, at its connection with the olfactory pit, it presents a second ganglionic swelling, fig. 7.
' Loc. cit., p. 20, and Plate II, figs. 17—19.
.
A point of very considerable interest, shown in the clearest possible manner by these figures, is that up to this date there is no indication of an olfactory lobe ; indeed, instead of a hollow process of the hemisphere at the point of origin of the olfactory nerve, there is at this point, as is shown by both figures, but especially by fig. 8, a slight external depression, with a very obvious internal projection of the wall of the hemisphere.
Fig. 9 represents a similar section, in a longitudinal and vertical plane, through the nasal region of a chick at the end of the seventh day ; passing through the cerebral hemisphere (c. h.), the eye (o. c), the anterior extremity of the ophthalmic branch of the fifth nerve (v. a.), the olfactory pit (olf.), and the olfactory nerve (i). The nerve itself presents the same histological characters as in fig. 7, /. e.[a. proximal ganglionic enlargement at its root of origin, a trunk consisting mainly of nerve fibres, but with a few ganglionic cells at intervals along its whole length, and a distal ganglionic expansion at its point of fusion with the olfactory epithelium. There is, however, one important difference between this figure and the two preceding ones ; opposite the point of origin of the olfactory nerve there is a small conical depression (ol. v.) of the inner wall of the cerebral hemisphere. From a comparison with fig. 20 there can be little doubt that this is the earliest appearance of an olfactory lobe. As in the dogfish, this lobe is formed at first entirely at the expense of the inner wall of the hemisphere, there being as yet no perceptible projection on the exterior of the brain.
The olfactory lobes, after their first appearance, grow rapidly. By the twelfth day they form a pair of small conical processes, about 1 millimetre in length, springing from the extreme anterior ends of the cerebral hemispheres : the two lobes lie side by side, their apposed surfaces being slightly flattened. Each lobe contains a prolojigation of the ventricular cavity of the corresponding hemisphere.
Fig 36 represents a longitudinal and vertical section through the olfactory lobe and the anterior part of the cerebral hemisphere of a twelfth day chick embryo : it shows how the ventricle of the hemisphere (c.h.) is prolonged to the extremity of the olfactory lobe (ol.v.) ; and also the mode in which the olfactory nerve arises from the end of the olfactory lobe as a series of bundles of nerve fibres.
Fig. 37 is a transverse section through the olfactory nerve of a chick embryo of the same age as fig. 36 ; it shows the bundles of nerve fibres, bound together by connective tissue, which together constitute the olfactory nerve ; it shows also how the majority of these bundles are arranged in a circle round the margin of the nerve, while a few smaller bundles lie in the centre.
Figs. 38 and 39 represent sections taken from the same embryo as the preceding figure. Fig. 38 is a transverse section through the olfactory lobe, and shows the laterally compressed ventricular cavity. Fig. 39 is a transverse section through the anterior part of the hemisphere : the outer wall of the hemisphere is seen to have increased greatly in thickness while the inner wall still remains thin ; so that the ventricle, which is greatly compressed laterally, no longer occupies the centre of the hemisphere, but lies close to its inner side.
In the adult fowl the olfactory lobe has much the same appearance as at the twelfth day : it is about two and a half millimetres in length, and still contains a central cavity, though this latter is relatively smaller than at the earlier date ; the relations of the olfactory nerve to the lobe are the same as at the twelfth day.
' In my former paper I stated that there is no trace of an olfactory vesicle at any i^eriod in the life of a chick} This statement my later work now shows to be erroneous ; the chick has an olfactory vesicle, but, as in the dogfish, this vesicle does not appear till an exceedingly late period of development.
The principal points then, in the development of the olfactory nerves in the chick to which I desire to direct attention are :
- The olfactory nerves arise from the forebrain, before the cerebral hemispheres have begun to be developed.
- They are at first connected with the dorsal surface of the forebrain, but on the appearance of the hemispheres become driven down to the ventral surface of the brain, and acquire a secondary connection with these latter.
- From their earliest appearance the olfactory nerves are solid, and present the same histological characters as the other cranial nerves.
- There is not the slightest indication of an olfactory lobe till the latter part of the seventh day of incubation.
Though these conclusions are in complete accordance with my earlier work, they are directly opposed to all other accounts with which I am acquainted, with one solitary exception, to which I shall refer immediately. As the date of appearance of the olfactory lobe is the point in which there is the greatest discrepancy between the descriptions of previous writers and my own, I have made sections in very various ])lanes in order to detect any appearance that could possibly be interpreted as an olfactory lobe at an earlier date than the seventh day, but have failed completely to observe any such.
1 Loc. cit., p. 20. VOL. XIX. NEW SER. X .
As far as I can ascertain, the earliest account of the development of the olfactory nerve is that given by Remak ; this description, which only occupies about three lines, and is unsupported by figures, is as follows : — " An ihrem Boden (Hemispharen) zeigen sich gegendas Ende des drittenTages jederseits kleine birnformige Blaschen (Geruchsblaschen) iiber deren weiter entwickelung ich keine Beobachtungen besitze."' This observation was repeated later on by von Baer, who, however, went further than Remak, and described this vesicle as the rudiment of the olfactory nerve ; he also described an olfactory pit distinct from this vesicle. Concerning these statements Remak speaks thus : — " Halte ich diese Angaben mit meineu eigenen Wahrnehmungen zusaminen, so wird es mir sehr wahrscheinlich, dass Baer am vierten Tage die Geruchsblaschen und die Nasengruben nicht gleichzeitigbeobachtet, dass er vielmehr dasselbe Gebilde bald als Anlage des Riechnerven, bald als Nasengrube gedeutet hat. Ich habe mich namlich iiberzeugt, dass die Geruchsblaschen, die zu Ende des dritten Tages auftreten, die nasengruben sind, und dass weder alsdann, noch bis zum fiiiiften Tagen ein entsprechender Auswuchs des vorderhirnes wahrzunehmen ist."[7] This very definite statement shows with perfect clearness not only that Remak recognised and corrected his original mistake, recognised, i. e. that what he had originally taken for outgrowths of the cerebral hemispheres were really the olfactory pits, a mistake doubtless due to his relying on surface view of whole embryos; but also that he discovered and recorded the fact that as late as the end of the fifth day there is no trace of an olfactory lobe.
Strange as it may seem, this exceedingly definite and accurate statement of Remak's has been completely overlooked, while his earlier, vague, brief, and avowedly imperfect observation actually furnishes the basis of the descriptions of the development of the olfactory nerve given in our text-books of embryology at the present day.
Thus, Professor Kolliker, in the second part of his textbook of embryology, published in the course of the present year, dismisses my previous account of the development of the olfactory nerve as " eine Angabe, die mit der Darstellung von Remak, d^pr zufolge die Lohi olfactorii des -Hiibnchens am Ende des zz. Tages als kleine birnforrnige Blaschen am Bodeii der Hemispharenblasen liegen, nicht zu vereineu ist." [8]Professor Kolliker's words show, beyond doubt, that he is quoting from Remak's earlier statement ; had he been acquainted with the latter part of Remak's work he would have known that my observations confirmed instead of contradicting Remak.
Again, Foster and Balfour describe the development of the olfactory nerve in the chick thus : — " At the under surface of each of the vesicles of the cerebral hemispheres there appears towards the end of the third day a small, somewhat elongated vesicle — the olfactory vesicle — which is the rudiment of the olfactory nerve or bulb."^ The authors make this statement on their own authority, but since the first part of their description is an almost literal translation of Remak's earlier account, it is, I think, a fair inference that they have fallen into the same error as Professor KoUiker. Remak, however, is not responsible for the statement that this olfactory vesicle is " the rudiment of the olfactory nerve or bulb."
Any further discussion of the literature of this subject would be unprofitable ; it is, however, only fair to add that at the time of writing my previous paper I had not referred to Remak's work, and was under the impression that my description was completely at variance with his account ; it is, therefore, a matter of great satisfaction to myself to find my statements corroborated by such high authority.
c. In the salmon and trout
Though my observations on Teleostean embryos are not nearly so complete as those I have just recorded concerning the chick and dogfish, yet, inasmuch as they have yielded definite, and in some respects important and unexpected results, I have thought it well to record them here.
The ova were obtained in the early part of last year from Mr. Capel, of the Foot's Cray Fishery ; for the opportunity of hatching them I am indebted to Mr. F. Buckland, to whom my best thanks are due for the liberal and courteous manner in which he met my requests. I am also much indebted to Mr. Edon, of the South Kensington Museum of Pisciculture, to whose care and experience I owe the successful hatching of the ova.
The early stages of development of the olfactory nerve are unfortunately even more difficult to investigate in Teleosteans than in either the chick or dogfish ; and my observations on these stages are exceedingly imperfect. The earliest stage at which I can speak with any confidence as to the existence of an olfactory nerve is shown in fig. 29, which represents a transverse section through the anterior part of the head of a trout embryo on the twenty-seventh day after the fertilization of the ova. The section passes through the forebrain (f.b.), and through the olfactory pits [olf-); on the left side of the section a small mass of cells, somewhat more compactly arranged and more deeply stained than the mesoblast cells, connects the upper part of the forebrain with the olfactory pit. This mass of cells (i) I believe to be the olfactory nerve, mainly from its relation to what is undoubtedly the olfactory nerve a few days later. I do not wish, however, to speak at all positively on this point.
Between the thirtieth and fortieth days the olfactory nerves, though still extremely short, can be easily recognised. Though my observations are far from complete, they suffice to establish the following points for both the salmon and trout :
- The olfactory nerves appear before the cerebral hemispheres, and are at first connected with the dorsal side of the forebrain.
- The nerves are, from the earliest period at which their existence can be determined with anything like certainty, solid ; i.e. there is no olfactory lobe.
- The connection between the olfactory nerve and the epithelium of the olfactory pit is acquired at a very early date.
Plate XIV, fig. 33, is taken from a transverse section through the head of a salmon embryo two days after hatching. The section, which is a little oblique, passes on the left side through the eye (o. c.) with the superior (r. s.) and inferior [r.i.) recti muscles; on the right side through the olfactory pit [olf.) The forebrain (_/.i.), which lies in the centre of the section, is seen to have a small vesicular cavity in its upper part; its roof is thin, its floor and sides very thick. From the lower part of its sides a pair of nerves (i) arises; these nerves run downwards for a short distance towards the ventral surface, then turn directly outwards, and the nerve on the right side is seen to divide into two branches, which can be readily traced to the thickened epithelium lining the bottom of the olfactory pit {olf.). It will be noticed that there is no trace of an olfactory lobe, and that the olfactory nerve presents no ganglionic enlargement at any part of its course. In its general relations, mode of origin, and course, the nerve agrees remarkably closely with the other cranial nerves, while in histological characters it is identical with them.
In fig. 34 the olfactory nerve (i) is seen in longitudinal and vertical section in a salmon embryo of the same age as that just described. This section shows well the relations of the olfactory nerve to the brain ; it also shows the roots of the optic nerves (ii), the infundibulum (e'w/.), and the trabeculce cranii {tr.).
It would seem, therefore, that if an olfactory lobe is present at any period in the life of a salmon or trout, it does not make its appearance till very late — so late, indeed, that it could have no claim to be considered as an embryonic structure at all : there is no trace of it at the time of hatching, or, indeed for some days afterwards.
d. In other vertebrates
In the Axolotl[9] the olfactory nerve is at first connected with the forebrain, not with the hemispheres. Throughout the whole period of embryonic development it is very short, and in the early stages exceedingly so : it is solid, and agrees completely in histological characters with the other cranial nerves. I have failed to detect an olfactory lobe in any of the stages I have examined; i.e. up to the time of hatching.
I have also made some observations on the earlier stages of development of the olfactory nerves in the frog, which show that in these stages the nerves are extremely short, and that there is no trace of an olfactory lobe. The resemblance between the frog and axolotl is, as might be expected, exceedingly close.
In some lizard embryos, for which I am indebted to Mr. Balfour, I have noticed the existence of solid olfactory nerves, with no indication of olfactory lobes, at stages apparently corresponding to the fourth or fifth day of incubation of the chick; and I believe I have succeeded in establishing the existence of olfactory nerves at still earlier stages, before the appearance of the cerebral hemispheres. In the later stages the olfactory lobes are more prominent objects than in the chick.
I will, in conclusion, quote from Professor Parker the following description of the development of the olfactory nerve in the green turtle : — " In embryos of the green turtle of the size of a horse-bean I find the nerves (olfactory) solid. When the embryos are two or three times that size, these nerves each acquire a large cavity proximally, from the fore wall of which the branches seem to spring. The foremost of these branches spring from the top of the vesicle ; they arose at first from the top of the forebrain."[10]
e. General considerations
Before proceeding to the development of the olfactory organ, I propose to summarise the results to which we have already been led, and to consider briefly certain questions of a more theoretical character.
The first point I desire to call attention to is the remarkably close agreement in the mode of development of the olfactory nerves presented by the several types examined, types which, it will be noticed, embrace examples from each of the vertebrate classes, with the exception of Mammalia. In all these types alike — dogfish, trout and salmon, axolotl, frog, lizard, turtle and chick — the mode of development is fundamentally the same ; while the resemblance between the dogfish and the chick, the most generalised and the most specialised of these types is, as I have already shown, complete. I would direct special attention to this agreetnent as affording very strong testimony of the correctness of my observations.
The fundamental points common to all the above types are the following : — 1, the olfactory nerves appear very early ; 2, they are at first connected with the forebrain, and iiot with the cerebral hemispheres ; 3, they are solid, and agree completely in histological characters with the other cranial nerves; 4, an olfactory lobe, when present at all, does not appear till an exceedingly late period of development.
Though the several types agree so closely in the above fundamental points, they present well-marked differences among themselves. The dogfish appear to form a central type round which the others may be grouped, and from which they may be supposed to be derived. Curiously enough, of the other types the chick appears to resemble the dogfish more closely than any of the others do, with the possible exception of the lizard and turtle, whose earlier stages are as yet unknown. The Amphibia are chiefly characterised by the extreme and long persisting shortness of their olfactory nerves, and are in no way intermediate between the dogfish and such Sauropsida as I have examined. Finally, the Teleosteans, if the salmon and trout may be taken as typical of that group, while they resemble the Amphibia in the extreme shortness of their olfactory nerves in the early stages of development, seem to differ somewhat from the -other types in the exceedingly late appearance of the olfactory lobes, and in the striking resemblance in general anatomical behaviour between the olfactory and the other cranial nerves.
The nomenclature of the olfactory nerve is, unfortunately, somewhat overburdened with synonyms, a never-failing source of confusion and inaccuracy. The " olfactory nerve " of an adult vertebrate is, })erhaps, best described as consisting of three pans; a ])roximal tractus olfactorius arising from the cerebral hemisphere, an intermediate ganglionic enlargement or bulbus olfactorius, from whose distal extremity the third part or tiervus olfactorius arises.[11] Of these parts the two former are commonly and correctly described as being properly parts of the brain, and as togetner con' stituting the rhinencephalon. By some authors, however, the term rliinencephalon appears to be limited to the bulbus olfactorius, the tractus olfactorius being then sooken of as the rhinenccphalic crus.[12] By olfactory lobe or olfactory vesicle is usually meant the hollow diverticulum of the forebrain or cerebral hemisphere in the embryo, from which both the tractus olfactorius and bulbus olfactorius of the adult are developed, and which has hitherto been erroneously supposed to be the earliest part of the " olfactory nerve " to be developed. It would, perhaps, be well to limit the term olfactory lobe to this embryonic structure ; Owen employs it in the adult as synonymous with bulbus olfactorius.
From the descriptions I have already given it follows that the nervus olfactoritis is the earliest of the three elements to be developed, and that it alone is the direct homologueof the other cranial nerves. The term olfactory nerve ought then to be strictly limited to the nervus olfactorius. Since, however, there is considerable inconvenience in disturbing established nomenclatures, it may ])erhaps be well to continue to use the term olfactory nerve in the ordinary anatomical sense, and to confine oneself to the term nervus olfactorius when wishing to speak of the third or distal element, the olfactory nerve proper ; in this case, however, it must be clearly understood that olfactory nerve and nervus olfactorius are by no means equivalent or mutually convertible terms.[13]
Though the hulbus olfactorius and tractus olfactorius are considered as together equivalent to the olfactory lobe of the embryo, it must be noticed that the proximal ganglion of the nervus olfactorius may fuse so completely with the bulbus, that it is, even in comparatively early stages, " rather difficult to fix on the exact line of demarcation between the bulb and the nerve."[14]
The three elements of the olfactory nerve, but especially the first and third, vary much in the relative proportions they attain in the adult. Thus, in the dogfish there is a large biilbus olfactorius, connected proximally with the hemispheres by a short, thick, tractus olfactorius, and giving origin distally to the numerous filaments of the nervus olfactorius. In the skate, while the bulbus and nervus retain much the same proportions as in the dogfish, the tractus olfactorius is of very great length. Among osseous fishes the variations are still greater; in the pike, salmon, perch, gurnard, &c., on the one hand, there is a very long nervus olfactorius, springing from a bulbus olfactorius which is in close contact with the hemispheres; on the other hand, in the cod, carp, &c., as in the skate, the bulbus olfactorius is situated near the olfactory organ, and is far removed from the rest of the brain, with which it is connected by a long tractus olfactorius.
A question of far more morphological interest is the rcdation of the olfactory nerve to the other cranial nerves. My observations, if confirmed, prove that in the chick up to the end of the sixth day, in the dogfish up to stage o, and in the salmon and trout, at any rate up to the time of hatching, the olfactory nerve agrees very closely in histological characters and in general anatomical relations with the other cranial nerves. I propose now to consider these resemblances more in detail, and specially in reference to the question of the segmental value of the olfactory nerve.
Certain of the cranial nerves — e.g. the facial and glossopharyngeal — have long been acknowledged to possess segmental value. If we consider the mode of development of these segmental cranial nerves, we find that they agree aniorg themselves, and differ sharply from other nerves or branches of nerves in the following embryological characters :
- They appear very early.
- They arise, at least in the chick, from the neural crest on the mid-dorsal surface of the brain.
- Shortly after their appearance their roots undergo a shifting downward of their points of attachment, so that they no longer arise from the dorsal surface, but from the sides of the brain.
- They present, at least in their early stages, ganglionic enlargements on or close to their roots of origin.
- Their course is at right angles to the longitudinal axis of the head.
- Finally, they have very definite relations to the segments of the head, as indicated by the visceral clefts, each nerve supplying the two sides of a cleft.
The true cranial segmental nerves, such as the facial and the glosso-pharyngeal, agree in presenting all these characters. On the other hand, the non-segmental nerves, or branches of nerves, though they may possess some of the characters above enumerated, yet never present all, and rarely more than one or two. This test suffices to dispose of the claims to segmental rank of the optic, the auditory, the fourth, and sixth nerves, and of the ophthalmic branch of the trigeminal nerve ; while, on the other hand, it serves to demonstrate the segmental value of the third nerve,
I propose now to apply this test to the olfactory nerve.
1. In all the types examined the olfactory nerve appears very early. Though the exact date of its first appearance has not been determined with certainty in any case, yet there is no reason for thinking that it arises later than the other cranial nerves. In all the types considered it appears before the cerebral hemispheres.^ In the dogfish it makes its appearance earlier than stage k, and in the chick there are strong reasons for thinking that it is "one of the first nerves in the body to appear."
2. I have already attempted elsewhere to prove that in the chick the olfactory nerve is developed from the neural crest. Though I have but little doubt on the matter myself, I have not yet succeeded in determining thispoint with absolute certainty in the case of the lizard.
Quarterly Journal Microscopical Science, Jan., 1878, p. 23.
Loc. cit., pp. 17 — 19. With reference to the extension forward of the neural crest in the chick to the forebrain, Prof. Kolliker suggests (op. cit., pp. 661 — 2), that 1 have been misled by certain folds which appear during closure of the medullary canal, and to which His lias already directed attenliou. With all due respect for Prof. Kolliker's authority, I cannot accept this explanation. My specimens leave no room for doubt that whatever may be its morphological importance, the neural crest is a perfectly continuous structure, extending in the chick at the twenty-ninth hour from the anterior end of the optic vesicles nearly to the end of tiie hindbrain. 1 am acquainted with folds such as Prof. KoUiker describes, but have only met with them in imperfectly-prepared specimens, and especially in specimens hardened in chromic acid, which, in my hands, at least. Las always proved a peculiarly unreliable hardening agent. ' Ballour, ' Elasmobraiich Fishes,' jip. I'Jl, l'J2, .
I have nothing to add to the arguments already given, though I am fully aware that the point is not yet proved. In the Elasmobranchs, the only other vertebrates in which the presence of a neural crest has been accurately described,* the anterior limits of this crest have not been fixed with certainty.
3. The shifting down of their roots of origin, one of the most striking features of the segmental nerves, is a very constant and well-marked point in the development of the olfactory nerves. It is well shown for birds in figs. 10 and 11, and for the dogfish in figs. 19 and 20. In the dogfish the displacement of the roots is less extensive than in the chick — a point obviously correlated with the greater development of the cerebral hemispheres in the latter.
4. The course of the segmental nerves in their early stages is, speaking within certain limits, at right angles to the longitudinal axis of the head at their point of origin. The facial and the postauditory nerves arise from a part of the head in which this axis is a straight line ; the nerves consequently run parallel to one another, as is seen in figs. 4 and 6. In front of the origin of the facial nerve the axis of the head is, owing to cranial flexure, no longer a straight line, but a curved one. The trigeminal nerve is disturbed only to a very slight extent, but it will be seen in fig. 4 that instead of running parallel to the facial, the two nerves converge slightly towards their distal ends. In the region of the midbrain the effects of cranial flexure are very well marked ; but fig. 6 shows that the course of the third nerve, the segmental nerve arising from the midbrain, is still at right angles to the longitudinal axis at its point of origin. Since the direction of the axis at this point is almost at right angles to its original direction, so also the third nerve is seen to take a course almost at right angles to the facial or glosso-pharyngeal. Similarly, the course of the olfactory nerve is at right angles to the longitudinal axis of the head at its point of origin; and its direction is such that if cranial flexure were corrected and the head straightened out the olfactory nerve would run parallel to the third, trigeminal, facial, and other segmental nerves. I have investigated very carefully this point, which I am disposed to regard as of some importance, and find that in the chick, at a time when cranial flexure has attained its maximum development, the angle formed by producing the direction of the olfactory nerve and of the facial or glosso-pharyngeal nerves until they meet, is almost identical with that which measures the amount of cranial flexure ; the angle in either case being about 120°. The course of the olfactory nerve in dogfish embryos is shown in figs. 17 and 18, and in the salmon in figs. 33 and 34.
5. This is a point of comparatively little importance, inasmuch as in the embryo ganglia, or local accumulations of nerve-cells, appear to be developed in a very irregular manner, and at very various points in the course of the nerves. Still it is a point not altogether destitute of weight, since those cranial nerves which appear for other reasons to have no claim to rank as segmental, are also peculiar in not possessing ganglionic enlargements at or near their roots of origin in the early stages. In the chick these ganglia are shown for the olfactory nerves in figs. 7, 8, and 9 ; for the third nerve in fig. 6 ; and for the trigeminal in fig. 4. In the dogfish the ganglia of the olfactory nerves are shown in figs. 20 and 21.
6. The discussion of the question whether the olfactory nerve is related to a visceral cleft in the same manner as the segmental nerves are to their respective clefts, will find a more suitable place after the development of the olfactory organ has been considered.
The distance between the root of the fifth nerve and that of the third is somewhat greater than that between the fifth and the facial, while that between the third and the olfactory is greater still. These facts, which are obviously correlated with the great hypertrophy of the anterior part of the brain, from which the nerves in question spring, can certainly not be used as arguments against the segmental nature of the olfactory nerve.
Though the olfactory nerve, from the earliest period at which it is recognisable as such, is thus seen to agree with the segmental nerves in all essential characters, it yet presents one or two minor points of difference. In the first place, owing to the close proximity of the forebrain to the nasal pit, the olfactory nerve is shorter than the other cranial nerves at the same age. Secondly, the olfactory appears to lag behind the others in development ; thus, at a time when the other nerves are fibrillar along the greater part of their length., and only present nerve cells in any considerable number at certain points, the olfactory nerve still presents nerve-cells along its whole length. This second difference appears, however, to depend on the first, since as the nerve elongates with age, we find it gradually taking on the histological characters of the other nerves, i.e. the greater part of its length becomes fibrillar, and the nerve-cells confined to the two extremities, where they form ganglionic swellings. The practical importance of these differences is, however, considerable, since, ovving to the olfactory nerve consisting for some time after its first appearance almost entirely of rounded cells, it is very difficult to distinguish from the surrounding mesoblast, and may, therefore, very readily be overlooked.
The olfactory nerves are by most authors considered as of totally different morphological value to the other cranial nerves.
According to Gegenbaur, "the cerebral nerves are seen to break up into two very distinctly marked divisions, when examined after the comparative method. One division, the larger, contains nerves which more or less agree with, or might even be derived from, spinal nerves, while the other contains those which have not the fai7itest resemblance to spi?ial nerves. This latter divison contains two specific sensory nerves, the olfactory and the optic. "^
Again, Prof. Huxley says, The greatest number of pairs of nerves ever given off from the vertebrate brain is twelve, including the so-called olfactory nerves and the optic nerves, which, as has been seen, are more properly diverticula of the hraiyi than nerves in the proper sense of the icord. The olfactory ' nerves ' {olfactorii) constitute the first pair of cerebral nerves. They always retain their primarxj connection with the cerebral hemispheres, and frequently contain, throughout life, a cavity, the olfactory ventricle, which communicates with the lateral ventricle."^
Finally, Balfour considers that the "very late appearance and peculiar relations" of the olfactory nerve " are, at least for the present, to my mind sufficient grounds for excluding it from the category of segmental cranial nerves. "^
1 have already attempted to show that the existence of an olfactory lobe or vesicle can in no way be s^^ud to militate against the establishment of a complete homology between the olfactory and the other cranial nerves. A structure that does not make the slightest appearance till the seventh day in the chick and stage o in the dogfish ; a structure that, in the chick, does not appear till long after the nerves have acquired their connection with the cerebral hemispheres, a connection which I must repeat is a purely secondary one, and not, as Prof. Huxley would have it, primary ; such a structure can hardly be deemed of sufficient morphological importance to outweigh the very obvious and striking resemblances between the olfactory and the other cranial nerves to which I have already referred.
1 'Elements of Comparative Anatomy,' English translation, p. 515. The italics are mine.
2 'Anatomy of Vertebrated Animals,' p. 7i. The italics again are mine.
3 Op. cit., p. 215.
.
Again, if my observations are correct, the olfactory nerves cannot be said to appear *' very late ;" while, if I may assume that I have fairly disposed of the olfactory vesicle difficulty, 1 fail to see what are the "^ peculiar relations" of the olfactory nerve that would justify its exclusion " from the category of segmental cranial nerves." .
The condition of the central nervous system appears to me to afford evidence of some value in favour of the ses'mental nature of the olfactory nerve. There is certainly no obvious reason why the anterior cerebral vesicle, or forebrain, of the embryo should be cojisidered to be of a different nature to the middle cerebral vesicle, or midbrain, or to any one of the vesicles of the hindbrain. The early embryonic stages afford no evidence whatever of a break of any kind between the fore and midbrains ; and, if the nerves arising from the mid and hindbrains have segmental value, there is surely a presumption in favour of the nerve that takes its origin in the forebrain having a similar and equivalent value j a presumption greatly increased in probability by the close similarity between the early stages of development of that nerve and of the nerves arising further back in the brain.
It still remains to be considered what is the morphological import of the olfactory lobe or vesicle ; but this is a question to which, in the present state of our knowledge, any answer that may be given must partake very largely of a speculative nature. The principal facts we have to guide us appear to be :
1. The very late appearance of the olfactory lobe.
2. The fact that though the olfactory lobe is obviously connected with the root of origin of the olfactory nerve, yet it has no relation to the original position of the root of the nerve, and does not appear till this root has acquired a new, and purely secondary position.
3. The fact that the olfactory lobe does not appear at equivalent periods in the development of different vertebrates. In the dogfish the olfactory lobe appears before the division of the forebrain into cerebral hemispheres takes place ; in the chick not till long after the appearance of the cerebral hemispheres ; and in the salmon, at any rate, not till after the time of hatching.
these facts would appear to indicate that the olfactory lobe is to be viewed rather as an adult or adaptative than as an embryonic or primitive structure ; a view that is materially strengthened by the great variations in relative size of the three elements of the olfactory nerve in various adult vertebrates, to which attention has already been directed.
One of the most remarkable features of the early stages of development of all vertebrates, is the enormous preponderance of the central nervous system to which at first everything appears to be subordinate, and which exercises a most important influence on the shape of the embryo. The rapid growth of the neural surface causes the body to become curved towards the ventral surface ; this curvature is naturally most marked at the free extremities of the body, and at the head end is the main, if not the sole, cause of cranial flexure. Owing to this cranial flexure the forebrain gets carried in front of the olfactory sacs, and, consequently, the olfactory nerves, which, as we have seen, acquire their connection with the olfactory sacs at a very early age, at first run in a direction downwards and backwards. Vide figs. 2, 15, 17, and 18.
Having attained this enormous relative development the nervous system stops for a while, and the face begins to grow more rapidly, causing the so-called rectification of the cranial flexure ; the olfactory sacs get carried further and further forwards, so that the olfactory nerves, instead of running downwards and backwards, now run directly downwards, or downwards and outwards as seen in fig. 33. The face still continuing to grow rapidly, while the brain undergoes little or no increase in length, the olfactory sacs get carried in front of the forebrain, so that the olfactory nerves now run downwards and forwards. A continuation of this process carries the olfactory sacs still further forwards, to an extent varying much in different vertebrates, so that the olfactory nerves ultimately run directly forwards as in most adult vertebrates. Vide fig. S6.
All the nerves of the body undergo during their development a considerable lengthening, owing to the gradual separation of their central and peripheral ends ; but while in the case of all the other nerves this is a gradual and continuous process, commencing with their earliest appearance, the olfactory nerves are somewhat peculiarly situated. In their early stages, owing to the close proximity of the olfactory sacs to the brain, the olfactory nerves are exceptionally short; and, owing to their origins being at first further forward than their insertions, the growth forwards of the face, carrying the olfactory sacs with it, does not at first cause any lengthening of the olfactory nerves. It is not till the sacs get in front of the forebrain that any lengthening is necessary, but no sooner does this occur than a sudden call is made on the olfactory nerves, which, previously quiescent, now have to commence growing rapidly in length and to continue so doing.
I would therefore suggest, without wishing to attach too much weight to the suggestion, that this elongation of the olfactory nerve, occurring under these exceptional conditions, may take place partly at the expense of the nerve itself, and partly at the expense of the brain ; and that it is in this way that the olfactory lobe is produced. It is certainly worthy of notice that in the two types — chick and dogfish — in which I have ascertained with precision the date of its first appearance, the olfactory vesicle comes into existence just about the time that the most rapid growth of the nose and snout occurs, and consequently just about the time when a sudden and rapid lengthening of the olfactory nerve becomes necessary. It is also a significant fact that the olfactory lobe grows very rapidly at first, the nerve itself remaining nearly stationary.
The above suggestion renders it easily intelligible that much variety should exist as to the relative lengths of the nervus and tractus olfactorius, even in nearly allied vertebrates; while it is quite possible that, at any rate in some forms, the skeletal elements may have an important share in determining the relative growth of nerve and brain.
II. The Olfactory Organ
a. Development of the olfactory organ
The consideration of the olfactory nerve having taken up far more space than I had originally anticipated, I shall be compelled to deal with the olfactory organ in a somewhat more summary fashion. The points to which I wish here to call attention are the remarkable resemblances that exist between the olfactory pits and the visceral clefts. As in the first part of the paper I shall deal first with matters of direct observation, and afterwards consider the theoretical side of the subject.
The vertebrate olfactory organs make their first appearance as a pair of slight thickenings of the external epiblast on the under surface of the forebrain, immediately in front of the mouth. . . . Each thickened patch of skin soon becomes involuted as a shallow pit."^
In the dogfish these thickenings appear " during a stage intermediate between L and k" (Balfour). Their condition during stage k is well shown in figs. 13 and 14 {olf.) ; the former figure being a longitudinal and horizontal section, the latter a vertical and transverse one. The exceedingly close proximity of the bottom of the olfactory pit to the brain is well shown by both figures. Fig. 13 shows also that at a time when the nose is in a very rudimentary condition, the eye (o. c.) has already made considerable progress in development, a point to which Balfour has already directed attention.
The communication between the visceral clefts and the exterior is established almost simultaneously with the first appearance of the olfactory pits. At stage L there are " three visceral clefts, none of which are as yet open to the exterior."" At stage k, according to Balfour, '^^ four visceral clefts are now visible, all of which are open to the exterior, but in a transparent embryo one more, not open to the exterior, wotild have been visible behind the last of these. "^ The visceral clefts, then, first become open to the exterior between stages L and k, and we have already seen that it is between these same two stages that the thickenings of the epiblast appear which form the earliest rudiments of the olfactory pits.
In the chick the early stages of development of the olfactory pits closely resemble those just described in the dogfish. Fig. 1 represents a longitudinal and horizontal section through the head of a fifty-four hours' chick embryo ; the section, which may with advantage be compared with fig. 13, shows on the right side the olfactory pit (o//".), formed by the thickened and involuted epiblast, and in close proximity to the forebrain (f. h.) ; on the left side the section, which is a little oblique, passes through the thickened epiblast forming the margin of the olfactory pit, and through the eye (o, c). Two visceral clefts {v. c.) are shown, both open to the exterior.
The earliest period in the chick at which I have noticed the thickening of the olfactory epithelium is about the forty-eighth hour ; a period almost identical, as in the dogfish, with the opening of the visceral clefts to the exterior.
Balfour, op. cit., p. 184.
2 Balfour, op. cit., p. 77.
3 Qp cit., p. 78.
In the trout the mode of development of the olfactory pits corresponds very closely with that occurring in the chick and dogfish ; and, as in these two types, their first appearance coincides almost exactly with the opening of the visceral clefts to the exterior.
The connection between the olfactory nerve and the bottom of the olfactory pit is, as already noticed, acquired exceedingly early, very shortly indeed after the appearance of the latter. The condition of the olfactory organ in the dogfish is shown at stage m in figs. 15, 16, and 19; and at stage o in figs. 17, 18, and 20. In the chick the olfactory organ is shown at the sixty-fourth hour in fig. 2, at the sixty-seventh in fig. 3, and at the ninety-sixth hour in figs. 5 and 6.
Throughout their early stages of development the olfactory organs present a striking resemblance to the visceral clefts, both in form, position, and general relations — a resemblance •which it will be necessary to consider in some detail, inasmuch as it has been very generally overlooked hitherto.
Fig. S represents a longitudinal and vertical section through the head of a chick embryo at the sixty-seventh hour. The section, which is taken in a plane not far from the surface, passes through the hind, raid, and forebrains, through the auditory vesicle (aucL), the eye (o.c), the trigeminal (v), and auditory (viii) nerves, through the anterior visceral clefts and arches, and through the olfactory pit (olf.). The olfactory pit is seen to bear a marked resemblance to the visceral clefts : like them it is situated on the ventral surface of the head ; it is open below ; its axis is at right angles to the longitudinal axis of the head, so that were the head straightened out it would be parallel to the clefts ; and its general appearance and relations are such as to strongly suggest the view that it is one of the same series of structures as the visceral clefts. It is indeed separated from the next cleft, that in front of the maxillary arch (Mz.), by an interval somewhat greater than that separating the hinder arches from one another ; but when we consider the enormous hypertrophy which the part of the brain with which it is connected has undergone, this becomes rather an argument in favour of than against the comparison.
Figs. 4, 5, and 6 are three sections taken from the same embryo, a ninety-six hours' chick. Of these sections, which are taken in a longitudinal vertical plane, that given in fig. 4 is the most superficial, that in fig. 6 the deepest of the three. Figs. 5 and 6 are drawn from consecutive sections, but between figs. 4 and 5 two sections intervened. These figures illustrate well the points to which I have just called attention ; they show that the visceral clefts form a continuous series of structures, of which the most anterior is, not the mouth cleft (between Mn. and 3fx.), but the cleft in front of the maxillary arch ; a cleft that, following Prof. Parker, T propose to speak of as the lachrymal cleft : they show further that just in front of the lachrymal cleft is the olfactory pit {olf.), and that the relations of the pit are such as to inevitably suggest that the olfactory organ is one of the same series of structures as the visceral clefts. Correct the cranial flexure, and straighten out the head, and the resemblance would amount almost to identity. I have only to add that, though these figures are semi-diagrammatical, yet as far as the outlines go, which alone concern us at present, they are as absolutely accurate as I have been able to make them.
The resemblance between the olfactory organ and the visceral clefts is quite as marked in the early stages of the dogfish as in the chick; but to convey anything like an adequate idea of it would require a much more extensive series of figures than I am able to give here.
Fig. 16 is taken from a longitudinal and vertical section through the head of a dogfish embryo at stage m. The section which is taken very near to the surface passes through the auditory vesicle (aud.), parts of the trigeminal, facial, auditory, glosso-pharyngeal and vagus nerves, the second head cavity {h. 2), the eye (o.c), the mandibular, hyoid, and first four branchial arches, as well as through the olfactory pit {olf.). The section is a little deceptive, inasmuch as, owing to the head being somewhat constricted just behind the eyes, the buccal and lachrymal clefts do not appear at all, while the constriction just referred to presents somewhat the appearance of a visceral cleft between the olfactory organ and the mandibular arch, and might possibly be mistaken for one on a superficial examination. The figure illustrates well the resemblance between the olfactory organ, which is larger than at a corresponding stage in the chick, and the visceral clefts.
Fig. 18 shows the same parts in a dogfish embryo of stage o; as in the preceding figure, and for the same cause, the buccal and lachrymal clefts do not appear in the section, but the figure shows the general resemblance in position and relations that exists between the olfactory organ and even the hinder visceral, or branchial, clefts.
In connection with this point, the study of whole embryos affords evidence fully as striking as that yielded by sections. I would here refer especially to certain of the figures given by Professor Parker in his monograph on the " Structure and Development of the Skull in Sharks and Skates," published in the ' Transactions of the Zoological Society ' for 18T9 : on Plate XXXIX side views of the heads of embryos of Raia maculata are given (figs. 1 and 2), in which the resemblance between the slit-like aperture of the olfactory organ and the gill slits is shown with remarkable distinctness. The direction of the slit forms an angle of about 120° with the hyomandibular or spiracular cleft, which angle is almost exactly that made by the longitudinal axis of the forebrain with that of the hind brain, i.e. is the amount of cranial flexure ; hence, but for cranial flexure, the external slit-like aperture of the olfactory organ would be parallel to the perf^-ctly similar gill slits.
The figures of dogfish embryos of stages K and L, given by Mr. Balfour on Plate VII of his monograph on Elasmobranch Fislies, illustrate the same points. The reference to these two works acquires additional weight from the consideration that the figures which I have named were drawn, I have reason to believe, without the slightest intention on the part of the authors to direct attention to the resemblance. Even in an adult skate the similarity between the olfactory organ and the gill slits is sufficiently striking.
The same points appear, if possible, still more clearly in axolotl and salmon embryos, especially in the former. I have found, however, that to give any adequate representation of these would require a large number of figures, which figures would also serve to illustrate other points in the development of the axolotl, which I hope to deal with on some subsequent occasion.
Fig. 30 represents a longitudinal and vertical section through the head of a trout embryo on the thirtieth day after fertilisation of the ova. The section passes through the hind and midbrains and the eye (o. c). The ventral surface of the section presents a series of undulatory folds, corresponding to the bases of the visceral arches, with their intervening clefts. The olfactory pit {olf.) is seen to form the most anterior of these depressions, and to differ from the hinder clefts in little but the greater thickness of its epithelium, and the somewhat greater interval between it and the next cleft. The cleft next but one to the olfactory pit is that over which the trigeminal nerve forks — i. e. the buccal or mouth cleft ; it is situated between the maxillary (Mx.) and mandibular (Mn.) arches. Behind the mandibular arch, between it and the hyoidean arch, is the cleft, the two sides of which are supplied by the facial nerve. Between the buccal cleft and the olfactory pit a cleft intervenes — the lachrymal cleft; so that the number of clefts in the trout agrees completely with that we have already found in the chick.
The resemblances between the olfactory pit and the visceral clefts are, however, not simply those of general appearance and relations ; they are of a far more intimate nature, and extend even into the details of histological structure. For studying these more intricate relationships the dogfish has proved the most suitable.
The olfactory organ of a dogfish does not long remain a simple pit; very soon after its first appearance its walls become thrown into a series of folds — the rudiments of the Schneiderian folds of the adult. I wish here to call attention to the resemblances between these folds and the series of folds which, arising from the sides of the visceral clefts, form the rudiments of the gills.
I have not myself observed the presence of the rudimentary Schneiderian folds in embryos younger than stage M, but Balfour has shown that they not only exist, but have acquired the characteristic adult arrangement in embryos *' a little older than k."i With regard to the gills, Balfour's description is as follows : — " Towards the close of stage k there arise, from the walls of the second, third, and fourth clefts, very small knob-like processes, the rudiments of the external gills. These outgrowths are formed both by the lining of the gill cleft and by the adjoining mesoblast."^ If, indeed, the times of appearance be not absolutely identical in the two cases, the correspondence is, at any rate, sufficiently striking.
Fig. 23 is a horizontal and longitudinal section through the head of a dogfish embryo at stage n, magnified twenty diameters ; it passes through the fore and hind brains, the notochord (w), the eyes (o. c), the oculo motor (iii) and trigeminal (y) nerves, and through the olfactory pits (olf.). The bottoms of these j)its are seen to be thrown into a series of small equidistant folds — the Schneiderian folds.
Fig. 24 is a transverse section through the body of the same embryo, taken a short way behind the head, and passing through one of the branchial arches on either side. The section which, like the preceding one, is magnified twenty diameters, sliows also the spinal cord, with the anterior and posterior roots of a spinal nerve, the notochord (n.), muscle plates {m. p.), pharynx {al.), parts of the vagus nerve, the cardiac and. dorsal aortse, and on either side the branchial arteries {h. a.) The free surface of each branchial arch presents a series of small equidistant folds, the rudiments of the gills {g.), which, even under this low magnifying power, have a close resemblance to the Schneiderian folds.
' Op. cit., p. 184, and Plate XIV, fig. 14. Op. cit., p. 211.
In order to show this resemblance more satisfactorily I have given figures of the parts in question on a larger scale. Fig, 25 represents the right olfactory pit of fig. 23, and fig. 26 the left gill of fig. 24; both figures are thus taken from the same embryo, and the magnifying power employed — ninety diameters — is the same in the two cases.
These figures show that the correspondence between the two structures is by no means confined to their coarser anatomy, but extends even to histological details. The folds are seen to be in the two cases — gills and Schneiderian folds — of the same width, and the same distance apart; in both cases, though consisting mainly of epithelium, they yet involve the underlying mesoblast to a certain, though slight, extent,^ but as nearly as possible to the same extent in the two cases. The epithelium that forms the greater part of the folds is of the same thickness in the two cases, and of the same histological character, consisting mainly of columnar cells in close contact Avith one another, and arranged, as a rule, in two rows.
The same folds are shown, at a somewhat later period, in figs. 27 and 28, the former representing the Schneiderian folds, the latter the gills of the same embryo. Though the resemblances are still strong, there are now well-marked differences between the two structures ; thus, in fig. 27 the epithelium is somewhat thicker than in the gills, while the mesoblast enters more largely into the gills than the Schneiderian folds. Most of the gill folds already present a central blood-vessel ; it is very difficultto satisfy oneself of the existence of distinct walls to these blood-vessels, which appear in many cases to be simply channels in the mesoblast forming the axis or core of each gill fold. Similar blood-vessels exist, especially at a rather later stage, in the Schneiderian folds, and their relations are similar to those in the gills.
Even in adult Elasmobranchs the Schneiderian folds resemble the gills closely in their great vascular supply, in the arrangement and distribution of the blood-vessels, and in the characters of their surface epithelium.
^ Balfour notes this in the case of the gills, but describes the Schneiderian folds as folds of epithelium. Op. cit., p. 184.
b. General considerations
Hitherto we have been concerned simply with matters of observation ; though, indeed, 1 have not attempted to give a complete account of the development of the olfactory organ, but have limited my description to certain developmental features, in which it strongly resembles the visceral clefts ; still I have dealt simply with facts, or what I believe to be facts. I propose now to consider the subject from a more theoretical point of view.
In the first place I would submit that the very close resemblance as to form, structure, general relations, time of appearance, &c., existing between the olfactory organ and the gill clefts, whether these be considered as wholes or in their separate parts, is sufficient to raise a strong probability that they are homologous structures.
This probability is strengthened by the complete absence of similar structures in any other part of the body at any period of development. Not only do the Schneiderian folds and the gills appear at the same time and agree completely in structure, but in no other part of the body do similar structures occur, either at this or any other period.
Again, this probability gains very material support from the conclusion arrived at in the first part of this paper, viz. that the olfactory nerve is a segmental nerve ; for we hav« seen that one of the most important diagnostic characters of a segmental nerve is its distribution to the two sides of a visceral cleft, and, since the olfactory nerve is distributed to the olfactory organ, and to that alone, if there be a visceral cleft with which it is in relation, the olfactory organ must be that cleft.
The conclusions, then, to which I have been led concerning the morphology of the vertebrate olfactory organ are — that the olfactory organ is the most anterior visceral cleft ; that the olfactory nerve is the segmental nerve supplying the tioo sides of that cleft i?i a manner precisely similar to that in which the hinder clefts are supplied hij their respective nerves ; and that the Schneiderian folds are homologues of gills.
The suggestion that the nasal organs are gill clefts was originally made by Dr. Dohrn, in his essay on the origin of vertebrates. In discussing the question whether the pair of gill clefts, which by their median fusion formed the vertebrate mouth, was the most anterior pair, Dr. Dohrn says : " Aber auch betreflfs der vorderea Kiemenspalten ist noch die Vermuthungzu aussern, ob nicht vielleicht in den Nasengruben ein Paar solchen Spalten^ freilicfi in wesentlich veianderter Function und, darum audi Structur, zu erkennen sei."[15]
Dr. Dohrn does not enter into any details concerning the suggestion thus made, and does not discuss the question of the olfactory nerve. I am not aware that he has since published any further observations on this point. The suggestion appeared to me, if not untenable, at any rate unprovable, so long as the ordinary account of the development of the olfactory nerve continued to find acceptance.
In addition to what has been already said there are, I think, many arguments in favour of this view. Even if we leave out of consideration the buccal and lachrymal clefts, it is well known that in all vertebrates above Amphioxus more or fewer of the visceral clefts undergo modification to a greater or less extent, and that this modification is first felt by the clefts at the two extremities of the series, especially by the anterior ones ; and it is a point worthy of notice that, while the posterior clefts tend simply to disappear, the anterior clefts with their gills are peculiarly prone to persist in a modified form. Thus, the first postoral or hyomandibular cleft is the only one which remains in Sauropsida and Mammalia. Among Ichthyopsida this cleft is apt to assume the modified form of a spiracle, while its gill loses its respiratory function, and persists as a pseudobranchia. Similarly, the carotid gland of the frog and the choroid gland of Teleosteans are probably other instances of the persistence, in an altered condition, of anterior gills. On the other hand, when reduction is effecte<l in the number of the gills from the posterior end of the series, as in nearly all fishes, the gills and their clefis usually disappear absolutely and completely.
Again, if the olfactory organ is a gill, we should expect to find the resemblance between the two structures strongest in the most primitive vertebrates. From what has been said already this obviously is the case. Of the various types of vertebrates examined it is in the dogfish alone that we find the intimate relation between the development of the gills and that of the Schneiderian folds.
Whatever view we may hold as to the ancestry of vertebrates, there can be little doubt that they have not inherited their olfactory organ as such. At any rate, we know as yet of no invertebrates that possess olfactory organs from which the vertebrate olfactory organ could possibly have been derived by inheritance. Hence it follows either that vertebrates must have acquired or developed an olfactory organ completely de novo, or else that their olfactory organ has been formed by gradual modification of some pre-existing structure with accompanying change of function.[16] The first of these alternatives may, I think, be at once dismissed as untenable, and then we are left with the second alternative.
I do not propose to enter here into a detailed discussion of the physiology of smell, but will only remark that what little we do appear to know definitely about it is quite in accordance with the view that smelling is only a modified form of breathing, and that no very violent physiological change would be necessary to convert a gill into an olfactory organ. On the other hand, the sense of smell is something of a totally different nature to sight or hearing ; the essence of these latter consists in the appreciation of the relative wave lengths of undulations conveyed by air or ether ; while smell appears to be due to direct chemical action on the nerve-endings, requiring the presence of free oxygen (Graham) .
In the first part of this paper I have attempted to show that the cranial nerves afford very definite evidence as to the segmentation of the anterior part of the vertebrate head — evidence, indeed, quite as definite as that which they have long been recognised as affording concerning the hinder part of the head. The second part of the paper has shown that the visceral clefts afford equally definite evidence on the same point. In a former paper[17] I have called attention to the fact that the early stages of the brain also afford evidence on this point.
It is a matter of considerable interest that the evidence yielded by these three types of structures respectively, as to the number and situation of the cephalic segments, is identical.
The brain consists in an early stage of a series of vesicular dilatations separated by slight constrictions ; of these vesicles the most anterior is the forebrain, and the next the midbrain ; while the succeeding vesicles, which form a series decreasing in size from before backwards, and of which the first two at any rate appear to possess considerable constancy, are spoken of collectively as forming the hindbrain.
From each of these brain-vesicles a segmental nerve arises : the forebrain gives origin to the olfactory nerve ; the midbrain to the third or oculomotor nerve;[18] from the anterior vesicle of the hind brain the fifth or trigeminal nerve arises, and the second hindbrain-vesicle appears to give origin fairly constantly to the seventh or facial nerve ; behind this I have not observed any definite relation between the cranial nerves and the brain-vesicles, which latter become very small and of doubtful constancy as to number and relations. This is, however, a matter of little moment, as the evidence afforded by the cranial nerves themselves and by the visceral clefts in the postauditory part of the head is of an unimpeachable character.
As to the clefts, the most anterior is the olfactory cleft ; vide figs. 3, 4, 5, and 6. Next to that is the cleft which, following Prof. Parker, I have spoken of as the lachrymal, i.e. the cleft in front of the maxillary arch : the relations of the third nerve to this cleft are well shown in fig. 6; it only remains to be added that, of the two branches into which the third nerve divides beyond the distal ganglionic swelling shown in fig. 6, one lies at first behind this cleft, the other in front of it.
We next come to the buccal or mouth cleft, between the maxillary and mandibular arches ; the relations of the fifth nerve to this cleft are well known. Behind this is the hyomandibular or spiracular cleft, supplied by the facial nerve ; and then the branchial clefts, supplied by the glossopharyngeal and by the several branches of the vagus.
These relations are exhibited more clearly in the following table. In the first column are the numbers of the segments, of which, if we count seven branchial clefts, the full vertebrate number, there are eleven in all; two of these being preoral, and eight postoral. The second column contains the brain vesicles, the third the segmental nerves, and the fourth the corresponding visceral clefts.
| Head Segments of Vertebrates | |||
|---|---|---|---|
| Segment | Brain-vesicle | Nerve | Cleft |
| Preoral 1 | Forebraian | I. Olfactory | Olfactory |
| Preoral 2 | Midbrain | III. Oculomotor | Lachrymal |
| Oral 3 | Hindbrain, 1st vesicle | V. Trigeminal | Buccal |
| Postoral 4 | Hindbrain,2nd vesicle | VII. Facial | Spiracular or hyomandibular. |
| Postoral 5 | Hindbrain | IX. Glosso-pharyngeal | 1st branchial. |
| Postoral 6 | Hindbrain | X. Vagus, 1st branch | 2nd branchial. |
| Postoral 7 | Hindbrain | X. Vagus, 2nd branch | 3rd branchial. |
| Postoral 8 | Hindbrain | X. Vagus, 3rd branch | 4th branchial. |
| Postoral 9 | Hindbrain | X. Vagus, 4th branch | 5th branchial. |
| Postoral 10 | Hindbrain | X. Vagus, 5th branch | 6th branchial. |
| Postoral 11 | Hindbrain | X. Vagus, 6th branch | 7th branchial. |
This table, which has been modelled after the one given by Balfour,[19] differs from this latter in some important points. The differences are most marked in the anterior part of the headj where I have added an olfactory segment, and have attempted to define more accurately the constituents of the second segment by removing the fourth and sixth nerves, and assigning, for reasons discussed elsewhere, the third nerve as the true segmental nerve. At the hinder end of the head I have added two segments for the two hinder branchial clefts of Notidanus and the Marsipobranchii, so that the table is intended to include the full number of head segments of which we have any definite indication in any vertebrate, excepting Amphioxus ; though it is by no means intended to exclude the possibility of additional segments having existed at the hinder end of the head in former vertebrates, or actually existing in some living forms.
It will further be noticed that I have placed the visceral clefts, and not the visceral arches, as indicating segments ; this is a point of some importance, but one which I do not think we are yet in a position to decide with certainty. The question is, whether the visceral clefts are to be viewed as intersegmental, i. e. as corresponding in position to the lines of separation of the original segments by the fusion of which we suppose the vertebrate head to have been formed; or whether they should be considered as inirasegtnental, as apertures formed in the substance of the segments ; in other words, whether the visceral clefts are formed between successive segments or through the middle of the segments.
Though the former of these views is usually assumed to be the true one, yet there are, I think, considerations of some weight in favour of the opposite view. In the first place, we must bear in mind that the original proto -vertebral segmentation does not extend to the head, and that the secondary, visceral cleft segmentation appears late, and differs totally from the segmentation of the body, inasmuch as, instead of starting on the dorsal side in the mesoblastic tissue on either side of the neural axis, it arises in, and is limited to, the lateral and ventral walls of the alimentary canal. Instead of arising primarily in the mesoblast, it is a segmentation in which the mesoblast takes no share whatever, except a purely passive one; it is a segmentation produced by ilie growth of diverticula from the alimentary canal, which come in contax't with, and fuse with the external epiblast, and, finally, by perforation of this latter open on to the exterior. Now, It is more in accordance with what we know of the occurrence of lateral diverticula of the alimentary canal iu Inveitebrata, that these should be segmental rather than intersegmental.
Again, while each cranial segmental nerve supplies two visceral arches, it only supplies one cleft; and, from the analogy of Invertebrata, we should expect that the distribution of each nerve would be to its own segment ; while it certainly would be a very remarkable fact that each segmental nerve should supply adjacent halves of two segments.
The distribution of the branchial arteries may also be cited as additional evidence in the same direction ; the corresponding vessels in Invertebrata do not occupy the middle of each segment, but follow the intersegmental septa, which septa, according to the view here advocated, would occupy the centres of the vertebrate visceral arches, and so correspond in position with the branchial arteries. On the same view the skeletal elements of the visceral arches would also correspond in position to the intersegmental septa, from which, indeed, they may conceivably have been derived.
This view acquires some additional interest in connection with Ur. Dohrn^s suggestion that the visceral clefts are homologues of segmental organs.[20]
The theory above propounded as to the morphology of the vertebrate head, will, I venture to think, throw some light on the nature of the skeletal elements of the head. I will here only notice briefly one or two points of importance. On the yiew here put forward the trabeculce cranii, which lie at the base of the brain, run parallel to the longitudinal axis of the head, and at right angles to the segmental structures, such as the third and olfactory nerves and the corresponding clefts, must be regarded as axial structures, and not as arches, whether neural or haemal.[21]
Again, my investigations appear to leave no room for doubt that the maxillary arch, the rudiment of the upper jaw, is as fully entitled to rank as a distinct visceral arch as the mandibular, hyoid, or branchial arches. Figs. 3 — 6 appear to me to afford conclusive evidence on this point.
The morphological nature of the labial cartilages has been matter of much dispute ; if the determination of the olfactory organ as a gill cleft be accepted, those at least of the labial cartilages which are grouped round the external aperture of the olfactory organ, and very possibly those* also in connection Avith the gape of the mouth, would appear to be homologues of the extra-branchial cartilages, a suggestion in which I find I have been anticipated by Professor Parker.[22]
I cannot refrain here from referring to the remarkable manner in which the views here put forward agree with the results arrived at by Professor Parker, and embodied in his latest paper .[23] He there expresses himself " satisfied that, in spite of the doubling up of the basis cranii, at the time of its greatest flexure, there are rudiments of three prseoral arches, related to ttvo prceoral clefts, the lachrymal and the nasal." " Thus we get four pre-auditory, and eight postauditory clefts, with their nerves ; if we add the twelfth (hypoglossal), of the ' Amniota,' we have obtained signs and proofs of thirteen cranial (segmental) nerves, all of these, except the last, forking over visceral clefts, and hedged in all hut the last by visceral bars. The first of the bars is in front of the first or tiasal cleft, the last, or thirteenth, is the hinder bar of the lamprey's branchial basket work." The italics in the above quotation are mine. Though I see no reason for regarding the hypoglossal as a segmental cranial nerve, this extract from Professor Parker's work shows that the study of the skeletal elements of the head leads to results almost identical with those at which I have arrived, and affords perhaps the strongest possible confirmation of these results.
Though in the above enumeration of the segmental cranial nerves I have left out the optic nerve, for reasons stated elsewhere,[24] it is quite possible that this nerve may ultimately prove to be of segmental value ; in which case it would indicate the existence of a cleft between the olfactory and lachrymal cleft. However, I have as yet completely failed to find any evidence of its segmental nature, and must, for the present, regard it as of a totally different nature to any of the other nerves. The case of the auditory nerve is very different, for there can be little doubt that this is to be viewed as merely a specialised branch of the facial.[25]
If the olfactory organs are really a pair of gill slits, then they must have originally communicated with the mouth cavity : and it becomes a matter of considerable interest to determine whether any traces of such a communication still exist. It is quite possible that the grooves which connect the nasal sacs with the angles of the mouth in the skate and other Elasmobranchs, and which form the rudiments of the posterior narial passages of higher vertebrates, are remnants of this communication. It is difficult to understand what function these grooves subserve in Elasmobranchs, and their apparently irregular presence or absence in closely allied genera would well accord with their being disappearing rudiments. In connection with this point some observations I have recently made on trout and salmon embryos, though incomplete, appear to possess some interest.
Fig. 31 is a transverse section through the anterior part of the head of a salmon embryo just about the time of hatching : the section passes through the anterior borders of the olfactory pits (o//".), through the cartilaginous plate formed by the fusion of the two trabeculse {tr.), and, on the ventral side, through a large flattened cavity {al\) ; this cavity is found, by a study of the sections in front of and behind the one figured, to be an anterior prolongation of the buccal cavity, extending forwards in front of the mouth, underlying the olfactory sacs, and reaching almost to the extreme anterior end of the head.
Fig. 33, which has been already described, is a section taken through the head of an embryo of about the same age as that in fig. 31, but a little further back; it shows this same cavity, which, however, is now not completely closed in the median ventral line, the section passing througli'the anterior part of the oral aperture.
In figs. 34 and 35 the same structure is shown in longitudinal and vertical section; fig. So, which is the more superficial of the two,^ shows the pharynx (al'.J, with the branchial arches and the anterior continuation of the buccal cavity {al'.) Fig. 34, which passes through the root of origin of the olfactory nerve, and therefore, as is evident from fig. 33, very close to the median line, passes also through the mouth; it shows very clearly the manner in which this anterior prolongation {al'} extends forwards in front of the anterior margin of the mouth,
I have unfortunately not yet succeeded in tracing the development of this prolongation, and do not even know for certain whether it appears before or after the formation of the mouth, or whether it is lined by hypoblast or epiblast.
At a stage a little later than that just described, when the growth of the anterior part of the head has carried the nose considerably farther forwards, this prolongation exists in the form of a pair of ccecal diverticula, stretching forwards from the anterior part of the buccal cavity towards the olfactory pits. These are well shown in fig. 32, a transverse section through the anterior part of the head of a salmon embryo about a week after hatching. The section passes through the extreme anterior end of the forebrain {f. b.) in front of the 'origin of the olfactory nerves, through the two eyes (o. c), the superior recti muscles (r. 5.), the trabecular plate (tt\), the hinder end of the two olfac tory pits {olf.), and the diverticula of the buccal cavity (aV.) close to their anterior terminations. At a stage a little later still, these diverticula appear to shrink and disappear; at least I have failed to recognize them in sections.
Whatever these diverticula may prove to be, their existence is certainly of some interest in connection with the visceral-cleft theory of the olfactory organ ; they show, at any rate, that there do exist diverticula of the alimentary canal towards the olfactory organs ; they may possibly be taken as indications of a former extension forwards of the alitnentary canal to the anterior end of the head ; while their paired condition, shown in fig. 32, may perhaps be an indication of relationship to the paired lateral diverticula of the alimentary canal, which form the rudiments of the hinder visceral clefts.
Again, if the olfactory organs are gill clefts and the Schneiderian folds gills, not only must these clefts have originally communicated with the buccal cavity, but the vertebrate mouth must originally have been in front of them. According to Dr. Dohrn the present vertebrate mouth is formed by the median coalescence of a pair of gill slits; my own investigations lead me to the conclusion that, though these gill slits do contribute to the formation of the mouth, there is in addition a median involution of the epiblast of the under surface of the head, as described by Balfour[26] and others, so that the mouth consists of three elements, a median epiblastic involution and a pair of gill slits. I am inclined also to believe that the fact of the olfactory organs appearing in front of the mouth is due to two causes ; firstly, the hypertrophy of the forepart of the head carrying the olfactory sacs forwards ; and, secondly, an actual shifting backwards of the median element of the mouth, of which I think there is a certain amount of independent evidence. The anterior end of the notochord is, as is well known, bent completely round on itself, through an angle of fully 180o, i. e. as Balfour has already noticed, to a much greater extent than cranial flexure alone will account for. Now, assuming that the notochord is a hypoblastic structure, and that its anterior end remains for a time in connection with the hypoblast, a shrinking back of the hypoblast of the anterior end of the foregut would at once account for this condition of the notochord, and would at the same time cause a displacement backwards of the mouth. It would appear therefore quite possible that the median element of the present vertebrate mouth is the original vertebrate mouth which has undergone a slight displacement backwards, and so has become severed from the olfactory organs.
Perhaps the most serious objection to the visceral-cleft theory of the olfactory organ that is likely to occur at first sight, is the fact that these organs are involutions of the external epiblast, while the visceral clefts are formed by diverticula of the hypoblast of the foregut. While fully admitting the force of this objection, I venture to think that the arguments I have brought forward — the evidence in favour of the segmental value of the olfactory nerve, the close relation, both anatomical and histological, between the olfactory organ and the visceral clefts, the fact that these relations are much more marked in the more primitive than in the more specialised vertebrates, the various identities in time of appearance and in histological structure, and the concurrent testimony of the various incidental circumstances to which I have alluded — are sufficient to outweigh this objection. Moreover, we must bear in mind that slight ingrowths of the external eplblast towards the hypoblastic outgrowtTrs of the pharynx may occur, and that it is still a matter of uncertainty whether some of the gills are not epiblastic rather than hypoblastic ;^ while, if the diverticula of the alimentary canal described above in the trout and salmon should prove to be hypoblastic, the principal differences between the olfactory organs and the gill clefts would be the gradual shrinking of the most anterior pair of diverticula of the buccal cavity, the ultimate failure on their part to reach the surface, and a corresponding exaggeration of the epiblastic surface involutions, which changes can readily be conceived as following on, and caused by, a slight displacement backwards of the mouth. Since the functional activity of the gills as such depends on the constant passage of a stream of water through the mouth into the buccal cavity and then out through the gill slits, it follows that if the mouth were changed in position so as to be situated behind instead of in front of, the first pair of gills, the function of these gills would be materially interfered with, while their position at the anterior extremity of the head and their consequent potential utility, would favour their preservation in a modified form, and with modified function.
Plates

Each figure has been drawn from a single section by the aid of a Hartnack camera. The numbers attached indicate, in diameters, the magnifying power employed in each case. The majority of the figures are of a semi-diagrammatical nature, the mesoblast being in nearly all cases omitted for the sake of clearness; the outlines are, however, strictly accurate in all cases. Figures 21, 22, 27, and 28 are drawn from sections kindly lent me for the purpose by Mr. Balfour ; the rest are from specimens of my own preparation.
Online Editor Note - original article had no images in the text, only appearing as figures in these two plates.
Figures
aC Alimentary canal. aV . Anterior prolongation of alimentary canal. aor. Dorsal aorta. a. r. Anterior root of a spinal nerve. mtcl. Auditory vesicle. b. a. Branchial artery. br. 1. First branchial arch. br. 2. Second branchial arch. c. h. cerebral hemisphere. f. b. forebrain, g. gill. h. 2. Second head-cavity. h . b. Hindbrain. hy. Hyoid arch. inf. Infundibulum, I. c. Lachrymal cleft. m. b. Midbrain. Mn. Mandibular arch. m. p. Muscle plate. 3f.v. Maxillary arch. u. Notochord. o. c. Optic cup or eye. olf. Olfactory pit. ol. V. Olfactory vesicle or lobe. r. i. Inferior rectus muscle. r. s. Superior rectus muscle. Sch. Schneiderian folds. sp. Spinal cord. tr. Trabecula3 cranii. v. c. Visceral cleft. I. Olfactory nerve. II. Optic nerve. III. Third or oculomotor nerve. V. Trigeminal nerve. V a. Opiithalmic branch of the trigeminal nerve, or ramus ophthalmicus profundus. V 3. Inferior maxillary branch of the trigeminal nerve. VII. Facial nerve. VII a. Ophthalmic branch of the facial nerve, or ramus ophthalmicus superficialis. VIII. Auditory nerve. IX. Glossopharyngeal nerve. X. Vagus or pneumogastric nerve.
Fig. 1. — Longitudinal and horizontal section through the head of a fifty-four hours' chick embryo, showing early stage in the formation of the olfactory pits, also the visceral clefts with their communication with the exterior. Picric acid, x 35 diameters.
Fig. 2. — Longitudinal and vertical section through the fore part of the head of a sixty-four hours' chick embryo, showing olfactory nerve and pit. Picric acid, x 27 diam.
Fig. 3. — Longitudinal and vertical section through the head of a sixty-seven hours' cliick embryo, showing the olfactory pit, the maxillary, man- dibular, hyoideau, and first three branchial arches, with the intervening clefts. Picric acid, x 20 diam.
Figs. 4—6. — Longitudinal and vertical sections through the head of a ninety-six hours' chick. Picric acid, x 15 diam.
Fig. 4. — The most superficial of the series ; shows the maxillary, mandibular and hyoidean arches, with their intervening clefts, and the trigeminal, facial, and auditory nerves.
Fig. 5 shows, in addition, the olfactory pit and the distal end of the olfactory nerve.
Fig. 6 shows the third, glossopharyngeal and vagus nerves.
Figs. 7 and 8. — Longitudinal and vertical sections through the cerebral hemispheres and nasal region of a six-day chiek embryo. Picric acid. X 15 diam.
Fig. 7 shows whole length of olfactory nerve.
Fig. 8 shows ganglion at root of origin of olfactory nerve, also internal projection of hemisphere at this point.
Fig. 9. — Longitudinal and vertical section through the nasal region of a seven-day chick embryo, showing whole length of olfactory nerve, and commencing rudiment of olfactory lobe. Picric acid, x 15 diam.
Fig. 10. — About half of a transverse section through the forebrain of a four-day duck embryo, showing the olfactory nerve and pit. Picric acid. X 50 diam.
Figs. 11 and 12. — Tranverse sections through the anterior part of the head of an eighty-hours' chick embryo, passing through the cerebral hemispheres, and the olfactory pits and nerves. Picric acid, x 27 diam.
Fig. 11 shows on left side the origin of the olfactory nerve.
Fig. 12 shows on right side the distal end of the olfactory nerve.
Fig. 13. — Longitudinal and horizontal section through the head of a dogfish embryo of stage k, showing early stage in development of the olfactory pits. Chromic and osmic acids, x 20 diam.
Fig. 14. — Transverse section through the anterior extremity of the head of a dogfish embryo of stage k, showing the forebrain, the olfactory pits and nerves. Chromic and osmic acids. X 20 diam.
Fig. 15. — Longitudinal and vertical section through the head of a dogfish embi-yo of stage m, showing the olfactory pit and nerve. Chromic and osmic acids, x 20 diam.
Fig. 16. — Longitudinal and vertical section through the head of a dogfish embryo of stage M, showing the olfactory pit, the mandibular, hyoid and branchial arches. Chromic and osmic acids, x 20 diam.
Figs. 17 and 18. — Longitudinal and vertical sections through the head of a dogfish embryo of stage o. Chromic and osmic acids, x 20 diam.
Fig. 17 shows olfactory pit and nerve.
Fig. I8 shows olfactory pit, with Schneiderian fold, olfactory nerve, forebrain, hyoid and branchial arches.
Fig. 19. — Transverse section through the forebrain of a dogfish embryo of stage M, showing olfactory pits and nerves, with absence of olfactory lobes. Chromic and osmic acids, x 27 diam.
Fig. 20. — Transverse section through the same region as in the preceding figure, but in a dogfish embryo at the commencement of stage o : shows olfactory pits with Schneiderian folds, olfactory nerve, and commencing olfactory lobe. Chromic and osmic acids, x 27 diam.
Fig. 21. — Transverse section through the forepart of the head of a dogfish embryo at a stage intermediate between o and r, showing the forebrain, the olfactory pits, and on the left side the olfactory nerve and olfactory lobe. X 17 diam.
Fig. 22. — Transverse section through the forepart of the head of a dogfish embryo of stage Q, showing the cerebral hemispheres, olfactory lobes, olfactory nerves, and olfactory pits with their Schneiderian folds, x 9 diam.
Figs. 23 — 26. — Sections through a dogfish embryo of stage n, showing the resemblances between the gills and the Schneiderian folds. Chromic and osmic acids.
Fig. 23. — Longitudinal and horizontal section through the head, showing fore- and hinclbrain, third and trigeminal nerves, eyes and olfactory pits. X 20 diam.
Fig, 24. — Transverse section through forepart of body, showing spinal cord, with anterior and posterior roots of a spinal nerve, vagus nerve, cardiac and dorsal aorta, branchial arteries, pharynx and gills. X 20 diam.
Fig. 25. — The right olfactory organ fi-om the same section as fig. 23 more highly magnified, showing the characters of the Schneiderian folds. X 90 diam.
Fig. 26. — The left gill from fig. 2i more highly magnified, showing histological characters of the gill folds, x 90 diam.
Figs. 27 and 28. — Sections from a dogfish embryo about stage o, showing resemblance between gills and Schneiderian folds at a rather later period. X 90 diam.
Fig. 27. — Through Schneiderian folds.
Fig. 28.— Through gill.
Fig. 29. — Transverse section through the anterior extremity of the head of a trout embryo on the twenty-seventh day after fertilisation of the ova, showing forebrain, olfactory pits, and on the left side the olfactory nerve. Picric acid. X 56 diam.
Fig. 30. — Longitudinal and vertical section through the head of a trout embryo on the thirtieth day, showing mid- and hindbrains, roots of visceral arches and clefts, and the olfactory pit. Picric acid, x 56 diam.
Fig. 31. — Transverse section through the anterior extremity of the head of a salmon embryo about the time of hatching, showing olfactory pits, trabecular plate, and anterior prolongations of the buccal cavity. Chromic and osmic acids. X 27 diam.
Fiig. 32. — Transverse section through the forepart of the head of a salmon embryo about a week after hatching, showing forebrain, eyes, olfactory pits, and paired-anterior diverticula of the buccal cavity. Picric acid. X 27 diam.
Fig. 33. — Transverse section through the forepart of the head of a salmon embryo two days after hatching, showing forebrain, and on the left side tlie eye, on the right the olfactory nerve and organ. Chromic and osmic acids, X 35 diam.
Figs, 34 and 35, — Longitudinal and vertical sections through the head of a salmon embryo of the same age as that in the preceding figure. Chromic and osmic acids. X 27 diam.
Fig. 34. — Section taken close to median line, showing root of olfactory nerve, optic nerves, infundibulum, and anterior prolongation of buccal cavity.
Fig. 35. — More superficial section, showing olfactory pit, eye, pharynx with its branchial arches, and the anterior prolongation of the buccal cavity.
Fig. 36. — Longitudinal and vertical section through the forepart of the cerebral hemisphere, the olfactory lobe and olfactory nerve of a twelve-day chick embryo. Picric acid, x 17 diam.
Figs. 37, 39 — Transverse sections through same region as preceding figure in a twelve-day chick embryo. Picric acid, x 17 diam.
Fig, 37. — Through the olfactory nerve.
Fig, 38, — Through the olfactory lobe.
Fig. 39. — Through the anterior part of the cerebral hemisphere.
Plate 13

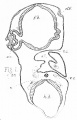
Fig 1.
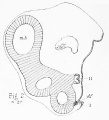
Fig 2.


Plate 14

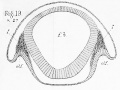
Fig 19.
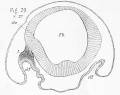
Fig 20.
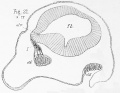
Fig 21.

Fig 22.
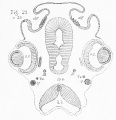
Fig 23.

References
- ↑ 1.0 1.1 I take this opportunity to make a slight alteration in the nomenclature adopted in my former paper. I have there suggested the term neural ridge for the longitudinal ridge of cells which grows out from the reentering angle between the external epiblast and the neural canal, and from which the nerves, whether cranial or spinal, arise. Since this ridge appears before closure of the neural canal is effected, there are manifestly two neural ridges, one on either side ; but I have also applied the same term, neural ridge, to the single outgrowth formed by the fusion of the neural ridges of the two sides after complete closure of the neural canal is effected, and after the external epiblast has become completely separated from the neural canal. I propose in future to speak of this single median outgrowth as the neural crest, limiting the term neural ridge to the former acceptation. Thus, while there are two neural ridges, there is only one neural crest, a distinction that will be at once evident on reference to my former figures.
- ↑ Quarterly Journal of Microscopical Science, January, I878, pp. 13— 16.
- ↑ 'Elasraobrauch Fishes,' p. 178.
- ↑ Op. cit., p. 179.
- ↑ ' Quarterly Journal of Microscopical Science January 1S78, pp. 17 — 23. To avoid repetition, I beg to refer the reader to the detailed account of the early stages contained in this paper.
- ↑ ' Journal of Anatomy and Physiology,' vol. xi, plate xxi, fig. 13.
- ↑ ' Untersuchungen iiber die Entwickelung der Wiibelthere.' Berlin, 1855, p. 33. 2 Log. cit., p. 74, note 55.
- ↑ Kolliker, ' Entwickelungsgeschiclite des Menschen uud der hoberen Tbiere ;' Zweite Auflage. Zweite Hiilfte, 1879, p. 609. = ' Elements of Embryology.' Part 1 , 1874, p. 117.
- ↑ For the opportunity of investigating the development of the Axololl, I am again indebted to Mr. Edon, of the South Kensington Museum.
- ↑ "On the Development of the Skull and its nerves in the Green Turtle {Chelone Midas):' 'Proc. Royal Society,' 1879.
- ↑ Vide Max Schultze. "UiitersuchunEjen iiber den Bau der Nasenschleimhaut bei dem Menscben und Wirbeltbiere." Halle, 1862, pp. 18, 19; and Stannius, ' Haudbucb der Auatomie der Wirbeltbiere,' 2 Aufiage, 1854, p. 165, seq.
- ↑ Owen, ' Anatomy of Vertebrates,' vol i, p. 283, 1866.
- ↑ Fide 'Quain's Anatomy,' 8th edition, vol. i, p. 526.
- ↑ Balfour, op. eit., p. 178.
- ↑ Ursprung der Wirbelthiere.' Leipzig, 1875, p. 23.
- ↑ Cf. Dolirn, op. cit., * Princip des Eiinctionswechsels.
- ↑ 'Journal of Anatomy and Physiology.' vol. xl, p. 510.
- ↑ For a full discussion of the reasons which have led me to consider the third nerve as a segmental nerve, and the fourth and sixth nerves as having DO claim to segmental value, vide 'Quart. Jouru. Micros. Sol.,' Jan., 1878, pp. 23 — 28, and pp. 32 — 33. the observations there recorded for the chick I have since verified in the dogfish.
- ↑ Op. cit., p. 216.
- ↑ Op. cit., pp. 10, 11.
- ↑ Cf. Parker, " On the Development of the Skull and its Nerves in the Green Turtle," ' Proc. Royal Soc.,' 1879.
- ↑ 'Trans. Zool. Soc.,' 1876, "On the Structure and Development of the Skull in Sharks and Skates," pp. 212 and 224.
- ↑ " On the Development of the Skull and its Nerves in the Green Turtle." 'Proc. Royal Soc' 1879. This vras read before the Royal Society on the same evening as the abstract of the present paper.
- ↑ ' Quart. Journ. Micros. Sci.,' January, 1878, pp. 23 — 27.
- ↑ Balfour, op. cit., p. 213, aud self, loc. cit., pp. 34 — 36.
- ↑ Op. cit., p. I89.
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - 1879 The Morphology of the Vertebrate Olfactory Organ. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_1879_The_Morphology_of_the_Vertebrate_Olfactory_Organ
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G