Paper - On the development of the membranous labyrinth and the acoustic and facial nerves in the human embryo
| Embryology - 28 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Streeter GL. On the development of the membranous labyrinth and the acoustic and facial nerves in the human embryo. (1906) Amer. J Anat. 6:139-165.

| Historic Disclaimer - information about historic embryology pages |
|---|
 |
On the Development of the Membranous Labyrinth and the Acoustic and Facial Nerves in the Human Embryo

{kind=link}
By
Associate, Wistar Institute of Anatomy.
From the Anatomical Laboratory of Johns Hopkins University.
With 2 Plates snd 8 Text figures.
- Preliminary reports concerning this investigation were read, and the models demonstrated, at the International Congress of Anatomists at Geneva, August, 1905, and at the meeting of the American Association of Anatomists at Ann Arbor, December, 1905.
Introduction
In the following paper some observations are reported concerning the embryonic morphology of the acoustic nerve and the development of the ganglion mass incorporated in its trunk. The differentiation of this latter mass, the ganglion acusticum, and its subdivision into the ganglion vestibulare and the ganglion spirale present several features of interest ; and deserving of especial attention is the additional light which the study of this process throws upon the question of nerve supply of the saccule, and the ampulla of the posterior semi-circular canal. It is found, namely, that these two portions of the membranous labyrinth are not supplied by the cochlear nerve, as described in English and German text books, but are supplied by the vestibular nerve, as has been maintained by some of the French writers. This brings all of the ampulls together with the utricle and saccule under control of the same nerve, and leaves the cochlear nerve as a specialized and distinct nerve for itself, supplying only the cochlear duct. This arrangement is one which should be gratifying to the physiologist, for it draws a definite line between that portion of the nerve complex which controls the analysis of sound and that which controls equilibrium.
This investigation was originally concerned only with the acoustic complex, later it was extended to the ear vesicle, and it IWS found possible to add several new features concerning the development of this structure and the formation of the membranous labyrinth to that which was already known from the work of His, Jr., 89,who, as far as could be learned, is the only investigator that has made a direct attack on this region in the human embryo since the introduction of wax plate reconstruction methods. It is, of course, to be remembered that in his work attention was mainly directed toward the nerve and ganglion masses, while the finer structure of the ear vesicle was not considered in detail.
The contributions here reported include both additional early stages in the development of the ear vesicle and further details in the formation of the individual parts of the labyrinth. Also some apparently fundamental errors in the work of the above investigator have been here corrected. One of these regards the saccule, which as represented by His, Jr., develops as a compartment pocketing out from the upper end of the cochlea, but which in our specimens develops as a compartment or subdivision of the utricle. Instead of the saccule developing from the cochlea, the cochlea develops from the saccule, though this occurs at a considerable time before the separation betmen utricle and saccule is complete.
The facial nerve, and especially its sensory division or pars intermedius, bears such a close relation to the auditory apparatus that it was found coiivenient to include it in some of the reconstructions. It was possible to identify conditions in the embryo confirmatory of what is now the generally accepted opinion as regards the adult, i. e., that the nervus intermedius is the dorsal and sensory root of the seventh, its fibers arising in the geniculate ganglion and continued peripherally in the chorda tympani and great superficial petrosal.
Material and Method
This work was made possible through the kindness of Professor lfall, who gave the writer, for the purpose of this investigation, free access to his large collection of human embryos. In the following list are tabulated the embryos which were selected for reconstruction:
| List of Embryos Reconstructed | |||||
|---|---|---|---|---|---|
| Number of embryo | Length in mm |
Probable age in days | Section | ||
| N. B. | V. B. | Thickness (micron) |
Direction | ||
| 148 | 4.3 | 3 | 20 | 10 | Coronal |
| B 17 | 6.5 | 6.6 | 26 | 15 | Sagittal |
| 2 | 7 | 6 | 26 | 15 | Coron-trans. |
| 163 | 9 | 9 | 30 | 20 | Transverse |
| 109 | 10.5 | 11 | 33 | 20 | Transverse |
| 175 | 13 | 13 | 36 | 20 | Transverse |
| 144 | 12 | 14 | 37 | 40 | Sagittal |
| 22 | 18 | 20 | 44 | 50 | Transverse |
| 229 | - | 21 | 44 | 50 | Sagittal |
| 86 | 20 | 30 | 54 | 50 | Coronal |
One or more wax plate reconstructions were made of each embryo after the method of Born. In most cases the models included the membranous labyrinth with the acoustic and facial nerves, and a portion of the central nervous system. Of these models seven were selected for illustration and are shown in Plates I and II. The form of the models has been controlled in all cases by dissections of pig embryoR of corresponding stages of development, prepared in the manner described in a previous paper (Streeter, 04, p. 87). Such comparison was of particular assistance in the study of the nerves and ganglion masses. The value of these dissections was greatly increased by previously staining the embryos, in toto, with alum cochineal (powdered cochineal 6 gm., ammonia alum 6 gm., and distilled water 200 cc.), which produces a brilliant differentiation of the tissues. In the same way that a microscopical section is improved by staining so is a stained microscopical dissection that much better than an unstained one. In studying these a strong, direct illumination of the specimen is necessary.
Whenever the size of an embryo is expressed by a single dimension it refers to its greatest length, and the age is that as determined by Mall's rule, i.e., age in days equals the square root of the greatest length times one hundred.
The drawings for Plates I and II were prepared under the guidance and assistance of Mr. Max Brodel, for which the author derives pleasure in taking advantage of this opportunity to acknowledge his appreciation.
Membranous Labyrinth
The auditory organ is generally described as developing phylogenetically from the lateral line organs of the marine vertebrate, which sink beneath the surface of the body and develop a cartilagenous or bony capsule, and become incorporated in the underlying head skeleton, the communication with the surface being maintained by a specially devised accessory apparatus.
In the embryo the first sign of the auditory organ, according to Krause, 03,and Poli, 97, consists of a thickening of the ectoderm, the auditory plate, which is seen lateral to the still open medullary groove in the region of the future third brain vesicle. In vertebrates having two layers of ectoderm the thickening involves the inner layer, the outer not being affected. Owing to the fact that the growth of cells shows greater activity in the deeper strata of the auditory plate it soon becomes converted into a cup shape depression and is then called the auditory fossa or auditory cup. By the folding in and closure of its edges the auditory cup is in turn converted into the auditory vesicle, which, however, remains attached to the surface for a longer or shorter period by means of an epithelial stalk or canal being finally separated from the surface, in mammals much earlier than in lower vertebrates.
It is at this point, just after the ear vesicle has been pinched off from the ectoderm, that my own observations begin. This stage corresponds to the “ primitive ear vesicle ” of Krause, 03, and will be described under that heading here.
The primitive ear vesicle
The reconstruction of the ear vesicle of an embryo 4.3 mm. long, No. 148, shown in Fig. a, Plate I, representa our youngest stage. This is considerably younger than the youngest human embryo described by His, Jr., 89. It is about the same age as shown in Krause’s, 03, Fig. 82, a model from a rabbit embryo, and is younger than the first stage of the series of models of the ear vesicle of the bat recently published by Denis, 02.
The ear vesicle consists at this time of a slightly elongated, oval sac, having the following diameters : dorso-ventral, .39 mm. ; caudo-cephalic, .26 mm., and transverse, .28 mm. It lies closely against the neural tube, and is connected with it by the acoustic ganglion, similarly as is shown by Mall, 88, in the dog, figured in his Fig. 4, Plate XX, and is surrounded on all sides by a thin layer of mesodermal tissue.
On the dorso-lateral surface, above that portion which is to become vestibular pouch and near where the endolymphatic appendage is to be separated off from the rest of the vesicle, there is a shallow groove. This groove, as seen in the sections, is cut transversely and consists of a seam, or the meeting point of the former edges of the auditory cup whose approximation completes the closure of the vesicle. This closure seam shows various degrees as regards the completeness of fusion, manifested by a difference in the thickness of the opposite edges, and the degree of obliteration of the line of juncture. The remainder of the vesicle wall is everywhere quite uniform in appearance, consisting of 2-3 layers of slightly elongated epithelial cells, without any apparent differentiation to indicate points of future nerve endings.

Fig.1. Profile reconstruction showing the membranous labyrinth and its relative size and relations to the brain and the flfth and seventh cranial nerves. Human embryo 14 mm. long, Mall Collection No. 144, magnified about 8 diams.
So epidermal stalk could be detected connecting the vesicle with the surface, or persisting beneath the surface epithelium, as observed in the rabbit by Krause, 03, p. 88. Evidently in the human embryo such a stalk must be either very temporary or else never present, as here we have to do with a vesicle whose closure and detachment from the surface must be regarded as only just completed.
The development of the endolymphatic appendage and its relation to the epithelial stalk formed during the detachment of the ear vesicle from the epidermis has excited a considerable controversy out of which certain facts have become definitely established. In the first place it is evident (Eeibel, 99; Blexander, 01;Krause, 01,and 03) that in the chick the appendage is formed out of the original union region between epidermis and labyrinth anlage, and corresponds to the closing place of the ear vesicle, and is its last point of attachment to the surface. On the other hand it is also established (Corning, 99; Peter, 00, and Krause, 01) that in reptiles and amphibia the tip of the appendage does not coincide with the point of detachment of the ear vesicle, but is situated somewhat more dorsal and proceeds in a course of independent development before the detachment of the vesicle is complete.
In the human embryo the endolymphatic appendage approaches in its development more nearly the type seen in amphibia than that in the chick. It is not developed until the epidermal stalk, if there ever is any such in man, has disappeared. Its anlage is formed by that portion of the vesicle wall just dorsal to the seam of closure, forming a rounded point on the dorsal edge of the vesicle, thus its tip cannot coincide with the point of detachment. Its situation is indicated by the external form before there is any apparent differentiation of the wall and can be seen in Fig. a, Plate I. By comparison of Figs. a-f, Plate I, it will be noticed how, by a process of extension, this diverticulum becomes converted into the endolymphatic appendage. In the second stage, Figs. b and c, the external form of the appendage is more distinctly outlined, as a short diverticulum opening widely into the rest of the vesicle. In the next older embryo, Figs. d, e, and f, by extension of the tip and constriction of its base the appendage begins to assume a typical form. The last step in its differentiation consists in the widening of the distal end into a flattened pouch or sac, in contrast to the remainder, which persists as a narrow duct connecting it with the vestibule, indicated in Figs. 1, m, n, Plate I, and well marked in Figs. a, b, c, Plate II. These are the two divisions of the appendage that are distinguished by the names endolymphatic sac, and endolymphatic duct.
During this process of expansion the wall of the appendage which originally, like the rest of the primitive vesicle, consists of an epithelium of 2-3 layers, is thinned out to a single layer. The thinning out commences in embryos of about 6 mm. It is at first limited to the lateral surface and the extreme tip of the appendage, while the median wall continues to be 2-3 cells thick. It is not until the embryo is about 18 mm. long that the whole appendage wall is thinned out to a single layer. It seems probable that the thick median wall in embryos of 6-18 mm. constitutes a germinating bed which furnishes ihe cells needed for the rapidly expanding appendage. It is only these cells that continue to multiply, and they can be imagined as moving around toward the lateral surface in a single layer in the order in which they are derived from their focus of growth.

Fig. 2. Profile reconstruction showing the membranous labyrinth and the flfth and seventh cranial nerves. The sensory part of the seventh is indicated by solid black. The great superflcial petrosal nerve extends from the geniculate to the spheno-palatine ganglion. Human embryo 30 mm. long, Mall Collection No. 86, magnified about 7 diams.
The diverticulum stage
Between the primitive vesicle just described and the labyrinth possessing cochlea, semi-circular canals, and accessory recesses, there is a stage through which the ear vesicle passes which can be characterized as the diverticulum or pouch stage. It is represented by the embryos 6.6 mm. and 9 mm. long, shown in Figs. b-f, Plate I. In these two embryos the vesicle may be said to consist of two pouches, a large, bulging triangular one above, with the endolymphatic appendage, the vestibular pouch, and opening into it from below the more slender and flattened cochlear pouch. Where these two pouches meet, there is a portion of the vesicle which is destined to form the utricle and saccule. It can be distinctly seen in Fig. f, Plate I. This was observed in the bat by Denis, 02, who called that part of it which projects toward the median surface the diverlicule utriculo-sacmlaire. The space concerned, however, involves also a part of the anterior and lateral walls of the vesicle and perhaps it would be advantageous to include this whole region under the concise and descriptive name atrium. This atrium is properly a subdivision of the vestibular pouch. It is in fact all that part of it which is left after the separation off of the semicircular canals and their ampulla. It is not to be confused with the cochlear pouch, which is phylogenetically a secondary diverticulum, which buds out from the atrial portion of the vestibular pouch. The embryonic relation is indicated in the following table:
| primary vesicle | endolymphatic appendage
|
atrium |
endolymphatic duct
endolymphatic sac
semicircular canals ampullae
saccule cochlea |
- primary vesicle
- endolymphatic appendage
- endolymphatic duct
- endolymphatic sac
- vestibular pouch
- canal pockets
- semicircular canals
- ampullae
- atrium
- utricle
- saccule
- canal pockets
- cochlear pouch
- cochlea
- endolymphatic appendage
If the words pars superior and pars inferior were substituted for the two pouches this conception would then be at variance with Krause, 03, only as regards the saccule which he describes as belonging to the pars inferior. This will be again referred to later.
The surface markings of the vesicle during this stage assume a significant character. In the first place the vestibular pouch at once takes on a triangular shape with the apex toward the appendage. The three borders of this triangle form the adages of the semicircular canals (see Fig. d, Plate I), which bear the same inter-relation as the canals in later stages. A second feature which is apparently constant and important is the sharp, vertical groove, which cuts in between the anlage of the posterior canal and the posterior end of the lateral canal. This we may call the lateral groove. It was not represented by His, Jr., 89, but can be seen in the model from the 8 mm. rabbit of Krause, 90, p. 296, and still better in models 3, 4, and 5 of Denis, 02, which were taken from the bat. The latter author mentions it in his text.
Ventral to the anlage of the lateral canal, on the lateral surface of the vesicle there is a rather large depression or fossa, which becomes more marked in proportion to the increasing projection of the lateral canal, which overhangs it like a shelf. This fossa forms the lateral wall of the atrium from which the utricle and saccule are to develop. The cochlear portion of the vesicle is limited. to its ventral tip and extends up along the rounded posterior border nearly to the prominent anlage of the posterior canal. There intervenes between them that portion of the wall thab is to become the posterior ampulla. The tip of the cochlea begins to bend forward practically as soon as the cochlear pouch can be distinguished as such.
The changes in the structure of the wall of the ear vesicle which accompany the pouch formation are limited to the thinning out of certain areas on the dorso-lateral surface of the vestibular pouch, and the lateral surface of the appendage as has already been referred to. The remainder of the vesicle wall is of the primitive type ; there were no areas that could be recognized as nerve endings. In embryo No. 163, 9 mm. long, however, protoplasmic nerve processes extend from the ganglion and lose themselves in the vesicle epithelium. The branch destined to become the posterior ampulla nerve could be seen with great distinctness; but where it ended there was no reaction to be seen on the part of the epithelium.
The period of semicircular canal formation is shown in Figs. g-k, Plate I. The process consists in the expansion of the edges of the vestibular pouch, i. e., the canal adages, and the coincident absorption of the intermediate vestibular walls, as was essentially described by Bottcher in his monumental work of 1869, and to some extent by other observers even previous to that. Since then further details have been worked out by various investigators, notably by Krause, 90, and 03,who approached the problem along the whole line of vertebrates. He demonstrated that the canals are formed one after the other in definite sequence, the superior first then the posterior and lastly the lateral. Our information concerning the human semicircular canals is based principally on the work of His, Jr., 89.
An interesting interval, which was left open by His, Jr., between his stages shown in his Figs. 6 and 7, Plate I, is filled in by my models, made from 11 and 13 mm. embryos (Figs. g-k, Plate 4). What is to be particularly noted is the change occurring in the structure of the vestibular wall which can be seen from a surface examination of the model. Those areas which are to persist stand out prominently and present a fairly definite outline of the future labyrinth, while the intermediate areas, which are destined to be absorbed, collapse before the advancing mesoderm; this is well shown in Figs. j and k. It might be thought that the absorption of epithelium in Fig. j had been completed as far as the superior canal is concerned, and that the remaining epithelium would go to make the canal wall, necessarily stretching out to obtain the diameter represented by the same canal in Fig. m. This, however, is not the case; it is only the thickened edge of the pockets of the vestibular pouch that becomes canal wall. In Fig. j there still remains a large area of epithelium that is to be absorbed before the inner rim of the superior canal is reached.
The histogenesis of the semicircular canal is shown in the accompanying Text Fig. 3, in which A, B, C, and D represent transverse sections of the superior canal in four stages of differentiation, taken at corresponding points and magnified the same number of diameters. The striking feature of the process is the persistence in the canal anlage of the primitive epithelium of 2-3 layers until after the canal is closed off, evidently being a factor in ita rapid growth. Section A is taken from the ear vesicle of a 9 mm. embryo, the same as shown in Figs. d, e, f, Plate I. Aa shows the entire section of which A is a portion, and Ab indicates the direction of the section as regards the ear vesicle. Section B is from a 11 mm. embryo. The entire section is represented by Ba, whose position as regards the ear vesicle is shown on Bb, which is from the same model shown in Figs. h, i, j, Plate I. Sections through the vestibular region at this stage are very interesting, as they show by the thickness of the wall which are the persistent areas; section Ba is made in such a way a8 to include the anlages of two canals, the lateral wall of the crus commune and a part of the utricle and the ductus endolymphaticus, all of which stand out prominently. The intervening vestibular epithelium, which is doomed to absorption, consists of a single layer of cuboidal cells, as shown in B, in contrast to the thick outer edge which is to become canal.

Fig. 3. Development of a semicircular canal. A, B, C, and D represent transverse sections of the superior canal (x240), taken at corresponding points from embryos No. 163, 9 mm.; No.109, 11 mm.; No. 229, 21 mm., and No. 86, 30 mm. Aa, Ab, Ba,and Bb are explanatory drawings of lower magnification to show the ear vesicle and the situation and shape of sections from which A and B are taken.
This process of absorption may be described histologically as a conversion of the definite epithelial membrane into a line of cells which seem to fuse with and cannot easily be distinguished from the adjacent mesodermal cells, the line finally becoming broken and irregular. The transition from one step in this procedure to the next is quite abrupt; thus in B the thin membrane is sharply cut off from the absorption focus. Several specimens were examined of about this age, and in one case, embryo No. lY5, 13 mm. long, it WM found that absorption of the epithelium was going on before the lateral and median walls of the vesicle had actually come together. So it is possible that during this process the vesicle cavity is in some cases left temporarily in open communication with the spaces of the adjacent mesoderm. The final curling in of the edges and closure of the canal tube repeats in a wag the procedure which we have already seen in case of the auditory cup duriing its conversion into the auditory vesicle. It is probably likewise mechanically brought about by the arrangement of the epithelial cells. Section C shows the canal after the formation of the closure seam, the so-called raphe of Hasse. The thickness of the epithelium of the outer edge and presence of division figures indicate that the activity of growth still continues. Section D shows a canal in an embryo 30 mm. long, the same stage as that shown in Pigs. a, b, c, Plate II. Here the epithelium is reduced to a single layer and division figures have disappeared. It can be seen, however, that traces still exist of the thickened outer edge and the raphe of Hasse. This stage differs from the adult canal practically only in its diameter, which there is 3-4 times greater. Doubtless this growth is in large part accomplished simply by the flattening out and expansion of the individual cells.
The formation of the ampullae can be seen by comparing the figures on Plates I and II. It will be noticed that their development proceeds simultaneously with that of the canals. In their histogenesis they resemble the canals, in having a thin single layer of epithelium on the inner rim and the thick 2-3 layered epithelium on the outer surface. It is out of the latter primitive epithelium that the maculze are developed, and they make their appearance before ampullze and canals are completely separated from the remainder of the vestibular sac; they can be seen in the 11 mm stage, but a high degree of differentiation is not found until we come to embryos 20 mm long. It will be remembered that His, Jr., 89, represents ampulla as forming on both ends of the superior and posterior canals. This was not confirmed in our models; the ends of these two canals where they unite to form the crus commune show no such enlargement. Each canal possesses but one ampulla.
The development of the utricle and snccule is dependent on the subdivision of the atrium into an upper and lower compartment. The atrium, ashas already been described is that ventral part of the vestibular pouch into which the endolymphatic appendage opens, and into which the cochlear pouch opens from below; in Fig. f, Plate I, it is marked utric-sacc., and in Fig. j the lateral surface of it is marked sacc., and in Fig. k a partial median view of it is marked utric. In Figs. j and k, though the canals and ampulle are already completing their Peparation from the vestibular pouch, the atrial region has not yet begun its subdivision. It, however, suggests by its outer form the future saccule and utricle. The actual subdivision begins in embryos between 18 and 20 mm. The initial ingrowth of the membranous partition can be seen in Figs. 1 and m, where it can be distinguished as a horizontal cleft which forms in front between the utricular and saccular parts of the atrium. Strictly speaking we cannot speak of a saccule and utricle until the intervening partition is complete. It is practically complete in Figs. a, b, c, Plate II; here it reaches back to the entrance of the ductus endolymphaticus. It later divides the orifice of that structure, thus affording it separate openings into the utricle and saccule, the two openings constituting the so-called ductus utriculo-saccularis.
In the meantime the utncle itself has developed a definite shape. As can be seen in the Figs. a, 6, and c, a transverse constriction divides it into an anterior or cephalic part and a posterior or caudal part. The anterior part constitutes the general utricular cavity, in the floor of which the nerve ends. In front, just ventro-median to the ampulla of the superior canal, a distinct diverticulum extends forward from it which is called the recessus utricularis. The posterior part consists of a central einus utriculi communis, into which opens from above the crus commune, laterally the sinus utriculi lateralis of the lateral canal, from below the sinus utriculi inferioris of the posterior canal, and on the median side the ductus endolymphaticus.
If one compares Figs. a, b, c, Plate II, with pictures of adult preparations such as found in the beautiful atlas of Schonemann, 04, it is apparent that the labyrinth of the 30 mm. embryo has practically completed its gross development. In its further expansion all parts of it become relatively more slender and the saccule draws away from the utricle and becomes flattened as well as biconcaved or saucer-shaped.
The cochlea as compared with the derivatives of the vestibular part of the ear vesicle is less complicated in its development, presenting only the peculiarity of spiral growth. The cochlea has already been referred to as the pouch which forms the ventral tip and part of the posterior border of the vesicle, as seen in Figs. b-f. In Figs. g, k, it is partly demarcated from the saccular region by a broad fossa. At 20 mm., Fig. I, a sharp constriction separates it from the saccule, and this becomes in the 30 mm. embryo the ductus reuniens, and in the meantime the cochlea has become a spiral of two turns.
As regards the relation of cochlea to saccule we differ from the description given by His, Jr., 89, who represents the saccule as budding off from the upper end of the cochlea, which is just the reverse of our own interpretation and what might be expected on the ground of the comparative anatomy of these structures. We know that in certain fishes the ear vesicle consists of a simple utricle into which the semicircular canals empty. In certain other fishes pockets bud out from the utricle analogous to the saccule. When we come to animals that leave the water, the amphibians, there develops from the saccule a secondary pocket, which in birds and reptiles takes on the characteristics which identify it with the mammalian cochlea. That is to say, first utricle, then utricle and saccule, and finally utricle, saccule, and cochlea. The phylogenetic development presents here, in discrete steps, the process which we find in the human embryo, but in the latter case it is a matter of simultaneous growth of all three structures.
A resume of the development of the labyrinth is presented in the form of a diagram in the adjacent Fig. 4, which illustrates the successive steps by which the simple ear vesicle enlarges and becomes differentiated into the group of connected individual compartments which characterize the adult ear.

Fig. 4. Diagram representing the growth and stages of differentiation of the human membranous labyrinth. At 3.5 weeks (6-7 mm) the ear vesicle consists of two simple pouches, into the upper of which opens the endolymphatic appendage. At 4 weeks (9 mm) there is at the base of the vestibular pouch an atrium, the space destined to form the utricle and saccule. At 5 weeks (12 mm) this space is circumscribed from the cochlear pouch below by a constriction corresponding to the ductus reuniens, and above from the rest of the vestibular pouch by the formation of the semicircular canals. At 6 weeks (20 mm.) an ingrowth of the wall of the atrium divides it into an upper part (utricle) and lower part (saccule). At 10 weeks (30 mm) this partition between the utricle and saccule is complete and extends inward in such a way as to split the orifice of the endolymphatic duct.
The ear vesicle very early (6-7 mm. long, 3.5 weeks) assumes the form of two communicating pouches, the vestibular pouch, with its endolymphatic appendage, and the cochlear pouch. The first gives origin to the semicircular canals, ampullze, utricle, and saccule. The semicircular canals, in consequence sf the approximation and absorption of the intervening wall of the vesicle, make their appearance between the fourth and fifth weeks, 9-14 mm. (only one is shown in the diagrams). That portion of the vestibular pouch that is not involved in the formation of the canals and their ampullae may be called atrium, to indicate that it forms at this time a cominon meeting place into which open the different compartments, including the endolymphatic appendage. At six weeks, 20 mm., the atrium becomes separated into an upper and lower division by an ingrowth of its wall, thus forming the utricle and saccule. This partition continues inward in such a way as to split the orifice of the ductus endolymphaticus, the divided ends of which form the ductus utriculo-sacculus. The cochlear pouch opens directly into the atrium, and as the development proceeds it can be seen that it is into that part of the atrium which is destined to form the saccule. At the fifth week, 14 mm., a beginning constriction appears between the cochlea and the saccular region. This constriction corresponds to the ductus reuniens and gradually narrows down until in the adult in many cases the communication between cochlea and saccule is obliterated. It is very apparent that the saccule is not developed from the cochlea, but the cochlea may be said in a certain sense to develop from the saccule.
N. Vestibularis and N. Cochlearis
The earlier anatomists described the auditory nerve as being made up of two main divisions. One of these, according to their plan, supplied the utricle, saccule, and the ampullre of the three semicircular canals, while the other division they considered to belong exclusively to the cochlea. This description prevailed up to the time the exhaustive monograph was published by Retzius, 84, upon the comparative anatomy of the membranous labyrinth and its nerves. This investigator, by means of careful dissection of a great variety of vertebrate material, was able to present a much more minute description of the n. acusticus than had previously existed. In mammals, according to his view, the anterior division or ramus vestibularis supplied the utricle, and the superior and lateral ampullze, while the posterior division or ramus cochlearis supplied the saccule, the posterior ampulla and the cochlea. This classification was substantiated not long after by His, Jr., 89, in his paper on the development of the human acoustic complex, in which he also represented the cochlear division as supplying not alone the ductus cochlearis but also the saccule and ampulla of the posterior canal. From that time until now the classification made by Retzius has been the one generally adopted by both English and German text books. Certain French writers (Cannieu, 94, 04, and Cuneo, 99), however, have come back to the original conception of the cochlear nerve and its individuality. They point out that Retzius fuses in his ramus cochlearis the inferior branch of the ramus vestibularis and the cochlear nerve proper. They admit that these two lie side by side and are closely united, but further than that deny any anatomical or physiological relation. A similar conclusion has also been reached by Alexander, 99, who studied serial sections of the acoustic ganglion mass taken from various adult mammals.
My own observations concerning the development of these structures in the human embryo are quite contrary to those of His, Jr., and as will be immediately seen, they seem to indicate that the cochlear division of this complex has nothing to do with the nerves to the saccule and posterior ampulla, but possesses its own specialized characteristics which distinguish it from all the rest of the acoustic mass. Embryologically, therefore, it seems well to follow Cannieu's, 94, lead and adopt the following classification :
| N. Octavus (N. Acusticus) | ||
|---|---|---|
| n. vestibularis | pars superior
|
r. ampul. sup. r. ampul. ext. r. recess. utric. r. sacc. |
| n. cochlearis | ramuli spirali | |
The form and branches of the acoustic mass in its different stages and its relation to the labyrinth is shown in the figures on Plate I and II. Two colors are added so that the cochlear division can be distinguished from the vestibular; the former is colored yellow and the latter light red. The same ganglion mass is shown more diagrammatically in the accompanying Fig. 5, showing its appearance in embryos 4, 7, 9, 20, and 30 mm long. The vestibular part is indicated by fine dots and the cochlear by coarse dots. The drawings on the left present a median view and those on the right a lateral view.

Fig. 5. Stages in the differentiation of the acoustic nerve complex. Vestibular ganglion shown by fine dots, and spiral ganglion by large dots.
In the youngest stage, embryos of about 4 mm, Mall collection, No. 148, the outlines of the ganglion mass are indefinite, particularly the peripheral border. The central end is more distinct and the protoplasmic cell processes can be seen leading to the wall of the neural tube. This is somewhat younger than the earliest stage of His, Jr., 89. In the next stage, embryos of about 7 mm., the outlines of the ganglion can be clearly made out. A section through such a ganglion is shown in Fig. 6. It lies closely against the front edge of the vesicle, its lower end migrating around on the median side. In its outer form it consists of an upper and lower part, pars superior and pars inferior, each of which develops its own separate group of peripheral nerve branches; the central root of the ganglion connecting it with the brain consists of a single stem. Owing to the proximity of the ganglion mass to the ear vesicle the nerves uniting them are at this time very short. His, Jr., 89, p. 6, regards that portion of the ganglion which we have called the pars inferior as the ganglion cochleare. What I regard as the ganglion cochleare or ganglion spirale does not make its appearance until a trifle later, in embryos of about 9 mm. There can be seen then a group of ganglion cells massing themselves on the ventral border of the pars inferior, which corresponds completely to the future spiral ganglion and may be considered as its adage. This adage develops into a derivative which buds off from the pars inferior and then follows an individual course of growth independent of the latter, and this is analogous to the way in which we have already seen the membranous cochlea bud off from the saccule and develop independently.
That part of the pars inferior which does not participate in the formation of the spiral ganglion remains closely related to the pars superior, and aupplies the saccule and posterior ampulla. It is this that His, Jr., describes in a later stage as the Zwischenganglion, and whose centripetal fibers he joins to those of the main cochlear trunk.
In embryos of 20 mm. (compare Figs. I, m, n, Plate I) the pars superior has increased greatly in size, and its peripheral nerves, which before were massed together, have become separate and distinct branches. The pars inferior, from which the spiral ganglion is rapidly separating, consists of a connecting strand of ganglion cells giving off separated branches to the saccule and posterior ampulla. The fibers extending to the posterior ampulla are at first (embryos of 11 mm.) loosely spread out and give the appearance of more than one nerve, but later, either by atrophy of some of them or by becoming bundled together more closely, they constitute a single compact nerve. It is possible that here we have to do with temporary fibers representing branches to the additional nerve endings which are €ound in this region in lower forms.
The cochlear nerve can be distinctly seen collecting its fibers from the spiral ganglion and extending up toward the brain. The exact manner in which this nerve reaches the neural tube proved difficult to determine. It apparently sprouts out from the spiral ganglion and travels up on the median surface of the vestibular ganglion until it reaches the brain. To be certain of this would require a greater number of stages between 8 and 10 mm. than were available. In the embryos studied the proximal end of the nerve could be made out almost as soon as the distal. So it is possible that the cochlear trunk consists originally of a column of ganglion cells connecting the anlage of the spiral ganglion with the brain, and the conversion of this column into fibroblasts produces the early fibers of the trunk; this would explain the abrupt appearance of the nerve trunk in all parts of its course at once.
Proceeding to embryos 30 mm. long, the same as seen in Figs. a, b, c, Plate I, we meet with conditions which are practically those found in the adult. There is the vestibular nerve, on whose trunk is situated its ganglion mass, consisting of an upper and lower division. The upper division is connected with the labyrinth by the branches supplying the anterior and lateral ampullce, and the utricle; the lower division gives off branches to the sacculus and posterior ampulla. In the adult the division between the pars superior and pars inferior is still more pronounced and the separation can be even seen in the trunk of the nerve. The ganglion mass is completely divided except for a bundle of anastomosing fibers, which according to Alexander, 99, may also be accompanied by a chain of ganglion cells-a persistence of the embryonic connection between the pars superior and pars inferior.
The cochlear nerve lies on the median surface of the two divisions of the vestibular ganglion, but is connected with them only by contiguity. This can be demonstrated by dissection methods in pig embryos; the cochlear trunk can be easily lifted off, leaving the vestibular ganglion mass and all of its branches in position, both of the pars inferior and pars superior. At the point where they enter the central nervous system the cochlear and vestibular trunks are in the 30 mm. embryo closely united; they run into each other slightly more than is shown in Fig. 5, and it is not easy to distinguish at what point the one stops and other begins. Their separation is brought about by the increase in size of the restiform body which develops in between them. During this process it could easily happen that some of the cochlear fibers should become grouped in with the vestibular, or that some of the vestibular should become grouped in with the cochlear; in both cases finally reaching their proper destination. If the individual fibers were traced in a large number of cases undoubtedly a considerable variation in this respect would be found. An important stride in this direction was made by Held, 93.
The twisted, rope-like character of the cochlear nerve, as indicated in Fig. 5, is easily identified in the 30 mm embryo. An effort was made in the pig to determine the number of turns and their relation to the number of turns of the spiral ganglion. There is apparently a certain amount of correspondence, but the fibers were too tightly adherent to admit of a satisfactory unrolling of the nerve.
In the summary of these nerves it should be emphasized that we have distinct points of difference between the vestibular and cochlear divisions of the acoustic complex, both as regards the ganglia and the nerve trunks themselves. The ganglia, belonging to the vestibular division, are spread along the trunk of the nerve; the ganglion of the cochlear division is situated at the extreme distal end of the nerve, and lies directly on the membranous labyrinth, being closely incorporated with it later in the cartilagenous capsule. The vestibular terminal branches develop as discrete and fairly long nerves; the cochlear terminal branches are short and freely anastomose. The main trunk of the cochlear division is characterized by the compactness of its fibers and their spiral arrangement; while in the vestibular division the fibers are less compactly bundled, showing a tendency to subdivision, and do not have the spiral character.
N. Facialis and Pars Intermedius
The facial nerve is so closely united in position with the acoustic ganglion that it was found advisable to include it in the reconstructions. For reason of simplicity it is not shown in Plates I and II, but its form and general position at two different stages is shown in the Text Figs. 1 and 2. It can be seen how it is divided into ventral and dorsal (motor and sensory) roots, and the situation of the geniculate ganglion on the latter.
Particular attention was given to the ganglion geniculatum and pars intermedius in order to bring their early morphology into accord with the conditions found in the adult. Though the “portio media inter communicantem faciei et nervum auditorium ’’ [1]was described by several writers over a century ago, and in the intervening time frequent reference and much speculation has been made concerning it; yet it has turned out that none of this was actually in advance of the original description until there appeared the paper of Sapolini, 83, who was the first to give a detailed report of its deep origin and terminal distribution. This investigator succeeded in dissecting out the pars intermedius in adult human material throughout its whole course. He describes it as arising in the floor of the fourth ventricle, from where it ascends as a nerve band which runs along the median edge of the acoustic area. At the level of the inferior cerebellar peduncles it makes its exit through the side of the pons, between the seventh and eighth cranial nerves, and then passes through the geniculate ganglion, and is continued into the chorda tympani, which in turn joins the lingual branch of the fifth nerve and forms a terminal plexus in the substance of the tongue. We are thus indebted to this writer for establishing the fact that the nerve of Wrisberg, or pars intermedius, and the chorda tympani, are two continuous parts of the same nerve, connecting the anterior part of the tongue with the floor of the fourth ventricle, an essential fact which has been very slow in getting into our text books. Further details regarding the central path of this nerve is given in a supplementary note in a paper of His, 90, in description of a 17 mm human embryo. He reports this nerve as extending from the geniculate ganglion as an independent bundle into the brain, where it can be seen running along the median edge of the acoustic area spinalwards until it reaches the entering fibers of the glossopharyngeus. It joins with these fibers and together with them takes part in the formation of the tractus solitarius. His felt justified in assuming that we have here to deal with a union within the neural tube of bhe taste fibers from the anterior and posterior portions of the tongue, one group coming through the chorda tympani and nerve of Wrisberg, and the other through the glossopharyngeus. This view has since then been vigorously supported by Dixon, 99, who pictures the facial nerve as a typical branchial nerve developed in connection with the ear cleft, having a motor part behind the cleft supplying the muscles developed from this arch, and a sensory part made up of the chorda tympani and great superficial petrosal which consist almost entirely of taste fibers. Reasoning from the development and comparative anatomy of the chorda tympani he points out the improbability of the presence of any taste fibers in the trigeminal nerve as had been maintained by many. That the fifth nerve pair contains few or no taste fibera has been further established by the careful observations of Gushing, 04, who, after the removal of the Gasserian ganglion, fund in all of his cases that the anterior part of the tongue retained in greater or less degree the sense of taste as well as common sensation.
If after the work of Sapolini there remained still doubt regarding the anatomical identity of the pars intermedius, geniculate ganglion, and chorda tympani, it was removed by the convincing dissections of Penso, 93. This worker investigated these structures in man and a considerable variety of other mammals, using principally dissection and teasing methods, though his observations were made also in part from serial sections. He showed conclusively that the geniculate ganglion is primarily connected with the motor trunk of the facial only by contiguity, its fundamental attachments being a central one, the pars intermedius, and two peripheral ones, the chorda tympani and the great superficial petrosal. He also describes a number of small anastoyoses existing between these branches and the surrounding structures, including the motor portion of the facial, the acoustic ganglion, the spheno-palatine ganglion, and the auricular branch of the vagus. He therefore represents the great superficial petrosal and chorda tympani as composite nerves, which are made up, in the first place of fibers from the geniculate ganglion, and in addition to these the fibers from the above-mentioned anastomosing branches. During the past year a paper confirming Penso's work in its essential points has been published by Weigner, 05, who studied serial sections of rodent and human material, and found it possible to trace to their destination the fibers of the n. intermedius by their histological character; that is to say, from the frequency of the sheath nuclei, the small size of the fibers, and Che presence of scattered ganglion cells lying along the course of the fibers.
The common identity of the pars intermedius geniculate ganglion and chorda tympani, as described by Sapolini and Penso, in the adult is less easily seen in the early embryo owing to the incomplete differentiation of these structures. In fact, His, Jr., 89, states that up to the third month no nerve is to be seen arising between facial and acoustic, and he concludes that the intermedius must until that time run in the trunks of these two nerves. In our embryos, however, it was found earlier than that. The Fig. 6 represents the pars intermedius and geniculate ganglion in an embryo of 33 weeks, and they can also be distinguished in embryo No. 148, Mall collection, which is about 20 days old. The ganglion can be made out first, and shortly after that the path of loose fibers connecting it with the neural tube. As is seen in Fig. 6, the ganglion and its proximal root are at this early period distinctly separate from the acoustic mass, and it is only as the acoustic mass.increases in size that it could be said to fuse with the geniculate ganglion; the existence even here of a real fusion is doubted, for no appearance was observed in any of our embryos that could not be explained by mere contiguity. The separation existing between these ganglia, in the early stages, merits the attention of those who would believe that they have a common origin, as was thought by the younger His.

Fig. 6. Sagittal section of a 7 mm human embryo (B. 17), showing the relations of the facial-acoustic complex.
In Fig. 6 the ventral or motor division of the facial nerve can be seen cut obliquely at the ventral edge of the geniculate ganglion. In this embryo the dorsal root or pars intermedius is fully as large as the ventral or motor root; but the proportion is gradually reversed as one looks through older stages, due to the more rapid growth of the ventral root. The pars intermedius is in this sense a more prominent structure in fetal life than in the adult, indicating that phylogenetically it has played a more important role in lower forms than in man. With the development of the connective tissue the geniculate ganglion becomes inclosed in a sheath, and is walled off from the motor root, against which it continues to lie, as may be seen in Fig. 8. In this figure the ganglion is cut somewhat obliquely, and at the two ends can be seen the central and peripheral fibers of the ganglion. To determine the destination of the peripheral fibers dissections were made of embryonic pigs and it waa found easy todemonstrate in 3-20 cm. embryos that the distal fibers leave at one corner of the ganglion by the great superficial petrosal nerve and at the other by a bundle that runs along the motor division until it leaves it as the chorda tympani.
Fig. 7 represents such a dissection made in a 20 cm. pig, showing the nature of the anastomosis with the auricular branch of the vagus, and following essentially the arrangement described by Sapolini and Penso.

Fig. 7. Drawing made from a dissection of the facial nerve in a 20 cm pig embryo. At this stage the sensory and motor divisions can be easily separated.

Fig. 8. Sagittal section through the geniculate ganglion and facial nerve of a 30 mm human embryo, No. 75, Mall Collection, showing how they are separated by a connective tissue partition.
In these dissections of the seventh nerve of pig embryos a ganglion cell mass, connected with the pons ganglia, was seen extending caudalwards as a surface ridge which could be traced beginning at the fifth nerve and then passing in between the seventh and eighth nerves and finally ending on the dorso-lateral surface of the restiform body. This is apparently the same structure that has been found by Mr. C. R. Essick, of the Johns Hopkins Medical School, in human adult material, and a full description of which he has now ready for publication.
References
ALEXANDER, G., 1899. - Zur Anatomie des Ganglion vetttibulare der Siiugethiere. Sitz. d. Kais. Akad. d. WiSS., math.-naturw. Classe, Bd. 108, Abth. 111, 1899.
- 1901. - Zur Entwicklung des Ductus endolymphaticus. Arch. f. Ohren- heilkunde, Bd. 52.
BOTTCHER A., 1869. - Ueber Entwick. u. Bau des Gehorlabyrinthes. Verhandl. d. Kaiserl. Leop . Carol. deutschen Akad. der Naturforscher., Bd. 86.
CANSIEU, A., 1894 .- Recherches sur le nerf auditif. These doctorat en medicine. Bordeaux. (Quoted from Cannieu, 04.)
- 1904. - Oreille interne. Trait d’anatomie humaine. Poirier et Charpy, Paris, Tome V.
CORNING,H. K., 1899. - Ueber einige Entwicklungsvorgange am Kopfe der Anuren. Morph. Jahrb,, Bd. 27.
CUNÉO, B., 1899. - Nerfs craniens. Traite d’anatomie humaine. Poirier et Charpy, Paris.
CUSHING, H., 1904. - The sensory distribution of the fifth cranial nerve. Johns Hopkins Hospital Bulletin, Vol. XV.
DENIS, P., 1902. - Recherches sur le developpement de I’oreille interne chez les mammifsres. Arch. d. Biol., Vol. XVIII.
DIXON, A. F.. 1899. - The sensory distribution of the facial nerve in man. Jour. of Anat. and Physiol., Vol. XXXIII.
HELD, H., 1893. - Die centrale Gehorleitung. Arch. f. Anat. und Physiol., Anat. Abth.
HIS, W., 1890. - Entwickelung des menschlichen Rautenhirns. Abhandl. d. S. Kgl. Sachs. Gesell. d. Wiss., Bd. 17.
HIS, W., Jun., 1989. - Zur Entwickelungsgeschichte des Acustico-Facialgebietes beim Menschen. Arch. f. Anat. 11. Physiol. Anat. Abth. Suppl.
KEIBEL, F., 1899. - Ueber die Entwicklung des Labyrinthanhanges. Anat. Anz., Bd. 16.
KRAUSE, R., 1890. - Entwicklungsgeschichte der hautigen Bogengange. Arch. f. mikr. Anat., Bd. 36.
- 1901 - Die Entwickelung des Aqueductus vestibuli s. Ductus endolymphaticus. Anat. Anz., Bd. 19.
- 1903 - Entwickelungsgeschichte des Gehororgans. Handbuch der Entwickelungslehre, Lief. 4 and 5,0.Hertwig, Jena.
MALL, F. P., 1888. - The branchial clefts of the dog. Studies from Biolog. Lab. of Johns Hopkins Univ., Baltimore.
PENSO, R., 1893. - Ueber das Gang. geniculi u. die mit demselben zusammenhangenden Nerven. Anat. Anz. 8.
PETER, K., 1900. - Der Schluss des ohrgrubschens der Eidechse. Arch. f. Ohren- heilk., Bd. 51.
POLI, C., 1897. - Zur Entwickelung der Gehorblase bei den Wirbelthieren. Arch. f. mikr. Anat., Bd. 48.
RETZIUS, G., 1884. - Das Gehorcrgan der Wirbeltiere. Stockholm.
SAPOLINI, 1883. - Etudes anatomiques sur le nerf de Wrisberg et la corde du tympan on un treizi6me nerf cranien. Journ. de Med. ’de Bruxelles. VOl. LXXVII.
SCHÖNEMANN, A., 1904. - Die Topographie des Menschlichen Gehororganes. Wiesbaden.
Streeter GL. The development of the cranial and spinal nerves in the occipital region of the human embryo. (1905) Amer. J Anat. 4(1):83–116.
WEIGNER, K., 1905. - Ueber den Verlauf des Nervus intermedius. Anat. Hefte. 87.
Footnote
- ↑ “portio media inter communicantem faciei et nervum auditorium ’’ - intermediate portion communicating between the central portion of the facial nerve opening
Description of Plates I and II
The reproductions shown. on these plates represent different views, in most cases lateral, front, and median, of seven selected models showing the membranous labyrinth and acoustic complex reconstructed from the following human embryos: No. 148, 4.3 mm; No. 817, 6.6 mm; No. 163, 9 mm; No. 109, 11 mm; No. 175, 13 mm.; No. 22, 20 mm.; No. 86, 30 mm. The colors, yellow and red, are used to indicate respectively the cochlear and vestibular divisions, and in general nerve flbers can be distinguished from ganglion cell masses by their lighter tone. The pictures represent a magniflcation of 25 diams.
The following abbreviations are used:
- absorpt. focus - area of wall where absorption is complete.
- amp. - ampulla membranacea.
- crus - crus commune.
- c.sc.lat. - ductus semicircularis lateralis.
- c. sc. post. - ductus semicircularis posterior.
- c. sc. sup. - ductus semicircularis superior.
- coch.orcochlea - ductus cochlearis.
- duct. endolymph. - ductus endolymphaticus.
- d. reuniens - ductus reuniens Henseni.
- endol. or endolymph. - appendix endolymphaticus.
- rec. utr. - recessus utriculi.
- sacc. - sacculus.
- sac. endol. - saccus endolymphaticus.
- sinus utr. lat. - sinus utriculi lateralis.
- utric. - utriculus.
- vestib.p. - vestibular pouch.


All Figures
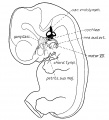
Fig 1 Membranous Labyrinth Human Embryo 14 mm
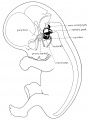
Fig 2 30mm Embryo

Fig 3 Semicircular canal

Fig 4 Membranous Labyrinth Growth
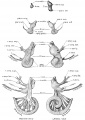
Fig 5 Acoustic nerve complex
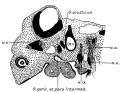
Fig 6 Facial-acoustic Complex Human Embryo 7 mm
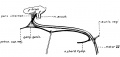
Fig 7 Facial Nerve Pig Embryo 20 cm
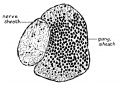
Fig 8 Geniculate Ganglion Human Embryo 30 mm
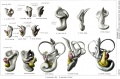
Plate 1. Membranous Labyrinth Human Embryo 4 to 20 mm
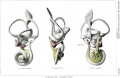
Plate 2. Membranous Labyrinth Human Embryo 30 mm
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Cite this page: Hill, M.A. (2024, April 28) Embryology Paper - On the development of the membranous labyrinth and the acoustic and facial nerves in the human embryo. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_On_the_development_of_the_membranous_labyrinth_and_the_acoustic_and_facial_nerves_in_the_human_embryo
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G