Introduction
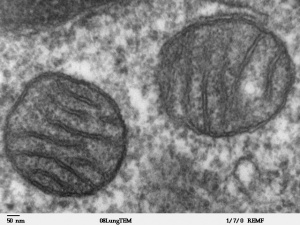
Electron micrograph of mitochondria.
Originally discovered in muscle by Kölliker in 1857, mitochondria are the "powerhouses" of the cell and the location where respiration occurs at the cellular level. These cytoplasmic organelles also contain their own DNA (mitochondrial DNA or mtDNA) that has been originally inherited only from the oocyte (maternal inheritance). The spermatozoa (paternal) mitochondria, required for energy for fertilization motility, can enter the oocyte but are generally destroyed during the first mitotic cell divisions. This pattern of inheritance has important implications for a variety of mitochondrial associated diseases, usually occurring in tissues requiring lots of energy (muscle, brain) and in the pathogenesis of common late-onset disorders. Recent experiments have employed swapping maternal mitochondrial DNA in mammalian oocytes, in order to overcome mitochondrial inherited diseases, raising both scientific and ethical discussion on mitochondrial donation.[1]
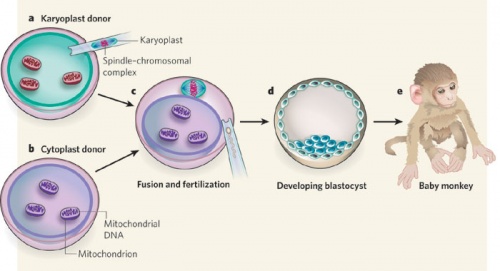
Swapping mitochondrial DNA mammalian oocytes[2]
Some Recent Findings

Spermatozoa Mitochondria in Early Mouse Embryos
[3]
- Segregation of mitochondrial DNA heteroplasmy through a developmental genetic bottleneck in human embryos[4] "Mitochondrial DNA (mtDNA) mutations cause inherited diseases and are implicated in the pathogenesis of common late-onset disorders, but how they arise is not clear. Here we show that mtDNA mutations are present in primordial germ cells (PGCs) within healthy female human embryos. Isolated PGCs have a profound reduction in mtDNA content, with discrete mitochondria containing ~5 mtDNA molecules. Single-cell deep mtDNA sequencing of in vivo human female PGCs showed rare variants reaching higher heteroplasmy levels in late PGCs, consistent with the observed genetic bottleneck. We also saw the signature of selection against non-synonymous protein-coding, tRNA gene and D-loop variants, concomitant with a progressive upregulation of genes involving mtDNA replication and transcription, and linked to a transition from glycolytic to oxidative metabolism. The associated metabolic shift would expose deleterious mutations to selection during early germ cell development, preventing the relentless accumulation of mtDNA mutations in the human population predicted by Muller's ratchet. Mutations escaping this mechanism will show shifts in heteroplasmy levels within one human generation, explaining the extreme phenotypic variation seen in human pedigrees with inherited mtDNA disorders."
- Obesity-exposed oocytes accumulate and transmit damaged mitochondria due to an inability to activate mitophagy[5] "Mitochondria are the most prominent organelle in the oocyte. Somatic cells maintain a healthy population of mitochondria by degrading damaged mitochondria via mitophagy, a specialized autophagy pathway. However, evidence from previous work investigating the more general macroautophagy pathway in oocytes suggests that mitophagy may not be active in the oocyte. This would leave the vast numbers of mitochondria - poised to be inherited by the offspring - vulnerable to damage. Here we test the hypothesis that inactive mitophagy in the oocyte underlies maternal transmission of dysfunctional mitochondria. To determine whether oocytes can complete mitophagy, we used either CCCP or AntimycinA to depolarize mitochondria and trigger mitophagy. After depolarization, we did not detect co-localization of mitochondria with autophagosomes and mitochondrial DNA copy number remained unchanged, indicating the non-functional mitochondrial population was not removed. To investigate the impact of an absence of mitophagy in oocytes with damaged mitochondria on offspring mitochondrial function, we utilized in vitro fertilization of high fat high sugar (HF/HS)-exposed oocytes, which have lower mitochondrial membrane potential and damaged mitochondria. Here, we demonstrate that blastocysts generated from HF/HS oocytes have decreased mitochondrial membrane potential, lower metabolites involved in ATP generation, and accumulation of PINK1, a mitophagy marker protein. This mitochondrial phenotype in the blastocyst mirrors the phenotype we show in HF/HS exposed oocytes. Taken together, these data suggest that the mechanisms governing oocyte mitophagy are fundamentally distinct from those governing somatic cell mitophagy and that the absence of mitophagy in the setting of HF/HS exposure contributes to the oocyte-to-blastocyst transmission of dysfunctional mitochondria."
|
| Older papers
|
| These papers originally appeared in the Some Recent Findings table, but as that list grew in length have now been shuffled down to this collapsible table.
See also the Discussion Page for other references listed by year and References on this current page.
- Unique insights into maternal mitochondrial inheritance in mice.[3] "In animals, mtDNA is always transmitted through the female and this is termed "maternal inheritance." Recently, autophagy was reported to be involved in maternal inheritance by elimination of paternal mitochondria and mtDNA in Caenorhabditis elegant. ...However, by using two transgenic mouse strains, one bearing GFP-labeled autophagosomes and the other bearing red fluorescent protein-labeled mitochondria, we demonstrated that autophagy did not participate in the postfertilization elimination of sperm mitochondria in mice. Based on these results, we conclude that, in mice, maternal inheritance of mtDNA is not an active process of sperm mitochondrial and mtDNA elimination achieved through autophagy in early embryos, but may be a passive process as a result of prefertilization sperm mtDNA elimination and uneven mitochondrial distribution in embryos."
- Recurrent tissue-specific mtDNA mutations are common in humans[6] "Mitochondrial DNA (mtDNA) variation can affect phenotypic variation; therefore, knowing its distribution within and among individuals is of importance to understanding many human diseases. Intra-individual mtDNA variation (heteroplasmy) has been generally assumed to be random. We used massively parallel sequencing to assess heteroplasmy across ten tissues and demonstrate that in unrelated individuals there are tissue-specific, recurrent mutations. Certain tissues, notably kidney, liver and skeletal muscle, displayed the identical recurrent mutations that were undetectable in other tissues in the same individuals. ...The most parsimonious explanation of the data is that these frequently repeated mutations experience tissue-specific positive selection, probably through replication advantage."
- Heteroplasmy of mouse mtDNA is genetically unstable and results in altered behavior and cognition[7] "Maternal inheritance of mtDNA is the rule in most animals, but the reasons for this pattern remain unclear. To investigate the consequence of overriding uniparental inheritance, we generated mice containing an admixture (heteroplasmy) of NZB and 129S6 mtDNAs in the presence of a congenic C57BL/6J nuclear background. Analysis of the segregation of the two mtDNAs across subsequent maternal generations revealed that proportion of NZB mtDNA was preferentially reduced. Ultimately, this segregation process produced NZB-129 heteroplasmic mice and their NZB or 129 mtDNA homoplasmic counterparts. Phenotypic comparison of these three mtDNA lines demonstrated that the NZB-129 heteroplasmic mice, but neither homoplasmic counterpart, had reduced activity, food intake, respiratory exchange ratio; accentuated stress response; and cognitive impairment. Therefore, admixture of two normal but different mouse mtDNAs can be genetically unstable and can produce adverse physiological effects, factors that may explain the advantage of uniparental inheritance of mtDNA."
|
Maternal Inheritance
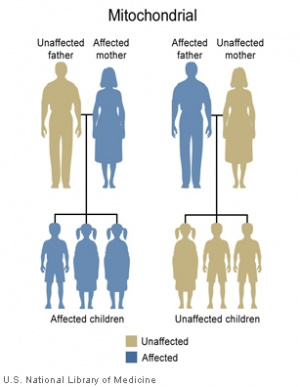
Mitochondrial genome inheritance
Most animals
- Oocyte mitochondria (maternal) are the only mitochondria inherited. (see genetics below)
- maternal mitochondrial genome inheritance.
- Spermatozoa mitochondria (paternal) can enter oocyte at fertilisation.
- Male spermatozoa are destroyed early in embryonic development (mechanism not yet elucidated)
- worm - (C. elegans) suggest ubiquitination occurs followed by autophagy.[8]
- mouse - suggest a more passive process, prefertilization sperm mtDNA elimination and uneven mitochondrial distribution in embryos.[3]
Paternal Mitochondria
| Spermatozoa organelles, including mitochondria, are destroyed in some species by the process of autophagy following fertilisation.[9]
Recent studies in mice suggest that additional mechanisms may be used in paternal mitochondrial elimination.[3]
|
|
- Links: spermatozoa
History Mitochodria
1857 Kölliker discovers mitochondria in muscle
1929 Karl Lohmann discovered ATP
1940s and 1950s ATP is formed in cell respiration in mitochondria and photosynthesis in chloroplasts of plants
1960 Efraim Racker and co-workers isolated, from mitochondria, the enzyme "F o F 1 ATPase" now call ATP synthase
1963 There’s DNA in those organelles DNA is directly visualized in first chloroplasts and then mitochondria, from the JCB Archive.
1992 Wallace identified degenerative disease caused by mtDNA mutations
1997 Nobel Prize in Chemistry - The three laureates have performed pioneering work on enzymes that participate in the conversion of the "high-energy" compound adenosine triphosphate (ATP).
- Paul D. Boyer and John E. Walker "for their elucidation of the enzymatic mechanism underlying the synthesis of adenosine triphosphate (ATP)"
- Jens C. Skou "for the first discovery of an ion-transporting enzyme, Na+, K+ -ATPase
Evolution Mitochondria
- primitive Eubacterium
- symbiotic relationship with eukaryotic cell
- circular DNA
- see antibiotic-induced deafness due to similarity of mitochondrial and bacterial ribosomes
- genes transferred to nucleus
- mitochondrial genome bp
- 366,924 Arabidopsis
- 16,569 Human
- 5966 Plasmodium
Mitochondrial Genome
Eukaryotic mitochondrial genomes
- In humans this genome is maternally inherited.
- Exists as multiple copies within the matrix of each mitochondrion within the cytoplasm of cells.
- In 1981 the human mitochondrial genome was sequenced.
- The genome is a small circular DNA molecule 16,568 bp in length containing 37 genes.
- 24 genes specify RNA molecules involved in protein synthesis (22 transfer RNAs (tRNA) and 2 ribosomal RNAs (rRNA))
- 13 genes encode proteins required for the biochemical reactions that make up respiration.
- Links: 16569 bp Homo sapiens mitochondrion | Mitochondrial DNA Deletion Syndromes
Spindle Transfer
(ST) An Assisted Reproductive Technology term referring to the transfer of the metaphase II-arrested (MII) spindle-chromosomal complex isolated as a karyoplast from a donor oocyte to the cytoplasm of a second recipient oocyte. The term cytoplast refers to the enucleated recipient oocyte. This technique has been suggested as a clinical solution to mitochondrial inherited disorders.
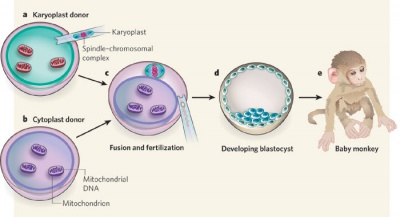
Swapping mitochondrial DNA mammalian oocytes[2]
- Links: Assisted Reproductive Technology | Mitochondrial DNA Deletion Syndromes | PMID 20539289
Oocyte Mitochondria
From a study of oocyte mitochondria during folliculogenesis in the cat ovary:[10]
- mitochondria DNA (mtDNA) copy numbers gradually increased as folliculogenesis progressed
- significant shift at the small antral stage (0.5 to <1 mm in diameter)
- location of mitochondria gradually shifted
- pre-antral oocytes - homogeneous cytoplasm distribution throughout
- advanced antral stage - pericortical concentration in the advanced antral stage
- progressive increase in mitochondrial activity in oocytes from the pre-antral to the large antral follicles.
Another study of obesity-exposed oocytes, show that they accumulate and transmit damaged mitochondria due to an inability to activate mitophagy.[5] Mitophagy is a term used for the selective degradation of mitochondria by autophagy.
- Links: ovary | oocyte | cat
Mitochondria Components
Outer Membrane
- porin - membrane channel, allows ions and metabolites into the mitochondria (<5000 daltons)
Intermembrane Space
- similar to the cytosol with respect to the small molecules it contains
- also enzymes that use ATP
Inner Membrane
- cardiolipin - phospholipid, makes membrane impermeable to ions (unique to mitochondria inner membrane)
- transport proteins - permeable to molecules required in the matrix
Cristae
- increase inner membrane surface area
- tubular, vesicular or flat cristae
- Adenosine triphosphate (ATP) synthase
- respiratory electron transfer chain proteins
- transport proteins
- Links: Model - inner boundary membrane and cristae membranes
Matrix
- metabolic enzymes of citric acid cycle (=Krebs) (100s of enzymes) (MH- do not need to know biochemical details of this cycle)
- genetic material DNA, tRNA, ribosomes
Mitochondria DNA
Eukaryotic mitochondrial genomes
- double stranded circular DNA (mitoDNA. mtDNA)
- 1981 complete human sequence (16,569 nucleotides)
- 37 genes
- encodes 13 polypeptides involved in oxidative phosphorylation
- remaining genes transfer RNA (tRNA) and ribosomal RNA (rRNA)
- multiple copies within the matrix
- maternally inherited
- remainder encoded by nuclear DNA
- proteins made in cytosol and imported into mitochondria
- Links: Home Reference - Mitochondrial DNA
Mitochondria Protein Synthesis
Many mitochondrial proteins are encoded by nuclear DNA
- synthesis begins in the cell cytoplasm
- imported into the mitochondria
- targeting similar to signal sequence for RER
- once in matrix signal sequence is cleaved (by Hsp70)
- protein then folds (by Hsp60)
- proteins for mitochondrial membrane or intermembranous space
- additional signal following matrix localization
Mitochondrial targeting signal (MTS) - alternating amino acid pattern (amphipathic helix) with a few hydrophobic amino acids and a few plus-charged amino acids at the N terminus.
Links: Replication and preferential inheritance of hypersuppressive petite mitochondrial DNA | Home Reference - Mitochondrial DNA
Mitochondrial Disorders
There has been recent discussion on the use of specialised Assisted Reproductive Technology (ART) techniques to allow replacement of maternal mitochondria with genetic abnormalities with a healthy donor mitochondria.
| Mitochondrial DNA disorder
|
Description
|
Prevalence
|
Life expectancy/morbidity
|
| Kearns–Sayre syndrome
|
Progressive blindness and blocked heart
|
Rare disease
|
Onset before 15
|
| Chronic progressive external opthalmoplegia (CPEO)
|
Progressive wastage of eyelids, eyes and sometimes facial muscles
|
Rare disease
|
Onset in young adulthood
|
| Pearson syndrome
|
Anaemia, pancreatic failure, muscle wastage
|
Very rare (less than 100 worldwide)
|
Early death
|
| Myopathy, encephalopathy, lactic acidosis and stroke (MELAS)
|
Stroke like episodes; muscle spasm; early dementia
|
Rare disease
|
Death before 40
|
| Myoclonic epilepsy and ragged-red fibres (MERFF)
|
Epilepsy, hearing loss, lactic acidosis, short stature
|
1 in 400,000 across Europe
|
Childhood onset
|
| Neurogenic weakness, ataxia and retinitis pigmentosa (NARP)
|
Muscle weakness, vision loss, learning disabilities
|
Rare disease
|
Onset in early childhood. Early death
|
| Maternally inherited Leigh syndrome (MILS)
|
Muscle weakness, heart and kidney failure, delayed development
|
Very rare
|
Onset in infancy. Death in early childhood
|
| Maternally inherited diabetes and deafness (MIDD)
|
Combination of all types of diabetes with deafness
|
Rare disease
|
Adult onset
|
| Leber hereditary optic neuropathy (LHON)
|
Rapid blindness
|
1 in 30,000 across Europe
|
Range from early childhood to 70s
|
| Myopathy and diabetes
|
Covers forms of muscular dystrophy
|
|
From infancy. Early death
|
| Sensorineural hearing loss
|
Covers a range of hearing loss through to deafness
|
Common condition but rarely caused by mitochondrial disease
|
Onset at any age
|
| Exercise intolerance
|
Range from lethargy to muscle wastage
|
1 in 8,000 but symptoms often combine with others
|
Onset in early life
|
Fatal infantile encephalopathy
(Leigh syndrome)
|
Brain and nervous system dysfunctions
|
Very rare
|
Onset in infancy. Death in early childhood
|
- Links: Mitochondria | Genetic Abnormalities | Zygote
|
Table based on Annex D, UK Mitochondrial Donation 2014.
|
- Links: Assisted Reproductive Technology
References
- ↑ Craven L, Murphy J, Turnbull DM, Taylor RW, Gorman GS & McFarland R. (2018). Scientific and Ethical Issues in Mitochondrial Donation. New Bioeth , 24, 57-73. PMID: 29529980 DOI.
- ↑ 2.0 2.1 Shoubridge EA. (2009). Developmental biology: Asexual healing. Nature , 461, 354-5. PMID: 19759608 DOI.
- ↑ 3.0 3.1 3.2 3.3 Luo SM, Ge ZJ, Wang ZW, Jiang ZZ, Wang ZB, Ouyang YC, Hou Y, Schatten H & Sun QY. (2013). Unique insights into maternal mitochondrial inheritance in mice. Proc. Natl. Acad. Sci. U.S.A. , 110, 13038-43. PMID: 23878233 DOI.
- ↑ Floros VI, Pyle A, Dietmann S, Wei W, Tang WCW, Irie N, Payne B, Capalbo A, Noli L, Coxhead J, Hudson G, Crosier M, Strahl H, Khalaf Y, Saitou M, Ilic D, Surani MA & Chinnery PF. (2018). Segregation of mitochondrial DNA heteroplasmy through a developmental genetic bottleneck in human embryos. Nat. Cell Biol. , 20, 144-151. PMID: 29335530 DOI.
- ↑ 5.0 5.1 Boudoures AL, Saben J, Drury A, Scheaffer S, Modi Z, Zhang W & Moley KH. (2017). Obesity-exposed oocytes accumulate and transmit damaged mitochondria due to an inability to activate mitophagy. Dev. Biol. , 426, 126-138. PMID: 28438607 DOI.
- ↑ Samuels DC, Li C, Li B, Song Z, Torstenson E, Boyd Clay H, Rokas A, Thornton-Wells TA, Moore JH, Hughes TM, Hoffman RD, Haines JL, Murdock DG, Mortlock DP & Williams SM. (2013). Recurrent tissue-specific mtDNA mutations are common in humans. PLoS Genet. , 9, e1003929. PMID: 24244193 DOI.
- ↑ Sharpley MS, Marciniak C, Eckel-Mahan K, McManus M, Crimi M, Waymire K, Lin CS, Masubuchi S, Friend N, Koike M, Chalkia D, MacGregor G, Sassone-Corsi P & Wallace DC. (2012). Heteroplasmy of mouse mtDNA is genetically unstable and results in altered behavior and cognition. Cell , 151, 333-343. PMID: 23063123 DOI.
- ↑ Hajjar C, Sampuda KM & Boyd L. (2014). Dual roles for ubiquitination in the processing of sperm organelles after fertilization. BMC Dev. Biol. , 14, 6. PMID: 24528894 DOI.
- ↑ Al Rawi S, Louvet-Vallée S, Djeddi A, Sachse M, Culetto E, Hajjar C, Boyd L, Legouis R & Galy V. (2011). Postfertilization autophagy of sperm organelles prevents paternal mitochondrial DNA transmission. Science , 334, 1144-7. PMID: 22033522 DOI.
- ↑ Songsasen N, Henson LH, Tipkantha W, Thongkittidilok C, Henson JH, Chatdarong K & Comizzoli P. (2017). Dynamic changes in mitochondrial DNA, distribution and activity within cat oocytes during folliculogenesis. Reprod. Domest. Anim. , 52 Suppl 2, 71-76. PMID: 28111812 DOI.
Reviews
Lima A, Burgstaller J, Sanchez-Nieto JM & Rodríguez TA. (2018). The Mitochondria and the Regulation of Cell Fitness During Early Mammalian Development. Curr. Top. Dev. Biol. , 128, 339-363. PMID: 29477168 DOI.
Articles
Floros VI, Pyle A, Dietmann S, Wei W, Tang WCW, Irie N, Payne B, Capalbo A, Noli L, Coxhead J, Hudson G, Crosier M, Strahl H, Khalaf Y, Saitou M, Ilic D, Surani MA & Chinnery PF. (2018). Segregation of mitochondrial DNA heteroplasmy through a developmental genetic bottleneck in human embryos. Nat. Cell Biol. , 20, 144-151. PMID: 29335530 DOI.
Boudoures AL, Saben J, Drury A, Scheaffer S, Modi Z, Zhang W & Moley KH. (2017). Obesity-exposed oocytes accumulate and transmit damaged mitochondria due to an inability to activate mitophagy. Dev. Biol. , 426, 126-138. PMID: 28438607 DOI.
Cieslak M, Pruvost M, Benecke N, Hofreiter M, Morales A, Reissmann M & Ludwig A. (2010). Origin and history of mitochondrial DNA lineages in domestic horses. PLoS ONE , 5, e15311. PMID: 21187961 DOI.
Search PubMed
Search Pubmed: Mitochondria
Terms
- maternal spindle transfer - (MST) An Assisted Reproductive Technology term. The “maternal spindle” is the group of maternal chromosomes within the oocyte, which are shaped in a spindle. The transfer technique involves removing the spindle from the mother’s oocyte before it is fertilised by the father’s spermatozoa. The spindle is then placed into a donor oocyte with healthy mitochondria, from which the donor’s spindle, and therefore her nuclear material, has been removed.
- Pro-nuclear transfer - (PNT) An Assisted Reproductive Technology term. The pro-nucleus is the nucleus of either a spermatozoa or an oocyte during the process of fertilisation after the sperm enters the egg, but before they fuse. Transfer involves removing the pro-nuclei from a newly fertilized oocyte that has unhealthy mitochondria. The pro-nuclei are then transferred into a donated embryo, with healthy mitochondria, that has had its own, original pro-nuclei removed.
| Cell Division Terms (expand to view)
|
meiosis | mitosis
- anaphase - (Greek, ana = up, again) Mitosis term referring to the fourth stage, where the paired chromatids now separate and migrate to spindle poles. This is followed by telophase.
- anaphase A - Mitosis term referring to the part of anaphase during which the chromosomes move.
- anaphase B - Mitosis term referring to the part of anaphase during which the poles of the mitotic spindle move apart.
- aneuploidy - (aneuploid) term used to describe an abnormal number of chromosomes mainly (90%) due to chromosome malsegregation mechanisms in maternal meiosis I.
- aster - (Latin, aster = star) star-like object visible in most dividing eukaryotic cells contains the microtubule organizing center.
- astral microtubule - spindle apparatus microtubule (MT) originating from the centrosome which does not connect to a kinetochore. These microtubules only exist during mitosis, the other spindle types are polar and kinetochore microtubules.
- autosomal inheritance - term used in hereditary diseases which means that the disease is due to a DNA error in one of the 22 chromosome pairs that are not sex chromosomes. Both boys and girls can then inherit this error. If the error is in a sex chromosome, the inheritance is said to be sex-linked.
- bivalent - (tetrad) a pair of homologous chromosomes physically held together by at least one DNA crossover.
- bouquet stage - meiosis term for when in prophase transition to the zygotene stage, the chromosome telomeres attachment to the inner nuclear envelope and form a cluster. This occurs before the onset of homologous pairing and synapsis. The name comes from the chromosomes resembling a "bouquet of flowers".
- diploid - (Greek, di = double + ploion = vessel) having two sets of chromosomes (2n), this is the normal euploidy state for all human cells, other than gametes that are haploid (n, a single set of chromosomes).
- diplotene stage- (diplotene phase, diplonema; Greek, diplonema = "two threads") meiotic stage seen during prophase I, the chromosomes separate from one another a small amount giving this appearance. In the developing human ovary, oocytes remain at the diplotene stage from fetal life through postnatal childhood, until puberty when the lutenizing hormone (LH) surges stimulate the resumption of meiosis. Prophase I, is divided into 5 stages (leptotene, zygotene, pachytene, diplotene, diakinesis) based upon changes associated with the synaptonemal complex structure that forms between two pairs of homologous chromosomes.
- euploidy - the normal genome chromosomal set (n, 2n, 3n) or complement for a species, in humans this is diploid (2n). The other classes of numerical chromosomal abnormalities include aneuploidy, polyploidy and mixoploidy.
- FUCCI - Acronym for Fluorescence Ubiquitination Cell Cycle Indicator a molecular tool for identifying the stage in the cell cycle. In G0/G1 cells express a red fluorescent protein and S/G2/M cells express a green fluorescent protein. (More? Tooth Development Movie)
- haploid - (Greek, haploos = single) Having a single set of chromosomes (n) as in mature germ/sex cells (oocyte, spermatozoa) following reductive cell division by meiosis. Normally cells are diploid, containing 2 sets of chromosomes. Ploidy refers to the number of sets of chromosomes in the nucleus of a cell.
- heteroplasmy - presence of more than one type of organellar genome. In humans this can refer to variations in the mitochondrial DNA (mtDNA). (More? PMID 26281784)
- homologous chromosomes - meiosis term for the two matching (maternal and one paternal) chromosomes that align during meiosis I.
- homologous recombination - meiosis term when DNA of homologous chromosomes is covalently exchanged to produce chromosomes with new allele combinations, and also links homologous chromosomes with each other to form a bivalent
- human genome - DNA within the 23 nucleus chromosome pairs and the cytoplasmic mitochondrial DNA.
- kinetochore - the protein structure formed on chromatids where the spindle kinetochore microtubules attach during cell division.
- kinetochore microtubule - spindle apparatus microtubule (MT) that attaches to the chromosome kinetochore by its plus end, the other spindle types are astral and polar microtubules.
- kinesin - a microtubule (MT) motor protein that exists in many isoforms and most move towards the MT positive end. Different isoforms have different functions within the spindle apparatus. PMID 20109570
- meiosis - reductive cell division required to produce germ cells (oocyte, spermatozoa) and for sexual reproduction. Note that only spermatozoa complete meiosis before fertilisation. Chromosome number is reduced from diploid to haploid, during this process maternal and paternal genetic material are exchanged. All other non-germ cells in the body divide by mitosis. (More? Meiosis | Spermatozoa Development | Oocyte Development | Week 1)
- meiosis I - (MI) the first part of meiosis resulting in separation of homologous chromosomes, in humans producing two haploid cells (N chromosomes, 23), a reductional division.
- meiosis II - (MII) the second part of meiosis. In male human spermatogenesis, producing of four haploid cells (23 chromosomes, 1N) from the two haploid cells (23 chromosomes, 1N), each of the chromosomes consisting of two sister chromatids produced in meiosis I. In female human oogenesis, only a single haploid cell (23 chromosomes, 1N) is produced. Meiosis II: Prophase II - Metaphase II - Anaphase II - Telophase II.
- meiotic silencing of unsynapsed chromatin - (MSUC) an aneuploidy protective mechanism for subsequent generations, during meiosis where chromosomes are silenced that fail to pair with their homologous partners.
- merotelic kinetochore - cell division abnormality in chromosomal attachment that occurs when a single kinetochore is attached to microtubules arising from both spindle poles. Normal chromosomal attachment in early mitosis, is by only one of the two sister kinetochores attached to spindle microtubules (monotelic attachment) later sister kinetochores attach to microtubules arising from opposite spindle poles (amphitelic attachment).
- metaphase - mitosis term referring to the third stage where mitotic spindle kinetochore microtubules align chromosomes in one midpoint plane. Metaphase ends when sister kinetochores separate. Originally based on light microscopy of living cells and electron microscopy of fixed and stained cells. A light microscope analysis called a "metaphase spread" was originally used to detect chromosomal abnormalities in cells. Mitosis Phases: prophase - prometaphase - metaphase - anaphase - telophase
- metaphase spread - In mitosis using light microscope analysis originally used to detect chromosomal abnormalities in cells, as chromosomes are only visible during cell division.
- microfilament - (MF) cytoskeleton filament normally required for cytoplasmic intracellular transport, motility and cell shape. Named by the actin monomers assembling into the smallest in cross-section of the three filament systems (microtubules and intermediate filaments). This system is disassembled and reassembled as the contractile ring for cytokinesis (cytoplasm division) following cell division mitosis and meiosis.
- microtubule - (MT) cytoskeleton filament normally required for cytoplasmic intracellular transport and motility. Named by the tubulin monomers assembling into "tubes", and are the largest in cross-section of the three filament systems (microfilaments and intermediate filaments). This system is disassembled and reassembled as the spindle apparatus during cell division.
- mitochondrial DNA - (mtDNA) multiple copies of a small circular DNA molecule located within the mitochondria matrix. In humans 16,568 bp in length containing 37 genes, originally inherited only from the oocyte (maternal inheritance).
- mitosis - (M phase) The normal division of all cells, except germ cells, where chromosome number is maintained (diploid). In germ cell division (oocyte, spermatozoa) meiosis is a modified form of this division resulting in reduction in genetic content (haploid). Mitosis, division of the nucleus, is followed by cytokinesis the division of the cell cytoplasm and the cytoplasmic contents. cytokinesis overlaps with telophase.
- p - chromosome short arm (possibly French, petit) and used along with chromosome and band number to indicate genes located on this arm of the chromosome. The chromosome long arm is identified as q (possibly French, tall) chosen as next letter in alphabet after p. These chromosomal arms are only seen when the chromosome is folded for cell division.
- polar microtubule - spindle apparatus microtubule (MT) that can arise from either pole and overlap at the spindle midzone. This interdigitating structure consisting of antiparallel microtubules is responsible for pushing the poles of the spindle apart. The other spindle types are astral and kinetochore microtubules.
- prometaphase - (Greek, pro = before) mitosis term referring to the second stage, when the nuclear envelope breaks down into vesicles. Microtubules then extend from the centrosomes at the spindle poles (ends) and reach the chromosomes. This is followed by metaphase.
- pronuclear fusion - (Greek, pro = before) the process of the fusion of the two haploid nuclear structures (pronuclei) contributed from the spermatazoa and oocyte to form the first diploid nucleus cell. Can also be called "fusion of pronuclei".
- pronucleus - (Greek, pro = before; plural, pronuclei) the two haploid nuclei or nuclear structures containing the genetic material from the spermatozoa and the oocyte. These two haploid nuclei will fuse together to form the first diploid nucleus cell, the zygote. Therefore the nuclear structures that exist "before the nucleus", the plural term is pronuclei.
- prophase - (Greek, pro = before) - mitosis term referring to the first stage, when the diffusely stained chromatin resolves into discrete chromosomes, each consisting of two chromatids joined together at the centromere.
- prophase I - meiosis term refers to the first phase of meiosis I, which together with meiosis II results in the reductive cell division only occurring gametes. Prophase can be further divided into a number of stages: leptotene zygotene, pachytene, diplotene, diakinesis.
- q - chromosome long arm (possibly French, tall), the next letter in alphabet after p, and used along with chromosome and band number to indicate genes located on this arm of the chromosome. The chromosome short arm is identified as p (possibly French, petit). These chromosomal arms are only seen when the chromosome is folded for cell division.
- S phase - during interphase of cell cycle where DNA is duplicated prior to second growth period (G2 phase) that is followed by mitosis (M phase).
- synapsis - (syndesis) meiosis term for the pairing of two homologous chromosomes that occurs during prophase I.
- synaptonemal complex - meiosis term for a protein structure essential for synapsis of homologous chromosomes. (proteins SCP3 and SCP1).
- telomere - region found at each end of the chromosome and involved in cellular ageing and the capacity for division. The regions consist of repeated sequences protecting the ends of chromosomes and harbour DNA repair proteins. In the absence of the enzyme telomerase, these regions shorten during each cell division and becoming critically short, cell senescence occurs.
- telophase - mitosis term referring to the fifth stage, where the vesicles of the nuclear envelope reform around the daughter cells, the nucleoli reappear and the chromosomes unfold to allow gene expression to begin. This phase overlaps with cytokinesis, the division of the cell cytoplasm.
- telomerase - the enzyme that maintains the chromosome end length, the telomeres, involved in cellular ageing and the capacity for division. Absence of telomerase activity leads to the chromosome ends shorten during each cell division, becoming critically short and cell senescence then occurs.
- tetrad - (bivalent) a pair of homologous chromosomes physically held together by at least one DNA crossover.
|
|
|
External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2026, July 5) Embryology Mitochondria. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Mitochondria
- What Links Here?
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G





