Book - Contributions to Embryology Carnegie Institution No.69
| Embryology - 28 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Streeter GL. Development of the auricle in the human embryo. (1922) Carnegie Instn. Wash. Publ. 277, Contrib. Embryol., 14: 111-138.

| Historic Disclaimer - information about historic embryology pages |
|---|
 |
Development of the Auricle in the Human Embryo
Carnegie Institution of Washington - Contributions to Embryology

(6 plates, 8 text-figures) 111-138 (1922)
Carnegie Institution of Washington, Department of Embryology.
Introduction
In order to obtain objective criteria for the determination of the age of human embryos, it has been found necessary to seek characteristics other than size. The practice of basing such determinations on the length of the specimen, which is the custom at the present time among anatomists, has proved in several respects unsatisfactory, particularly as young embryos vary greatly in length according to the posture in which they happen to undergo fixation, and for the further reason that, when placed in formalin or other fixing solution, embryos become distended by the solution to a degree that adds considerably to their length and weight. This increase in volume varies according to the size of the specimen and the condition of its tissues. Smaller specimens undergo a greater relative increase than the larger ones and fresh specimens greater than macerated ones. Furthermore, this acquired distention gradually disappears and hence the size or weight of a given specimen will vary according to the time that has elapsed since its fixation.
These sources of inaccuracy, which are of disturbing importance in the case of young embryos, are of less importance in larger fetuses, because in these it is possible to standardize more accurately the measurements and to control fully the posture of the specimen. Also, in large fetuses the factor of distention by the fixative is of less moment ; the increasing imperviousness of the integument retards the absorption of the fixative solution and the weekly increment in size reaches proportions that render the fixative distention a factor of progressively diminishing importance.
The period during which length is particularly unreliable as an indication of the age of a specimen, and for which we are in the greatest need of more accurate criteria of development, is the first two months; that is, from the earliest stages up to about 30 mm. length. This was pointed out by Mall (1914), who proposed the subdivision of this period into stages, based upon the development of external features, such as the branchial arches, arms, and legs.
In attempting this standardization it soon became apparent that it would be necessary to survey the details of the external form more carefully than had
previously been done. This meant the study of more specimens and better photographic records, so planned as best to display individual regions. This is particularly true of the human embryo, where the difficulty of distinguishing between real and accidental differences is increased by the varied conditions under which the material for study is obtained. It was, in fact, the recognition of such a need that led Spaulding (1921) to make a detailed study of the steps in the differentiation of the external genitalia. The successful outcome of his investigation testifies to his wisdom in limiting his attention to a definite region. It is clear that before a satisfactory series of developmental stages, based on external form, can be arrived at, it will be necessary to study separately each part of the body and establish the normal sequence of differentiation, region by region.
What was done by Spaulding for the external genitalia I have endeavored to do for the branchial region, and it is my purpose in the following pages to outline what seem to be the significant morphological features in the transformation of the tissues in the neighborhood of the first gill-cleft into the definitive auricle.
Historical
Most of the investigators who have published accounts of the development of the auricle have shown a lively interest in the branchial hillocks and have placed great emphasis on them as the essential factors in the acquirement of the final form of the auricle. I am of the opinion that too much importance has been attributed to these hillocks, and that the auricle, instead of being a composite structure the fused product of a group of separate and discrete masses comes into existence as an intact and continuous primordium, which, by the ordinary processes of differentiation, gradually becomes elaborated into its final form. It arises, for the most part, from the mesenchymal cells of the hyoid bar; the overlying ectoderm, also, may play an important role in its determination. It is possible that it is entirely of hyoid origin and that the mandibular elements are nothing more than the product of cells that have migrated forward into that region. In support of this idea is the fact that the mandibular parts, when first seen, are mostly in the deeper levels. However that may be, as soon as one can begin to outline the condensed tissues constituting its primordium, the whole auricle is continuous and exhibits the essential contours of the mature structure.
Before entering into this subject more fully, it might be well to outline the principal steps in our present knowledge regarding the development of the auricle.
To make the history brief, condensed abstracts of the more significant observations will be given in chronological order, as far as I have been able to follow them.
Moldenhauer (1877), in a careful study of the development of the middle and external ear of the chick, discovers the occurrence of two pairs of hillocks on the first and second branchial arches, which he terms colliculi branchiales externi. He regards these as connected with the development of the external auditory meatus, the tragus being derived from the first arch and the anti-tragus from the second arch. They are present on the sixth and seventh days of incubation, and on the eighth day they become transformed into the definitive parts of the meatus. Thinking of the head as erect, with its longitudinal axis in the vertical plane, the author speaks (p. 118) of the hillocks in front of the first gill-cleft as "superior" and those behind the first gill-cleft as "inferior." The ventral pan he calls "anterior" and the dorsal pair "posterior." Thus, the hillocks of the mandibular bar become, respectively, colliculus posterior superior and colliculus anterior superior, and the hillocks
of the hyoid bar become colliculus posterior inferior and colliculus anterior inferior.
His (1882), in describing the external form of human embryos between 12 and 30 mm. long, briefly mentions the occurrence of branchial hillocks around the first gill-cleft, similar to those found by Moldenhauer in the chick. Instead of four, however, he finds six. There are two hillocks on the mandibular arch, the lower one becoming the tragus, the upper becoming the spina helicis. At the upper end of the first gill-cleft is the colliculus intermedius, which forms all of the helix with the exception of the spina. On the hyoid arch he finds three hillocks, the upper one becoming the anthelix and the next lower the an ti tragus; the lowest one seems to disappear and become covered in by the tragus.
His (1885), in describing the development of the ear, gives a classical account of the auricular hillocks (or tubercles, as he names them) which has dominated all
subsequent literature. He describes the auricle as arising from the nodular edges that surround the first gill-cleft, very early showing a subdivision into six hillocks, which he numbers consecutively 1 to 6. He divides the mandibular arch into a dorsal and a ventral portion. On the ventral portion is the first hillock (tuberculum tragicum). The remainder of this portion takes no further part in the formation of the auricle, becoming the lip-ridge and jaw-ridge, the latter subsequently covering in and fusing with hillock 6. Hillocks 1 to 5 form a plump ring surrounding the first gill-cleft, which thus becomes the fossa angularis. In this process hillocks 1 and 2, also 2 and 3, partially fuse. Between 3 and 4 there is a deep furrow; 3 is continued as a tail caudal to 4 and loses itself in the neighborhood of hillock 6. The eventual helix is formed by the union of hillocks 2 and 3, together with the tail-like process extending from the latter. The anthelix is derived from hillock 4, the lobule from hillock 6. The taenia lobularis is a remnant of 6. The tragus is derived from hillock 1, the antitragus from hillock 5. In addition to these hillocks, the author describes a tuberculum centrale, which takes the form of a transverse elevation in the floor of the fossa angularis, separating the upper and lower depressions. It consists of a connective-tissue pillow or swelling of the closure plate of the first gill-cleft. The cartilaginous strand belonging to the second arch extends into it. It contains a small blood-vessel, the stapedius artery. The furrow between hillocks 1 and 5 he designates as the sulcus antitragicus, while the lower end of the fossa angularis he calls the incisura inter tragicus. The crus, or spina helicis, is derived from a fusion of hillocks 2 and 4.
Kastschenko (1887), in a study of the fate of the mammalian gill-clefts, in which he concerns himself particularly with the thymus and thyroid, describes the external auditory canal of the pig, which, he points out, is a secondary formation, its tip only being a true remnant of the first epidermal pocket. He pictures five auricular hillocks, as seen in 12, 13, and 15 mm. specimens, but does not clearly trace them into the eventual ear. Kastschenko's figures correspond fairly well with the description given by His for the human, with the exception of the fifth and sixth tubercles. Kastschenko's hillock 5 seems to correspond to His's hillock 6.
Tartaroff (1887) reports a relationship between the character of the skin and
the underlying cartilage covering the auricle, particularly as to the presence of hair
and subcutaneous fat. The growth of the cartilage results in tension of the skin,
which he regards as the cause (pressure atrophy) of the lack of fat and the disappearance of hair, and it is inferred that the resistance of the skin may explain the folding of the ear cartilage.
Gradenigo (1888) presents a study of the development of the auricle in a series
of mammals, including man, pig, cat, guinea-pig, rabbit, sheep, and cow. In the
last three his studies were controlled by serial sections. Like the previous authors,
he found six hillocks, three on the mandibular arch and three on the hyoid arch.
According to Gradenigo, the hillocks flatten out and tend to fuse together. The
four lower ones, by closing in around the lower hyomandibular groove, form the
external auditory meatus and the floor of the future fossa angularis. In doing this
the hillocks disappear for the most part. The hillocks. do not form the auricle
proper; this arises through the fusion of two elevations immediately adjacent to
the hillocks, which the author names helix hyoidalis and helix mandibularis. These
elevations appear at about the time of the flattening out of the hillocks. In later
stages these two elevations fuse above and below, thus surrounding the region of
the hillocks and thereby forming the auricle. The hillock region corresponds to
the fossa angularis and becomes the future concha and the entrance to the external
auditory meatus. In tracing the formation of these elevations from which the auricle
is derived, Gradenigo points out that the helix hyoidalis first makes its appearance just behind the middle hyoid hillock and from there spreads behind the other
two hyoid hillocks. Its upper end arches forward over the region of the hillocks.
At this stage in its development we have a structure resembling the cauda of the
third hillock of His. The helix mandibularis makes its appearance somewhat later
than the helix hyoidalis, its upper part being better developed than the lower part.
The lower part forms the tragus. In addition to fusing above and below, the helices
develop processes which extend transversely across the fossa angularis. One of
these becomes the eventual crus helicis, and another forms part of the cms inferius
anthelicis. The other processes become lost. The paper is not very well illus-
trated, so that it is difficult to follow the author's description in detail. However,
he reviews the pathology of this region and gives an account of a variety of teratological conditions. He points out that the lobule makes its appearance later in
man than in other mammals and that it is derived from the growth of the lower end
of the helix hyoidalis.
His (1889) gives a morphological description of the adult auricle in man. He goes into particular detail regarding the lower part of the ear, especially the lobule.
Schwalbe (1889), in the first of a series of important papers on the development of the auricle, briefly describes the form of the auricle in human fetuses ranging
from 60 to 180 mm. sitting height, and in doing so he introduces the more accurate technique of physical anthropology. He points out that the crown of the ear
(satyr tip) is not the same as the Darwin angle. The Darwin angle is the true eartip and first makes its appearance in the human fetus about the middle of the third
month. It becomes less distinct in the later months, due to its thickening and the rolling in of its edge. The rolling in of the ear he regards as a reduction process.
Schwalbe (1891a), in his next paper, discusses the Darwin tubercle (i. e., the true ear-tip) as it occurs in adult man. He describes six degrees of its occurrence, varying from the most pronounced type, resembling the Macacus form, to the least marked, where no trace of the ear-tip can be recognized. He explains the increase in ear dimensions, occurring with advancing age, as due to the flattening out of the various folds of the auricle. This he regards as connected with the loss of elasticity of the elastic fibers of the skin and cartilage, and as related to the wrinkling of the skin which accompanies loss of elasticity in the aged.
In his next paper, Schwalbe (1891b) points out the significant fact that in reptiles that lack an external ear (lizard and turtle) there occur distinct hillocks in the embryo,resembling those in vertebrates that develop an auricle. These hillocks undergo degeneration and are reduced to the level of the surrounding skin. He finds in both birds and reptiles hillocks corresponding to the tragus and antitragus hillocks of His. These animals have one hillock (Auricularkegel), situated dorsal to the first cleft, which seems to represent a more primitive apparatus than is present in mammals, although it may be related to the helix system. In Salachians it possesses a spiracle.
Schaeffer (1892-1893), reviewing the embryonic stages of the auricle, endeavors to trace them to their phylogenetic representatives in adult mammals. He describes the six hillocks as found in the embryo and notes their change in form in the 18-mm. embryo, which change he regards as due to opacities of the covering skin. The opacities are produced by cell accumulations, which usher in the fibro-cartilage of the auricle. The first part of the auricle to make its appearance is the inferior-posterior part of the helix. This is followed by the tragus and antitragus and finally (20 mm.) by the crus helicis. Schaeffer points out that the anterior cms of the anthelix is present in all mammals. The folds of the anthelix, which can be seen in the 50-mm. embryo, are present only in Primates. The lobule is a later acquisition and is found only in anthropoids and man.
In 1897 Schwalbe published an account of the development of the auricle in the human embryo which ranks with that of His (1885) in having dominated all subsequent descriptions. He describes the six hillocks substantially in the same manner as was done by His. The auricle, however, he regards as quite separate in origin from the hillocks. It appears as a fold of skin, resembling an eyelid, caudal to hillocks 4 and 5. (This fold of Schwalbe's corresponds fairly closely to the helix hyoidalis of Gradenigo.) From the region corresponding to hillocks 2 and 3 is formed the helix ascendens, the lower end of which becomes the crus helicis. Above, the helix ascendens is continuous over the first gill-cleft with the main ear-fold, the point of union being sharply kinked and corresponding to the crown angle (satyr tip) of the mature ear. The helix ascendens does not exactly correspond to the helix mandibulars of Gradenigo, in that the tragus is not derived from its lower end. Schwalbe derives the tragus from hillock 1, as did His; the antitragus
he derives from hillock 6. Like Gradenigo, he derives the lobule from the lower end
of the ear-fold (helix hyoidalis). He traces hillock 4 into the anthelix system,
especially into the inferior crus of the anthelix. The crista anthelicis inferior is
probably derived from hillock 5. With the further development of the free ear-
fold, three important angles can be recognized along its margin: (1) at the junction
of the helix ascendens and the ear-fold, the crown angle or satyr point; (2) in the
middle of the ear-fold, the posterior angle or Darwin point; and (3) at the lower end of the ear-fold, where it merges into the lobule, the posterior-inferior angle.
Schwalbe points out that one can draw a straight line separating the hillock region
from that of the free ear-fold. This line falls above the upper end of the helix
ascendens and passes down, posterior to the antitragus, to the point of junction
of the lobule and the free ear-fold. The hillock region lies in front of this line and
is more or less constant in all types of auricles; the free ear-fold lies posterior to
the line, and the degree of development of this part of the ear is the chief factor
in producing the different types of ears found in various mammals The author
describes the occurrence, during the fourth month, of ridges in the free ear-fold,
which he regards as the temporary presence of the longitudinal folds that become
permanent in some of the other mammals. His paper is accompanied by an instructive table in which are listed the separate hillocks, their embryological desig-
nations, and the part each takes in the formation of the definitive auricle, including
the terminology of His and Gradenigo.
Miinch (1897) describes the morphology of the auricular cartilage in human embryos 20, 48, 57, 96, and 142 mm. long, as seen in wax-plate reconstructions. In studying its histogenesis, the author notes the close relationship existing between the cartilage and the ectoderm and describes the characteristic appearance of the ectoderm over the auricular region. He alludes to the relatively large size of the spina helicis in early stages and its subsequent tendency to become pinched off. It seems never to become completely detached in man, but does so in other animals. It is then designated scutulum.
Ruge (1898) presents a comparative anatomical study (Ornithorhynchus and Echidna) of the cartilage of the auricle. He bases his argument on its adult
connections, regarding the cartilage of the auricle as a derivative of the hyoid arch.
The tympanic end of the cartilage of the external auditory meatus is most closely
connected with the hyoid by connective tissue and common musculature. In
tracing it peripherally, its medial terminal part becomes the concha, and the
lateral terminal part becomes the tragus. The author emphasizes the unity of
the external auditory meatus and the auricle.
Hammar (1902), in describing the development of the middle ear and external auditory meatus in man, points out that the fossa concha? (angularis) certainly arises directly from the first branchial cleft, and thus we have as derivatives of the
first cleft the incisura intertragicus, cavitas conchae, and cymba concha?. All the
other furrows of the auricle are secondary. In referring to the hillocks, the author
states that he does not find that they take part in the formation of the floor, but
rather that the auricle is derived from two ridges that are independent of the
auricular hillocks, somewhat as described by Gradenigo (helix mandibulars and
helix hyoidalis) . The hillocks are not so sharply marked as has been indicated by
previous writers. They consist only of slight thickenings of a more or less uniform
subepidermal connective-tissue layer. Hammar regards it as artificial to describe
them as independent structures which shove over and fuse with one another.
He makes the important observation that the hillocks are more or less absorbed
in the swellings from which the auricle is derived.
Schmidt (1902) made a comparative anatomical study of the auricle, with examples from the following orders: Primates, Prosimise, Rodentia, Perissodactyla, Artiodactyla, and Carnivora. The paper is accompanied by a limited number of excellent drawings. The author regards the human auricle as rudimentary and finds that most of its morphological characteristics can be recognized in the ear of other mammals.
Baum and Dobers (1905) describe the development of the auricle in the pig and sheep. In the early pig embryo six auricular hillocks are found, corresponding
closely to the His description for man. Hillock 1 can not be recognized in the sheep ;
in the pig it becomes the tragus. The ear-fold is derived from hillocks 4, 5, and 6.
Hillocks 2 and 3 acquire cartilage and form the cms and helix ascendens. Hillocks
4, 5, and 6, in addition to forming the ear-fold, become elongated into three longitudinal ridges which constitute the anthelix. Hillocks 2 and 5 fuse and create a
transverse ridge which divides the fossa angularis into a dorsal part (scapha) and a
ventral part (concha), which is continuous with the external auditory meatus.
This paper is accompanied by very few figures of the earlier stages, so that it is not
possible to follow accurately the transitions referred to by the authors. They
describe the development of the scutulum and find that it has the same origin as
the auricular cartilage and is a derivative of it. They regard it as identical with
the spina helicis of man, which has become detached by the pull of the massive
anterior auricular cartilage.
Keith (1906) gives the results of an anthropological study of the mature auricle, with the view of determining the relation of one group of people to another, his records extending to 8,567 males and 6,577 females, belonging to Germany, Scotland, England, Wales, and Ireland, and including representatives of the insane, criminal, and vagrant classes. He regards it as unlikely that we shall obtain any light on racial affinities from the study of the form of the auricle.
Henneberg (1908) describes the development of the auricle in the rat, rabbit, and pig. His descriptions are accompanied by a series of excellent illustrations, which give the principal stages of development from the time of the formation of the auricular hillocks until the auricle has acquired its mature characteristics. The fate of the individual hillocks appears to be the same in the three forms studied. Henneberg differs from Schwalbe chiefly in regard to hillocks 4 and 5, which, according to him, give origin directly to the ear-fold (primitive scapha). By the fusion of hillocks 1 and 6 the first gill-cleft becomes converted into the fossa angularis. Through the undermining of the surrounding wall this fossa becomes converted into the concha, while the wall itself gives origin to the tragus, antitragus, helix, and parts of the definitive scapha. In all of the three animals studied, the inner surface of the scapha shows the presence of longitudinal ridges which are derived from the hyoidal hillocks. In the rodents these disappear, but in the pig they remain as the permanent longitudinal folds.
In 1910 Henneberg made a study of the function of the auricle, in which special attention is given to the closure mechanism as it occurs in a variety of mammals. He believes that in man the auricle serves not only as a sound collector but also as a closure mechanism whose function has remained rudimentary. The presence of the anthelix, the small size and rolled-in character of the helix, and the rudimentary character of the auricular muscles are all regarded by him as evidences of the reduction of the auricle in man.
Boas (1912) published the results of a comparative anatomical study of the mammalian adult auricle and its contained cartilage. The work is accompanied by an excellent series of plates illustrating the matter exhaustively. The author has perfected a method of preparing the ear-cartilage so that it can be completely flattened out, thus greatly increasing the possibilities of comparing one form with another. He introduces a new terminology which simplifies the analysis of the different parts of the cartilage. The term plica principalis, used for the cms inferius anthelicis, is a term that will surely be of the greatest value.
Schwalbe (1916), in a comparative anatomical study of the primate auricle, summarizes and extends his previous studies on this subject. He still regards the auricular hillocks as the basis, in all mammals, for the form of the outer ear. The fact that they are present in reptiles he regards as proof that the organ, which first reaches its characteristic form in mammals, may make its appearance in earlier stages of phylogenetic development. In human embryos the hillocks become modified into a hillock region, whereas there is a fold back of hillocks 4 and 5 from which is formed the free ear-fold or scapha. The variations in this free ear-fold account for the chief differences in ear-tips. The author points out that in those animals that live in water, in subterranean burrows, or in trees the ear-fold is reduced, whereas these forms retain the hillock region, which serves to protect the entrance to the external auditory meatus. The free ear-fold is greatly increased in nocturnal animals. It is of interest to note that similar types of ears may occur in diverse forms living under similar conditions.
Sera (1917) maintains that the human auricle, with the folded helix and without the Darwin tubercle, constitutes the original and primary form. The unfolded ear with the Darwin tubercle represents an arrest of development and has no phylogenetic significance.
Terminology
The terminology of the external ear now in general use is a purely descriptive one and is based upon the form usually met with in the human adult. In its establishment scant attention has been given to the embryonic stages and as little to the ear of other animals. It is therefore not surprising than one finds the terminology more or less inadequate for any critical analysis of the auricle or for the study of any other ear than that of adult man. When the appropriate time comes, the nomenclature of the external ear will benefit, as much as that of any other part of the body, by a thorough rt -deration. In this paper I shall depart but little from the prevalent terminology and then only where it seems unavoidable. As can be seen in figure 2, the following new terms have been utilized: fossa articularis superior, for fossa triangularis; fossa articularis inferior, for cymba concha?; plica principalis (introduced by Boas, 1912), for cms inferius anthelicis; crus helicis, to include all that part of the helix derived from the mandibular arch ; on the median side of the cartilage corresponding to the articular fossae: eminentia articularis superior, for eminentia fossa triangularis; and eminentia articularis inferior, for eminentia conchae.
It may be pointed out here that the two articular eminences (fig. 1) are continuous with each other anteriorly, and that together they constitute a relatively
rigid, bowl-shaped base from which the auricle is suspended. It is this part only
of the auricular cartilage that offers a contact surface suitable for its attachment
to the skull, and it may therefore be designated as the pars articularis. It is chiefly
the inferior eminence that contributes to the surface, although the anterior and lower portions of the superior eminence also take part. The band-like fenestrated cartilage surrounding the external acoustic meatus likewise has a bony attachment, but
this is quite different in character from the pars articularis; it may be compared
rather to the tracheal rings, serving as a mechanism to prevent collapse of the
meatus. In structure and position it offers little if any support to the auricle.

Figs. 1 and 2. Human adult auricle, illustrating terminology used in this paper. In figure 1 the auricular cartilage is viewed from the median side, thus showing the two eminences which constitute its main area of contact with the skull. In figure 2 can be seen the cavities (fossae articulares) of these eminences and the plica principalis projecting between them as a strengthening ridge.
For the convenience of the reader I am appending a glossary containing the principal terms met with in the literature dealing with the development of the auricle. In some instances the author who introduced the term is mentioned.
Glossary
Note some of the terminology below is historic and may not be currently used.
Anteron
(Boas]. By macerating the cartilage of the auricle and auditory canal the whole system can be unrolled into a flat plate. When this is done the plate presents, along its anterior and posterior margins, and particularly in its proximal half, a scries of incisures which divide the contour into a corresponding series of processes. The processes along the anterior margin are designated anteron 1, anteron 2, etc., numbering from the base of the cartilage. The processes along the posterior margin are designated posteron 1, etc. In the typical mammalian ear, anteron 5 corresponds to the spina helicis, posteron 4 to the tragus, and posteron 5 to the antitragus.
Anthelix
The rounded brim of the concha, from which the secondary part of the. auricle flares out as the scapha-helix. (See fig. 2.)
Antitragus
The thickened ventral rim of the concha, situated between the incisura intertragica and the anthelix. Apparently a part of the closure mechanism.
Cartilago-scutiformis
or cartilago-scutularis. See Scutulum.
Cauda helicis
(1) Term applied to the terminal process of the cartilage of the helix, which is separated from the conchal cartilage by a cleft (fissura antitragico-helicina). His designated that part of it forming the skeletal part of the lobule as lingula auricula. (2) The term applied by His to the fold found in the embryo, extending from the third auricular hillock, directly posterior to the fourth and fifth hillocks. According to that author, the adult helix is derived from it.
Cavitas conchas
See Concha.
Colliculi branchiales externi
[Moldenhauer]. The name originally given to the hillocks that appear in the embryo on the first and second branchial arches.
Concha
The shell-shaped primary part of the auricle immediately surrounding the meatus. As previously used, the term included only the cymba conchas and the cavitas concha?. In this paper I have extended the term to include also what has been known as the fossa triangularis. The contour of the concha thus is outlined by the tragus, incisura intertragica, antitragus, anthelix, and crus helicis.
Crista inferior anthelicis
[Schwalbe]. Used synonymously with crus inferius anthelicis or plica principalis.
Crus helicis
Formerly restricted to the horizontal portion of the helix, forming a transverse ridge in the floor of the concha. In this paper the term is extended to include all that part of the helix derived from the mandibular arch. (See fig. 2.) It constitutes the lateral free edge of the pars articularis concha?, differing in structure and development from the remainder of the helix.
Crus inferius anthelicis
Fold in the auricular cartilage extending forward from the anthelix and separating the fossa triangularis from the cymba concha?. Equivalent to plica principalis, which is a better term.
Crus superius anthelicis
Ridge limiting the upper border of the fossa triangularis (fossa articularis superior). In using the term concha to include this fossa, the crus superius anthelicis becomes merely the upper end of the anthelix itself.
Crus supertragicum
[His]. A process sometimes extending forward from the crus helicis to the region just above the tragus. Also called antitragicum [Gradenigo].
Cymba conchae
See Concha.
Darwin's tubercle
See Tuberculum auricula.
Eminentia articularis inferior
See EminenUa articularis superior. Formerly known as eminentia concha.
Eminentia articularis superior
Same as eminentia fossae triangularis. The pars articularis of the concha, as viewed from the median side, presents two eminences which constitute the chief area of contact of the auricle with the skull. In this paper these are designated, respectively, eminentia articularis superior and eminentia articularis inferior. (See fig. 1.) The groove between them is the sulcus corresponding to the plica principalis.
Fissura antitragico-helicina
Cleft separating cartilaginous cauda helicis from conchal cartilage.
Fossa angularis
[His]. Name applied to the first branchial cleft when modified by the formation of the auricular hillocks, five of which form a plump ring around it.
Fossa articularis inferior
Same as cymba concha. See Fossa articularis superior.
Fossa articularis superior
Same as fossa triangularis. When the pars articularis concha? is viewed from the lateral side, its floor presents two fossa? (superior and inferior) separated by the plica principalis. (See fig. 2.)
Fossa conchae
[Hammar]. Essentially the same as fossa angularis.
Fossa intercruralis
Same as fossa triangularis, or, as used in this paper, fossa articularis superior.
Fossa scaphoidea
See Scapha.
Fossa triquetra
Same as fossa triangularis, or, as used in this paper, fossa articularis superior.
Free ear-fold
or freien Ohrfalte [Schwalbe]. The ridge representing first appearance of definitive auricle. Same as helix hyoidalis [Gradenigo], cauda helicis [His], or primitive scapha [Henneberg].
Helix
In adult man the rolled-in margin of the auricle, when viewed as a whole from the lateral side, resembles in outline a coiled spring and on this account it was termed helix. Included under it are parts that are quite different, both embryologically and structurally. Furthermore, it is not applicable to the auricle of other animals. If the term scapha be used for all of the auricle peripheral to the anthelix, and the term helix used for the rolled edge of the scapha, where this occurs, the difficulty is then largely removed. It is so used in this paper, and under scapha-helix will be designated only those parts of the secondary auricle derived from the hyoid arch. The crus helicis is a different structure. The lobulus auricula? is a part of the secondary auricle and bears a similar relation to the concha as does the scapha. (See fig. 2.)
Helix ascendens
[Schwalbe]. The anterior portion of the helix which is derived from the third auricular hillock of the mandibular arch. Partially synonymous with crus helicis, as used by me.
Helix hyoidalis
[Gradenigo]. That portion of the helix derived from the hyoidal arch, from a fold posterior to the fourth, fifth, and sixth auricular hillocks. Same as cauda helicis [His] and helix posterior [Schwalbe].
Helix mandibularis
[Gradenigo]. Fold found in the embryo directly in front of the third auricular hillock and extending down in front of the second and first hillocks. According to Gradenigo, this fold gives origin to the anterior end of the helix, crus helicis, and tragus.
Helix posterior
[Schwalbe]. That portion of the helix derived from the hyoidal arch. Same as helix hyoidalis [Gradenigo].
Incisurae cartilaginis meatus auditorii externi
[Santorini]. Clefts in cartilage of external meatus, somewhat analogous to the clefts between the cartilaginous rings of the trachea.
Incisura intertragica
The cleft between the tragus and antitragus. A derivative of the lower end of the first branchial cleft.
Lamina tragi
Cartilaginous plate supporting tragus.
Lingula auricula
[His]. See Cauda helicis.
Lobulus auriculae
The free edge of the auricle below the antitragus continuous with the scapha helix. See Helix.
Margo oralis helicis
[Baum and Dobers]. Anterior free border of auricle, particularly in such animals as the pig. In a similar way the posterior border is referred to as aboral.
Pars articularis concha
The upper half of the concha. It includes the two articular fossa? (eminentia?), plica principalis, crus helicis, and spina helicis. (See figs. 1 and 2.)
Plica auricularis longitudinalis cranialis
[Henneberg]. The most cranial of the three longitudinal folds of the scapha in such animals as the pig. The others are designated medialis and caudalis, respectively.
Plica principalis
[Boas]. Equivalent to crus inferius anthelicis. Introduced because it is more accurately applied, particularly to the auricle of mammals other than man.
Ponticulus
Ridge on inner surface of conchal cartilage downward from the inferior articular eminence. It appears to be concerned with the ligamentous attachment of the auricle.
Posteron
[Boas]. See Anteron.
Rima helicis
[Albinus]. Perforation of the cartilage of the crus helicis.
Satyr-tip
[Schwalbe]. The tip of the auricle toward the crown of the head. Also called crown-lip or crown-angle.
Scapha
Concave surface of the free portion of the auricle lying between the anthelix and the helix. Term applied by Henneberg to the entire free auricle from the anthelix to the free border. He applies the term helix to the unwrinkled border of the scapha.
Scapha primitiva
[Henneberg]. Same as free ear-fold.
Scutellum
See Scutulum.
Scutulum
(Also known as scutellum, cartilago-scutiformis, or cartilago-sculularis.) This is supposed by some writers to be simply an enlarged spina helicis which has become detached. According to Schmidt, it is an accessory cartilage, connected with the complicated muscular apparatus, which is provided for the auricle of some mammals and is entirely absent in man. See Spina helicis.
Spina helicis
Cartilaginous process extending forward from the pars articularis concha?. (See fig. 1.) It is not in reality a part of the helix. It is supposed that this structure is enlarged and becomes detached in some mammals to form the scutulum.
Taenia lobularis
The fold attaching the lobule to the parotid region. In the embryo it appears before the lobule itself, being derived from the ventral end of the hyoid bar below hillock 6. It is the extension and widening of the taenia as a free fold, to join the lower end of the helix, that produces the lobule.
Torus marginalis
or Randwulst [Henneberg]. The rounded border inclosing the fossa angularis. It makes its appearance as the hillocks disappear. The latter contribute in part to its formation.
Tragus
The thickened margin of the anterior wall of the concha, situated between the incisura intertragica and the crus helicis. Regarded as a part of the closure mechanism.
Tuberculum anthelicis
[His]. Auricular hillock No. 4.
Tuberculum arterius
[His]. Auricular hillock No. 2, the middle hillock of the mandibular arch.
Tuberculum auriculae
The so-called Darwin's tubercle. Corresponds to the true ear-tip of the long-eared mammals [Schwalbe].
Tuberculum centrale
[His]. Transverse elevation in floor of fossa angularis, separating it into an upper and a lower depression, the lower becoming the auditory meatus. It arises as a bulging of the closure plate of the first gill-cleft.
Tuberculum innominatum
Small cartilaginous anterolateral elevation at junction of horizontal portion of crus helicis with the helix ascendens, i. e., mandibular portion of helix.
Tuberculum intermedius
[His]. Auricular hillock No. 3, the one at the top of the first branchial Cleft.
Tuberculum supratragicum
[His]. Term applied to the accessory elevation that sometimes is found at the upper edge of the tragus. In these cases the tragus may be regarded as two-lobed. The separation of the tragus into two lobes occurs in varying degrees of distinctness.
Tuberculum tragicum
[His]. Auricular hillock No. 1, the lowest hillock of the mandibular arch, giving origin to the tragus.
Topography
In very young embryos (up to 12 mm. long) the branchial area involved in the formation of the external ear constitutes a considerable portion of the ventrolateral surface of the head. Growth in this area is precocious and in advance of the surrounding structures. As the elements of the face and cranium later undergo differentiation, the auricular area becomes relatively smaller, and at the same time it appears to migrate dorsolaterally from near the median line, until it finally occupies its adult site on the lateral surface of the head. The transition in relative size and position of these structures can be traced through figures 9 to 12 (plate 1), in which the auricular area at different stages is shown in blue.
If we start with the primitive branchial arrangement existing in a 6-mm.
embryo, a condition is met with such as is shown in figure 9. Specimens at this
early period, when fixed in formalin, are moderately transparent, and thus it is
difficult to make out their true form. By slightly staining the specimen, as was
done in this case, it is possible to distinguish more clearly the surface modeling
and to represent these structures accurately. In order to display completely the
face region, the greater part of the trunk was removed, leaving only the pericardial
dome and cut end of the aortic trunk.
The drawing which we are considering was made directly from the specimen
and presents a three-quarter view of the four branchial arches of the left side. It
is only by tracing backward from older stages, where the auricular area is pronounced, that one can outline it at this stage. For this purpose actual specimens
were compared, as were also enlarged models in which the branchial region was
completely exposed and in which analogous parts could be identified. When the
same proportionate area is plotted in this way on the mandibular and hyoid arches,
one obtains the result shown in blue in figure 9. Practically the whole surface of
the hyoid arch subsequently takes part in three thickenings, known as the auricular
hillocks numbers 4 to 6. In the same way the greater part of the surface of the
mandibular arch enters into the formation of the first three hillocks. It is of
interest to note how closely the auricular areas of the right and left sides approach
each other in the midventral fine. It is from the small interval between them that
the mandible and its associated soft parts must be derived. It is true, there was
some difficulty in determining the boundary line between the auricular area and
the midventral segment of the mandibular arch, as the line of junction is not characterized by any surface marking, nor can any histological difference be yet recog-
nized in serial sections. The area as outlined, however, agrees in form with that
seen in the next older stage and is probabfy accurate.
When the topography of the auricular region in the stage shown in figure 9
is considered, it can be readily understood that failure on the part of the mandible
to develop would leave the external ears near the median fine in front of the upper
part of the neck. The literature records cases of agnathia or synotia which are of
this nature. Two of these are reproduced in text-figures 3 and 4. In them the
early position of the auricles is retained, owing to the fact that there was nothing to
wedge the two auricles apart, as is normally done by the growing mandible.
In embryos of 8 to 11 mm. the component parts of the mandible have begun
to express themselves, and with their increase in size there is a corresponding
spreading apart of the auricular areas of the two sides, as shown in figure 10. This
drawing was made from a reconstruction model, which, because of the development
of the auricular hillocks, shows very clearly the surface area that enters into the
formation of the external ear. In the 6-mm. embryo we were dealing with a series
of four simple branchial bars; here these bars have partially lost their identity.
The first two have undergone marked development, whereas the third and fourth
have become much less conspicuous. The first or mandibular portion is broken
up into (1) the part that will form the lower jaw and (2) a more lateral part, whose
surface forms the three mandibular auricular hillocks, two of which can be seen
in the figure. These hillocks and those of the hyoid bar have caused deflections in
the first gill-cleft, whose ventral termination will eventually be represented by the intertragal incisure. The surface of the hyoid bar is entirely taken up with its
three auricular hillocks, all of which show in the figure. The small third branchial
bar can be seen partly exposed, but the fourth is entirely covered in.

Figs. 3 and 4. Figure 3 shows (a) ventral and (b) lateral views of an agnathous specimen illustrated in Forster's Atlas of Malformations, 1865 (plate 13, figs. 19 and 20). Figure 4 is copied from a case of cyclopia and agnathia from the Pathological Institute at Heidelberg, described by Schwalbe (1909, p. 615). In both of these cases there is a complete arrest in the development of the greater part of the mandibular arch, with the result that the auricles retain their original median position.
Between the stages of 10 and 14 mm. there is rapid progress in the formation of the face, as can be seen by comparing figures 10 and 11. Figure 11 is drawn from a model to show the details of the face region and the topography of the auricular area, the latter shown in blue. The mouth at this time is fairly well outlined, and one can recognize the region between it and the auricular area which is to form the cheek and jaw. As this region enlarges it will result in the further lateral and dorsal displacement of the auricular area. In the preceding stages the latter still extended downward on the ventral surface of the head, whereas now it is entirely on the lateral surface, and the whole area can be seen in a profile view of the embryo. At this stage the six auricular hillocks show their maximum prominence. The three mandibular hillocks, which at first covered a large part of the mandibular bar, now cover only its caudal margin. The three hyoid hillocks still represent the whole surface of the hyoid bar excepting that part which has been molded into the first cleft. It can be seen in figure 11 that this cleft is much wider than in the younger stages, and we can now speak of a distinct fossa angularis. The ventral third of this fossa becomes relatively deeper to form the external auditory meatus, while the remainder is eventually taken up in the formation of the auricle.
In embryos 16 to 18 mm. long the relations are such as are shown in figure 12, which is a drawing of a model posed similarly to those shown in figures 9 and 11. Owing to the foreshortening in a three-quarter view of this kind, one is apt to get a false impression as to the height of the head; in a true profile view the distance between the eye or ear and the dorsal midline over the midbrain or cerebellum would be much greater. At this time the auricular hillocks, except those continued as the tragus and antitragus, have lost their identity and have been molded into the early form of the definitive parts of the auricle. The beginning helix can be definitely outlined, and less distinctly the crus, the former being entirely a derivative of the hyoid bar, the latter a derivative of the mandibular bar. A fact of interest is that, whereas the crus and tragus form a relatively small part of those adult surface structures that are derived from the mandibular bar, the scapha-helix and antitragus (eventually, also, the anthelix and lobule) constitute the only permanent surface representatives of the hyoid bar.
With the topography of the auricular area thus identified in the four stages just represented, a comparison of these stages discloses certain general facts. Only two gill-bars take any prominent part in the formation of the surface structures of the lower jaw. Of these, the first or mandibular bar contributes by far the greater amount, the second or hyoid bar supplying only a portion of the auricle. The third and fourth bars have no permanent surface record of their existence. The auricular area, relative to the size of the head, covers at first a large surface, but as we pass from simple gill-bars to the stage of hillocks and then to the definite auricle it becomes progressively smaller. Were we to trace it to the stage of 20 to 30 mm., when the face is more fully formed, we would find it still smaller. There- after, the increased growth and spreading character of the free auricle counteract the previous relative decrease in size.
Another and perhaps the most conspicuous feature in the topography of the
developing auricle is its lateral and dorsal migration. In the stage of simple gill-bars the two auricular areas nearly meet in the midventral fine, but, as can be seen in figures 9 to 12, they are gradually crowded sidewise coincidentally with the development of the mandibular apparatus and the structures at the base of the skull. A true profile of figure 12 would show the auricle higher on the side of the head than
it there appears. It is to be remembered that this migration is relative rather than
real. At all stages the mouth line is in a plane roughly intersecting the middle
of the auricle; the appearance of an upward migration is due chiefly to the growth
of the angle of the jaw and the elongation of the neck.
The Branchial Hillocks
In embryos 4 to 6 mm. long the mandibular and hyoid bars are each subdivided by a transverse groove into a dorsal and a ventral part, as can be seen in figures 13 and 14 (plate 2); also figure 9 (plate 1). These are not to be confused with the branchial hillocks. The significance of the subdivision of these bars has never been determined; we shall see, however, that the closure mechanism is derived from the ventral portions, while the articular and sound-collecting mechanisms are derived from the dorsal portions. His (1882), in describing the mandibular bar in young embryos, mentions the existence of a root part (Wurzelstiick) as distinguished from the more ventral portion, which he describes as divided longitudinally into a lip ridge (Lippenwulst) and a mental ridge (Kinnwulst). Careful examination of figures 10 and 11 (plate 1) will show that the ventral part of the mandibular arch is roughly subdivided into two ridges, somewhat as described by His. These ridges do not, however, correspond exactly to the eventual chin and lip, as His first thought. The more anterior one (lip-ridge) in reality gives origin to the greater part of the jaw, the Up being a much later derivative of it. The more posterior ridge (Kinnwulst) corresponds to the soft parts beneath the jaw.
The origin of the branchial hillocks and their fate are shown in figure 5. This figure is intended as a diagrammatic interpretation of figures 13 to 27 (plates 2 and 3). For convenience I have lettered these as a series of successive stages. By comparing them it will be seen that definite hillocks make their appearance in embryos about 10 mm. long, reach their full development in embryos about 14 mm. long, and disappear for the most part between 16 and 18 mm. At stage B, when they first appear (cf. fig. 14), one finds on the dorsal segment of the hyoid bar two opaque elevations corresponding to hillocks 4 and 5. Hillock 4 is strongly suggestive of a facial placode, but on tracing it into the succeeding stages (figs. 15 to 18) it becomes evident that this can not be the explanation. On the ventral segment of the hyoid bar in stage B can be seen an opaque thickening representing the first appearance of hillock 6. At stage C (cf. fig. 15) the three hyoid hillocks are clearly indicated, and at the same time the first indication of hillock 1 can be recognized on the ventral segment of the mandibular bar. The dorsal segment of this bar still forms a round mass corresponding to the Wurzelstiick of His.
At stage D (cf. fig. 16) hillocks 4 and 5 are sharply rounded and have reached their maximum development. Hillock 6 becomes subdivided, as indicated in the diagram, and, as will be seen, it is hillock 6' that eventually forms the antitragus. At this time the dorsal part of the mandibular bar shows the first evidences of hillocks 2 and 3. Along with the appearance of these hillocks the hyoid cleft is widened to form a definite fossa, the fossa angularis of His.
Stage E (cf. fig. 17) represents the hillocks at their maximum development, and it is then appearance at this time that led to the classical description of His and to the numbering of the hillocks serially 1 to 6. Furthermore, it is this appearance that we find duplicated in embryos of other mammals and which resembles also the condition found in buds and reptiles. Microscopic examination of sections through the hillocks at this time shows that they consist of rather sharply outlined masses of condensed mesenchyme cells closely packed against the covering ectoderm. The ectoderm itself is in active proliferation and is much thicker than that of the surrounding regions. In embryos about 11 mm. long the ectoderm can be seen to consist of two layers a more superficial, flattened membrane one cell thick, beneath which is a layer of closely packed cuboidal cells with large round nuclei. It is this deeper layer that appears to be chiefly involved in the process of proliferation. In slightly older specimens it becomes several cells thick, and in some specimens one finds, at the point where it abuts against the mesoderm, a clear white line consisting of the elongated cell-bodies of the proliferating ectoderm. The changes in the ectoderm are most marked over the areas where the condensation of mesenchyme is greatest and more marked over the hyoid bar (hillocks 4 to 6) than over the mandibular bar (hillocks 1, 2, 3). The whole auricular region, however, exhibits this phenomenon and stands out in strong contrast to the adjacent portions of the head. The evidence of activity on the part of the ectoderm of the auricular region is very striking and appears to be closely related to the changes in the subjacent mesenchyme. In appearance it resembles very much the ectoderm of the arm and leg buds in their earlier stages. I shall refer to this subject later.

Fig. 5. A diagrammatic interpretation of figures 13 to 27 (plates 2 and 3), showing the advent and disappearance of the branchial hillocks and the coincident changes in the mandibular and hyoid bars. These stages cover the period of transition from a state of simple branchial bars to the establishment of the primitive auricle. The hillocks are interpreted by the author as foci of more active proliferation of the condensed mesenchymal primordium of the auricle. A to C, embryos 5 to 11 mm.; D to G, embryos 13 to 14 mm.; H to K, embryos 15 to 18 mm.; L to O, embryos 18 to 33 mm. Varying magnifications were adopted so as to bring the structures to about the same size.
The branchial hillocks never reach the same degree of development on the mandibular arch that they do on the hyoid arch, making their appearance later and disappearing earlier. At stage F (cf. fig. 18) they can still be recognized, although they are less distinct than they were in the preceding stage. The specimen selected to illustrate this stage exhibits an anomaly of the fossa angularis, in that a relatively large ridge appears to extend from the region of hillock 2. This is not to be mistaken for the cms helicis; it is due apparently to some peculiarity of this specimen. Hillocks 6 and 6' are very characteristic at this stage. The latter curves inward and forward, forming a different planefrom hillock 6. The two are still, however, partially connected.
At stage G (cf. fig. 19) hillock 3 is beginning to disappear, and hillock 2 is crowded to a more ventral point by the change that has taken place in that region of the mandibular bar, which is preliminary to the formation of the cms helicis. The fossa angularis now forms a rather roomy quadrilateral depression whose floor in this and the next succeeding stages bulges out slightly, corresponding to the development of the tissues in the neighborhood of the head of Meckel's cartilage.
At stage H (cf. fig. 20) hillocks 1, 2, 6, and 6' are still clearly defined. Hillock 5 can still be recognized but is becoming less distinct. Hillocks 3 and 4 can scarcely be outlined, but in their place is a ridge which forms the rounded contour of the upper end of the fossa angularis. The tissue lying under the other hillocks has been constantly increasing in amount, having the effect of increasing the depth of the fossa.
At stage I (cf. fig. 21), coincidentally with the gradual disappearance of the hillocks, the raised margin of the fossa angularis begins to take the form of definitive
parts of the auricle. One can see in the region formerly occupied by hillock 3 that the first evidence of the cms helicis is making its appearance. In the region corresponding to hillock 4 the upper end of the fold in which will form the helix can readily be recognized. The last traces, however, of hillocks 5 and 6 are to be seen. Hillocks 1 and 2 are still quite definite. The relative sizes of hillocks 1 and 2 appear to vary, as does also the degree of separation between them.
At stage J (cf. fig. 22) the conditions are much the same as in the preceding specimen, although, the fold of the helix appears to be a little more pronounced and the last vestige of hillock 5 has disappeared.
In studying these hillocks I find that the angle from which they are viewed and the method of illumination have a great deal to do with their appearance. It has also proved necessary to make considerable allowance for the condition of the tissues and the manner of fixation. In the specimens selected for illustration I have attempted to include only the normal and average ones, but even with this precaution I am conscious of the possibility of having introduced examples that are not necessarily typical. I am somewhat doubtful regarding figure 22, as well as figure 18, the peculiarity of which has already been mentioned. In figure 22 the thick fossa angularis is somewhat exaggerated, as is also the fold of the helix. The embryo is absolutely normal, but the tissues seemed a little shrunken at the time the drawing was made. I may mention at this point that all of these drawings were made directly from the specimens by Mr. J. F. Didusch. In most of them the embryo was stained slightly in order to define more clearly the surface markings.
In stage K (cf. fig. 23) the period of branchial hillocks may be regarded as having passed. The remnants of hillocks 1, 2, and 6' can still be recognized; otherwise, the borders of the fossa angularis are now made up of the sloping surface of the crus helicis and the primitive ear-fold or scapha-helix. Microscopic examination of a transverse section through the ear-fold at this time shows it to be due to a mass of condensed mesenchyme, although differing from the condition found during the hillock period in that there is now a precartilaginous outline of the auricular cartilage, the contours of which can be made out along the posterior edge of the condensed tissue. From the outset this precartilage assumes the typical outlines of the auricular cartilage.
On coming to stage L (cf. fig. 24) we can speak only of remnants of hillocks 1 and 6'. The crus helicis is becoming more distinct and the primitive ear-fold more prominent. With the formation of the crus helicis the fossa angularis loses its identity, and in its stead there is the early form of the concha, divided by the crus into an upper and a lower half.
Stage M (cf. fig. 25) shows a rather marked primitive ear-fold, which is probably a peculiarity of this particular specimen. It may be assumed that any extreme characteristics of the adult ear would have begun to express themselves at this time, and it may be that in this case we would have had an ear with a prominent tip. The tendency toward a pointed process of the ear-fold, however, is an artifact of preservation.
The specimen used to illustrate stage N (cf. fig. 26) is somewhat fuller than the preceding specimen and is more characteristic. The transition from stage N to stage (cf. fig. 27) brings us to a condition that may be regarded as the definitive auricle. We can now recognize the tragus, antitragus, anthelix, scapha-helix, and, distinctly separate from the latter, the crus helicis. In tracing the hillocks up to this point, it is found that the only ones that can be said to persist are hillock 1 (as the tragus) and hillock 6' as the antitragus. All of the others lose then identity in the transition of the tissues forming the margins of the angular fossa into the definitive auricle. Sections through the auricle at this time disclose the fact that the condensed mesenchyme, which heretofore made up these elevations, is now entirely resolved into the cartilaginous plate representing the auricular cartilage and the looser subcutaneous tissues, including the muscles and ligaments of the
Elaboration of the Auricle
On plates 4, 5, and 6 I have arranged a series of photographs showing the auricle at different stages of fetal development. It is thus possible to trace the development of its different parts by following them through these photographs. The increase in the size of the auricle holds only for the individual plate, the photographs on plate 4 being enlarged 10 diameters, those on plate 5 being enlarged 6 diameters, and those on plate 6 being enlarged 4 diameters. In studying them, one should keep in mind the considerable variation which occurs in the form of the ear in adults, for this appears to be expressed in the earliest developmental stages. The photographs, however, are sufficiently numerous to make it possible to separate the constant characteristics from the incidental variations due to normal differences in the auricle and differences in the preservation of the specimens.
The photographs on plate 4 are specimens from the third month of intrauterine life. The first two (figs. 28 and 29) overlap the oldest stage shown on plate 3; most of the parts of the auricle can here be clearly recognized, although they are still very simple in form. Derived from the mandibular bar are the tragus and the crus helicis; as derived from the hyoid bar, one can recognize the anti tragus and the ridge-like primitive ear-fold or scapha-helix. The incisura intertragica, at the entrance of the concha, still bears a resemblance to the hyoid cleft from which it was derived. The concha does not acquire its concave, shell-like character until
later in development, due to the relatively thick and swollen character of the surrounding parts. Figures 30 to 32 differ from the preceding ones only in the increasing prominence of the ear-fold. At this time there is very little surface evidence of the anthelix as distinct from the scapha-helix. However, if sections through this region are examined microscopically, it will be found that the cartilaginous auricle is already characteristically folded into a helix, scapha, anthelix, and concha, the free edge of the helix coming into close contact with the surface of the auricle.
Figure 33 was taken from a slightly different angle and thus exaggerates the
taenia lobularis. In the earlier stages the taenia stands out more prominently.
The lobule forms a free fold between the taenia and the lower end of the helix,
principally at the expense or as an elaboration of the taenia. The latter thus becomes relatively less conspicuous.
In figures 34 and 35 the anthelix makes its appearance on the surface of the
auricle for the first time, and as it does so a groove develops between it and the
free edge of the auricle, representing the early scapha. A lobule can also be recognized as a rounded expansion from the taenia. The small tubercle on the posterior
edge of the helix in figure 35 is due to a thickening of the skin and is to be regarded
only as a peculiarity of this particular specimen. Figures 36 to 38 show a distinct
increase in the size of the auricle. In these there is some differentiation of the
scaphal groove and a corresponding prominence of the helix. The specimen shown
in figure 39 is from a fetus larger than any of the preceding specimens. It falls in
this place because the photographs are arranged in the order of fetal length; the
auricle, however, shows a somewhat retarded degree of differentiation and in form
resembles the specimen illustrated in figure 35. In size it corresponds fairly
closely to its neighbors, and we may perhaps assume that if the fetus had gone on
to term it would have had a simplified type of auricle and possibly a prominent
taenia lobularis or an attached lobule. The method of illumination in making the
photograph shown in figure 40 exaggerates the prominence of the antitragus. I
am introducing it on this account, in order to illustrate the marked differences in
appearance one can secure by a modification of the illumination. The auricle shown in figure 41 is from a fetus from the end of the twelfth week, and aside
from a poorly defined concha it represents most of the elements of the mature
auricle.
On looking back over the auricles illustrated on this plate, one can see that in all of them the mandibular derivatives - the crus and tragus - are relatively large and prominent as compared with the hyoid derivatives. In the further development of the auricle this proportion gradually decreases. It will be further noted that the crus helicis is always a distinctly separate structure from the helix proper; the line of demarcation between them persists in the adult.
The photographs shown on plate 5 represent the changes occurring in the
auricle during the fourth month of intrauterine life. As compared with the photographs on plate 4, the principal change is a relative decrease in the size of the crus
helicis and tragus. Corresponding to this, it is possible to recognize a conchal
cavity which has heretofore been nothing more than a cleft. The concha in the
first two photographs (figures 42 and 43) appears to me a little exaggerated, due,
probably, to the shrinkage of the auricle. Judging from the preceding and succeeding photographs, the average auricle at this time would be somewhat plumper
in appearance. Owing to the fact that these are thinner, one can see for the first
time the presence of the plica principalis.
The specimen shown in figure 44 exhibits the average fulness in the region of
the anthelix, with a tendency to be thrown into transverse ridges. These ridges
occur in this region throughout the fourth and fifth months, depending, apparently,
upon the amount of fulness in the subdermal connective tissue. The helix of this
specimen is characterized by the presence of a moderately well developed tuberculum (Darwinii). In the next specimen (fig. 45) the helix shows a distinct crown
angle (satyr-tip), which doubtless would have persisted in the adult. Although
the concha is still not much more than a cleft, one can make out the presence of a
plica principalis near its upper end. The condition shown in figure 46 is an interesting example of the flat type of auricle with a prominent tuber culum. The
tendency toward obliteration of the helix appears to be due partly to the surplus
tissue in the region of the anthelix, which is thrown into corresponding transverse
folds. In figure 47 the transverse folds are absent and in their stead is a prominent
plica principalis. The auricle shown in figure 48, although of the same size as its
neighbors, is of a more rudimentary type and resembles the specimen in figure 39.
It is probable that both of these would have resulted in small ears had the fetuses
gone on to term.
Figure 49 shows a marked development of the transverse folds in the region
of the anthelix, which were first described by Schwalbe and interpreted by him as
temporary representatives of the longitudinal folds seen in some of the long-eared
mammals. The fact that they are so irregular in occurrence, however, as can be
seen by this and the next plate, makes it doubtful whether these folds can be safely
interpreted as phylogenetic rudiments. I am inclined rather to attribute them to
a redundancy of the soft tissues of the anthelix. This specimen illustrates very
well the difference in character between the auricular derivatives of the mandibular bar (cms helicis and tragus) and the derivatives of the hyoid bar (scapha-helix,
anthelix, antitragus, and lobule). In the last specimen on this plate (fig. 50), in
contrast with figure 49, there are no distinct transverse folds, but the plica principalis is more prominent.
The photographs shown on plate 6 are taken from specimens in the fifth month
of development, with the exception of the last, which has a menstrual age of 23
weeks. Figures 51 and 52 represent flattened types of auricles, such as that depicted
in figure 46 (plate 5). One might think that this was due to flattening of the ear
by handling of the specimen, but such is not the case; the specimens were in good
condition and had not been subjected to any mechanical damage. In my opinion
they can be interpreted only as early exhibitions of a poorly marked helix so commonly seen in the adult. Both of these specimens show a tendency toward a
double tragus. In one the larger segment is above and in the other below. The
specimen shown in figure 53 is similar to the type shown in figure 49 and is characterized by a marked development of the transverse ridges across the anthelix.
In figure 54 the auricle has a well-defined helix at its upper end, together with a
tendency toward a satyr-tip. The lower half of the helix is less well marked.
In this respect it represents a type seen in adults and known as the Cercopithecus
type, as described by Schwalbe (1891). In this ear, as in all the succeeding ones,
the plica principalis can be clearly recognized. Figure 55 shows a very perfect
type of auricle, the one most usually seen, and for the first time we meet with a
well-defined concha, its upper half subdivided by the plica principalis into a superior
and an inferior articular fossa.
The specimen in figure 56 is interesting, in that it still shows the remnants of
transverse folds over the anthelix. The fact that there is a tendency toward
similar folds along the margin of the helix is strongly indicative of their being
nothing more than a temporary expression of the condition of the soft tissues.
The auricular cartilage never takes any part in their formation. The specimens
shown in figures 57 and 58 both have a well-marked helix. In figure 58 the scapha
is somewhat larger and there is a distinct tuberculum.
Figure 59, which closes the series, shows an auricle having all the essential characteristics of the mature ear. In comparing figure 59 with the first figure on this plate it will be seen that in the course of a month the auricle has about doubled in size. This was true also in the two preceding plates. The auricle in figure 59 is of a simple type, having a marked helix only along its upper border. There is now a distinct concha the definite parts of which can be clearly identified. The hair follicles are well developed over the whole of its surface. In comparing this with the auricles shown on plate 4, the marked difference in the relative sizes of the mandibular and hyoid derivatives is very evident.
Summary
In describing the development of the auricle, most investigators have traced its origin to the six branchial hillocks, which make then- appearance at the fifth week as rounded nodules on the mandibular and hyoid bars adjacent to the first branchial cleft. These hillocks, since the early paper of His, have received much attention and have been variously designated, and descriptions have been given of how, by their unequal growth and subsequent coalescence, the eventual auricle comes into existence. The hillocks have been so interpreted in spite of the fact that it was known that they present much the same appearance in mammals that have, in the adult stage, very different types of ears from those of man, and that they are present even in birds and reptiles, which never acquire a distinct auricle.
From what has been stated in the foregoing pages one is forced to the conclusion
that the hillocks, as such, are of a transitory character and are incidental, rather
than fundamental, to the development of the auricle. Probably of more significance, as far as the derivatives of these parts are concerned, is the division of the
mandibular and hyoid bars into ventral and dorsal segments, the closure mechanism
being derived from the former, the articular mechanism and scapha helix from the
latter.
The essential histological change which inaugurates the formation of the
auricle (embryos between 4 and 14 mm.) consists of a proliferation and condensation of the mesenchyme. The mesenchymal change is accompanied by evidences
of marked activity of the ectoderm over the whole auricular area. The deeper
layer of ectoderm cells enlarge, proliferate, pile up two or three cells thick, and at the
same time develop elongated, cylindrical bodies or processes which project toward
the abutting mesenchyme, thus forming a narrow, clear cytoplasmic band at
the mesenchymal junction. Directly beneath the ectoderm the mesenchymal
cells are crowded into a compact line of proliferating elements from which great
numbers of cells can be seen streaming into the deeper levels. The condensation
of the mesenchyme is thus most intense at the ectoderm and gradually becomes
less marked in the looser tissues of the central part of the bar. This phenomenon
of ectodermal and mesodermal activity takes place over the whole surface of the
hyoid bar, and in a less degree over the posterior half of the mandibular bar, in
which the condensed mesenchyme soon becomes localized in the deeper layers.
It is more prominent in those parts where the auricular cartilage arises, and almost
from the first gives the outlines of the cartilage in its primitive form, so that we may
speak of it as the primordium of the auricle. The relation of the branchial hillocks
to the auricular primordium appears to be that they are merely foci in which the
mesenchymal proliferation is temporarily most rapid; they do not represent the
entire auricular primordium. This is particularly evident in hillocks 4 and 5.
In the hillocks of the mandibular bar (1, 2, 3) the mesenchyme is not so compact,
although there also it is in active proliferation.
The proliferation and condensation of the branchial mesenchyme constituting
the primordium of the auricle and the rearrangement of the mesenchyme where
the condensation is less marked produce a change in the surface form of the gill-bars. The narrow hyoid cleft thereby becomes converted into a broad fossa angularis. The width of the fossa is increased by a relative sinking in of those portions
of the bars adjacent to the cleft. This depression is not so much an actual sinking
in as an elevation of the surrounding parts, especially of the auricular rim, made up of those condensed parts of the hyoid and mandibular bars that constitute
the auricular primordium. The widening of the angular fossa can be partly
accounted for by the spreading apart of the auricular rim through the growth of the
tissues composing its floor (closure plate), in which can be seen forming the head of
Meckel's cartilage.
In embryos up to 16 or 18 mm. the condensed mesenchyme forming the primordium of the auricle is fairly uniform in appearance, but at about this time one
can begin to see clearly the auricular cartilage separating itself from the less dense
tissue as a lamina of precartilage cells. As soon as it can be recognized, this lamina
is found to be folded in a manner essentially like that of the adult cartilage. The
scapha-helix stands out prominently, the free edge of the helix remaining in contact
with the ectoderm. The anthelix is also indicated almost from the first, whereas the concha is less sharply defined, and it is not until the embryo has reached a
length of 40 to 50 mm. that the cartilage may be considered to have acquired its
definitive adult form. In this respect, however, it is much in advance of the surface form of the auricle. It is quite evident that the folding of the cartilage is not
produced mechanically by resistance to its expansion on the part of the ectoderm, as
has been maintained; the surrounding tissues are loose enough to make folding
unnecessary. Furthermore, the folding is relatively as great at first, when the
cartilage is small, as it is in the later stages. The auricular cartilage clearly acquires
its form with all the precision and individuality shown by the other cartilaginous
parts of the body.

Fig. 6. and Fig. 7.
Fig. 6. Lateral views of left auricular cartilage, taken from reconstructions of human embryos of the Carnegie Collection: No. 460 (21 mm.), No. 417 (32 mm.), No. 88G (43 mm.). X14.
Fig. 7. Reconstruction of left auricular cartilage of a 50 mm. fetus (No. 84, Carnegie Collection). X 14. A model of the external form of the auricle was made, in conjunction with the cartilage, to give the topographical relations. The edge of the helix in contact with the ectoderm is indicated by cross-lines. Compare with figures 35 and 38.
The transition from the arrangement of typical branchial bars to an auricle of primitive type takes place during the period represented by embryos from 4 to 16 mm. By that time the nodular elevations caused by the hillocks are for the most part smoothed out, and we find the angular fossa inclosed by a rounded border which consists of parts corresponding to the elements of the auricle. The most conspicuous of these are the scapha-helix, tragus, and antitragus. The angular fossa has the form of an elongated depression. Its anterior margin is entirely of mandibular origin and its posterior margin of hyoid origin. The greater part of this posterior margin is taken up with the scapha-helix, or the so-called free ear-fold, at the lower end of which is the modified remnant of hillock 6, which persists as the antitragus. Hillock 1 has become directly converted into the tragus, whereas the crus is slower in making its appearance; not until the embryo has reached a length of 18 or 20 mm. does this structure become evident. It arises from the mandibular tissue in the region formerly occupied by hillocks 2 and 3 and forms an oblique ridge which, enlarging, encroaches upon the angular fossa and converts it into a narrow cleft.
The transformation from the more primitive type of auricle, as just described, into the adult ear may be easily followed in figure 8. This figure is intended as a diagrammatic analysis of the changes illustrated by the photographs on plates 4, 5, and 6. The parts of the auricle derived from the mandibular bar are shown in a lighter tone, while the parts derived from the hyoid bar are stippled. It is interesting to note that the mandibular derivatives are relatively very large in the earlier stages, and this is also true of the derivatives of the lower end of the hyoid bar; in other words, those parts of the auricle concerned with the closure mechanism and the attachment of the auricle to the head are more precocious than the scapha-helix and anthelix. The latter two structures merge directly into each other. Their approximate point of junction, however, is indicated by a dotted line.

Fig. 8. Drawings showing the development of the auricle and its primitive form to the adult type. Those parts derived from the mandibular bar are indicated in lighter tone and are relatively larger in the younger stages; the parts derived from the hyoid bar are stippled; the broken line represents the approximate junction of the anthelix and scapha-helix.
In the younger stages (for example, 85 mm.) the soft tissues of the auricle give the appearance of fulness and tend to be thrown into folds. These should not be confused with the longitudinal folds seen in the adult scapha of some of the long-eared animals. As the cartilage expands, the subcutaneous tissue becomes relatively more scant, particularly in the region of the anthelix and scapha-helix. The histological appearance of the crus helicis is quite different from the hyoid auricle (anthelix and scapha-helix). This difference consists chiefly in the presence of a great number of hair follicles and a considerable amount of subcutaneous fat. These are almost absent in the scapha-helix. The form of the concha, particularly of its articular portion (superior and inferior fossse and plica principalis), does not make itself conspicuous until after the fetus has reached a crown-rump length of 135 mm., although the complete outline of these parts can be recognized in the cartilage in embryos of less than 50 mm.
If one studies a great many specimens, covering the period from 30 mm. to full term, it will be found that there is great variation, just as exists in the adult ear, and that the individuality of the ear is expressed early, as soon as the respective parts can be identified. The tragus may consist of a single lobe or may tend to be subdivided into two lobes; the form of the antitragus varies considerably, and still more does the lobule. The part that varies most, however, is the scapha-helix, particularly as regards its extent and the character of folding of the helix. The least variable is the articular portion, including the crus helicis, the two articular fossae, and the plica principalis.
Bibliography
Baum and Dobees, 1905. Die Entwicklung des ausseren Ohres bei Schwein und Schaf. Anat. Hefte, vol. 28, p. 587-688.
Boas, J. E. V., 1912. Ohrknorpel und ausseres Ohr der Saugetiere. Kopenhagen.
Gbadenigo, G., 1888. Die Formentwickelung der Ohrmuschel, niit Riieksicht auf die Morphologie und Teratologie derselben. Centralbl. f. d. med. Wiss., vol. 26, p. 82-86 and 113-117.
Hammak, J. A., 1902. Studien fiber die Entwicklung des Vorderdarnis und einiger angrenzenden Organe. I. Abtheilung: Allgemeine Morphologie der Schlundspalten beim Menschen. Entwicklung des Mittelohrraumes und des ausseren Gehorganges. Arch. f. mikr. Anat., vol. 59, p. 471-628.
Hennebero, B., 1908. Beitrage zur Entwicklung der Ohrmuschel. Anat. Hefte, vol. 36, p. 107-187.
- 1910. Ueber die Bedeutung der Ohrmuschel. Anat. Hefte, vol. 40, p. 95-147.
His, W., 1882. Auf Stellung von Entwickelungsnormen, zweiter Monat. Anatomie menschlicher Embryonen, Part II, p. 55 and 60-62.
- 1885. Die Formentwickelung des ausseren Ohres. Anatomie menschlicher Embryonen, Part III, p. 211-221.
- 1889. Zur Anatomie des Ohrliippchens. Arch. f. Anat. u. Physiol., Anat. Abth., p. 301-307.
Kastschenko, N., 1887. Das Schicksal der embryonalen Schlundspalten bei Saugethieren. Arch. f. mikr. Anat., vol. 30, p. 1-26.
Keith, A., 1906. The results of an anthropological investigation of the external ear. Proc. Anat. and Anthrol. Soc. Univ. Aberdeen, 1904-1906, p. 217-239.
Mall FP. On stages in the development of human embryos from 2 to 25 mm long. (1914) Anat. Anz., 46: 78-84.
Mall, F. P., 1914. On stages in the development of human embryos from 2 to 25 mm. long. Anat. Anz., vol. 46.
Moldenhauer, W., 1877. Die Entwicklung des mittleren und des ausseren Ohres. Morph. Jahrb., vol. 3, p. 106-151.
Munch, F. E., 1897. Ueber die Entwicklung des Knorpels des ausseren Ohres. Morph. Arbeiten, vol. 7, p. 583-610.
Ruge, G., 1898. Das Knorpelskelett des ausseren OhreB der Monotremen. Morph. Jahrb., vol. 25, p. 202-223.
Schaeffer, O., 1892-93. Ueber fbtale Ohrentwicklung, die Haufigkeit fotalen Ohrformen bei Erwachsenen und die Erblichkeitsverhaltnisse derselben. Arch. f. Anthropol., vol. 21, p. 77-132; also p. 215-245.
Schmidt, Joh., 1902. Vergleichend-anatomische Untersuchungen uber die Ohrmuschel verschiedener Saugetiere. Berlin.
Schwalbe, G., 1889. Das Darwin'sche Spitzohr beim menschlichen Embryo. Anat. Anz., vol. 4, p. 176-189.
- 1891a. Beitrage zur Anthropologic des Ohres. Internat. Beitr. z. wiss. Medicin. Virchow Festschr., vol. 1, p. 95-144.
- 1891b. Ueber Auricularhbcker bei Reptilien; ein Beitrag zur Phylogenie des ausseren Ohres. Anat. Anz., vol. 6, p. 43-53.
- 1897. Das aussere Ohr. Handb. d. Anat. d. Menschen, Herausg. K. von Bardeleben, vol. 5, part 2, pp. 125-131.
- 1916. Beitrage zur Kenntnis des ausseren Ohres der Primaten. Ztschr. f. Morphol. u. Anthropol.. vol. 19, p. 545-668.
Sera, G. L., 1917. E la forma dell' orecehio umano antica o recente? Giornale per la morfologia dell' Uomo e dei Primati, vol. 1, p. 109-125.
Spaulding, M. H., 1921. Development of the external genitalia in the human embryo. Contributions to Embryology, vol. 13, p. 69-88. Carnegie Inst. Wash. Pub. 276.
Tataroff, D., 1887. Ueber die Muskeln der Ohrmuschel und einige Besonderheiten des Ohrknorpels. Arch. f. Anat. u. Physiol., Anat. Abth.
Description of Plates
- In-text Figures: Figure 1 and 2 | Figure 3 and 4 | Figure 5 | Figure 6 and 7 | Figure 8 | Text | Glossary
- Plates: Plate 1 | Plate 2 | Plate 3 | Plate 4 | Plate 5 | Plate 6 | Plates | Glossary
- Figures: 1. Auricle cartilage | 2. External ear | 3. Agnathia | 4. Agnathia+cyclopia | 6. Auricular cartilage embryo 21, 32 and 43 mm | 7. Auricular cartilage 50 mm fetus | 9. Embryo 6 mm | 10. Embryo 12 mm | 11. Embryo 14 mm | 12. Embryo 18 mm | 13. Embryo 1380, 5 mm | 14. Embryo 1767, 11 mm | 15. Embryo 1461, 10 mm | 16. Embryo 562, 13 mm | 17. Embryo 1232, 14 mm | 18. Embryo 475, 15 mm | 19. Embryo 899, 13 mm | 20. Embryo 434, 15 mm | 21. Embryo 492, 16.8 mm | 22. Embryo 576, 17 mm | 23. Embryo 547, 18 mm | 24. Embryo 955, 17 mm | 25. Embryo 1584, 18 mm | 26. Embryo 1134e, 21.3 mm | 27. Embryo 1358b, 33.2 mm | 28. Embryo 1535, 28 mm | 29. Embryo 2163, 36 mm | 30. Embryo 1980, 37 mm | 31. Embryo 1840a, 38.5 mm | 32. Embryo 2075, 40 mm | 33. Embryo 2144, 45.5 mm | 34. Embryo 642, 49 mm | 35. Embryo 2170, 50 mm | 36. Embryo 2095, 52 mm | 37. Embryo 2095, 52 mm | 38. Embryo 2066, 53 mm | 39. Embryo 2079, 56.5 mm. | 40. Embryo 1561, 57 mm | 41. Embryo 218, 62.5 mm. (R.) | 42. Embryo 1724, 66.2 mm | 43. Embryo 2328, 65 mm | 44. Embryo 2118, 69 mm | 45. Embryo 981, 85 mm | 46. Embryo 1845, 87 mm | 47. Embryo 1449, 87.3 mm | 48. Embryo 2003, 103.5 mm | 49. Embryo 1858, 100 mm | 50. Embryo 2274, 113 mm | 51. Embryo 2185, 113.5 mm. | 52. Embryo 9526, 114 mm. | 53. Embryo 1811, 114 mm | 54. Embryo 1716, 119 mm. Fig. 59. 1742, 191.2 mm | 55. Embryo 19576, 119 mm. | 56. Embryo 1782, 135.6 mm | 57. Embryo 1702, 150 mm | 58. Embryo 1708, 154 mm | 59. Embryo 1742, 191.2 mm | Figures
- Related Notes: Outer Ear Development | Carnegie Contributions to Embryology
Reference
Streeter GL. Development of the auricle in the human embryo. (1922) Carnegie Instn. Wash. Publ. 277, Contrib. Embryol., 14: 111-138.
Cite this page: Hill, M.A. (2024, April 28) Embryology Book - Contributions to Embryology Carnegie Institution No.69. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Book_-_Contributions_to_Embryology_Carnegie_Institution_No.69
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G
Plate 1

Ventrolateral view of the head in a series of human embryos, showing the change in topography of the auricle in the course of its development. The surface area of the mandibular and hyoid bars entering into the formation of the auricular primordium is colored blue. These figures show the lateral and dorsal migration of the auricle coincident with the formation of the mandible.
Fig. 9. Drawing made directly from an embryo 6 mm long, No. 1787 Carnegie Collection. X 22. The olfactory disk and the lens of the eye are outlined by dots.
Fig. 10. Reconstruction model of an embryo 12 mm long, No. 1121 Carnegie Collection. X 15.
Fig. 11. Reconstruction model of an embryo 14 mm long, No. 940 Carnegie Collection. X 15. Here the parts belonging to the jaw are clearly separated from what are to be the soft parts of the upper neck by a groove, which might be called the mental groove.
Fig. 12. Reconstruction model of an embryo 18 mm long, No. 1390 Carnegie Collection. X 12.3.
- Streeter (1922) Plate 1
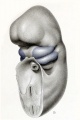
Fig. 9. Embryo No. 1787 6 mm
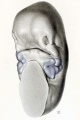
Fig. 10. Embryo No. 1121 12 mm
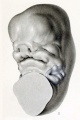
Fig. 11. Embryo No. 940 14 mm long
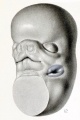
Fig. 12. Embryo No. 1390 18 mm




Plate 2

Drawings of human embryos, showing the region of the first branchial cleft and its transformation into a fossa angularis. Coincident with this transformation the mesenchyme of the hyoid and mandibular bars undergoes proliferation and becomes condensed to form the primordium of the auricle. Foci of more active proliferation show on the surface as branchial hillocks. Specimens are from the Carnegie Collection.
| Fig. 13. No. 1380, 5 mm. long. X 34. | Fig. 14. No. 1767, 11 mm. long. X 24. |
| Fig. 15. No. 1461, 10 mm. long. X 20. | Fig. 16. No. 562, 13 mm. long. X 20. |
| Fig. 17. No. 1232, 14 mm. long. X 17. | Fig. 18. No. 475, 15 mm. long. X 17. |
- Streeter (1922) Plate 2
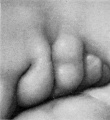
Fig. 13. Embryo No. 1380 5 mm
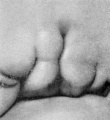
Fig. 14. Embryo No. 1767 11 mm
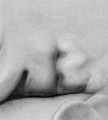
Fig. 15. Embryo No. 1461 10 mm
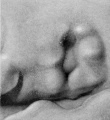
Fig. 16. Embryo No. 562 13 mm
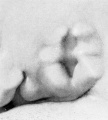
Fig. 17. Embryo No. 1232 14 mm
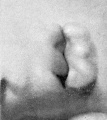
Fig. 18. Embryo No. 475 15 mm






</gallery>
Plate 3

Drawings of human embryos, in series with the preceding plate, and showing the disappearance of the branchial hillocks and the completion of the auricle in its primary form. Specimens are from the Carnegie Collection.
| Fig. 19. No. 899, 13 mm. long. X 24. | Fig. 20. No. Template:CE434, 15 mm. long. X 27. | Fig. 21. No. 492, 16.8 mm. long. X 27. |
| Fig. 22. No. 576, 17 mm. long. X 17. | Fig. 23. No. Template:CE547, 18 mm. long. X 22. | Fig. 24. No. 955, 17 mm. long. X 24. |
| Fig. 25. No. 1584, 18 mm. long. X 24. | Fig. 26. No. 1134E, 21.3 mm. long. X 24. | Fig. 27. No. Template:CE1358b, 33.2 mm. long. X 24. |
- Streeter (1922) Plate 3
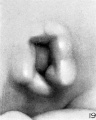
Fig. 19. Embryo No. 899 13 mm. long. X 24.
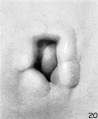
Fig. 20. Embryo No. Template:CE434 15 mm. long. X 27.
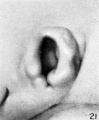
Fig. 21. Embryo No. 492 16.8 mm. long. X 27.
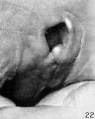
Fig. 22. Embryo No. 576 17 mm. long. X 17.
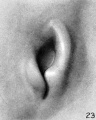
Fig. 23. Embryo No. Template:CE547 18 mm. long. X 22.
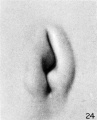
Fig. 24. Embryo No. 955 17 mm. long. X 24.
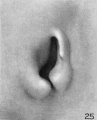
Fig. 25. Embryo No. 1584 18 mm. long. X 24.
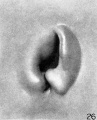
Fig. 26. Embryo No.1134e, 21.3 mm long. X 24.
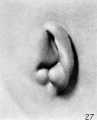
Fig. 27. Embryo No. 1358b, 33.2 mm long. X 24.









Plate 4

Photographs of the auricle of the human fetus during the third month, all being taken at an enlargement of 10 diameters. In some cases the right ear was selected and reversed for convenience in comparison. These are indicated by the letter R. All specimens are from the Carnegie Collection, and length given is crown-rump.
- Streeter (1922) Plate 4
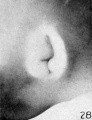
Fig. 28. No. 1535 28 mm
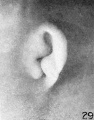
Fig. 29. No. 2163 36 mm
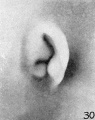
Fig. 30. No. 1980 37 mm
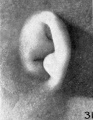
Fig. 31. No. 1840a 38.5 mm (R)
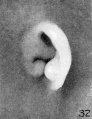
Fig. 32. No. 2075 40 mm (R)
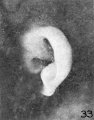
Fig. 33. No. 2144 45.5 mm (R)
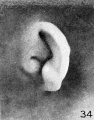
Fig. 34. No. 642 49 mm
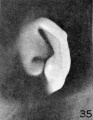
Fig. 35. No. 2170 50 mm
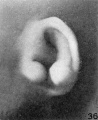
Fig. 36. No. 2095 52 mm (R)
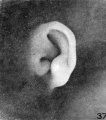
Fig. 37. No. 2095 52 mm
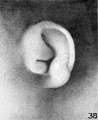
Fig. 38. No. 2066 53 mm (R)
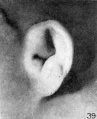
Fig. 39. No. 2079 56.5 mm
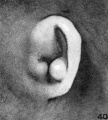
Fig. 40. No. 1561 57 mm
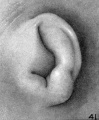
Fig. 41. No. 218 62.5 mm (R)














Plate 5

Photographs showing changes occurring in the auricle of the human fetus during the fourth month. In some cases the right ear was selected and reversed for convenience in comparison. These are indicated by the letter R. All the photographs are taken at an enlargement of 6 diameters. Specimens are from the Carnegie Collection, and length given is crown-rump.
- Streeter (1922) Plate 5
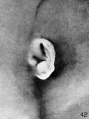
Fig. 42. No. 1724 66.2 mm
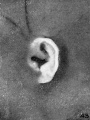
Fig. 43. No. 2328 65 mm
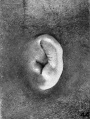
Fig. 44. No. 2118 69 mm
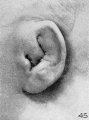
Fig. 45. No. 981 85 mm
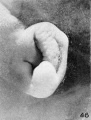
Fig. 46. No. 1845 87 mm
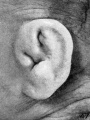
Fig. 47. No. 1449 87.3 mm (R.)
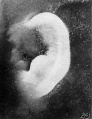
Fig. 48. No. 2003 103.5 mm
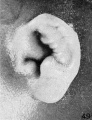
Fig. 49. No. 1858 100 mm (R.)
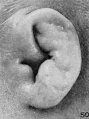
Fig. 50. No. 2274113 mm (R.)









Plate 6

Photographs showing the form of the human auricle during the fifth month of intrauterine life, with the exception of specimen shown in figure 59, which has a menstrual age of 23 weeks. The photographs are all shown at an enlargement of 4 diameters. Specimens are from the Carnegie Collection, and length given is crown-rump.
- Streeter (1922) Plate 6
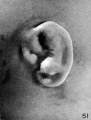
Fig. 51. Embryo No. 2185 113.5 mm
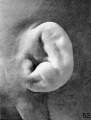
Fig. 52. Embryo No. 9526 114 mm
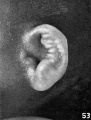
Fig. 53. Embryo No. 1811 114 mm
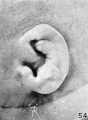
Fig. 54. Embryo No. 1716 119 mm
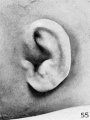
Fig. 55. Embryo No. Template:CE19576 119 mm
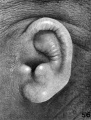
Fig. 56. Embryo No. 1782 135.6 mm
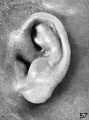
Fig. 57. Embryo No. 1702 150 mm
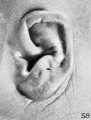
Fig. 58. Embryo No. 1708 154 mm

Fig. 59. Embryo No. 1742 191.2 mm









Figures
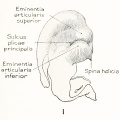
Fig. 1.
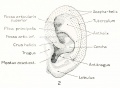
Fig. 2.
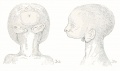
Fig. 3. Agnathia
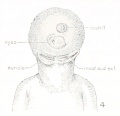
Fig. 4. Agnathia+cyclopia
Fig. 6. Auricular cartilage embryo 21, 32 and 43 mm
Fig. 7. Auricular cartilage 50 mm fetus
Fig. 9. Embryo drawing 6 mm
Fig. 10. Embryo model 12 mm
Fig. 11. Embryo model 14 mm
Fig. 12. Embryo model 18 mm
Fig. 13. Embryo No. 1380, 5 mm
Fig. 14. Embryo No. 1767, 11 mm
Fig. 15. Embryo No. 1461, 10 mm
Fig. 16. Embryo No. 562, 13 mm
Fig. 17. Embryo No. 1232, 14 mm
Fig. 18. Embryo No. 475, 15 mm
Fig. 19. Embryo No. 899, 13 mm
Fig. 20. Embryo No.434, 15 mm
Fig. 21. Embryo No.492, 16.8 mm
Fig. 22. Embryo No. 576, 17 mm
Fig. 23. Embryo No.547, 18 mm
Fig. 24. Embryo No. 955, 17 mm
Fig. 25. Embryo No. 1584, 18 mm
Fig. 26. Embryo No.1134e, 21.3 mm
Fig. 27. Embryo No. 1358b, 33.2 mm
Fig. 28. Embryo No. 1535, 28 mm.
Fig. 29. Embryo No. 2163, 36 mm.
Fig. 30. Embryo No. 1980, 37 mm.
Fig. 31. Embryo No. 1840a, 38.5 mm. (R.)
Fig. 32. Embryo No. 2075, 40 mm. (R.)
Fig. 33. Embryo No. 2144, 45.5 mm. (R.)
Fig. 34. Embryo No. 642, 49 mm.
Fig. 35. Embryo No. 2170, 50 mm.
Fig. 36. Embryo No. 2095, 52 mm. (R.)
Fig. 37. Embryo No. 2095, 52 mm.
Fig. 38. Embryo No. 2066, 53 mm. (R.)
Fig. 39. Embryo No. 2079, 56.5 mm.
Fig. 40. Embryo No. 1561, 57 mm.
Fig. 41. Embryo No. 218, 62.5 mm. (R.)
Fig. 42. Embryo No. 1724, 66.2 mm.
Fig. 43. Embryo No. 2328, 65 mm.
Fig. 44. Embryo No. 2118, 69 mm.
Fig. 45. Embryo No. 981, 85 mm.
Fig. 46. Embryo No. 1845, 87 mm.
Fig. 47. Embryo No. 1449, 87.3 mm. (R.)
Fig. 48. Embryo No. 2003, 103.5 mm.
Fig. 49. Embryo No. 1858, 100 mm. (R.)
Fig. 50. Embryo No. 2274, 113 mm. (R.)
Fig. 51. Embryo No. 2185, 113.5 mm.
Fig. 52. Embryo No. 9526, 114 mm.
Fig. 53. Embryo No. 1811, 114 mm.
Fig. 54. Embryo No. 1716, 119 mm. Fig. 59. No. 1742, 191.2 mm.
Fig. 55. Embryo No. 19576, 119 mm.
Fig. 56. Embryo No. 1782, 135.6 mm.
Fig. 57. Embryo No. 1702, 150 mm.
Fig. 58. Embryo No. 1708, 154 mm.
Fig. 59. Embryo No. 1742, 191.2 mm.






{kind=link}
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2024, April 28) Embryology Book - Contributions to Embryology Carnegie Institution No.69. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Book_-_Contributions_to_Embryology_Carnegie_Institution_No.69
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G