Paper - On the factors concerned in causing rotation of the intestine in man
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Frazer JE. and Robbins RH. On the factors concerned in causing rotation of the intestine in man. (1915) J Anat. 50(1): 75-110. PMID 17233053
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
On the Factors Concerned in Causing Rotation of the Intestine in Man

{kind=link}
By J. Ernest Frazer, F.R.C.S.,
St Mary’s Hospital, Professor of Anatomy in the University of London; and R. H. Robbins, M.D. Cantab., Senior Demonstrator of Anatomy in the Medical School of St Mary’s Hospital.
Introduction
It has been recognised for a very long time that the disposition of the intestinal canal in the adult human subject is the result of a process of rotation which it undergoes in the course of embryonic development, the tube being changed from an early condition, in which it is described as being straight and median in position, into one in which the distal part is said to be rotated round the proximal portion in a direction from left to right figures illustrating the nature and extent of this rotation are common in all text-books of anatomy, and variations in the phenomenon are constantly assumed to account for cases showing abnormal disposition of the intestinal tube in some small or large part of its course. But one does not meet with much success in an endeavour to obtain a clear and detailed notion of the nature of the rotation and of the extent of gut taking part in it. This is particularly the case when light is sought on the origin and cause of the movement; in fact, we have not found any satisfactory or coherent account of the process from this point of View in the books we have consulted.
In the absence of reliable information, we have endeavoured to work the question out for ourselves, and to build up a reasonable and connected theory of cause and effect in association with the rotation of the bowel. In the nature of things, any explanation of the changes which occur must be theoretical, but we have, we hope, been successful in formulating a coherent hypothesis which is in accordance with the facts that we have directly observed. It is founded on the examination, by the microscope and by reconstructions, of embryos from the fourth week to the third month, and by dissection of specimens from this time up to birth: we have made some eighteen models,[1] large and small, as well as other reconstructions, and propose to give in this paper the results of our study of these and of our other specimens.
An example of the intestine placed in the median sagittal plane may be taken from the embryo shown in fig. 1. The embryo is one of 5 mm. in which a window has been cut in the left abdominal wall to expose the intestinal tube: this, disposed in what is practically the middle line of the cavity, forms a nearly right—angled V, the apex of which is continuous with the vitelline duct. The vitelline vein lies in the cavity in front of the proximal limb of the V, running up to join the sub-hepatic anastomosis above, and down to reach the vitelline duct externally. It is worth noting that the hinder limb of the V is continuously curved, corresponding with the umbilical arteries, on and between which it lies.

Fig. 1. 5 mm embryo, simplified from models, showing belly cavity through window in wall. Course of vitelline duct indicated by dotted lines on further side of cut; body stalk. Shows gut lying practically in median sagittal plane.
A great change must take place in the disposition of the intestines if the adult state is to be attained from the simple conditions seen in this embryo. We think it is convenient to divide this process of development into three stages: the first of these extends to the period of return of the intestines to the abdominal cavity; the second begins with this return and lasts till the caecum comes into relation with the dorsal wall of the abdomen; while the third goes on from this until some time after birth. Each of these stages has its special characters: the first stage is essentially that of an umbilical loop with two limbs lying beside one another, the second is the stage of rotation, in which the planes of the adult condition are reached ; and the third exhibits the extension in those planes by which the various parts of the tube attain their usual individual position.

Fig. 2. 8 mm embryo, simplified from model. Seen from the ventral side, and rather from the left. V, continuous with the apex of the loop, contains not only the epithelial remains of the vitelline duct, but also the vitelline artery, and the vein which joins it just be ond the apex of the loop. The “ umbilical cord” passed to the right of the tail this, with the oblique view-point, gives the appearance of the umbilical loop being directed to the right.
First Stage
fig. 2, from an embryo of 8 mm., exhibits the essential points of the first stage. There is a loop consisting of two limbs, proximal and distal, lying in the umbilical sac; the sac is not shown, having been removed with the front wall of the belly. The proximal limb is placed to the right of the middle line, and, in the sac, to the right of the distal limb.
The presence of the loop in the umbilical sac seems to be the natural result of its growth in a cavity too small or too much taken up to contain it: the apex of the V in fig. 1 is held in the umbilical opening by the attachment to it of the vitelline duct, and it is only to be expected that it will be guided, so to speak, by this duct into the opening when the elongation of the loop occurs.
In a specimen of 7.5 mm., in which the proximal limb of the loop is not turned over and depressed to the same degree as in the embryo from which fig. 2 was drawn, but lies more on the level of the rounded ventral surface of the mesentery, the concavity of the limb is occupied by the continuation of the left umbilical vein on the visceral aspect of the liver, with which is a small prolongation from the right lobe of that organ. These structures that occupy the concavity are along the upper edge of the umbilical opening. In the 8—mm. embryo, and to a greater degree in those of about 9 and 10 mm., the right lobe has very much increased in size and the limb’ is correspondingly depressed, while the rounded mesenteric surface, concave from above down, is in contact with the liver tissue surrounding the venous channel. On the other hand, the umbilical vein in the 5—mm. specimen passes up mainly to the left of the comparatively small liver, but anastomoses on the caudal aspect of the organ with the vitelline system: the specimen is not in good enough condition to permit closer examination of the nature of the vitelline anastomoses. As a result of the study of the models of these embryos we suggest that the turning over of the proximal limb of the loop to the right is directly due to its close relation to the vein and liver. When the secondary left umbilical vein is formed as the result of the vitello—umbilical junction, the new venous channel lies on the visceral surface of the liver and is carried with this across the ventral aspect of the proximal limb of the loop of gut: the vein runs up from the left side of the umbilical opening to the right lobe of the liver, and as this descends it carries down the upper part of the vein, swinging it, as it were, on its lower umbilical end, with the effect of turning the proximal limb over to the right. The schemes in fig. 3 may perhaps make this plainer.

Fig. 3. Schemes to illustrate the effect of the vitello-umbilical anastomosis and descent of the liver on the proximal limb of the loop, represented as on frontal section. In the first figure the left umbilical vein passes up mainly to the left of the liver but is also sending offshoots below it which join the vitelline system. In the second figure this anastomosis forms now the main left umbilical drainage, and is carried down on the visceral aspect of the liver. the hepatic end of the vein swinging in the direction of the arrow. The result is that the proximal limb is folded over to the right.
The loop in an early specimen, such as in fig. 2, is a freely movable tongue-shaped projection, directed ventrally from a fixed base.‘ The mesentery of the loop is continuous with the median mesentery attached to the dorsal abdominal wall, but in addition to this the two limbs of the gut, where they become continuous with the upper and lower parts of the rest of the tube, are relatively fixed by thick areas of mesentery. These can now be considered under duodenal and colic headings respectively.
(a) The Duodenal fixation
If the proximal loop is looked at from the right, as diagrammatically represented in fig. 4, it is seen to form a curved loop, concave ventrally, placed beside the median mesentery. At its cranial end a thick pad is seen on the dorsal surface of this end of the limb, standing out from the general level of the median mesentery. The portion of the gut which is fastened to the dorsal wall by this conglomeration of tissue is the future duodenum, and the pad itself is the dorsal duodenal mesentery or mesoduodenum : its substance is continuous on the left with that of the general mesentery, extends up to the foramen of Winslow, and presents below a rounded blunt falciform edge, deep to which a shallow recess lies between it and the median mesentery.
If a frontal section were to be made, running cranio-caudally through the mesentery a little ventral to its dorsal attachment and passing through the foramen of Winslow above (along the interrupted line in fig. 4), the mesenteric structures, seen from the dorsal side, would present somewhat the aspect which is schematically shown in fig. 5. In the first figure the line of the median mesentery is seen to be interrupted above by the opening into the lesser sac, and just below this the thick mass of the mesoduodenum is apparent, standing out to the right and carrying the duodenum on its ventral surface.
These figures are only schemes, but they are founded on the reconstructions of the parts: the thick hook-like arrangement of the mesoduodenum is a striking feature in sections through the parts, and the. hollow of the hook, the little inter—mesenteric recess, is to be found throughout the greater part of the first stage. It is in this mesoduodenum that the pancreatic outgrowths occur and enlarge, the upper one passing below the foramen of Winslow and so into the lower or right wall of the small sac, while the lower outgrowth enlarges rapidly in the mesoduodenum itself.

Fig. 4. Diagram of proximal limb of loop seen from the right. It shows the duodenaumbilical loop turned over to the right and depressed, so that it lies beside the median mesentery. The ventral surface of the limb and the mesentery is in contact with liver, which should occupy the space (black) between them and the belly wall. The interrupted line gives the imaginary line of section for the schemes in Hg. 5. Cf. fig. 6, where part of the limb is cut away.
These are indicated in the schemes. The formation of the duodenal curve has nothing to do directly with the occurrence of rotation in the intestinal loop, but, as the lower end of the duodenum affords a fixed point of attachment for the proximal limb of the loop, a short account of its production, as it presents itself to us, may not be out of place.
In the first scheme in fig. 5 the duodenum is seen on the further or ventral side of the mesoduodenum, in which the head of the pancreas is indicated; the second drawing shows that this head has increased in size and has curved out the intestinal tube, a process which has progressed still further in the third figure, but the duodenum throtoghout is held by its mcsoduoolemom. This account denotes shortly the manner in which we believe the form of the duodenum to be attained— curved out to the right by the growth of the head of the pancreas, and remaining fixed throughout to its mesoduodenal base. The earlier growth of the head of the pancreas appears to be mainly in its upper part, so that the proximal portion of the axopfiagus duodenum is raised and curved, then the middle and lower parts enlarge and the corresponding portions of the tube are bowed out (see figs. 10 and 11}. It follows from this conception of the duodenum that the muscle of Treitz is formed in the lower part of the mesoduodenum : its place is indicated in the schemes, but we have not been able to discover undoubted evidence of its presence before the 35-mm. stage, when the duodenal curve is practically complete. It may be that the method of staining of our specimens makes more diflicult the recognition of the band before this time, or perhaps its definite formation and recognition may be associated with the increasing strain thrown on this part of the gut with the later downward growth of the head of the pancreas.

Fig. 5. Schemes to illustrate the formation of the curve of the duodenum The mesentery is supposed to be viewed from the dorsum, having been cut away from its dorsal attachment. The dorsal mesoduodenum is seen as a thick mass projecting to the right immediately below the protrusion to the left of the small sac. The alimentary tube is shown by interrupted lines running down in the front wall of the small sac, turning to the right, and becoming continuous with the duodenum which passes down in front of the mesoduodenum. The head (H) and body (B) of pancreas grow into the mesoduodenum: observe that B can pass almost directly into the wall of the small sac. The other figures show how, while B extends into this wall, H grows in the mesoduodenum and by its enlargement curves out the duodenum round it: thus the duodenum is attached all the time to the mesoduodenum. X is the position of Treitz’ band, put in its hypothetical place in the first two figures.
We have no doubt that the duodenal formation comes about in the way outlined above. But we are not so certain about the terminal piece of this part of the bowel, whether there might not be some secondary adhesion present here at a later stage. For various reasons we are inclined against this view, but the question is of little importance relative to our main object, so we do not propose to consider it further.
The formation of the duodenal curve proceeds throughout the first stage, progressing more rapidly in the latter part of the stage, like the other modifications that occur in the simple conditions of the 8-mm. embryo. The mode of formation, in relation with the visceral surface of the liver, leads to the rectification of the originally oblique position (fig. 11) of the duodenum, and it comes into a more nearly frontal plane behind this organ, while its distal part must be directed inwards and must therefore form a sharp duodeno—je_junal curve forward close against the right side of the median mesentery (see fig. 10)
It is convenient here to distinguish between that part of the proximal limb of the intestinal loop which extends in the abdomen from the duodenum to the umbilical opening, and that part which is in the umbilical sac : the first can be referred to as the duodeno—umbilical loop and the other as the umbilical part of the proximal limb.
The superior mesenteric artery passes downwards and forwards in the mesentery to the left of the middle of the mesoduodenum to gain the mesentery of the loop, and the band of Treitz is continuous with the condensation which surrounds the arterial stem.
(b) The Colic fixation
The fixation of the base of the distal limb of the loop is simple, but has not, we believe, been hitherto described. The continuous curve of this limb, as seen in fig. 2, is no longer visible in fig. 3 Where the colon is seen to be bent at a sharp angle which we may term the “colic angle”: this angle, of course, marks the junction of the “ midgut ” with the “ hindgut ” in the older descriptions. The appearance of this angle suggests that it is produced by the attachment to the point of the angle of a band or other structure which holds it up, but allows the loop to turn freely forward, and examination of the specimens reveals at once the existence of such a band.
Its position can be understood from fig. 6, where the structures are viewed from the right: the greater part of the duodeno—umbilical loop is cut away, exposing the median mesentery and the colic angle, while the cut edge of the mesentery is clearly visible. The position of the band, which we term the “retention band," is indicated by the interrupted lines: the direction of the lines does not imply that any such direction of disposal is to he made out, for in structure it is at this stage, of course, merely a band of much condensed mesenchyme. Traced towards the colic angle, the greater and thickest part of the band leads to the point of the angle, but a thinner part passes behind the angle to become continuous with the thick mesocolon of the “ hindgut,” and another thinner part runs in front of the angle to the distal limb and reaches the dilation just in front of the angle which marks the situation of the future caecum. Traced in the other direction, the band joins the concentration round the mesenterie vessels into which the band of Treitz has already been stated to pass. A section through the situation A, just above the angle, shows the band as a marked thickening in the median Inesentery, while the mesenteric Vessels are turned over with the duodeno-umbilical loop and lie on its right side: a section further back, at B, shows that the band is no longer in the mesocolic area but is now merged in the thickened mesenchyme round the superior mesenteric vessels, in the axis of the common mesentery, where, though it may be supposed to he as shown in the figure, it is not distinctly marked off‘.

Fig. 6. Diagram modified and simplified from models. 7.5 mm. Duodeno-umbilical loop cut away largely, exposing the median mesentery, on which the position of the retention hand is marked b interrupted lines. (If. fig. 4. A, B, lines of sections a and b, which, lhowever, are trace directly from sections through the model. In these sections the retention band is stippled.
As stated above, the band presents on examination only the evidence of condensed mesenchyme in the early stages, with one or two minute vessels among the cells ; in later stages nerves are distinctly visible with the vessels, and, as the band gets smaller, actually or proportionately, the vessels and nerves become more marked and the condensation has the appearance of being of secondary importance to them. The obliquity of the band and its connexion with the band of Treitz high up suggest the possibility that they may he remains of a more extensive structure which had some connexion with the more cranially-placed roof of the widely open intestinal sac of early stages. Discussion of such an interesting question would be outside our province, even if we possessed material young enough and in sufficiently good preservation to enable us to speak from direct acquaintance with the subject: as it is, we have no desire to make any suggestion concerning its origin nor any wider statement about its distribution or possibilities.
But although we have no views to advance about the morphological position of the retention band, concerning its practical effect on the colon of the human embryo we do not think there can be any doubt. The constant presence of the colic angle is enough, in our opinion, to justify the presumption that traction is being exercised on that part of the gut, and when this thick band is found in the exact situation that would be suggested by a consideration of the angle, it seems to us that no further evidence is required to permit us to describe the band as exercising such traction. But when we use the word “traction” we do not mean to imply an active approximation of the colic angle to the region of the duodenum : measurements on the models show us that the distance between them remains the same nearly to the end of the first stage, so there is no actual approximation. But the relative approximation is very great, as it is hardly necessary to point out. The band holds the piece of gut to which it is attached in the same place while the body caudal to it is growing, and for this reason we have termed it the retention band.
The value of the retention band is evident when we consider the great growth of the post-umbilical segment of the body which occurs in the first stage. In this growth the colic angle is held in place by the band, and consequently the “hindgut” of the older descriptions must be drawn out between the angle and cloaca. This is an interesting reflection when considered with von Berenberg-Gossler’s hypothesis, founded on the examination of a very rare and striking teratological specimen, that not only the rectum, but also the colon, caecum, and terminal part of the ileum are derived from the cloacal walls. However this may be, we think there is no doubt that the band, by holding the colic angle, relatively approximates the angle and duodenum and at the same time draws out the gut distal to the angle so that its length at any time is in correspondence with that of the postumbilical part of the belly.
We can now appreciate the condition of the complete intestinal and mesenteric complex—-from our point of view it may be simply pictured as in the scheme in fig. 7. In the first of these schemes the whole system is represented as flattened out, and it is. possible to divide descriptively the mesentery into that of the loop and that in the abdomen; or, since the basal part of the loop remains in the abdomen, into that of the loop and that of the remainder of the gut: this last part can be simply called the median mesentery, The median mesentery has a part below the retention band and a part above the band of Treitz: the upper part is continuous on its right side with the mesoduodenum, and need not concern us further, but the lower part, which has to do with the appearance of rotation of the bowels, can be referred to when necessary as the median mesocolon.
In the loop the arterial axis can be taken as dividing the area of mesentery into a proximal mesentery and a distal or mesocolon of the loop. The retention band, then, can be described as separating the median mesocolon from that of the loop.

Fig. 7. Analytical schemes of the mesentery. A, at the beginning of the first stage. B, toward the end of this stage. Observe that the narrow strip (mesocolon) between the arterial axis and the distal limb remains unaffected, whereas the mesentery of the proximal limb widens very considerably with the growth in length of the limb.
So far, the loop with which we have dealt is characterised by the more or less equal growth of its parts, so that its limbs are about of the same length and the depth of mesentery between them and the arterial axis is nearly equal on both sides, as appears in the scheme. But, as the first stage goes on, modifications appear in these details, although the essential features of the stage—the double-limbed loop, with the depressed proximal limb to the right of the distal—remain until the return of the bowel to the abdomen. The end-products of these modifications chiefly concern us from our present standpoint, and we will deal with these later in some detail, but it can be pointed out now that, so far as the loop is concerned, the modification is in the direction of great and disproportionate growth of the proximal limb and of its mesentery, so that coils are formed to the right of the distal limb, which remains relatively very short and with only a narrow strip of mesocolon between it and the artery. Such differences might be expressed as in fig. 7, B.
Within the abdomen increase takes place in cranio-caudal length and dorso-ventral depth of the median mesocolon commensurate with the growth of the hinder part of the embryo. The growth is most marked in the latter half of the second and beginning of third month, and the result is as shown in fig. 8, where the median mesocolon a little time before the intestinal return is viewed from the left side. The abdominal colon lies in relation with the ventral wall, and the mesocolon extends as a median septum in the cavity between this and the dorsal wall.

Fig. 8. Sagittal section to left of middle line of abdomen in embryo of 35 mm. Reconstruction. Seen from the left. The coils are left in sitd in the opened umbilical sac. The abdominal colon and mesocolon form a median septum extending dorso-ventrally and up to the liver and attachment of omental bursa above. The small figures are tracings from sections through the model showing the increase in size of the umbilical colon in the caecal region. The sections pass through the colon at about the levels indicated by the arrows: the dotted surfaces are those of the cut intestine, which include part of the ileum in one section, while the position of attachment to mesocolon is indicated by dotted lines.
The less extensive area of mesocolon in a 28-mm. embryo is shown in fig. 9, and its small size in one of 22 mm. can be estimated from fig. 11, where, however, the abdomen is viewed from the right. A 26-mm. model shows an intermediate stage, so that the rapid increase may be said to begin between the 22-min. and 26-mm. stages. The thin area of mesocolon between the retention band and the thick pelvic mesentery (seeifig. 7) appears to be enlarged and drawn o11t to supply the increased surface: this assumption seems to be justified by an examination of the distribution of the vessels of the part, indicated in fig. 8. The ascending branch of the inferior n1esenteric elongates with the growth, showing that the corresponding length of gut is drawn out, the situation of the colic angle being marked by a branch of the superior mesenteric which joins the other vessel here.
The colic angle is found in the intestinal tube up to a fairly late stage, becoming gradually less well marked. This is in accordance with what occurs in the parts of the retention band. The originally thickest part of this band, that which went to the colic angle, becomes smaller and presumably weaker as the mesocolon grows, about the end of the second month, and in the stage represented in fig. 8 it is only to be found as a slight thickening beside the branch of the artery which runs to the region of the angle. The portion of the band which passed dorsal to the angle to reach the thick pelvic mesocolon becomes attenuated in a similar way, and the only part which presents any appearance of strength is that part wl1icl1 is shown in fig. 7 passing to the caecal region: this seems to become more developed as growth proceeds, and is drawn out along the growing distal limb of the loop, being well marked in the 26-mm. specimen and easily recognised in that of 35 mm. as a fairly thick but small band in the mesocolon between the mesenteric vessels and the colon, as far as the caecum.

Fig. 9. Model of 28-mm. embryo, seen from the left side. Stomach and bursa omentalis have been cut away in part, exposing the median mesocolon : when in position they reach down as far as the genital structures below the suprarenal body. The vitelline vein has also been cut away.
In fig. 8 the stomach and bursa omentalis have been cut away at their attachments to the median structures. They are suspended to the left side of the mesocolon, and are shown partially cut away in fig. 9: when in position in this model they reach the floor of the false pelvis. By the end of the first stage they have become relatively smaller ailil are well overlapped by the liver in front and below, this organ coming into relation with the colon.
The duodeno—umbilical loop is placed on the right side of the mesocolon, between it and the right lobe of the liver. This loop increases in length also, forming a curve the lowest part of which rests on the pelvic brim, apparently depressed, as it grows, by the liver. The deep and sharply cut markings on the liver are enough to show that this organ exercises a certain amount of pressure on the structures with which it comes into contact, and retains them in position.

Fig. 10. View of the abdominal and umbilical contents from the right. 35-mm. embryo. Observe that the duodeno-umbilical loop is sharply kinked just beyond the duodenum to form the duodeno-jejunal flexure, the course of the gut being indicated by interrupted lines. The rounded elevation of the head of the pancreas is seen in front of the duodenal curve and below the cut vitelline vein. The mesoduodenum is visible behind the duodenum and below the foramen of Winslow. Cf. fig. 8.
The condition present on the right side toward the end of the first stage is seen in fig. 10. In this figure the duodenal curve is represented round the head of the pancreas, which is visible as a slight rounded prominence within the curve. Just distal to the duodenum the duodeno-jejunal flexure is to be found: its position is evident in the figure, and the course of the hidden piece of gut is indicated by the interrupted lines. The bend lies against the median mesocolon, between it and the plane of the mesentery of the proximal limb of the loop, and in the concavity made by the continuity between these two (see fig. 15). The position, on the right, of the plane of the mesentery of the proximal limb is probably the result of the pancreatic growth : this portion of the mesentery of the loop is continued at its base on to the front of the duodenum and the head of the pancreas, and it is carried somewhat to the right by them as they grow, thus affording an opportunity to the flexure to turn up under it, between it and the median mesocolon. The flexure itself may also come into existence as an indirect result of the pancreatic growth with its consequent bending in of the distal end of the duodenum. This, with the comparative narrowness of the mesentery at the base of the loop (fig. 7, B), would lead to an upward and inward turning of the next succeeding part of the gut.

Fig. 11. View from right of abdominal contents of 22-mm. embryo, the liver being removed to the middle line. Modified from model. The interrupted line shows the position of the duodeno-jejunal bend lying against the median mesocolon. Structures are cut short at the umbilical opening. Head of pancreas is not apparent.
A corollary to this position of the flexure would be the placing on its right, potentially, of the mesenteric vessels, lying as they do in the upper part of the mesentery of the proximal limb.
The flexure is apparent in the 22-mm. stage (fig. 11) and is also seen in the 20-mm. specimen. In the 28-mm. embryo it is represented by a slight kink in the intestine below the duodenum.
Between the flexure and the umbilical opening the abdominal part of the proximal limb is curved beside the mesocolon. All these structures, simple modifications of the duodeno-umbilical loop, lie, with their mesentery, between the right lobe of the liver and the median mesentery.
Summing up, then; the conditions which hold in the abdomen at the end of the first stage, we may say that there is a median septum, movable on its dorsal attachment and made by the median mesocolon and colon, placed between the omental bursa and stomach on the left and duodenoumbilical coils on the right, and that it is separated by these structures from the left and right lobes of the liver respectively. These conditions are all the natural and inevitable results of growth occurring in the positions assumed at the beginning of the stage (see fig. 2), and there is no alteration in the essential character of the intra-abdominal conditions seen at that period.

Fig. 12. Schemes to show (A) the position of the structures below and between the lobes of the liver before the return of the bowel, and (B) the alteration caused by the return. The median septum, which is displaced to the left in B, is the median mesocolon, the colon being supposed removed with the front part by the section, which is transverse. The abdominal conditions are represented schematically in fig. 12, A.
Second Stage
The second stage, that of the return to the abdomen, is also that in which rotation occurs. The rotation is the effect of the return on the loop itself and on the contents of the abdominal cavity into which it passes: the intra-abdominal conditions have already been described, but it remains to examine the state of the umbilical loop and the mode and cause of its return before we consider the result attained by this phenomenon.
To take the last of these first. Various suggestions have been made as to the responsible factor which may be at work in the reduction of the herniated intestine. Without entering into a discussion of these, we may say at once that our observations lead us decidedly to favour the view that the intestines are, to use Mall’s term, “sucked back ” into the abdomen: we can find no evidence of any traction directly exercised on the loop or its mesentery, nor any reason to suppose that there can be any indirect traction brought to bear on it.
The process of being “sucked back” is, of course, one in which, as a result of relatively greater external pressure, the contents of the sac are really pushed back into the abdomen. This accords with what we know of the conditions inside that cavity. Jackson has shown that after the 31—mm. stage the liver decreases in proportionate bulk, and, as the abdominal cavity is growing larger-—perhaps disproportionately——it follows that there must be a fall in intra—abdominal tension: the effect of this fall on the amniotic pressure need not be regarded practically, so that the pressure exercised by the amniotic fluid on the umbilical sac which it surrounds must be relatively increased. In our view, there must come a time when the increasing pressure will force the intestine out of the sac into the belly, and that this does not occur pawl passil with the fall in internal pressure may be due to resistance offered by the greater bulk of the structures in the sac when compared with that in the passage: it seems very probable that such enlargement would hinder the return, but when once the resistance is overcome by the relatively increasing pressure of the sac, there is nothing further to interfere with the movement, which goes on rapidly to its termination.
Such appears to us to be a reasonable view to take of the circumstances attending the reduction of the extruded gut, but in adopting such a theory there are certain points which present themselves for consideration, and about which a few remarks may not be out of place. firstly, there is the question of the meaning of reduction of liver mass. Judging from our own specimens, we think it must be very difficult to base accurate estimations of the actual size of the liver on microscopic sections and reconstructions made from them. In our experience, however well prepared and apparently fresh the embryo may have been, there is yet always more or less retraction of the organ. For example, in the embryo from which the model shown in fig. 9 was made, though the preservation and histological condition appear to he very good, yet if one were to judge the state present from the actual sections, the liver did not occupy the lower part of the abdominal cavity at all. But the model of the liver shows at once that this would be a most fallacious view, for it carries on it the sharply cut markings of the structures with which it was in contact, and these can be read on it with absolute certainty: they show conclusively that the organ was in contact with the Wolffian remains and gubernaculum where these lie in the extreme caudal floor of the cavity, or false pelvis, although no indication of such relation could be gathered from the sections directly. The model gives one an idea of the extent to which the liver has contracted, and the question arises whether, seeing how much or how little this contraction may be in any individual liver, it is possible to compare and estimate the bulk of livers of various stages with more than approximate accuracy.
We would suggest, from the study of our models, that more trustworthy figures relating to the actual size of the liver might be obtained from casts of the general abdominal cavity, but we have not ourselves taken up this investigation. By the expression “actual size” we mean the presumed size in the living embryo.
But, whatever may prove to be the result of further research in this matter, we suppose that there can be no doubt that the relative decrease of the liver can be expressed in other terms as “decrease in the rate of growth,” and it is this decrease in growth that leads to the fall in intra-abdominal pressure. But to say that the pressure falls in the belly is not to say that a space occurs there. No doubt some of the fall in pressure is met by the partial collapse of the belly wall, which is also under amniotic pressure, and in this way a potential space is provided which, when necessary, can be distended by the returned bowel: reconstruction favours the idea that the lower part of the wall is retracted before the return of the intestine, but comparison with later conditions is difficult, as allowance cannot be made for individual differences in the specimens. Even if we allow this possibility, however, there is also some change in the liver to be accounted for: it fills the spare cavity before the return, as shown by the reconstruction of the liver in the 35-mm. specimen, but after the return it is separated from the false pelvis by the coils of intestine and has acquired a different set of markings. A possible explanation of this and allied points has occurred to us as a result of consideration of liver reconstructions.
It is plain, from the nature of the markings, that the retraction of the organ about which we have already spoken as occurring in sectioned specimens has taken place at the time of employment of the hardening and fixing agents, so that there can be no question of absolute decrease of liver substance, and the difference between such a liver and its bulk before the retraction took place must admit practically of expression in terms of loss of fluid. If this is so, the liver before retraction, the living organ filling the cavity, contains more fluid, e.g. blood, than afterwards: in other words, it fills the cavity, although its rate of growth may be decreasing, by virtue of holding more fluid in its meshes, and in this way has a potential power of retraction which comes into play \vhenever circumstances allow it to retract by taking up some of the space it occupies and permitting it to discharge some of this fluid from its bulk. Thus it will be seen that, although we are doubtful of the value of reconstructions for showing the actual size and form of the living organ, yet they may be taken as perhaps expressing more accurately the potential difierences in bulk of liver substance than would such models if they were to show the actual size only.
A fruitful comparison, then, can be made between the retracted liver of the first stage and that of the second stage. In the first, the retraction is effected not by loss or atrophy of liver substance but by loss of fluid from the whole mass, and in the second the arrival of the intestines in the abdomen allows the fluid to leave the organ, so that it retracts. Thus in some ways and with evident reservations the liver might be compared with the lung in the pleural cavity—it is in a condition of undue expansion, and, as soon as an opportunity is given to it, it retracts towards its base of attachment. In this connexion it is interesting to note that the opening out of the liver under reagents, and the general direction of its visceral surface, is similar to that observed in the organ after the return of the bowel to the abdomen.
We bring forward these suggestions as purely hypothetical explanations of the points raised above, for with our material we have not been able to devise any means of putting it definitely to the proof.
Having provided hypothetically for the factors causing return and for accommodation for the intestines in the abdomen, it is now necessary to inquire into the act of return itself. As is admitted, the act is practically sudden and complete, and we have offered above a more or less satisfactory reason for the sudden and ‘complete nature of the action following the commencement of the return.
It is not possible, we think, for the umbilical coils to return en masse: the shape and size of the umbilical opening forbid this, and when we remember that the edge of the central notch in the liver corresponds with the upper and side margins of the opening, with which it is in close contact, and that the abdominal recti are also in the immediate neighbourhood, the sense of impossibility becomes a positive certainty. If, then, the coils do not return en masse, there must be a movement of each limb into the abdomen in continuity, each moving with its corresponding mesentery, and it becomes necessary to decide whether these limbs return together or one before the other. Considerations of the conditions found in the sac lead us to believe that the distal limb returns after the proximal one.
In the umbilical sac the colon is seen (fig. 8) to be placed along the left side of the collection of coils, in a more or less straight line. It is a narrow tube when it enters the sac, but just before it reaches the caecal region it increases markedly in diameter, and this enlargement is still further increased by the ileum which runs into it at an acute angle: the outlines of the gut in the figure do not show this difference very well, but the tracings from sections of the model at the levels indicated give a better notion of the increase in calibre. If, now, We think of the conditions present in the sac when the external pressure is moving the contents into the abdomen, it would seem certain that this shape of the colon must operate against its passage through the narrow neck of the sac. For the neck must be completely filled by a mass of mesentery and gut, pressed together in its narrow confines, and under such circumstances the caecal enlargement must be retained in the sac. It does not appear to us that we can escape from this conclusion as a result of a theoretical consideration of conditions inside the sac.
We have not been able, at the moment, to obtain a fresh specimen at the proper stage of development on which we could verify this conclusion experimentally, and, so far as we know, only one case (Mall’s) has been reported in which the intestinal loop has been found in a state of partial reduction: in this case, while the proximal limb of the loop was in the abdomen, the caecum still lay in the umbilical sac. Here, therefore, we have an experiment which bears out the view we have advocated and, so far as it goes, confirms us in that view.
We may call attention here to another observation we have made which might be perhaps of some importance in this connexion. A system of large intercommunicating venous sinuses exists in the mesentery close to the caecum, becoming noticeable toward the end of the second month and apparently increasing in size after this. It is conceivable that venous return might be hindered at the neck of the sac when the mesentery is engaged in it, that these sinuses would therefore be distended, and that in this way further resistance to the passage of the caecum through the neck would arise. This possibility has presented itself to us, but we do not desire to lay any emphasis on it, for we think that the retention of the caecum is a necessary consequence of facts which are open to observation and there is no need to call in to its aid a factor which rests on an imagined condition.
We have not investigated the conditions in other mammals, so cannot speak about any corresponding influences in them: we hope to be able to do so on a future occasion.
Holding the view, then, that the caecum is retained to the last in the sac, we must assume that the proximal limb of the loop returns in successive lengths, slipping back rapidly with its mesentery into the belly. We can now see that the condition of the mesentery shown in fig. 7, B, fits in with the scheme of return. The mesentery of the proximal limb is deep enough to permit its gut to lie in the abdomen though the distal limb remains in the sac. On the other hand, the narrowness of the strip of umbilical mesocolon which exists between the colon and the arterial axis entails the necessity of the mesenteric vessels also remaining in the sac With the colon, for practically they can only move with the distal limb.
We can now imagine the mechanism in motion under the conditions we have described. The proximal limb and its mesentery slip back into the abdomen, the more proximal part first and so on, and thus find themselves entering that cavity to the right of the median mesocolon, which we have described as forming a median septum with its abdominal colon. This septum extends above to the central notch in the .liver, out of which the colon passes forward to reach the umbilical sac. The returning coils, entering below the right lobe of the liver, will fill the lower part of the abdomen: in so doing they push the median septum to the left and dorsally, swinging it back on its dorsal attachment, so that the coils pass ventral to it and thus come to lie below the left lobe of the liver, though separated from it in part by the stomach and omental bursa. The general idea of these changes is illustrated in the scheme in fig. 12, B. At the same time, as suggested above, there becomes possible a certain amount of retraction of the liver, with raising and opening out of its visceral surface, so that the coils can gather below it. The extension of coils to the left takes place in front of the displaced median septum, comes into relation with the bursa omentalis in front of this portion of the colon, and in front of the bursa lies below the left lobe of the liver directly: it also reaches this lobe beyond the colon and bursa and stomach.

Fig. 13. Schemes to show how the returning proximal limb of the loop passes below the umbilical colon and mesenteric vessels. Supposed to be transverse sections looked at from above.
Moreover, in passing to the left, the coils must necesmrily go below the level of the ventrally olirected distal limb of the loop and of the vnesenteric vessels, as these run forward from the notch of the liver to the umbilical opening: these structures can be described, then, as passing forward to the opening above the mass of coils and their mesentery. This is shown in ii . 13.
When the colon leaves the sac and enters the cavity of the abdomen, it must lie, therefore, with the main vessels on the top of the mass of coils and their mesentery. In considering this matter it seemed to us that the comparative shortness of the umbilical colon—i.c. the distance between the abdominal end of the distal limb and the cmcum—compared with the length of the mesentery of the coils of small intestine would bring the caecum into position on the coils, not reaching as far as their ventral limits but lying between them and the liver. We were therefore interested in observing the conditions in specimens shortly after the return of the bowel, and our theoretical expectations were borne out by what was found in these. fig. 14 gives illustrations of some of these specimens, with descriptions of them, and the differences between them appear to us simply to indicate that there is a certain amount of chance or individual variation in the immediate effect on the returning colon of the influences under which it comes when it enters the abdomen. The walls of the colon are thicker than those of the small intestine at this stage, and its lumen smaller, so that it would resist bending or kinking with greater power than the small bowel, as a thick rubber tubing tends more to assume a straight direction than thin tubing. So one would expect that the umbilical colon, when it enters the abdomen, would naturally tend to come into line with the part of the abdominal colon with which it is directly continuous, i.e., it would be disposed to turn toward the right, as can be understood from fig. '13. But such a movement would be hampered by its entanglement between the liver and the intestinal coils, and to us the differences in the specimens in fig. 14: are only individual differences in the arrest of the movement toward the right.

Fig. 14. Two specimens of 45 mm. , Intestines exposed by raising the liver carefully and depressing the pelvic parts: in A the liver is represented as divided transversely, and the drawing is somewhat diagrammatic. In A the ca:cum lay as shown in a curved state on the coils a. little to the right of the middle, but in B it was more to the right, though still delinitely resting on the coils. St. is the stomach. The inesocolon of the loop is seen in A but not in B. The overliaiiging projecting part of the omental bursa has been raised to show the coils, but the left transverse colon is too deep to be shown in this way. Altogetlier live specimens of this period were examined. Of the others, one ofubout 40 mm. was like A in its caecal relations, another of 42 mm. resembled B, but in one of 39 mm. the caecuin lay to the right of the coils. in this case the whole mass of coils seemed to he carried more than is usual to the left, so that the caecum was not much beyond the level of the duodenum although it lay to the right of them, and the right lobe of the liver was perhaps larger than usual.
The immediate result of the return of the colon, then, is that it lies on the coils of small intestine, with the caecum wedged in from behind between these coils and the liver. There does not appear to be any reason to suppose that peristaltic movements take place at this time in the intestine, but a cursory observation of the abdomen shows at once that there is at any rate a rapid increase in mass of these coils, and the tendency of the mass as a whole would be to keep at the full length of its mesentery_ Under these circumstances, it seems to us that there would be a constant inclination to press the caecum back from between the growing coils and the solid liver, so that it and the colon would come to lie where there is more freedom from pressure, namely, over the smaller bulk of the neck of the mass. Such a position is, of course, transverse to the long axis of the mesenteric “neck” of the mass of coils: the disposition would be a consequence of the backward move of the caecum from between the coils and the liver, for as it is pressed back the colon behind it naturally comes into line with the transversely directed piece of the abdominal colon immediately succeeding it, the process going on until the originally umbilical colon is placed from left to right across the root of the mesentery of the loop, with the caecum pointing to the right. In this way we explain the change from a position above the coils, with a more or less dorso-ventral direction, to one transversely placed across the mesentery behind the coils.
In the 55-mm. embryo the caecum is in contact with the dorsal wall to the right of the mesentery. It is not in any way attached to the wall, in fact it hardly touches it, but it is behind the plane of the coils of small intestine, and the attainment of this position can be taken as marking the end of the second stage in the evolution of the human type.
Glancing back at the description we have given of the progress of the change, as we conceive it, in this stage, we may say that the fall in intraabdominal tension leads to the return of the intestine, that this takes place quickly but not en masse, that the positions of the descending colon and left part of the transverse colon in their relations to the small intestine are consequences of the first part of the return, and that the position in proper plane of the remainder of the colon is a delayed consequence of the last part of the return. That is to say, that the rotated state, even though the final positions are not yet reached, has been attained in its essentials during this stage, as a result of the return of the bowel from the umbilical sac. It is fitting, therefore, that we should inquire at this stage into the nature and extent of this rotation.
It has already been said that the rotation occurs in the loop and does not involve the duodenal region nor that of the median abdominal colon, so that the descending and left portion of the transverse colon are not concerned in the twist, although they appear to be included in it. The diagrams in fig. 15 may be of assistance in following out the process. In the first diagram the conditions of the earlier part of the first stage are represented, the loop having been cut away. In B the conditions at the end of the first stage are shown: the duodenal curve has appeared, widening out the attachment of the mesentery of the loop toward the right, so that the duodeno-jejunal bend is seen turned up to the left of the axis of the mesentery of the loop, while the stomach lies to the left of the median colon and mesocolon but separated from them by the thin-walled bursa omentalis. C is the same as B, but the proximal limb of the loop is left in position with its mesentery, only the distal limb being supposed to have been removed with its mesocolon: it is evident that when the proximal limb returns it must do so to the right of the median mesocolon. The result of this return on the mesocolon is seen in D : the colon and the mesocolon are pushed to the left and backwards, so that they would have the coils of small intestine in front of them. If the position of the structures at the bases of the limbs of the loop is noted, it call be seen that these do not change their relations to each other during the second stage 2 in B the proximal limb is cut through beyond the duodeno-jejunal flexure, whereas in D the division goes through that part; but if this is allowed for, the relative position of the bases of the two limbs is seen to be practically the same after as before the return of the intestine. The lower end of the duodenum is below the colic angle: this is partly the persistence of the essential conditions of the first stage, and partly the result of depression due to the pancreatic growth, but no effect seems to be produced on its level by the return or rotation of the loop. The section of the colon is supposed to be made in the region of the colic angle, and this is seen to remain in position although the gut distal to it is turned backwards and to the left.
As a matter of fact, the region of the earlier angle does not remain absolutely central in position, but is carried a little to the left of the middle line; but, as the end of the duodenum and the duodeno-‘jejunal bend are carried with it in that direction, the relative positions of all these structures are practically unchanged. The movement of this part of the colon towards the left is probably to be explained as a part of the swinging of the abdominal colon to that side (as can be understood from fig. 13), and may be made possible by return of part of the umbilical colon from the sac: there is nothing to hinder the return of the unenlarged colon from the sac, only the caecum being held back as a result of its size.
Thus it becomes apparent that there is no tendency to rotate on the part of the colic angle: whatever movement takes place is not in the direction of rotation round the loop, but is in the other direction toward the left. This being so, there can be no movement of rotation affecting the gut distal to the angle, and it can be seen in D (fig. 15) that there is no actual twist of this portion of intestine, it being simply laid back from its median attachment while the coils pass in front of it.
When the proximal limb enters the abdomen, the upper part of the small intestine returns first. It is to be expected that this part, continuous with the duodeno—jejunal flexure, will lie against the median mesocolon, and the next succeeding part as it enters the abdomen will lie ventral to the first part and rather to its right. So, in a general way, we might say that the coils which lie deepest in the abdomen and more to the left, pushed there by those which enter subsequently, might be expected to be those that return first, i.e. the upper part of the bowel. This agrees with general experience, and calls for no further study. These proximal coils, as they pass to the left, carry with them their mesentery and vessels, and twist these below the main vessels which we have seen (fig. 13) must remain in the umbilical sac: this can be taken as an indication of rotation of this part of the loop. But those coils which lie to the right of the middle line do not exhibit any indication of rotation, using the relation of their vessels to the main Vascular stem as the test of its occurrence. So also when the caecum reaches the abdomen and lies on the coils, those coils which are on its right obtain their blood from the arterial stem on their left, and, judged by this standard, have not yet “rotated.” The mesocolon containing the main vessels can be seen in fig. 14, A, lying with the colon on the coils: it was not visible in specimen B without disturbing the coils.
When the caecum passes to the right of the mesenteric neck of the mass of coils, however, the fundamental adult arrangement of right and left branches of the main vessels is attained, and the rotation is complete. Looking at it in this way, and judging the rotation from its effect on the branches of the mesenteric vessels, the process can be said to commence with the first passage of coils to the left, and to end when the caecum reaches the dorsal Wall—in other words, the rotation continues throughout the second stage of our description.
A complete twist of this sort, through half a circle, involves the whole of the free loop. The base, as We have seen, is fixed, and just where the movable joins the fixed part the amount of rotation is not so great, but it rapidly passes from the state of the original to that of the acquired condition.
Examination of the vessels indicates the area of rotation very clearly. The models make plain that the relations between artery and vein in the fixed region of duodenum, dorsal to the junction of vitelline and superior mesenteric veins, are practically the same in the 8-mm. embryo as in the adult body, but the relation between the vessels in the loop is reversed during the second stage. Thus the rotation, judged by its results, is confined to the loop’ and occurs only during the second stage.
A rotation of such sort is conveniently spoken of as occurring round the arterial axis, but it must not be assumed that the main vessel actually forms an axis round which the loop twists. If the account we have given has been properly understood, it will be evident that, although the first stage can be schematically represented with an arterial axis and the completed second stage shows the reversed branches round this axis, yet the intervening rotation takes place without reference to any fixed axis. When the first coils go to the left they pass below the artery because it is temporarily fixed above them, but they do so without turning on it and as a result of their surrounding relations: in the last part of rotation the caecum is carried to the right and brings the artery with it, so that the “ axis ” is really swung to the right on the remaining coils and not these to the left on the artery as an axis. The point is perhaps one of minor importance, but it is necessary to understand it if one wishes to have a clear comprehension of the process of rotation.
Before proceeding to the third stage in the evolution of intestinal position, it may be as well to deal with certain structures which we have mentioned in the foregoing account of the two first stages. The retention band is shown in fig. 6 as comprising a main part extending to the angle, and two subsidiary parts going to the caecum and pelvic mesocolon respectively. The main part, as already stated, diminishes in thickness toward the end of the second month and, at the time of return and rotation, is quite an insignificant thickening in the mesocolon which would only be noticed if looked for in the proper situation. We have not been able to find any certain representative of it in the full—term foetus. The descending subsidiary part also thins away as the median colon elongates, and is present only as a slight condensation along the ascending branch of the inferior mesenteric artery in the second stage. The caecal prolongation from the band, however, becomes better marked and thicker as the first stage proceeds, so that it is present as a strong and prominent band in the 26-min. embryo and is well marked, though not so thick, in the 35-mm. specimen. We have no means of ascertaining what relation, if any, there may be between the presence of this band and the comparative slowness in the growth of the distal limb of the loop, nor have we made any investigation into its ultimate fate: we have not so far found definite evidences of its existence at birth.[2]
The colic angle depends for its presence on the colic attachment of the retention band, and, presumably in consequence of the atrophy of this band, the angle is less marked toward the end of the second month and is practically non-existent when the return of the bowel is due. Its position, however, can be easily recognised from the fact that the left branch of the middle colic artery reaches the gut here, or, for practical purposes, it can be placed on the gut opposite the jejuno-mesocolic fold, which represents nearly enough the narrow part of the neck of the mesentery of the loop. Generally speaking, therefore, it may be said that the descending colon and less than the left half of the transverse colon are derived from the median abdominal colon, distal to the colic angle, while the remainder of the transverse and the ascending colon come from the distal limb of the loop.
The vitelline duct loses its connexion with the intestine at an early stage—in one of our embryos, however, labelled as 12.5 mm., the duct was still continuous with the intestinal wall, although no definite epithelial strand of connexion could be made out. Keibel and Elze have reported the connexion in an embryo of about the same size, and Thyng in one a little larger.
The vitelline vein is to be found as a small channel subsequent to the intestinal return; we cannot say definitely when it finally disappears.
The vitelline artery was present on the right side of the loop of intestine in all our specimens which were examined on this point. It was still to be found connected with the mesentery, but in a very attenuated state, in the 28-mm. embryo, but not later than this.
The bursa omentalis lies as a thin-walled sac behind the stomach, and between it and the median mesentery. It is only fixed by its “neck,” on the left side of the common dorsal mesentery, opposite the opening (foramen of Winslow), and lies free below this with the stomach developing, so to speak, in its front wall (see figs. 5 and 8). In the first half of the second month it begins to project ventrally between the stomach and median colon, and by the end of this month it forms a definite irregularly folded projection in this situation, as may be seen in the model in fig. 9, and in the diagrams in fig. 15. This hollow projection of the excessively thinWalled sac is, of course, the early indication of the great omentum, and it lies in close contact with the abdominal colon at the end of the first stage (fig. 15, C) but is not attached to it in any way. When the coils enter the abdomen the median colon is turned to the left as in fig. 15, D, and not only raises the bursa and the stomach, but is also pushed back below the bursa so that the great omentum comes to lie on its ventral face to some extent, and here is in contact with the coils of small intestine. The small intestine seems to be responsible for raising the bursa and stomach both indirectly through the colon and directly by its own mass as it gathers on the left side of the abdomen. The future left half of transverse colon and splenic flexure are now invaginated to some extent into the omental bursa from below, but there is not, at the end of the second stage, any adhesion between the structures. The further changes in this region belong to the third stage of our description.

Fig. 15. Four diagrams to show the positions of the bases of the limbs of the loop before and after the return of the bowel. The abdominal colon is shown as a. median septum with its mesocolon, deflected to the left (in D) after the return. The colon is supposed to be divided at about the region of the colic angle, and the proximal limb in front of the duodeno-jejunal bend in A and B, and through the bend in D. Observe how the pancreatic growth carries the base of attachment of the mesentery of the loop out to the right, allowing room for the flexure in its lower concavity, and making possible the relation of the vessels to the transverse part of the duodenum.
The great omentum comes into evidence along the whole length of the bursa and thus extends to the right as far as the first part of the duodenum : when the colon of the loop ends the second stage by swinging across the neck of the mass of coils, its distal part comes into a relation with this right extremity of the bursal projection which is comparable with that of the abdominal gut with which it is continuous (see fig. 16). At the end of the second stage the omental projection covers the front of the left part of the transverse colon and the upper left coils, and is not quite disposed as in fig. 14, where it has been raised to exhibit the coils.
Third Stage
The third stage is one of extension and fixation of the colon in the plane it has reached at the end of the preceding stage. Thus it is not really a stage concerned in the actual rotation of the loop and does not call for a very detailed description here.
When the caecum comes into relation with the dorsal wall it touches it at or about—i.e. just above—the crest of the ilium, and just below the lower end of the kidney. As already stated, we found this position attained in the 55-mm. specimen, but it seems to us highly probable, when the individual variations found are taken into account, that in some cases this standard would be departed from to a considerable extent. The length of the second stage might be shortened or, in some cases, lengthened.
We first found adhesion of the colon in its new position in an embryo of 63 mm. fig. 16 is drawn from the 63-mm. specimen, the small intestine of the loop having been removed with the greater part of its mesentery to give an exposure of the colon. The (originally umbilical) colon, passing transversely across the neck of the coil—mass, has been laid down against the pancreas, duodenum, and inner part of the kidney, and its reversed strip of mesocolon is also laid against the dorsal structures and stretches between the colon and the arterial axis, as in the earlier stages.
The adhesion in the 63-mm. foetus is in the region of the duodenum, the caecum and intervening part being still free.
On the left side the colon and mesocolon lie free on the dorsal wall, but they cover a larger area here than at the beginning of the second stage: at that time the “descending colon” barely reaches the inner border of the left kidney, but now it runs down the middle of that organ. The left “transverse colon” is also at a somewhat higher level than in the second stage: i.e., it more decidedly invaginates the lower and back wall of the omental bursa and passes higher up behind the stomach.
Our meaning in describing this part of the colon as “invaginating” the bursa may perhaps be better understood from a glance at fig. 17. This represents in a schematic fashion what would be found in a longitudinal dorso-ventral section along a line such as at in figs. 13 and 16. The plane of mesocolon is seen applied to the dorsal wall, with the transverse gut cut through at its upper end and the descending colon divided below. The upper one is seen to be projecting into the bursal sac, invaginating its lower wall and thus coming to lie behind and below the stomach. Small intestine is indicated in front of the colic plane. The second figure shows how, by elongation and fusion occurring at a much later period, the definitive condition can be attained in this region.

Fig. 16. Foetus of 63 mm. in which the small intestine has been cutaway. The caacum is in contact with the dorsal wall, but not yet adherent, and the mesocolon of the loop can be recognised, laid with the colon across the duodenum. The “jejuno-mesocolic fold " is clearly seen, and, being practically opposite the old colic angle (see fig. 15, D), can be taken as marking the base of the mesentery of the loop. Proximal to this the colon has come secondarily into relation with the omental bursa, in line with the left transverse colon.
It is unnecessary to go into the particulars of our findings in individual specimens throughout the rest of foetal life; a good idea of the progress in this stage can be obtained by observing the relative positions of the great gut in a few foetuses of different ages, such as are combined in fig. 18.
In this figure the positions of the colons in various stages are marked Factors concerned in causing on a chart of the dorsal wall. No. 1 represents the gut at 45 mm.: it is laid down on the dorsal wall and just reaches the inner edge of the kidney, stretching its mesentery, a, to its full extent, while its caecal end is shown in interrupted lines to indicate that it is not in contact with the wall behind. The condition at 63 mm. is shown in 2, where the mesocolon on the left side is seen to be extended so that the gut rests on the left kidney, while its upper part, in relation with the bursal sac, is represented in interrupted lines: on the right side its caecal end is in contact with the dorsal wall just above the iliac crest below the right kidney, and its mesocolon, b, is reversed and laid down across the duodenum and on the wall as shown. The arrow indicates approximately the position of the one-time colic angle beyond which the mesocolon of the right side proper extends. Nos. 3 and 4- are stages in foetuses of 125 mm. and 160 mm. respectively: the peripheral extension is evident on the left, and to a less extent on the right. The mesocolon exhibits some attachment in the duodenal region in the 63-mm. stage and a little later the caecum is found to be adherent, so that these parts are relatively fixed and the extension on the right occurs slowly between them. No. 5 is the position of the caecum and colon on the right in a full-term foetus; its position on the left practically corresponds with 4 and is not shown. The great growth in length of the ilio—pelvic colon in the later stages is not shown. The position of kidneys and suprarenals is not quite as shown in the later stages, as these organs assume a rather higher level, but this need not be regarded from our present point of view.

Fig. 17. The first figure is a schematic section down the left side of the abdomen, as along the line a. in fig. 16 or fig. 13, to show the relation of the upper part of the abdominal colon to the omental bursa after it has been turned to the left. The abdominal mesocolon is cut at m, and p is the body of the pancreas in the back wall of the bursa. The second figure shows how the adult condition can be reached from this: the elongating mesocolon fuses with the bursa to form the transverse mesocolon, below this it is fixed to the wall, and at its lower end may be free more or less for the iliac colon.
Looking at this series of positions at different ages, it is evidently to be concluded that the whole colon is elongating, its left curve is increasing to the left and upwards, and its right curve to the right and upwards, though to a less extent, probably owing to the larger resistance of the right lobe of the liver. The increasing curve of the colon is only possible when there is a corresponding increasing breadth of mesocolic area. This is simply seen on the left side, where the mesocolic sheet is fixed in the middle line, and its increasing breadth or depth is only a continuation of the process which forms the median septum in the growing abdomen of the second stage. In the figure the added areas of mesocolon are shown by the dotted lines. On the right side, however, the mesocolon is at first free, and it is only after it has become fixed that it is necessary to have an increasing mesocolic area to keep pace with the slowly increasing colic curve. The fixation of the structure takes place fairly soon : the mesocolon, b, in the figure becomes adherent to the back wall shortly after the 63—mm. stage, so that the mesocolic areas of the succeeding stages are added to it. Thus we may say that the mesocolon of the loop differs from that of the left side in that it does not broaden before the intestinal return and only slowly and to a less extent after this has taken place.
So far, then, it may be said that a general progressive widening of the right and left mesocolons goes on pawl passzt with the elongation of the colon and the increase of its curves.
We have worked out some of the details of the process, and think that there is some reason to believe that the widening of the mesocolon, at any rate on the right side, is not directly dependent on the growth of the colon. It is not necessary, however, to go into that point, and it will suflice to look on the two enlargements as associated and more or less corresponding with each other.
As the two mesocolic areas widen they become adherent to the dorsal structures. Without dwelling on certain local peculiarities, it may be said that the mesocolon in a general way shows adhesion increasing from the centre toward the periphery, so that there remains a broad strip of free membrane between the gut and the adherent portion, and this strip is necessarily that in which active increase must be going on at the moment.
In this way the mesocolon becomes gradually fixed to the dorsal structures, a fate which overtakes the gut itself when the growth of the intervening strip ceases. In the case of the left upper colon the contact and adhesion is with the wall of the omental bursa, and has already been mentioned.

Fig. 18. Plan to show the position of the colon at different periods. For explanation see text. The abdominal mesocolon is seen at a, and that of the loop at 5.
It seems to us, then, that the activities which constitute the changes seen in the third stage may be said to be confined to the colon and mesocolon, the latter extending in its plane and fixing itself as it extends. It follows from this View that there is no fixation of the mesentery of the small intestine. In fig. 18, for example, the mesocolic area, b, is fixed between the gut and the straight dotted line: to the left of this the mesentery would hang free, so that the obliquely directed “attached border of the mesentery” is really the extreme left limit of the area of adhesion of the mesocolon of the loop.
The caecum during foetal life is very constant in its level, about the crest of the ilium. As the liver recedes the colon above this is bowed out to form the bend which is ultimately known as the hepatic flexure.
We have not carried our investigations into post-partum development, and therefore have no suggestions to offer concerning the subsequent shifting of any portion of the intestines.
Summary
The greater part of this paper is occupied by an account made up mainly of descriptions of a large number of inter-connected observations, and it would be almost impossible to bring these into the form of an abstract or summary. We propose, therefore, to limit our remarks under this heading to a statement of the main or large conclusions to which we have come, passing over the many smaller matters which may be found in the paper itself.
We have divided the evolution of the adult type into three stages :—
- The stage in which an umbilical “hernia” of the bowel exists, lasting from the condition of the “median” intestine to the time of return to the abdomen.
- The stage of return and rotation, occurring about the tenth week, and lasting for a short but variable time, coming to an end when the whole length of the colon is in its proper plane relative to the small intestine.
- The stage of extension of colon and its mesocolon in that plane, lasting till after birth. This stage is not really one in the course of rotation proper, for this is confined to the second stage, the first stage being a preparation for it.
The essential character of the first stage is the presence of an umbilical loop with its proximal limb lying to the right of the distal limb. The position is brought about by the depression of the proximal limb as a result of the enlargement and downgrowth of the liver carrying with it the vitello-umbilical venous anastomosis on its visceral surface.
Towards the end of the first stage there is rapid growth of the proximal limb and its mesentery, so that a mass of coils occupies the umbilical sac, along the left side of which the distal limb, consisting in part of caecum and a portion of colon, is placed without coils.
The second stage starts with the somewhat sudden return from the umbilical sac to the abdomen. The return is due to the fall of intraabdominal pressure owing mainly to relative decrease in liver mass: thus the extra—abdominal (intra-amniotic) pressure pushes back the contents of the sac. The return is not en masse, but the proximal limb returns first in continuity of length, the caecum being retained to the last in the sac owing to its larger size compared with the colon immediately continuous with it. The sudden and complete nature of the return may be due to resistance to the movement at first holding the coils in the sac; the resistance would be owing to the size of the mass of mesentery just inside the sac compared with that passing through its opening. As the intraabdominal tension is falling all the time, it seems probable that, when the slight resistance is at last overcome, the return could go on to its end without stop.
When the coils of the proximal limb return they must occupy the lower part of the abdomen below the liver, and they pass first into the cavity on the right of the intra-abdominal colon and mesocolon: these form, at the end of the first stage, a median “septum” which extends in the cavity from the liver above to the pelvis below. As the coils spread out below the liver they pass to the left, pushing this septum before them to the left and backwards, so that the colon and mesocolon lie against the dorsal wall behind the coils. Moreover, in going to the left, the coils have passed below the continuity of the abdominal colon with the part still remaining in the umbilical sac, and also below the main mesenteric vessels which also remain in the sac with the colon. Thus, when the caecum returns with these vessels toward the end of the movement, it must lie on top of the coils, between them and the liver. In this way the rotation is partly accomplished, a large part of the small intestine having passed to the left below the upper part of the colon.
The caecum, at first wedged in between the liver and the intestine, is forced back from this position by the pressure of the growing mass of coils, and thus comes to lie to the right of the mesentery of the coils and behind them, with the rest of the originally umbilical colon placed transversely across the mesenteric neck of the mass. This essentially completes the rotation, brings the caecum against the back wall on the right side, and closes the second stage.
Rotation is limited to the bowel which constitutes, with its mesentery, the umbilical loop. It does not involve the duodenum, which is curved out separately as a result of the growth of the head of the pancreas on its left side. Nor does it include the abdominal colon, which with its mesocolon makes the “septum” already mentioned: this colon is continuous with the umbilical colon at the “colic angle” held up by the “retention band” in the mesocolon. The position in the adult of the original colic angle is rather to the left of the middle of the transverse colon, so that the actual rotation of bowel may be said to affect only the intestine lying between the duodeno-jejunal bend proximally and the left middle of the transverse colon distally.
The third stage shows the occurrence of widening in the mesocolic areas applied to the dorsal wall, in association with growth of the gut attached to them: thus there is an increasing length and curve of intestine and width of mesocolon. As the mesocolic areas widen adhesion takes place: iii a general way it may be said that the fixation spreads peripherally, following the enlarging curve of the gut. This process occurs, of course, behind the plane of the coils of small intestine and does not influence the amount or nature of the rotation. It goes on for some time after birth and in its course the splenic and hepatic flexures are produced.
We may conclude this short summary of our main conclusions by pointing out the great influence exercised by the liver in the production of the final conditions. No doubt this organ, with the Wolffian bodies, is largely responsible through its growth for the early entrance of the gut into the umbilical sac, as well as for the depression and turning to the right of the proximal limb of the loop. It occupies all the available space in the abdominal cavity and holds the median visceral structures in place up to the intestinal return. By a relative decrease ‘in its rate of growth it leads to the ultimate return of the loop, and it probably helps to accommodate the returned coils by a11 actual decrease in its size. It lies in contact centrally with the retention band and colon passing from this region to the umbilicus, and in this way keeps the returning coils down in the lower part of the cavity so that they pass to the left below the colon and mesenteric vessels. And in the third stage the slowness of the peripheral spreading of the colon and mesocolon is without ‘doubt associated with the gradual decrease of the relative size of the liver.
References
- ↑ These models were shown at the June meeting of the Anatomical Society. figures and separate accounts of each of the models would be a tedious addition to an already long paper, and would serve no useful purpose: we therefore have decided to dispense with such formal descriptions.
- ↑ We are indebted to Professor Wood Jones for a reference to a paper by Rost (Arch. f. klin. Chir., 1912) in which he describes bands of involuntary muscle fibre in relation with the proximal part of the transverse colon and elsewhere. It is possible that these may represent remains of the structures we have described, but we ave not gone into the matter and cannot speak definitely about it.
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Figures
- Intestine Development
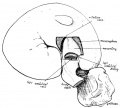
1 embryo 5 mm
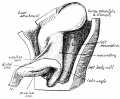
2 embryo 8 mm
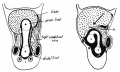
3 vitello-umbilical anastomosis
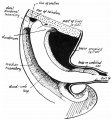
4 proximal limb of loop
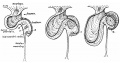
5 curve of the duodenum
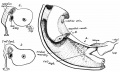
6 embryo 7.5 mm
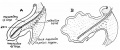
7 mesentery
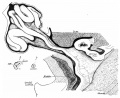
8 embryo 35 mm
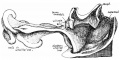
9 embryo 28 mm
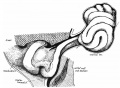
10 embryo 35 mm
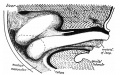
11 embryo 22 mm
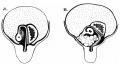
12 liver lobes and bowel
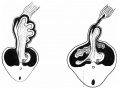
13 returning proximal limb of loop
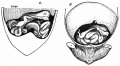
14 embryo 45 mm
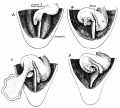
15 before and after bowel return

16 fetus 63 mm
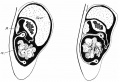
17 left abdomen
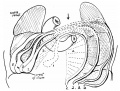
18 colon at different periods
Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - On the factors concerned in causing rotation of the intestine in man. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_On_the_factors_concerned_in_causing_rotation_of_the_intestine_in_man
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G