Paper - Observations upon Young Human Embryos
| Embryology - 18 Jul 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Wilson JT. Observations upon young human embryos. (1914) J Anat Physiol., 48(3): 315-51 PMID 17233002 PMC1288949
| Online Editor |
|---|

|
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Observations upon Young Human Embryos

James Thomas Wilson
Challis Professor of Anatomy in the University of Sydney, Australia.
Part 1
The appearance in recent years of Keibel and Mall's Manual of Human Embryology[1], following upon Keibel and Elze's Normentafein[2], marked an epoch in the formulation of our knowledge of specifically human development.
The comprehensive summary there offered of our knowledge of the earlier human ontogenetic processes provided for the firsttime a more or less connected account of these phenomena, but it also served to accentuate the still very sketchy and incomplete character of that knowledge.
Much of our belief in regard to the method of establishment of the human blastocyst is stil quite hypothetical, even if probable; and as regards the appearance of the earliest rudiments of the body itself, our knowledge isbased on a very few human specimens separated by intervals which it is important to fill in with the aid of intermediate or allied stages.
In the present paper I propose to give an account of the three youngest human embryos in my collection.
Previous writers have described and figured specimens of a stage of development more or less similar to those exhibited by the two older of the embryos which form the subject of this communication. Nevertheless, well-preserved specimens, of ages nearly corresponding to these, are of such comparative rarity that for some time to come it will still be desirable to have accurate records published of the form and structure of any that may become available for detailed examination. As a matter of fact, no two specimens hitherto described, however apparently similar in stage of development, have proved to be precisely identical in detail. It will appear in the course of the paper that each of the individual embryos under consideration presents features entitling it to independent description.
A more special interest attaches to the youngest of the three specimens, inasmuch as it would seem to exhibit a phase of development hitherto unrepresented in the records of early human embryos.
It possessed probably two, possibly three, pairs of somites, and may thus be determined as occupying a position in the gap between stages 2 and 3 of Keibel and Elze's Norrnentafel. These stages are represented respectively by Spee's embryo " Gle " [3]
Hitherto, or as far as I am aware, no human embryo has been recorded as exhibiting a smaller number of somites than five (in " Klb ").
I am inclined to believe that the embryo "E," No.1 of His' Normentafel (4), would have turned out to be of very similar character to that now about to be described, inspire of the somewhat greater length of the former (2.1 mm. as against 1.68 mm.).
Eternod's well-known embryo of 1-3 mm. this "No. 7 Vuill.") (5) may well represent a somewhat earlier phase.
The youngest embryo now to be described -the first of the three referred to- appears in my list of human embryos under the designation of "Hdr." I shall, however, refer to it in future simply under its catalogue number "H 3."
History of the Human Embryo "H3"
The specimen[4] was received by me so long ago as 25th May 1898, from my late friend Dr H.V.C. Hinder of Sydney, who had obtained it from a case of abortion on the previous day. The unopened chorionic vesicle had been placed in diluted alcohol. It was to outward appearance well conserved and perfectly intact when I received it, and it was at once transferred to picrosulphuric acid and then passed through graded alcohols.
From notes procured at the time by my friend Dr A.E. Mills, who was also associated with the case, it appears that the last menstruation period had begun on 12th April 1898 and ended on 16th April 1898. Abortion actually took place on 24th May 1898, but hemorrhagic discharge had appeared on 22nd May 1898. Thus the period that had elapsed since the beginning of the last menstruation up to the commencement of abortion was 40 days, or 36 days from the end of the last menstruation. The period that had elapsed since the due date of the lapsed menstrual period was 12 days.
According to present-day criteria the age of this embryo may be estimated as included in the period 18-21 days, and probably in the earliest part of this period.[5], p.90) quote Born's estimate of the age of embryo "Klb" as10-14 days. But if we take into account the more recently accepted criteria of age in early embryos, this age must be judged to be considerably underestimated. I shall show reason to regard embryo " Klb" as distinctly more advanced in development than embryo "H 3."
As originally received by me, the specimen was to al appearance in excellent condition. No defect in its preservation was recognisable throughout the period of its examination as an entire specimen prior to embedding, except that a portion of the yolk-sac was accidentally broken away during manipulation.
After embedding in paraffin a complete series of sections at 10 , was obtained. Unfortunately the histological condition of the sections was most disappointing. The attempt at an adequate fixation must, after all, have been too belated. Possibly also there may have been some over-heating in the paraffin oven. So unsatisfactory was the result at the time, that after a somewhat cursory examination the series was put aside for a considerable number of years. Now, however, on re-examination in connexion with the investigation of more recent specimens, ithas appeared to me to be well worth while to describe this early embryo in some detail. More especially perhaps do the photographic records of the entire specimen, which shows no external sign of structural deterioration, constitute original documents of some value for comparative purposes. And even the sectional series, although not fully adequate for the purposes of plastic reconstruction, turns out, on closer examination, to be of no little interest and value.
Characters and Dimensions of Chorionic Vesicle
The chorionic vesicle of "H3 " is illustrated in the photograph reproduced in PL. I. fig. 1, which was taken after the vesicle had been rendered transparent in cedar oil. It was flattened in its polar axis and measured 5mm. in its (practically a villous) polar diameter. Its equatorial diameter was about 8.5mm., inclusive of the vili; or, without vili, 6.4mm. in its longer and 5.7 mm. in its shorter equatorial diameter.

The chorionic dimensions were thus rather smaller than those of Spee's embryo "Gle" and almost identical with those of His' embryo "E."
As just indicated, the vili were unequally distributed over the surface of the vesicle. There was a richer equatorial villous zone, whilst the polar areas were freer from vili, though at no place completely bald. Oneof the polar areas, the antembryonic, was barer than the other.
The vili showed a very moderate degree of branching (cf. P1. I. fig.1).
After having been examined and photographed from both polar aspects, the vesicle was opened and the portion of its chorionic wall carrying the attachment of the body-stalk was separated from the rest. Theembryonic rudiment with its associated appendages was then subjected to closer examination and sketched and photographed from various points of view. (P1.I. fig.2, taken in cedar oil; also text-figs. 1-3.)


Micrometer Measurements
The following micrometer measurements were obtained of the embryo and its immediate appendages. The measurements were taken while the specimen was in cedar oil.
- Maximum cranio-caudal length of amnion - 1.78 mm.
- Maximum cranio-caudal length of yolk-sac - 2.26 mm.
- Apparent cranio-caudal length of embryo - 1.64 mm.
- Dorso-ventral extent of amniotic cavity from its dorsal convexity to line of reflection of somatopleure in the head region - 0.6 mm.
- Dorso-ventral extent o fyolk-sac 1.66 mm.
N.B. The yolk-sac was partly collapsed and crumpled, as shown in the photograph (fig. 2).
The specimen was next embedded in paraffin and cut in series of 10 micron sections in a plane intended to be transverse to the long axis of the embryo, but which turned out to be distinctly oblique. The sections constitute a practically unbroken series, but their histological condition is not very satisfactory. They are tolerably well stained in haematoxylin. The series was both cut and numbered in caudocranial succession. When the sections and figures are viewed with the dorsal embryonic surface away from the observer, the right and left embryonic surfaces are right and left, respectively, to the observer.
Form and Characters of Embryo and its Appendages
In PI. I. fig. 2 the embryo, along with its immediate appendages, amnion and yolk-sac, is seen attached to a fragment of the chorion by the body-stalk. The latter is rather acutely reflexed in a cranial direction, thus arching over a considerable extent of the amnion so as to form the actual roof of the caudal portion ofthatcavity. Theserialsections,when followed from behind forwards, show that the amnion only gradually becomes free from super incumbent cellular tissue of the stalk. Rather more than the caudal third of the amnion is thus intimately connected with the body-stalk, whose vascular mesodermal tissue, indeed, spreads out over this portion of the amnion like a hood.
The lateral portions of this hood gradually thin out as they clothe the sides of the amnion. In consequence of this extension of body-stalk tissue over the sides of the amniotic sac, the amnion exhibits in this region the appearance of a more or less vascular membrane (text-figs.4 and 5).


Within the amniotic chamber the outline of the embryo may be discerned (Pl. I. fig. 2), with its cephalic expansion elevated bilaterally into two prominent medullary folds, separated by a deep and wide medullary groove (cf.text-fig. 6).
There is either no dorsal flexure or kink of the embryonic body, or only a faint indication of one (cf. text-fig. 1).
Behind the broad cephalic region the embryo shows marked constriction and then appears to widen out into a foliate expansion, which forms the hinder third of the apparent embryonic body.


The superficial appearance of the foliate expansion suggests a widening and opening-out of the medullary plate in this region. This,however,is not the exact condition met with. The still widely open neurenteric aperture, text-fig. 7, is situated near the junction of the middle and posterior thirds of the embryonic region. Itspositionnearlycoincides with the ventrally open angle recognizable in PI. I. fig. 2, and also in text- fig. 1, at b, where also an arrow points to the site of the neurenteric aperture. The open angle at b in text-fig.1 really marks the anterior limit of the leaf-like expansion of the hinder part of the embryonic region. Immediately in front of the neurenteric aperture, the sections show the medullary plate as stil markedly infolded so as to form a deep and not very wide medullary groove.
Behind the aperture there is no shallowing out, but, on the contrary, the primitive-streak formation is here actually depressed into a deep cleft which continues backwards between two elongated, bolster-like, caudal swellings containing mesoderm (text-fig. 5).
It is these caudal swellings which form the lateral wings of the posterior foliate expansion of the embryonic region. The ectoderm covering them is not at al, or only slightly, thickened, whilst the primitive- streak ectoderm lining the deep dorsal furrow between them is thick and columnar like that of the medullary plate in front.
Apart from the elongated caudal cushions bounding the deep sulcus in the primitive-streak region, there is no posterior tail-prominence. The partoftheembryonic region containing the caudal swellings is,however, placed slightly at an angle with the rest of the embryonic area. Thismay be recognized in the photograph, fig. 2, PI. I., and is well shown in the outline in text-fig. 1.
The caudal cushions gradually fade away posteriorly in the floor of the hinder region of the amniotic cavity. This latter tapers into a narrow-pointed prolongation and ends at the plane of the hinder limit of the root ofthebody-stalk. It has no continuation into the body-stalk. Itsflooris formed solely by the continuation backwards of the floor of the vanishing primitive groove (text-fig. 8).


Close to the termination of the amniotic cavity there is a thick " cloacal membrane" connecting with the entoderm at the base of the allantoic duct (text-fig.9). The position of this cloacal membrane maybe compared with that described and figured recently by Grosser in a younger embryo (8, Taf. 27). There was no caudal stalk-like prolongation of the amnion, other than the above, to correspond with that described by Eternod in his 1P3 mm. embryo.
As there is no tail-prominence proper, the posterior limit of the embryo has been reckoned as if coincident with the hinder limit of the furrowed primitive-streak region,although the extremity of the future tail would undoubtedly be formed by hypertrophy of the bilateral caudal cushions far in front of this point. Reckoning, then, from the posterior limit of the primitive-streak region to the cranial limit of the cephalic medullary plate, the length of the embryonic area included 143 sections at 10 micron. In the table of measurements the apparent length of the embryo prior to embedding was 1.64mm.; the difference just indicated is doubtless to be accounted for mainly by shrinkage during embedding.
In text-figs. 1, 2, and 3 are reproduced the outlines of freehand sketches, from various points of view, that were made in the course of examination of this embryo, in toto, in cedar oil.
Text-fig.1 is the outline of an early profile view. It may be looked at along with the photograph, fig. 2, PI. I., which represents a dorso- lateral view of the specimen. Thetext-figure shows the degree of vertical curvature of the several regions of the embryo. The approximate position of the neurenteric aperture is indicated by the arrow. The region a to b is the primitive-streak region: the angle of its inclination to the main embryonic axis is to be noted.


If this figure be compared, e.g., with figs. 19 and 20 of Eternod's monograph L'ceuf human (5) (Geneve, 1909), illustrating respectively Selenka's outline figure of HylobatesRaflesi,and Eternod's outline of his 1-3 mm. human embryo ("No. 7Vuil."),it will be seen that the most outstanding difference is due to the acute ventral flexure of the primitive-streak region in these embryos ascompared with my specimen "H3." And this difference in flexure is correlated with the difference observable in relative position of the body-stalk. The absence of a ventral flexure of the primitive-streak region in embryo "H 3" permits of a rather acute reflexion, in a cranial direction, of the body-stalk.
Practically the same comparison may be made with embryo "Klb" (Kroemer-Pfannenstiel) of Keibel and Elze's Normentafel (2), which possessed 5-6 pairs of somites and must represent a somewhat similar developmental phase to that of the embryo under consideration. That " Klb" was slightly more advanced is indicated by its possession of a well- marked tail-prominence, of a closed-in hind-gut, and by several other features to be later commented upon.
The possibility cannot be entirely excluded that the extended position of the hinder part of the embryonic area of " H 3 " is somewhat abnormal, especially as the flexed attitude of the primitive-streak region was already attained in Spee's embryo " Gle" (3), which is certainly less advanced and is probably perfectly normal.
In any case, it is plain that the absence of ventral flexion of the primitive-streak region in "H 3" involves a reconsideration of the true length of the embryo. Ifareliable comparison is to be instituted with other human embryos of approximately the same stage, then we must deduct from the apparent embryonic length given above as 1-64 mm., nearly the whole length of the primitive-streak region behind the neurenteric aperture, since, in the embryos referred to, the neurenteric aperture lies quite close to the angle of a ventrally flexed tail-end of the embryonic body.
The deduction here suggested as necessary includes nearly the whole of the primitive-streak region. In the total of 143 sections of the embryonic area, inclusive of primitive streak, no fewer than 47 lie behind the plane of the caudal boundary of the neurenteric aperture.
Allowing for the slight caudal convexity behind this which is recognisable in comparable embryos, I estimate the true " embryonic length," for comparative purposes,to have been not over 1.25 mm. The actual proportion of the total of 1.64 mm. in front of the neurenteric aperture is almost exactly 1 mm.
Text-fig. 2 represents the outline of embryo "H 3,"as viewed from above and slightly from the caudal direction. The irregular area which has been hatched in the drawing represents the small portion of chorion connected with the body-stalk. The stippled area represents the dorso-caudal aspect of the body-stalk, and shows how it spreads out laterally over the hinder portion of the amniotic sac. Some indications of the chorio-placental vessels may be noted, but their detailed arrangement has not yet been worked out from the sections.
Text-fig. 3 represents an outline sketch giving the frontal "elevation" (Norma frontalis) of the amnion and head of the embryo, as viewed from the front and very slightly from the ventral side. The wide bulging area below the amniotic sac corresponds to what may be termed the "pericardial plate," although at the present stage the pericardium does not actually reach quite to its surface.
Mesodermal Somites
Not withstanding careful and prolonged examination of this embryo while in cedar oil prior to embedding, I was unable to detect the presence of differentiatedsomites. The other features of the embryo seemed to indicate that some at least of the earliest somites should be already in existence.
Examination of the serial sections has, in fact, confirmed this conjecture. Unfortunately,the histological condition of the paraxial mesoderm is not so satisfactory as to permit of a wholly reliable determination of the number ofsomitesrepresented. I have only been able to recognize in the section seriesonelineofsegmentalcleavagewithtolerablecertainty. I dare not assert that it is the only one present, and a critical examination of good silver prints of the photograph here reproduced on P1.I.fig.2 suggests the possible existence of up to three pairs of somites.
In any case, it is evident from the photograph in question that the number of somites could only have beensmall. Andinthisconnexionit must be pointed out that only a distance represented by about 25 sections intervenes between the tolerably abrupt posterior limit of the crescentic curve of the cephalic medullary fold and the anterior margin of the neurenteric aperture.
The distance between the probably corresponding points in the second of my specimens, "H 98," which possesses 9-10 pairsof somites,includes no fewer than 65 sections of similar thickness.[6]
We havethereforetoreckoninembryo"H 3"with25sections,against 65inthelatterstage("H 98"),asrepresentingthepossiblelongitudinalfield ofsomitedifferentiation.
Now, an inspection of figures of other early embryos suitable for com- parison will show that the differentiated somites do not extend close up to the anterior, and certainly not nearly to the posterior, limits as above defined, so that we have to deal with a lesser number than 25 sections, which could pass through mesodermal primitive segments.
In the Kroemer-Pfannenstiel embryo, for example (fig. 11D of Taf. I., Keibel and Elze's Normentafel (2), the five pairsofsomnitestherepresentdo not occupy much more than half the distance between the neurenteric apertureandthehinderendofthecephalicmedullaryplate. Fromtheir figure it would appear that the distance from the cranial limit of the first pair of somites to the neurenteric aperture was about 0.7 mm., and the actual segmented zone occupied only about 0.35.
Inasmuch,then,as inembryo "H 3" the entire available length from cephalic medullary plate to neurenteric aperture is represented by only about 25 sections,the actualsegmentedportionwas quiteprobablynot more than about half of that length-say 12 or 13 sections in extent.
I find in the older embryo "H 98" that the somites there met with extend over an average distance of about 5 sections each. If,asishighly probable,there is no great increase in size of the individual somites in these early stages, the conclusion may be drawn that only two, or at most three, pairs of somites could possibly have been differentiated in embryo "H 3."
I have already stated that one cleavage line may be definitely established in the sectional series. I can find no positive evidence in the series of any other, and I see little reason to doubt that here we have an example of a human embryo at about the stage of progressive differentiation of the first two, or possibly three, pairs of somites.
Chorda
The chorda is still in the stage of intercalation in, and is indistinguishable from, the entoderm.
Not only isthere no chord as such, but there isno unequivocal indica- tion of strict delimination, even of a chorda-plate, except for a distance of 3 or 4 sections immediately in front of the neurenteric canal. Itispossible that in the pharyngeal region the longitudinal zone of entoderm, which coversthedorsalmedian prominence,due to thegrooved medullary plate, may actually represent definite chorda-entoderm. But, except that a median strip of entoderm is thus more or less accidentally delimited, there is nothing to suggest the specific character of just that precise area. Text- fig. 13 illustrates the 4th section in front of the anterior margin of the neurentericaperture. Evenherethechorda-plateisevident,notsomuch by its differentiation from the rest of the entoderm, as by the appearance here of an entodermal indentation which leads caudally into the neurenteric opening itself.
Excretory Apparatus
There is no ascertainable pronephric rudiment, nor would one expect to find any at this early stage. In any case, the histological conditions are unfavourable to any critical verification.
Medullary Plate
The general characters of the medullary plate have already been alluded to in connexion with the form of the embryo. I.I.fig.2,alongwith text-figs. 1, 2, and 3 and the various sectional text-figures, will sufficiently illustrateitspresentphaseofdevelopment. Thephotomicrographictext- fig. 18 will further elucidate its structural arrangement so far as its rather poorhistologicalconditionwillallow. Text-fig.4illustratestheappearance of the medullary plate and groove a short distance (4-5 sections) caudal to the cephalic expansion of the plate, and 21 sections in front of the anterior lipoftheneurentericaperture. Nowhereisthereanycloserapproximation of the medullary folds than is shown in this section.
The histological conditions are not favourable enough for reliable observations either on neuromeric segmentation or on neural crest formation.
Absence of Sense Organs
There is no trace whatever of optic groove formation nor of auditory areas, although there is a diffuse and rather extensive thickening of the head ectoderm in the posterior cephalic region, which may possibly foreshadow the appearance of such areas.
Oral Region
On the ventral aspect of the head, immediately in front of the line of reflexion of the ectoderm from the free head to the "pericardial plate" swelling (text-fig.1), the ectoderm forms a plate, slightly recessed bilaterally, the depressions being separated from one another by a slight median prominence.
This gently recessed area alone represents the oral sinus (stomodaeum), which is thus very imperfectly formed.
The ectoderm of this oral sinus is separated from the entoderm of the pharynx by abundant mesoderm. Nowhere do ectoderm and entoderm even closely approach one another, so that the "primary pharyngeal mem- brane" ishere quite thick (0.06 mm.), and composed of al three germ layers. In fact, the mesoderm is here disposed as a specially thick compact mass of cells between the ectoderm and entoderm (text-fig. 11), and is continued backwards as such to the very caudal limit of the region of the rudimentary oral sinus. The area of this imperfectly formed oral sinus extends through sections 159-153 inclusive.
There are no evidences of any hypophysial formation.
Atlimentary Canal
The fore-gut shows a general agreement in character with that of the important Kroemer-Pfannenstiel embryo "Klb" of Keibel and Elze's Normentafel. The pharynx of this embryo has been described, modelled, and figured by Grosser (6) (also in the Keibel-Mall Manual, vol i. pp.446-7). In embryo"H3"thefore-gut is closed in for a distance represented by 30 sections, in front of the plane of its continuity with the yolk-sacatthe"Darmpforte." In the "Klb" embryo, the fore-gut was closed in throughout the extent of 32 sections. Grosser states (6) that its actuallength,allowingforcurvature,was400micra. Thefore-gutin H3"represents the anterior portion of the future pharynx. It has the usual laterally expanded form (text-fig. 6). It gradually diminishes in width when traced forwards until the cranial limit of the first primary pharyngealpouchisreached. Herethereisamore abruptdiminutionin width (text-fig. 12) and then a tolerably rapid tapering into the attenuated blind extremity of the gut,stil retaining its dorso-ventrally compressed character. Thistaperingterminalsegmentextendsinacranialdirection for several sections beyond the ascertainable anterior limit of the oral sinus. No such marked ventral flexion of the narrow terminal segment of the gut as that seen in embryo "Klb" is recognizable. At a distance of 5 sections from its cranial limit, however, the contact which has up to that point been maintained between the dorsal wall of the pharynx and the medullary plate suddenly disappears.' This separation corresponds with the now rapidly shallowing character of the medullary groove, as it is traced intherostraldirection. There is no recognisable trace of any Seessel's pouch.


I have made a rough wax-plate reconstruction of the entire pharynx, but unfortunately certain dislocations of structure detract from its value for purposes of publication. In most respects, however, it is reliable enough, and it brings into prominence the tolerably close similarity in various features between embryo "H 3" and embryo "Klb" in respect of the pharynx.
Throughout nearly its entire length the ventral pharyngeal wall is depressed into a median V-shaped furrow. On the exterior of the model this expresses itself in the ventrally keeled appearance which the pharynx thuspresents. Though present throughout practically its entire length, the keel-like prominence varies in its degree of salience at different levels. It is already visible even in the region of the "primary pharyngeal membrane." Caudally from the level of the oral sinus it becomes accentuated where it overlies the paired pericardial cavities (text-fig. 12). But it is recognizable even where the dorsal pericardial wall is perfectly level. Nevertheless, text-fig. 18 inevitably suggests that this keel-like form of the early ventral pharyngeal wall is somehow a result of the bilateral constitution oftheheadregion,as are the paired pericardial cavities themselves.
In the reconstruction model two maxima are apparent for the keeled ventralpharyngealwall. One of these (text-fig. 6) appears to coincide with that in embryo " Klb" in the region between the first and second pharyngeal pouches, which Grosser has determined as the thyreoid rudiment(6).
The second maximal development of the median ventral pharyngeal keel lies further forward (cf. text-figs. 12 and 18), in the region opposite the anterior end of the first pharyngeal pouch, and it is this accentuated portion of the keel which seems related to the imperfect interpericardial septumseeninthesetext-figures. Insucceedingsections(proceedingina caudal direction) there is a diminution of prominence of the ventral keel until it somewhat abruptly develops the second or thyreoid prominence seen in text-fig. 6. This then continues in a caudal direction until it merges in the median cranial boundary of the yolk-sac opening ("Darm- pforte"). Itthusappearsthatthethyreoidrudiment,thoughperhaps tolerably definitely localized in the cranial direction, has at the present stage no definite caudal limit.
There is seen on each side of the thyreoid rudiment in text-fig. 6, a "ventral pharyngeal groove," in Grosser's sense (cf. (6), pp.274-75). This, however, begins more abruptly and perhaps more caudally than in embryo "Klb." It makes its appearance in the same section (142) which shows the first trace of the definite thyroid rudiment. This is the eighth section behind the cranial limit of the first pharyngeal pouch. It may be noted that this bilateral " ventral pharyngeal groove," appearing as it does very abruptly, shows no sign of being continued from the ventral aspect of the first pharyngeal pouch, as described by Grosser in embryo "Klb." Further, it is more sharply defined, is of less cranio-caudal extent, and is somewhat more medial in position than would appear to have been the case in the latter embryo.
The first primary pharyngeal pouch is first met with in section 150 (text-fig. 12), in the form of an abrupt lateral extension from the side of the pharynx, exactly as in embryo " Klb " (Grosser's figs. 1 and 2 (6), and in Keibel-Mall(1),figs.314-5). The latter dilatation of the pharynx thus arising, is continued caudally, showing no definite limit in this direction. It does not exhibit the pronounced and progressive diminution in its dorso- ventral dimension visible in Grosser's model, but there is a slight indication of expansion inthe region corresponding to the "second pharyngeal pouch" of embryo " Klb, " as shown in that model.
The dorsal wall of the pharynx is bulged towards its lumen along three longitudinal zones, corresponding respectively to the grooved medullary plate medially, and the two dorsal aorta lateraly. Grosser'sdescription seems to imply that dorsal aortic impressions on the dorsal pharyngeal wall were absent in embryo "Klb"; but Keibel and Elze's fig. 5a, p. 19 (2), shows them quite definitely.
It has already been stated that, from a point 5 sections behind the anterior limit of the pharynx, the dorsal wall of the pharynx is in close appositionwiththegroovedmedullaryplate. Thiscontactdeterminesthe median of the three longitudinal prominences which are evident along the dorsalwallofthepharynx. The entoderm covering this median prominence isthatofthechorda-plate(cf.text-figs.6,14,etc.),butitishardly,ifatal, distinguishablefromtherestoftheentodermatthisstage. Intheserial sections there are some ruptures of the continuity of the pharyngeal entoderm, but this chorda-entoderm has very generally preserved its intimate relation to the medullary plate, which has afforded it a firm support. Thereisasyetnoclosed-inhind-gut. Thisconditionistobe correlated with the absence of any true caudal prominence and the non- flexed condition of the entire primitive-streak region. Underlyingthe extremehinderend ofthelater,thecavityoftheyolk-sacisprolonged, unilaterally, in a dorsal direction on the right side, into an asymmetrically placeddorsalentericgroove. Posteriorly (text-figs. 8 and 9), this groove leads into the somewhat dilated proximal portion or vestibule of the allantoicduct. This turns dorsally (text-fig.9),andrapidlynarrowsinto a canal of practically uniform fine calibre, being carried at first forwards into the cranially reflexed body-stalk.
The more medial wall of the asymmetrically placed dorsal enteric groove of the yolk-sac aforesaid exhibits, close to the proximal end of the allantoic vestibule, a well-marked depression which is closed by the thick cloacal membrane already referred to.
Allantoic Duct
The proximal portion or vestibule of the allantoic duct has already been referred to, as also its continuation into a narrow canal of uniform calibre prolonged not the reflexed body-stalk. Afterpassinginacranio-dorsal directionforsome considerabledistance(025 mm.),itbendsfurtherdorsally and then finally in the caudal direction and is now very slightly more attenuated. At a further distance of about 0.15 from its latest flexure, it now ratherrapidlyincreasesinsize,andends byexpandingintoa distinct vesicle. Throughout its entire tubular portion,it is lined by relatively thick cubical entoderm. Atitsexpansion,this quite abruptly passes into very thin entodermal epithelium with which the terminal vesicle is lined. The maximum diameter of the lumen of the vesicular expansion is about eight times that of the tubular allantoic canal. Eternodhasnotedthatthe allantoic cord of his 1'3 mm. embryo had "no appreciable vesicular enlargement." A distance of 14 sections, or 0.14mm., intervenes between the commencement of the dilatation and the terminal fundus of the vesicle. The total length of the allantois, exclusive of what I have termed the allantoicvestibule,but including the terminal vesicle and allowingforthe flexure of the canal is over 0.6 mm. It maybe said to follow a U-shaped course,the concavity of the U looking int he caudal direction.
Pericardum
The pericardium lies chiefly ventral to the pharynx, but it extends for a distance of about 005 mm. in front of the cranial limit of the later. It consists of paired cavities which are in actual communication with one another for a distance of only 9 sections,whilst it is extremely probable that in the case of the three more cranial of these sections the communica- tion is artificial and due to rupture of a delicate septum (cf. text-fig. 12).
The separation, partial or total, of the two pericardial cavities is effected differently in different regions of their extent.
Cranially and for a distance backwards of 15 sections (sections 152-137) there exists a genuine septUm proprium interpericardiacum, which is approximatelymedianin position. In no one section does this appear as wholly complete,but there is little doubt that it was actually completein its most cranial portion, as already indicated (text-fig. 12). As itistraced caudallythisseptumproprium becomeslesandlescomplete(text-fig.14), and gradually undergoes reduction (text-fig. 6) to the condition of a mere vestigial ridge of the ventral pericardial wall.
ut already, considerably in front of the plane at which there is any marked reduction of the septum, the appearance of the heart rudiments, with their dorsal and ventral mesocardial connexions (cf. text-figs. 14 and 6), effects the complete temporary separation of the more caudal regions of the two pericardial cavities.


These now, as they are traced further in the caudal direction, diverge gradually (text-fig. 15, right side) more and more from one another and are continued as the parietal (pleuropericardial) recesses.
The parietal recesses end blindly without establishing any communica- tion with any other coelomic cavity such as occurs later, e.g. in Mall's embryo, No. 391 (Dandy (7), pl.ii. fig.11).
Their lumina are only traceable for a distance of about 8 sections behind the termination of the crescentic curve of the cephalic medullary plate, or for a distance of about 12 sections behind the present posterior limit of the foregut (cranial margin of the "Darmpforte").
The-paired pericardial cavities are linedthroughoutbya definitelayer of ccelomic mesothelium, which in many of the sections has shrunk away to -some- extent from the mesoderm surrounding it (text-figs. 16 and 17).


The most cranialportionsofthe-rightand leftpericardialcavities.which are unoccupied by and wholly in front of the heart, occupy a position in thethick,plate-likemass of mesoderm uniting amnion and yolk-sac (cf. text-figs.1, 12, and 16). The dorsoventral thickness of this mesodermal mass is maintained through out at about 0.3 mm. In the cranial direction, it merges in an accumulation of loose vascular mesoderm which lies in the angle between yolk-sac and amnion, supporting, dorsally, the cephalic fold ofthelater,andcontinuous,ventrally,withthevascularmesoderm ofthe yolk-sac.
The presence of this mesodermal mass occasions a slight bulging ventral to the head of the embryo between amnion and yolk-sac (cf.text-figs. 2 and 3). Thismesodermalmassmayappropriatelybestyledthe"pericardial plate," inasmuch as it soon gives place, and probably with great rapidity at this juncture,to a bulging pericardium as found in such a closely succeeding stage of development as that represented by embryo"H 98,"v.infra,oras in Mall's embryo No. 391, F. T. Lewis' reconstruction, fig. 232, Keibel and Mall, vol.2.
In the present case of embryo " H3 " the pairedpericardialcavities have already appeared in this pericardial plate (text-figs. 16 and 17), but they have not yet so completely excavated it as to form a bulbous pericardial protrusion anteriorly, as is the case in the succeeding stage.
The pericardial cavity is first met within the serial sections at a plane lying 5 sections caudally to that of the anterior end of the medullary plate. Here (text-fig.16) the colomic mesothelium lining the fore end of the right pericardialcavityisfirstencountered. Thattherightcavityappearsas much as 12 sections before the left (text-fig. 17) is largely, but perhaps not wholly, to be accounted for by the obliquity of the sectional plane, which is considerable.
As the serial sections are traced in the caudal direction, the pericardial cavities are found to expand so as to occupy the place of the thick mesoderm of the " pericardial plate," and they gradually supplant that tissue, the interval between the amniotic and yolk-sacs remaining practically unaltered (text-fig. 14).
Heart
The heart itself is still almost completely duplex as regards not only its endothelial but also its myoepicardial components. The duplicity of the latter may be recognised from a comparison of-text-figs. 6 and 14. Text- fig. 6 shows portions of both right and left heart-tubes (myo-epicardial). Text-fig. 14' shows only the right heart. The plane of the latter section lies quite in front of the looped arterial end of the left heart, which is cut through in the section shown in text-fig. 6. To establish the connexions of this looped arterial left hear tone must pass caudally from the plane of the last-named text-figure.
The marked dissimilarity of the right and left hearts in that figure is due mainly to the obliquity of the plane of sectional ready referred to. It is somewhat difficult to state accurately the extent of this obliquity, but it may be indicated by the statement that the cranial limit of the first primary pharyngeal pouch on the right side lies some 8 or 9 sections in front of the corresponding point on the left. Were it not for this obliquity the right and left heart-tubes would appear tolerably symmetri- cally arranged, though both of them already show a marked degree of individual flexure. The relations of each heart-tube to the interpericardial septum may be easily recognised in the text-figs.6 and 14. It is evident that each heart-tube,in so far as it is separate, occupies its own pericardial cavity.

I have not yet been able to undertake a plastic reconstruction of this highly interesting state in cardiac development. This I hope to see carried out later on, although I fear that the degree of accuracy attainable will only be approximate. The section series is complete and quite intelligible, but there are histological imperfections which may be troublesome. This will be more especialythe case with the endothelial heart-tube and the aortic arch system.
The arrangement of the latter I have not been able thus far to clear up satisfactorilyfrommere examinationofthesectionseries. Sofarasmy observationsgoatpresent,Iam disposedtoregardtheconnexionsbetween the arterial heart-tubes and the dorsal aortme as stil of a more or les plexiformcharacter. Perhaps this relationship would also best explain the condition met with by Eternod in his 1.3 mm. embryo. And I can well imagine the cardiac condition I find in embryo "H 3 " to be immediately preceded by some such duplex tubular cardiac condition as that figured by Eternod in that interesting specimen. Inanycase,it is clear to me that there are several highly interesting pages of the history of early human cardiac development which are yet unwritten.
It is clear from the conditions met with in embryo "H 3" that the ordinarily accepted account of the origin of the single pericardium and heart in humandevelopmentwillnot hold good without considerable modification. Thus Tandler,in Keibel and Mall's Manual (English edition, vol.2), says:-" On the closure of the fore-gut ventrally the hitherto symmetrical pleuropericardial cavities come together anteriorly and fuse in this region, the median partition between them, the mesocardiuqm anterius, disappearing, while the me8ocardium posteriuos persists for sometime longer. The closely approximated but not yet fused endothelial tubes are now surrounded by a continuous myo-epicardial mnarntle (Molier). Finally, the two endothelial tubes come into contact, their partition wall disappears, and the unpaired heart cylinder is formed from the paired heart-tubes. This stage of the development of the heart occurs in the Kromer-Pfannenstiel embryo 'Klb,' etc. etc."
The final result here referred to as manifest in embryo "Klb " may be quite correctly described, but the hypothetical preliminary phases leading up to itare in alprobability neither so simple nor so direct as Tandler represents them.
The fusion of the right and left pericardial cavities is, as we have seen, for a time incomplete anteriorly, and the imperfect septum separating them isnota "mesocardium" atal in the ordinary sense,butaninterpericardial septum of mesoderm, quite independent of the heart-tubes themselves. Further, there occurs, prior to the state of single and continuous myo- epicardial mantle, a stage of an at least nearly complete duplex heart in which both right and left hearts possess myo-epicardial mantles which are, at least throughout the greater part of their extent, independent of one another, and which cannot be regarded as mere semitubular elements completing a simple median heart-tube by their coalescence.
I may say here that through the courtesy of Professor J. P. Hill,F.R.S., I have had the opportunity of looking through a superb section-series of an embryo of Peraineles in which the stage of cardiac development- evident with diagrammatic clearness-is obviously essentially identical with that which obtains in my embryo"H 3."
The condition in Perameles clearly shows that fore-gut closure in a mammalian embryo does not of itself at once determine median fusion of the primitively paired heart-tubes, for in this mammalian embryo one inds a completely closed fore-gut very greatly expanded in the transverse direction, and, attached to its floor but widely separated from one another, there are seen independent right and left myo-epicardial tubes. These appear to be of essentially similar character to those of embryo "H 3," and, like them, each shows a definite and more or less symmetrical (in Perameles apparently quite symmetrical) primary flexure, definitely marking the venous and arterial subdivisions of the tube.
No further discussion of either cardiac or vascular systems can be undertaken on the present occasion. Many details of the vascular arrangements have not yet been ascertained with suficient precision.
It may be stated that the vascular network in the wall of the yolk-sac is not only well established, but many of its channels show clear lumina with a definite endothelial lining.
Relative Position of Embryo "H 3" in the Human Embryonic Series
The relative position, in a "Normentafel" of human embryonic development, to be assigned to embryo " H 3," has already been discussed in an earlier section of this paper. I place it in the interval between stages 2 and 3 of Keibel and Elze's Normentafel (2).
Although His' embryo "E" (No.1 of his Normentafel (4) is only known to us by its external characters, I believe that, not withstanding its greater length, it offers the nearest parallel to "H3." Both of these embryos I believe to be slightly more advanced than Eternod's "No. 7 Vuill." of 1.3mm. length.
I have attempted to summarise the characteristics of embryo " H 3 " on the general lines of the paradigm of Keibel and Elze's Notrmerntafel, with a few modifications, so that the corresponding characters of various known embryos may be readily compared. That"H3"isdistinctlyinadvance of stage 2 of the Normentctfel (Spee's embryo " Gle ") is at once evident on comparison of the figures and descriptions. In particular the medullary plate and the pericardium and heart in " H 3" show a marked advance on the earlier stage.
The more outstanding characters whichseem todifferentiate"H 3,"as earlier, from embryo " Klb," representing stage 3 of the Normentafel, are the following:-
- In " H 3 " the embryonic length, as interpreted in the body of this paper,is slightly less than that of "Klb."
- In the former embryo no brain subdivisions were apparent in the otherwisewell-markedcephalicmedullaryplate. Ontheotherhand,the brain region seems rather more prominent in " H 3 " (fig. 2, PI. I.) than inthepublishedfiguresof"Klb."
- The neurenteric aperture was widely patent, and was both relatively andabsolutelyclosertotheheadregionin"H 3"thanin"Klb."
- In embryo "H3" there was no hind-gut. Ontheotherhand,the fore-gut, both in its dimensions and its degree of differentiation, differed onlyslightlyfromthatof"Klb."
- The chorda rudiment is in a much more backward stage in "H 3." Hardly any indication of it is visible, even in the form of a chorda-plate.
- The somites, although not quite conclusively determined in "H 3," were certainly fewer (not more than three) in number.
- The pericardium in "H 3" is stil in the condition of bilaterally paired and nearly symmetrical cavities, separated by a septum proprium interpericardiacum. The extent of the intercommunication between the two pericardial cavities is very limited.
- The heart of "H 3" is still largely duplex, consisting of right and left myo-epicardial mantles.
Summary of Characters of Embryo "H3"
Desqgnation. - Human embryo "H 3"(Hdr). CollectionJ.T.Wilson.
Dimensions.-Chorionic vesicle6.4 x 5.7 x 5mm., ex.vili. , , 85 x5,imld.,
Length, amniotic sac 1.78 mm., yielding 151 sections. embryonic region 1.64 mm., yielding 143 sections. yolk-sac (distorted) 2.26 mm.
Depth, yolk-sac (dorsoventrally) 166 mm.
Length (cranio-caudally) of cephalic medullary plate, estimated as to the caudal limit of its prominent fold, 0.6mm. i (plus or minus).
Age.-Period elapsed since beginning of last menstruation to commencement of abortion, 40 days. Periods ince end of last menstruation, 36 days. Period elapsed since due date of lapsed menstruation, 12 days. Estimated age of embryo, 18-21 days. No conjugal history. Menstrual history as above.
Body Form.-No dorsal bend of embryonic body.
Primitive-streak region (= tail end) deeply furrowed, and forming only a very slight angle with the body proper. No.tail-prominence other than caudal cushions bordering the furrow of the primitive-streak region.
Headendofbrain-plateflexedventrallyoverapericardial-plate-prominence, which is already visible.
Primitive-Streak Region.- Neurenteric canal stil widely patent Primitive-streakregion tolerably lengthy (047mm. in the section series) and deeply furrowed, and bordered by elongated, cushion-like caudal swellings.
Entire primitive-streak region still appears as the nearlydirectcaudal continuation of the embryonic body proper. Cloacal membrane posteriorly.
Mesodermal Segments.-Somites not visible on external examination; certainly fewer than five pairs, probably not more than two pairs, and not less than two.
Chorda.-Chorda entirely absent as such. Median strip of entoderm forming chorda-plate only recognizable with any degree of certainty just in front of the neurenteric aperture. But probably also in head region.
Nervous System.-Medullary plate expanded in front to form a brain- plate elevated laterally into prominent crescentic folds and flexed ventrally atitsanteriorend.
Medullary groove well formed throughout but no where closed. No clear evidence of intrinsic brain segmentation. No traces of commencing formation of eye, ear, or nose.
Hypophysis.-Noindicationofhypophysis.
Mouth Cavity.-Veryfaintindicationoforalsinus. Primarypharyn- geal membrane shows abundant mesoderm between ectoderm and entoderm. DigestireTract.-Fore-gutendsblindlyatcranialend. Consistssolely
ofwidepharynx. Extendsfor30sections(0-3mm.)infrontof"Darm- pforte.'" Hind-gutnotyetclosedin. Aproximaldilatationor"allantoic vestibule" at root of allantoic duct opens widely into a unilateral dorsal groove of the yolk-sac, or "dorsal enteric groove," whose medial wall exhibits a well-marked depression closed by a cloacal membrane.
Pharyngeal Derivatives.-First primary pharyngeal pouch is differenti- ated, but is separated from the ectoderm of the head by thick layer of mesoderm. Thereisageneralventralmediangrooveofthepharyngeal wall, whose maximal point of depression answers to the median thyreoid rudiment. "Ventralpharyngealgrooves"(paramedian)arealsopresent. There is a possible faint indication of the appearance of a second primary pharyngeal pouch.
Urogenital System.-No indication of any urogenital apparatus. Heart, Pericardium, and Vascwtar Systemr-Heart largely duplex; right and left myo-epicardial mantles occupy paired pericardial cavities, which are imperfectly separated by an incomplete septum propriuminter- pericardiacumn. Right and left epimyocardial hearts each exhibit sub-division into venous and arterial segments. Paired dorsal aortae.
Blood-vessels of yolk-sac contain corpuscles, but many show clear lumina lined by endothelium.
Amnion.-Amniotic sac extends for a distance of 8 sections in front of anterior.endofhead. It tapers at its hindered, first gradually,then abruptly, into a narrow-pointed recess floored by the continuation of the primitivestreak. The cloacal membranous connexion with the ectoderm lies at the right dorsolateral wall of this recess, very nearly at its apex. Mesoderm of the body-stalk extends like a hood over the hinder third of the amnion, and even in-front of the body-stalk, the mesoderm of the amnion is thickened.
Allantois.-The allantoic duct opens by a dilated " allantoic vestibule" intoa"dorsal enteric groove."
Sinuous in its course, it ends in a definite thin-walled, terminal vesicle. The total length of the canal,including the vesicle, is about 0.6mm.
Second Embryo "H 98"
For the second of my specimens, to be referred to as "H 98," I am indebted to the kindness of Dr F.P. Sandes, who obtained itasthe resultof abortion in a patient who had exceeded her menstrual period by 10 days.
| Online Editor Note H98 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Carnegie Collection - Stage 10 | |||||||||
| Serial No. | Pairs of somites | Size (mm) | Grade | Fixative | Embedding Medium | Thinness (µm) | Stain | Year | Notes |
| 7251 | 8 | E., 1.27 | Good | Formol | C-P | 10 | (Stain - Haematoxylin Eosin) | 1941 | "Singapore embryo." Univ. Cambridge No. H 98. Studied by Wilson (1914)[7] |
Technique
Dr Sandes brought the perfectly fresh and intact specimen direct to my laboratory, where itwas at once placed in isotonic 10 per cent. solution of formalin. After fixation in this solution the specimen was transferred to alcohol. After complete dehydration, it was cleared en bloc in cedar oil, when the embryo in its interior became clearly discernible.
The chorionic vesicle was then carefully opened up under the stereo- binocular microscope, and the embryo, with body-stalk, amnion, and yolk-sac, was isolated from the greater part of the chorion. Micrometer measurements were made of the various dimensions of the embryo, etc.,and the specimen was subjected to very careful and prolonged examination under the stereo-binocular microscope. It was also repeatedly photographed. On completion of the examination of the specimen in toto, it was double- embedded in cedar-oil-celloidin and paraffin, cut in faultless series of 10 AL sections transversely to its long axis, in caudo-cranial succession, mounted on albuminised slides, and double-stained in Benda's hoematoxylin, followed byeosin. The quality of the sectional series thus obtained is exceptionally good for early human embryonic material. A rent in the right wall of the yolk-sac is visible in the sections, but this must have occurred prior to fixation, since there is a small collection of effused blood adherent to one area of the interior of the yolk-sac wall.
The sections being cut in caudo-cranial succession and mounted with their latest cut surfaces downwards, it will be evident that when the sections are looked at with the dorsal aspect away from the observer, the right embryonic surface is to his right,and the left to his left. This relationship is preserved in the photomicrographs and drawings of the sections.
It should be noticed that the sections, though cut in caudo-cranial succession, are not numbered in this order as they are in "H 3." The numbers of the sections in "H 98" run cranio-caudally.
Chorionic Vesicle of "H98."
The chorionic vesicle of "H 98" (stereographic fig. 3, PI. I.) was somewhat oval and flattened, and whilst stil in the formalin fixative, it measured 6.6 x 5.3 x 4mm., exclusive of the vili. Ifthevilibeincluded inthemeasurements, the dimensions were 9 x 8 x 5 mm.
The chorionic wall was more or less villous throughout, except over part of one of the polar areas. Asissofrequentlythecasewithvesicles of approximately the same size, the vili were more luxuriantly developed around the equatorial zone of the flattened vesicle. As is also usual, the comparatively bald polar area of the vesicle was ant embryonic.
The stereographic fig.3,PI.I.,shows the chorionicvesicle,trans- lucent in cedar oil,viewed by transmitted light. The embryo with its yolk-sac is visible in the interior.
When the chorionic vesicle was opened the interior was found to be filled with the normal "magma reticularis," clearandtransparentandof semigelatinousconsistency.
Appearance and Configuration of the Embryo and its Immediate Appendages
As was to be expected from the circumstances of its reception and subsequent treatment, the condition of the embryo, etc., when isolated from the general chorion, appeared to be quite perfect. This judgment was borne out by the subsequent microscopical examination of the sectional series.
Apart from the one fact of its having been procured from a case of abortion (causation undetermined), there is no ground for regarding the embryo as in any way abnormal.
Figs. 4, 5, and 6,PI. I. and I.,illustrate the appearance presented bytheembryo,etc.,afterisolationfromthegeneralchorion. Infig.4, PI. I., is reproduced a stereo-photograph obtained by enlarging the negatives taken with Zeiss' "a 3" paired objectives in the Braus-Druner camera.
Fig.5,PI.I.,represents a photomicrograph of the same at a magnifica- tion of 28 diameters; whilst fig. 6, PI. I.,represents a very careful drawingbyMr HerbertBeecroft,made with the aid of various photographs both stereoscopic and other.
Figs. 4, 5, and 6,PI. I. and I.,illustrate the appearance presented bytheembryo,etc.,afterisolationfromthegeneralchorion. Infig.4, PI. I., is reproduced a stereo-photograph obtained by enlarging the negatives taken with Zeiss' "a 3" paired objectives in the Braus-Druner camera.
Fig.5,PI.I., represents a photomicrograph of the same at a magnification of 28 diameters; whilst fig. 6, PI. I.,represents a very careful drawingbyMr HerbertBeecroft,madewiththeaidofvariousphotographs both stereoscopic and other.
Dimensions of Embryo "H 98."
- Length from anterior limit of amniotic collaring front of head to posterior limit of body-stalk at chorionic attachment - 1.8 mm.
- Length of embryo, without body-stalk, from tip of anterior end of brain-plate to tip of tail, measured in a straight line - 1.27
- Greatest cranio-caudal length of amniotic sac - 1.38
- Greatest cranio-caudal length of yolk-sac - 1.46
- Greatest dorsoventral extent of amnion in head region - 0.86
- Greatest dorsoventral extent of yolk-sac measured from line of reflection of amnion - 1.32
From the illustrations given, it will be seen that embryo "H 98" possessed 8 pairs of somites completely differentiated. My original notes of the examination of the specimen contain the statement that there are "probably 9 or 10 pairs of somites, of which the first pair appear to ne very small."
The figures, cf. especially fig. 6, PI. I., show indications of the existence of 10 pairs. But a plane reconstruction of the segmented mesoderm, since made, indicates that neither the first nor the last of the 10 segments is definitely segmented off from the axial mesoderm in front of, and behind, the somites. I can therefore only indicate the stage of somite formation by the statement that there are 8-10 pairs.
In Mall's embryo No. 391, reconstructed and described by Dandy (7), and also by F. T. Lewis (Keibel and Mall's Manual, vol. 2, fig. 232), there were 7 pairs of somites differentiated. Embryo"H98" thus appears to be more advanced than the latter embryo, which is otherwise remarkably similar to it-to the extent of the appearance of one additional somite pair.
A feature of considerable interest in embryo., 'H 98" is its manifestation of a well-marked, dorsally concave body-flexure (the "dorsal kink," "dorsal flexure," or "Knickung"). This was present in almost as high degree in the Mall embryo No.391. In Dandy's opinion it was "partly natural and partly an exaggerated postmortem condition." He remarks (loc.cit.) that "we should naturally expect a dorsal concavity due to the greater develop- ment of the structures in both the anterior and posterior regions of the embryo," and then proceeds to account for the supposed post-mortem accentuationofthischaracter. IcannothelpfeelingthatDandyhasbeen influenced in his attitude towards the question of this dorsal flexure or kink by the weight of Keibel's authority against considering it as a normal feature. I therefore desire specially to emphasise the following considerations in regard to the same feature as occurring in embryo "H 98":--
(a) The chorionic vesicle was obtained fresh and unopened; (b) it was fixed in 10 per cent. isotonic formalin, in accordance with the recommendations of Mann, in order to obviate or minimise to the utmost the tendency to tissue distortion; (c) the photographs taken through the walls of the cleared, but as yet unopened, chorionic vesicle already showed the well-marked dorsal flexure; (d) a consideration of the later photograph (fig. 4-6, P1.I.andI.) will, I think, convince the unprejudiced observer that the existence of the developmental phase of the heart and pericardium shown in the photograph must almost necessarily have determined the temporarily erect attitude of the cranial end of the body, or, more strictly speaking; of the cephalic portion of the medullary plate. And it is this erect attitude of the latter which is the main (though not the only) factor in the production of the dorsal kink. The other factor, as indeed has been already implied by Dandy, is the elevation of the tail end. And this is in turn no doubt due to the ventral attachment of the embryonic stalk.
It is, of course, open to anyone to throw doubt upon the normal characteroftheseandsimilarembryos. Butitwillnotdotobasesuch doubts solely upon the ground of the existence of the flexure, whose validityasanormalfeatureissubjuice. Anditistobenotedthatthe Mall-Dandy embryo, although aborted, was from a case of traumatic abortion, in which there was therefore no reason to suspect any natural or inherent abnormality of the ovum.
With regard to the normal occurrence of the flexure in question in the human embryo, Keibel has repeatedly expressed himself as skeptical. In hislatestutteranceon thissubject(Manual,vol.1,1910,pp.66-7)he again records his judgment that the normal occurrence of a dorsal flexure in embryos of the stage o fKollmann's "von Bulle" embryo, or later, has been disproved. But he still leaves open the question of itsoccurrenceasa normalfeatureinembryosoffrom6to12pairsofsomites. At the same time, he regards the evidence for its normal occurrence even in these stagesasinsufficientlybased upon Eternod's 211 mm. embryo and Spee's embryo with 7 somites.
But, in the light of the condition met with in the Mall-Dandy embryo, andnowagaininmy specimen"H 98,"Isubmitthatthenormalexistence of the dorsal flexure during stages of from 7 to 10 pairs of somites should beadmitted. Itistruethatthesetwolatestspecimensarenotbythem- selveswhollyunimpeachable,beingabortedspecimens. Butthetraumatic character of the abortion in the one case, and the extreme care taken to avoid the possibility of distortion in the other, tend to establish the naturalnessoftheappearance. Eachcorroboratestheother. And,inview of the facts as now before us, it is rather too much to ask us to set aside the positive evidence of flexure in a whole series of cases, viz. Spee's embryo with 7 somites, Eternod's embryo of 8 somites, the Unger embryo of Keibel himself with 9 somites, the Mall-Dandy embryo of 7somites, and "H 98" with 8 to 10 somites,aseachandalabnormal,whereas the fact of the matter now is that, so far as I am aware, no embryo of the stage of 7 to 10 somites has ever been recorded without the flexure in dispute.
Further,embryo"SR" No.2,ofHis'Normentafel,is(Ithinkun- warrantably) set aside by Keibel as probably abnormal, apparently solely onaccountofitspossessingadorsalflexure. Now,thedimensionsofthis embryo would tend to indicate that, although His does not show nor describe somites in this stage, the embryo must have possessed them. Embryo " SR," in fact, shows a tolerably close resemblance, so far as one cantel,to the stage of 7 to 12 somites,andmay beconfidentlyaddedto the list of embryos of this stage exhibiting the dorsal flexure.
Other features which may be noted in the figs.4 to 6, PI. I. and I., are (1) the differentiation of the cephalic portion of the medullary plateintofore-,mid-,andhind-brainregions. Itistobenotedthatwhat appears to be a primary cerebral flexure is at this stage intrinsic to the fore-brainregion. Themid-brainshowsnotraceofflexureatal. One can recognise the same characteristic in the Mall-Dandy specimen and in Kollmann's"vonBulle"embryo; perhapsalso,butmoredoubtfully,inthe Kroemer-Pfannenstielembryo"Klb." Ihavenotseenattentiondrawnto this distinction, which is very striking in view of the fact that, later on, what we cal the "primary cerebral flexure" lies in the region of the mid-brain.
(2) The contour of the fore-gut may be followed in the figs. 5 and 6 from its blind anterior extremity to its communication with the yolk-sac throughthewidevitellinestalk. The posterior continuation of the hind-gut from the caudal limit of the vitelline stalk into the tail region of the embryo may also be perceived. Incidentally, in connexion with the problem of the flexure, attention has been directed to the extremely prominentpericardialdevelopment. Thisconditionrepresentsadistinct advance on that seen in Mall's embryo No. 391, cf. especially F. T. Lewis' fig.232intheKeibel-MallManual,vol.2. Butitisanadvanceinthe samedirectionasthatoftheprogressfromthestageofmy "H 3"through thatofembryo"Klb"tothatoftheMallembryoNo.391. Itisaprogress which consists of a continuing process of excavation of what I have called the "pericardial plate" in embryo "H 3," and when that process of excavation has reached its limit, of a further process of bulbous expansion ofthepericardialcavity.
I imagine that the condition seen in "H 98" must represent the maximumstageofthisrapidexpansion. WhenthestageofKollmann's embryo"von Bulle"isreached,thedevelopmentoftheheadhasalready overtaken the pericardium and the latter is forced ventrally by the now moreorlessoverhanginghead. Theentireprocessseemstomeaperfectly obvious and intelligible swaying in the balance of competing developmental processes. Idonotevenseethatoneisforcedtobelievethatthe equilibrium isalways preciselysimilarlymaintained at alcorresponding movements in the otherwise perfectly normal course of development.
It is quite likely that another factor in the maintenance of develop- mentalequilibriuminthisregionistherateofamnioticexpansion. We know perfectly well that, at least in later stages, the rate of growth and expansion of the amnion varies within tolerably wide limits without being intheleastabnormal.
Iseelitlereasontodenythatthesemaybequite normal variations in a similar sense, though of lesser degree, even at the earlyperiodnow underconsideration.
The carrying out of an adequate structural analysis of this embryo from the serial sections has been largely in suspense during my personal absencefromSydney. EreIleftIhaddonesomeworkinthedirection of reconstruction of the heart, but I do not propose now to enter upon the discussionofthesectionalanatomyofthisembryo. Thiswil,Ihope,form a portion of the subject-matter of Part I. of this contribution to our knowledge of early human embryos.
The sequel will also, I hope, contain a descriptive account of the third embryo of the series, viz."H 86" in my list.
List Of References
- ↑ KEIBEL and MALL, Manual of Human Embryology, Philadelphia and London, 1910-12.
- ↑ KEIBEL and ELZE, Normentafeln zur Entwicklungsgeschichte des Menschen, Jena, 1908.
- ↑ SPEE, v., "Neue Beobachtungen fiber sehr friuhe Entwickluiigsstufen, etc.," Arch. f. Anat. u. Physiol., Analt. Abt., 1896., and the Kroemer-Pfannenstiel embryo " Klb."
- ↑ A lantern demonstration of slides of this embryo was given at a meeting of the Anatomical Society of Great Britain and Ireland at a meeting held on 16th January 1914.
- ↑ Keibel and Elze (Normentafel KEIBEL and ELZE, Normentafeln zur Entwicklungsgeschichte des Menschen, Jena, 1908.
- ↑ It is perhaps necessary here to point out that although in the later embryo the neurenteric aperture was closed, yet it is quite easy to determine the site corresponding to it, for here the chorda merges in mesodermn which cuts into the overlying medullary plate like an inverted keel (text-fig. 10).
- ↑ Wilson, J. T. 1914. Observations upon Young Human Embryos. J. Anat, 48, 315-351.
(3) SPEE, v., "Neue Beobachtungen fiber sehr friuhe Entwickluiigsstufen, etc.," Arch. f. Anat. u. Physiol., Analt. Abt., 1896.
(4) His, W., Anat. menschl. Embryonen, 1880-5, Taf. x.
(5) ETERNOD, L'wrufhuman, Genbve, 1909.
(6) GROSSER, O., "Zur Entwicklung des Vorderdarmes menschlicher Embryonen,etc.," Sitzungsber. d. kaiserl. Akad. d. Wiss. in Wien. Math.-Naturw. Klasse, Bd. cxx.,Abt.i.,1911.
(7) DANDY, W. E., " A Human Embryo with Seven Pairs of Somites, measuring about 2 mm. in Length," American Journal of Anatomy, vol. x. p. 85, 1910.
(8) GROSSER, O., "Ein menschlicher Embryo mit Chordakanal," Anat. Hefte, Heft 143, p. 653, 1913.
Explanation of Text-Fig. 18.
Photomicrograph(x 100)ofsectionNo.149ofhumanembryo"H3,"atthe level of the cranial limit of the 1st right primary pharyngeal pouch.
The section is ratherobliquelycut. Itpasses throughthecephalicportionof themedullaryplate,whoseedgesare welldefined. In contact with the median ectoderm of the floor of themedullary groove may be seen the adherent strip of pharyngeal entoderm which represents the chorda-plate. The pharynx shows its medio-ventral keel immediately dorsal and corresponding to the septum proprium inteirericardiacum (see text). The latter nearly completely separates right and left pericardial chambers, which show a very definite coelomic epithelial lining. The right chamber contains portions of tissue from tangential grazing of the cranial bulgingoftherightmyo-epicardialtube. Thedorsalaortrearevisibledorsalof thepharynx.
Note the entoderm lining the roof of the yolk-sac in the lower part of the figure, and the amnion on either side in the upper part.
Explanation Of Plates I.-III. Figs. 1-6.
Photomicrograph of chorionic vesicle of human embryo "H3." x 7 (reduced from x14).
Zeiss' 35 mm. micro-projection objective.
The photograph was taken after slight surface staining with hematoxylin and subsequent cleaving of the entire vesicle in cedar oil.
Note the very slightly branched vili on the exterior of the vesicle. Theembryo, with its more intimate appendages, amnion and yolk-sac, are visible in the interior. The yolk-sac is the larger crumpled sac mainly to the left of the embryo. The sharply defined clear outline of the more anterior portion of the amnion may be seen to the right and (in the figure) above the nearly vertically directed embryonic rudiment.
Plate I. Fig. 2.
Photomicrograph(x155)ofhuman embryo"H3,"withitsamnion,yolk-sac, body-stalk, and a portion of its chorion, viewed from the right dorsolateral aspect.
Zeiss' 35 mm. micro-projection objective.
The somewhat collapsed and crumpled yolk-sac is seen in the lower part of the figure. The patch of chorion giving attachment to the embryo, etc., in the upper part of the figure presents its edge, for the most part, to the observer and shows no detail. Descending from this towards the embryo is the body-stalk and tail-half of the amnion. This latter appears as if separated from the anterior portion of the amnion by a deep crease of the membrane. The body-stalk is evidently directed dorsally and cranially from the caudal end of the embryonic rudiment. Thie medullary folds of the embryo are visible within the amnion anteriorly, and the leaf- like are a traversed by the primitive streak posteriorly.
Plate II. Fig. 3.
Stereo-photomicrograph of chorionic vesicle of human embryo"H98." The photograph was taken from the specimen in cedar oil by transmitted light, under the Braus-Druner camera, Zeiss' stereo-objectives a', magnification x 2-8.
The vesicle is translucent, and shows the embryo and yolk-sac in its interior.
Plate I. Fig. 4.
Stereo-photomicrograph (x16) of human embryo "H 98," with amnion, body-stalk, and yolk-sac. Enlarged from negative taken with Zeiss' stereo-objective a2 at four diameters. The specimen was photographed in cedar oil by transmitted light.
Compare with figs. 5 and 6, P1. III.
The body-stalk is practically complete, although the portion of clhorion which had been left in connexion with it became detached whilst in the cedar oil. The dark patches in the body-stalk represent the blood content of vessels. The pericardium and heart form a prominent bulging mass ventrally to the head, which is, in consequence, erect in attitude. The amnion and yolk-sac are both almost wholly free from folds, and there is no evidence of shrinkage.
Plate III. Fig. 5.
Photomicrograph (x 28) of human embryo " H 98," with amnion, body-stalk, and yolk-sac.
See description of fig. 4 and text, and compare with figs. 4 and 6, PI. II. and III.
Plate III. Fig. 6.
Human embryo " H 98," with amnnion, body-stalk, and yolk-sac.
Drawn from photographs and stereo-photographs by Mr Herbert Beecroft, Sydney. See description of fig. 4 and text, and compare with figs. 4 and 5, P1. I. and III.

Fig. 1.
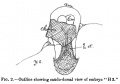
Fig. 2.

Fig. 3.
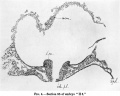
Fig. 4.
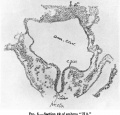
Fig. 5.
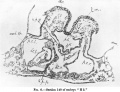
Fig. 6.
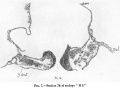
Fig. 7.
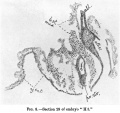
Fig. 8.
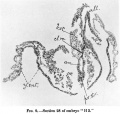
Fig. 9.
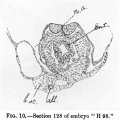
Fig. 10.

Fig. 11.

Fig. 12.

Fig. 13.
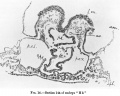
Fig. 14.
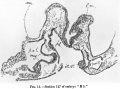
Fig. 15.
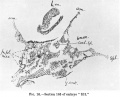
Fig. 16.
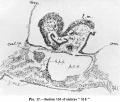
Fig. 17.

Fig. 18.
- Wilson Paper Images: Fig 1 | Fig. 2 | Fig. 3 | Fig. 4 | Fig. 5 | Fig. 6 | Fig. 7 | Fig. 8 | Fig. 9 | Fig. 10 | Fig. 11 | Fig. 12 | Fig. 13 | Fig. 14 | Fig. 15 | Fig. 16 | Fig. 17 | Fig. 18
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Reference
Wilson JT. Observations upon young human embryos. (1914) J Anat Physiol., 48(3): 315-51 PMID 17233002 PMC1288949
Cite this page: Hill, M.A. (2026, July 18) Embryology Paper - Observations upon Young Human Embryos. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_Observations_upon_Young_Human_Embryos
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2026, July 18) Embryology Paper - Observations upon Young Human Embryos. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_Observations_upon_Young_Human_Embryos
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G