2017 Group Project 2
| 2017 Student Projects | |||
|---|---|---|---|
|
Kidney
The kidneys are two bean-shaped organs located in the abdomen that play an important role in the renal system (also known as the urinary system). They arise from the intermediate mesoderm and begin their development (nephrogenesis) in week 4 of gestation which is usually completed in week 36.
Postnatally, the kidneys will continue to mature and are responsible for the filtration of blood and subsequent reabsorption of water and other nutrients according to what the body needs. Filtration is carried out in a special compartment called the nephron and in humans, the number of nephrons in each kidney ranges from 200,000 to 2.5 million.[1]
It is quite common to experience developmental abnormalities relating to the kidneys, with about 1 in 500 babies being born with kidney defects[2] such as horseshoe kidney, Wilms' tumor and renal genesis. Research is currently being conducted to see whether nephron number has an impact. Also, there is ongoing research being conducted into possible stem cell therapies.
As mentioned above, the kidneys fall under the renal system however this wiki page is solely dedicated to kidneys. If you would like more information on the renal system, please expand:
| General info on the renal system |
|---|
| The renal system consists of the kidneys, ureters, bladder and urethra. The four organs function collaboratively in the production, storage and excretion of urine, in which liquid waste products from blood is eliminated from the body. Other purposes of a renal system includes the regulation of blood pH, blood volume, blood pressure and the levels of metabolites and electrolytes in blood, A healthy human produces an estimated volume of 800mℓ to 2000mℓ urine daily. The volume of urine produced varies depending on fluid intake and renal function, which is measured by glomerular filtration rate. [3]
Bladder: The urinary bladder stores urine before it is excreted from the body. In human adults, the bladder has the capacity to hold about 300mℓ to 500mℓ of urine. Urine is expelled into the urethra from the urinary bladder via micturition, which involves both voluntary and involuntary muscles. Incontinence refers to a lack of voluntary control over micturition.[5] Urethra: The urethra is a passageway that leads from the urinary bladder to the outside of the body, through which urine is expelled from the body. The length of urethra differs in males and females. Ejaculation of semen in males also passes through the urethra.[6] |
Anatomical Position

Unlike other abdominal organs, kidneys lie retroperitoneally in the abdomen, on either side of the vertebral column. As shown in figure 1, they are typical located between the transverse processes from Thoracic 12 (T12) to Lumbar 3 (L3) of the vertebrae, however the right kidney sits slightly more superior due to the large size of the liver.

Kidney Structure
The kidneys have external coverings which involves complex layers of fascia and fat. From deep to superficial, the layers are as follows:
Renal capsule – Tough fibrous capsule
Perirenal fat – Collection of extra peritoneal fat
Renal fascia – Encloses the kidneys and the suprarenal glands
Pararenal fat – Mainly located on the posterolateral aspect of the kidney.
Their inner structure can be divided into 2 main areas: the outer cortex, and the inner medulla. The extension of the cortex into the medulla gives rise to renal pyramids, with the apex known as renal papilla. Each renal papilla is associated with structures known as minor calyx, which collects urine from the pyramids. Several minor calyces join to form a major calyx, where urine passes before it reaches the renal pelvis and into the ureter.
Kidney development
Kidney development (nephrogenesis) in humans begins in week 4 and commonly ends in week 36 of gestation. This is a brief timeline of the developmental processes.
| Week | Development |
|---|---|
| 4 (early) | Pronephros begins (first stage of nephrogenesis) |
| 4 (late) | Mesonephros begins (second stage of nephrogenesis) |
| 5 | Metanephros begins (third last stage of nephrogenesis)
Development of renal vasculature begins |
| 6 | Ascension of kidneys from pelvis area begins |
| 8 | Functional kidney is formed. First nephrons are formed, a process that continues until week 36 |
| 9 | Kidneys complete their ascension and now sit just below the adrenal glands |
| 10 | Kidneys now ready to perform filtration |
| 15 | Renal vasculature development is completed |
| 36 | Nephrogenesis completed. No more nephrons formed from this point on. |
| Postnatal | Kidney and its structures continue to mature. |
Development of the kidney is called nephrogenesis and it arises from the intermediate mesoderm in the metanephric blastema. Here are the three main stages of nephrogenesis which begins in week 4 of gestation and ends in week 36. It must be noted that whilst nephrogenesis does not continue beyond week 36, maturation of the kidney and its functional units does continue.
Before reading the steps below, it may be helpful to consult this video first.
<html5media height="300" width="500">https://www.youtube.com/watch?v=81yCpyF2VhQ</html5media> [7]
Nephrogenesis
Nephrogenesis involves two transitory stages, pronephros and mesonephros, which end in a final stage (metanephros) giving us the final product of a functional kidney. It is not to say that kidney is ready and functional once metanephros is reached, but rather, it has reached the stage where its structures are set and can continue to mature into a proper functional kidney that can sustain life.

1. Pronephros
The earliest nephric stage in humans (week 4), arising from the intermediate mesoderm near the pharyngeal arches and extend from the 4th to the 14th somites and consists of 6-10 pairs of tubules. These spill into a pair (a 'pair' because there are two kidneys) of primary ducts (nephric or mesonephric duct, also known as the Wolffian duct) that are formed at the same level and go on to extend caudally. The pronephros is a transient structure that disappears completely by the 4th week of human embryonic life, its degradation can be seen in the diagram.
2. Mesonephros
This stage sees the continuation of the nephric duct caudally, with mesonephric tubules arising laterally from it. Together, these structures are known as the mesonephros and whilst still a transient structure, it has important excretory functions during early embryonic life (4—8 weeks). Gradually the top two thirds of the mesonephros go onto form the genitals, however the last third continues to form the functional kidney. This part of the mesonephros that goes on to form the kidney is known as the nephrogenic chord which is essentially mesenchymal tissue.
Towards the end of week 4 of development, the nephrogenic chord starts to move away from the nephric duct as seen in the diagram, whilst the nephric duct continues to grow caudally. NOTE: It is important to understand that the nephrogenic chord and nephrogenic duct are two different things. See Figure 3.
3. Metanephros
At the fifth week of development, a lateral projection called the ureteric bud develops from the nephric duct whilst the nephrogenic chord has now detached itself from the nephric duct to form the metanephric blastema, which has been described as a cloud of mesenchymal cells [8]. After receiving relevant signalling, the metanephric blastema and ureteric bud interact by way of the blastema 'clouding around' the bud. This cloud shapes into the traditional kidney-bean shape and just gets bigger as time goes on.
Inside the metanephric blastema, the ureteric bud bifurcates to form the calyces, pelvis, ureter and collecting tubules of the kidney, where as blastema's mesenchymal cells go on to form numerous vesicles which develop into glomeruli and Bowman's capsules. Their interaction allows for the formation of the nephron. However, as general overview, the ureteric bud differentiates to create the renal tubule section of the kidney (reabsorption compartment of kidney), while the metanephric blastema's mesenchymal cells differentiate to create the renal corpuscle section (filtration compartment).
The permanent and functional kidney is now ready at week 8. Nephrons are still being made until approximately week 36 as they are incredibly important for life and therefore, there needs to be a lot of them.
| Renal Overview |
| Page | Play |
Here is an animation showing the renal development described in this section.
Genes Expressed
Current literature highlights two stages during organogenesis in which signalling molecules play a crucial role in the development of the functional kidney. The first is the outgrowth of the ureteric bud and the second instance being the inductive signal from the ureter that dictates differentiation of the mesenchyme. During these major signalling cascades, there are other more complex mechanisms which serve to ‘fine-tune’ this development through the regulation of cell proliferation, apoptosis, differentiation and motility. Without these growth factors and their respective receptors, outgrowth of the ureteric bud and differentiation of the mesenchyme will become severely affected.
The GDNF (Glial cell derived neurotrophic factor) / RET (REarranged during Transfection) pathway is the primary signalling complex that regulates ureteric bud growth. RET is expressed in the Wolffian duct from E8 to E11.5 and eventually in the ureteric bud as it emerges from the metanephric mesenchyme. During E13.5-17.5 expression of RET is confined to the growing tips of the ureteric bud epithelium. The RET protein acts cell autonomously by receiving a signal from the mesenchyme, that activates the proliferation and branching pathway of the ureteric bud epithelium. Targeted inactivation of RET results in a failure of the ureteric bud to emerge and respond to signals in the metanephric blastema.
Notable genes and their roles:
| Gene | Function |
|---|---|
| BF-2 (transcription factor belonging to the Winged Helix family) | Regulates kidney stromal cell formation[9] |
| Bmp-7 (also known as OP-1) | Regulator of nephron maturation. Plays a role in the differentiation of metanephric mesenchyme.[10] |
| FGF-2 (fibroblast growth factor) | Produced by ureteric bud. Prevents mesenchyme apoptosis and induces regulation of transcription factor WT-1. FGF-2 also plays a role in tubulogenesis.[11] |
| GDNF (gilial cell derived neurotrophic factor) | Partners with RET to form the GDNF/RET pathway which is the primary signally complex that regulates growth of ureteric bud[12] |
| Hox genes | The current understanding is that Hox genes provide the positional information during development.[13] |
| LIF (leukaemia inhibitory factor) | Combines with FGF-2 to form nephrons[14] |
| p53 (a tumor suppressive gene) | p53 is an important regulator of cell growth. In mice, it has been found that taking away p53 does not have any significant developmental effect, however over expression does. Too much p53 can result in a smaller kidney and therefore a smaller number of nephrons also.[15] |
| Wnt-4 (of the Wnt gene family) | Regulator of nephron maturation. Wnt-4 is required for epithelium formation in the metanephros. [16] |
| WT-1 | Important transcription factor that is essential during different stages of renal development. Absence of WT-1 prevents the metanephrogenic mesenchyme from differentiating into nephrons. It also regulates other genes.[17] |
Nephron development
In a mature kidney, nephrons function as a complex epithelial network of blood filtration units which work to remove nitrogenous waste metabolites and regulate homeostasis of water and electrolytes in the body. Mammalian nephrons are generated exclusively during late embryonic and early postnatal development, with very limited cell turnover as opposed to intestine, stomach and skin epithelia which constantly renew throughout and individual's lifetime. [18] Although damaged nephrons are capable of regeneration, extremely damaged nephrons are lost and can not be replaced.
In the kidney, epithelial tubules develop from cell types of distinct embryonic origins using different cellular mechanisms (Little et al., 2010). Both the mesonephric blastema and ureteric bud contribute to the formation of the nephron and its two main units, the glomerulus and Bowman's capsule. The ureteric bud forms as an outgrowth of a pre-existing tubule and undergoes many rounds of branching to form the renal collecting system [19]
| Nephron |
| Page | Play |
Here is a quick animation depicting nephron development. The steps are summarised below.

Stages in nephron formation:
1) Metanephric mesenchyme cells condense into a tight pre tubular aggregate near the ureteric bud.
2) The compacted cells then undergo a mesenchymal-to-epithelial transition, forming a sphere of polarised epithelia
3) The sphere of epithelial cells called the renal vesicle, elongates to form comma shaped bodies and primordial tubule called the 'S' shaped body
4) The proximal slit of the 'S' shaped tubules forms the glomerulus, and the distal pole connects to the tip of an adjacent bud tubule.
The ureteric bud is plays a main role in producing factors that promote mesenchymal survival, as well as secreting proteins that induce condensation of mesenchymal cells and differentiation into nephrons. One of the proteins that are produced by the ureteric bud is FGF-2, which was shown to prevent apoptosis in isolated rat mesenchyme and induce upregulation of the important transcription factor WT-1. Leukaemia inhibitory factor (LIF) combines with FGF-2 to form nephrons within 7 days in culture, and addition of TGFβ made by stromal cells speeds up the process to just 2-3 days. [21]
WT-1 is a zinc finger protein, which is essential at various stages of renal development. It is also implicated to regulate genes such as Pax-2, syndecan, and E-cadherin, which are significant during the later stages on renal development. Absence of WT-1 prevents the metanephrogenic mesenchyme from differentiating into nephrons, leading to apoptosis [22].
Ascension
During metanephros, the ureteric bud forms the ureter. The ureter of each kidney descends from the kidney complex and connects to the urinary bladder. At week 6, the ureters ascend upwards as the torso of the foetus continues to extend. At week 9, they assume their permanent and proper anatomical position under the adrenal glands. During the process of ascension, the kidneys also develop their vasculature as depicted in the animation.

| Renal Vascular |
| Page | Play |
This animation depicts the kidneys during ascension. You can also see vasculature development.
Renal vasculature
Although kidneys are relatively small as compared to other organs, they are responsible for filtering about 20% of the blood output from the heart. [23] A constant and stable blood flow is important for the tissues to carry out respiration, thus ensuring the normal functioning of the kidneys. Renal infarction [24], that is, interruptions to the supply of oxygenated blood to the kidneys may result in kidney failure and subsequently the loss of kidney functions. A person with a damaged kidney is able to survive with the other functional kidney. In the case where both kidneys fail, dialysis or kidney transplant is needed to filter metabolic waste from blood.
Angiogenesis and vasculogenesis
Blood vessels in kidneys are formed via angiogenesis and vasculogenesis. During ascension, the kidneys receive blood from the dorsal aorta. The first endothelial cells in early gestation are formed by vasculogenesis while those in later gestation are formed by both angiogenesis and vasculogenesis. The co-expression of Vascular Endothelial Growth Factor (VEGF) and its receptors (VEGF-R) during kidney organogenesis stimulates the development of renal blood vessels and is important in regulating vascular permeability. High levels of VEGF and VEGF-R are expressed in the kidneys during both embryonic development and adulthood. The high expression of VEGF and VEGF-R by the glomerular endothelium supports the hypothesis that VEGF and VEGF-R play an important role in the regulation of vascular permeability. When VEGF and VEGF-R levels are high, vascular permeability increases. Conversely, when VEGF and VEGF-R levels are low, the blood brain barrier permeability decreases. Hypoxia induces the production of VEGF by glomerular epithelial mass and the expression of VEGF-R by endothelial precursor cells. VEGF and/or anti-VEGF are believed to be therapeutically useful in treating many disorders. [25]
Glomerulus
The glomerulus is a specialised network of blood capillaries that filters metabolic waste products in blood carried via the afferent blood arterioles. Metabolic waste products are filtered through fenestrae, which are small pores with diameter of 50nm to 100nm, on endothelial cells lining the glomerular capillaries. The resultant glomerular filtrate of water and soluble solutes is transported to the Bowman’s capsule and subsequently, to the renal tubule of the nephron to form urine. The glomerular capillaries converge into efferent arterioles in which filtered blood is carried away from the glomerulus. The juxtaglomerular cells lining the walls of the afferent arterioles secrete renin and regulate the volume and pressure of blood flow via the renin-angiotensin system. The efferent arterioles have high resistance that generates hydrostatic pressure that is sufficient for ultrafiltration within the glomerulus. The glomerulus serves as the connection between the vascular system and the nephron. The glomerulus and Bowman’s capsule form the filtration unit of the kidney known as the renal corpuscle. The glomerular filtration rate is the rate at which blood is completely filtered through the glomerulus and is a measure of the renal function. [26]

Juxtaglomerular apparatus
The juxtaglomerular apparatus regulates the renal blood flow (volume and pressure) and glomerular filtration rate. Figure 6 displays the three different types of cells, namely macula densa, juxtaglomerular cells and extraglomerular mesangial.[27]
- Macula densa: The macula densa is a specialized area of the distal convoluted tubule where afferent arterioles enter the glomerulus and the efferent arterioles leave the glomerulus. The macula densa is able to detect changes in the levels of sodium chloride in the distal tubule of the nephron by the tubuloglomerular feedback loop.[28]
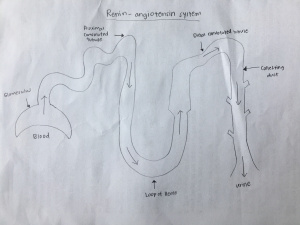 Figure 7. Renin-angiotensin system. Glomerulus, proximal convoluted tubule, loop of Henle, distal convoluted tubule and collecting duct are labelled. This figure shows the flow of blood through the renin-angiotensin system and the resultant filtrate excretion as urine.[29]
Figure 7. Renin-angiotensin system. Glomerulus, proximal convoluted tubule, loop of Henle, distal convoluted tubule and collecting duct are labelled. This figure shows the flow of blood through the renin-angiotensin system and the resultant filtrate excretion as urine.[29]

- Juxtaglomerular cells: Juxtaglomerular cells are responsible for the synthesis, storage and secretion of renin in the kidneys. They line the walls of the afferent arterioles and regulate the volume and pressure of blood flow via the renin-angiotensin system. Juxtaglomerular cells secrete renin in response to (1) stimulation by macula densa cells when the concentration of sodium in tubular fluid decreases, (2) stimulation of β1 adrenergic receptors by epinephrine or norepinephrine and (3) a decrease in renal perfusion pressure. Poorly perfused juxtaglomerular cells activate the renin-angiotensin system.[30]
- Extraglomerular mesangial: The specific function of extraglomerular mesangial is not well understood.
Renin-angiotensin system
A decrease in plasma sodium level leads to the conversion of prorenin into renin by the juxtaglomerular cells. Renin secreted into the blood cleaves a short peptide of 10 amino acids called angiotensin I from plasma protein angiotensinogen. Angiotensin-converting enzyme converts angiotensin I into angiotensin II. Angiotensin II constricts arterioles, thus increasing renal blood pressure. It also stimulates the release of hormone aldosterone from the adrenal cortex, thus increasing the reabsorption of sodium ions into the blood.[31]
Developmental abnormalities
As mentioned in the Introduction, every 1 in 500 newborns suffer from congenital abnormalities of the kidney and urinary tract (CAKUT). Studies have shown that certain CAKUT increases the risks of developing hypertension and cardiovascular diseases at adulthood.
An increase in anti-α smooth muscle actin (α-SMA), vimentin and fibronectin expression in renal tissue [32] [33] as well as a decrease of the proximal tubule cubulin receptor [34] have been associated with disruptions in renal development. The proximal tubule receptor is important as it characterises the epithelial-mesenchymal transition (EMT) process, which is a physiological process that occurs during early embryogenesis, tissue repair, and pathology. [35]
Vimentin and α-SMA are only expressed before the differentiation or transdifferentiation processes in epithelial cells. During this process, cells can proliferate, migrate and produce extracellular matrix. Therefore, these proteins can be utilised as a marker of cell indifferentiation.
Studies have shown that the renin-angiotensis system (RAS) participates in renal development, and that exposure to RAS blockers resulted in the presence of acute kidney injury, chronic kidney disease, and tubular dysfunction in children. These studies demonstrate that inhibition of the RAS causes an increase in the relative interstitial area from the renal cortex, high levels of apoptosis, decreased cell proliferation and impaired expression of growth factors in the kidney. Furthermore, Chen at al.'s treatment of neonatal rats with losartan for 2 days promote down regulation of genes encoding cytoskeletal and extracellular matrix ECM components, which results in ECM malformation and cell-cell and cell-matrix interaction dysfunctions. [36]
Although calcitriol is widely known for its important role in the homeostasis of calcium homeostasis and bone metabolism [37][38], recent studies have shown that it is also involved in the homeostasis of other cellular processes. These processes include the control of of autoimmunity, inflammatory process as well as blood pressure. Furthermore, calcitriol regulates the cell proliferation and differentiation processes as well as the renin gene. [39]
Kidney developmental abnormalities are diverse and they correspond to defects at different stages of the kidney development. Renal vascular anomalies are defects involving renal arteries and renal veins while fusion anomalies results in conditions such as fused pelvic kidney, crossed fused renal ectopia and horseshoe kidney.
Common congenital kidney defects include:
- Horseshoe kidney (fusion of two kidneys during development)
- Wilms tumours (kidney cancer)
- Renal agenesis (absence of one or both kidneys)
- Multiple ureters (more than one ureter draining a kidney)
- Hypoplastic kidneys (underdevelopment of the kidneys)
- Dysplastic kidneys (abnormal development of kidneys which arises from tubules failing to branch out completely)
- Patterning defects

Horseshoe kidney
- Common congenital abnormality of the kidneys.
- The horseshoe kidney can be T-, horseshoe, or L-shaped, depending on the manner of fusion of the two kidneys. It occurs during development, when the left and right kidneys fuse at their lower poles by a parenchymal isthmus located ventral to the abdominal aorta, forming a "U" shape.
- It is usually located in the lower lumbar position (L3 to L5), in front of the aorta and inferior vena cava, posterior to the inferior mesenteric artery [40]. The inferior mesenteric artery blocks the isthmus, preventing the ascension of the kidneys and causing them to remain at a lower position.
- It shows a wide variation of arterial blood supply. [41]
- During migration from the sacral region, the two metanephric blastemas can come into contact, mainly at the lower pole.
- The ureters pass in front of the zone of fusion of the kidneys.
- Despite the abnormality, kidneys and ureters are still able to function. However, there is increased chances of developing upper urinary tract obstruction of infection.
- Horseshoe kidney is typically associated with other congenital defects, including Turners Syndrome, Wilms' Tumour, duplicated ureter, and Trisomy 18.

Wilms' tumour
- Most common intraabdominal cancer in children [42]; 9 out of 10 kidney cancers in children are Wilms' tumours.
- Most Wilms' tumours are unilateral, meaning they occur in one kidney. In some cases, they can occur bilaterally.
- Transcription factor Wilms' Tumour 1 (WT-1) is known as a classic suppressor gene in Wilms' tumour. [43] Although the exact function of WT-1 is still unclear, research suggests that gemline mutations of WT-1 is the main cause for Wilms' Tumour.
- A classic Wilms' tumour has three main cell types (stromal, epithelia, blastomal), giving it a triphasic appearance.

Renal agenesis
- Renal agenesis is a congenital abnormality occurring when there is a failure of development of the kidneys and ureter. It is induced by a lack of interaction between the ureteric bud and the metanephric mesenchyme.
- This defect usually occurs around 5 weeks of embryonic life. [44]
- It can occur in two forms: bilateral or or unilateral renal agenesis.
- Renal agenesis occurs when the ureteric bud fails to become a ureter, the renal pelvis and the collecting ducts and the mesenchyme to form nephrons. [45]
- Renal agenesis patients frequently have extra-renal anomalies, such as cardiac, genital or gastrointestinal malformations. [46]
- It is usually associated other congenital defects including oligohydramnios, as well as facial abnormalities including, wide set eyes and low-set ears, and a broad flat nose.
Animal Models
Most of our current understanding of molecular regulation of kidney development has been derived from either using genetically altered (knock-in) or knock out mice. To obtain knock-out mice, homologous recombination of a gene of interest and a reporter gene like green-fluorescence protein (GFP) or LacZ is used. Using knock-out mice with a reporter gene makes it possible for the researchers to detect and visualize transcriptional control and gene expression of the gene of interest during development. However, during to many cases of embryonic motility when knocking out a gene, conditional knock-out mice are used instead. The most commonly used method to create conditional knock-out mice is the use of Cre/loxP recombination.
Cre-recombinase is an 38kB enzyme, which is produced by bacteriophage P1. It recognizes a 34-bp sequences called loxP and can mediate DNA recombination between two lox sites. Since Cre recombinase is not found in mammalian cells, this makes the technique very specific. [47].
Below we have listed some of the transgenic mouse models which have been used to detect and visualize different kidney structures during development:
| Model | Description |
|---|---|
| Hoxb7/GFP transgenic mice | The model was developed in 1999 by Srinivas et al. [48] and is one of the best known reporter mouse model. In this mouse model, the GFP is under the control of Hoxb7 promotor, and it is used to examine the genetic control of renal branching morphogenesis, therefore allowing visualization of the Wollfian duct, ureteric tree and its derivatives (collecting ducts, calyces, renal pelvis, ureter) [49] |
| Ksp-cadherin (Ksp1.3/BgEGFP) transgenic mouse model | Ksp-cadherin is a tissue-specific cell adhesion molecule which is only expressed in tubular epithelial cells in the kidney and developing genitourinary tract. It is therefore a good model to visualize the ureteric bud, Wolffian duct, Müllerian duct, and developing tubules in the mesonephros and metanephros during development . [50] |
| Sall-1 knock-in mice | This model uses GFP to visualize the Sall1 gene expression, which is expressed in metanephric mesenchyme prior to bud invasion and in cells of mesenchymal origin (glomerular, proximal and distal tubule epithelia). This model has been used together with other techniques to determine which genes are expressed in the metanephric mesenchyme during development. [51] |
Current Research
Can hypertension be associated with nephron endowment?
Nephron endowment is the total number of nephrons an individual has at birth – and only depends on embryonic development. Back in 1988, Doctor Barry Brenner hypothesized that there is an inverse relationship between nephron endowment and the risk of developing hypertension. He suggested that the kidney compensate for a reduction in nephron number by increasing the glomerular surface, which in the long run contributes to hypertension because the sodium and fluid retention is increased. These changes then lead to an increase in arterial and glomerular capillary pressure, glomerular hyper filtration and injury which ultimately increase arterial pressure. [52] . This hypothesis is based on a study he conducted in hypertensive patients and many more have supported Brenner’s hypothesis. These studies have all shown an association between hypertension and low nephron number. However, no correlation has been demonstrated, because most kidney have been obtained from death individuals. So blood pressure and nephron number has not been measured at the same time. [53] Also, the methods to measure nephron number in humans have been complicated, not so accurate and there are not very many of them. Often you would measure the nephron number indirect by measuring kidney mass or volume, or you would do a kidney biopsy even though it only gives an estimate. So, some of the more current research has been to develop an experimental model with reduced nephron number to allow direct testing of Brenners hypothesis and see if there is evidence for this hypothesis [53] One recent study has made a hypothesis, that the use of high blood pressure (BHP2) mice can be used to examine the relationship between nephron endowment and hypertension. [53] BHP2 mice is an old mouse model, and was already produced back in 1974. Back then the selective breed eight common mice strains and the off-spring was screened and selected based on their blood pressure phenotypes. This resulted in hypotensive BPL1 (low blood pressure), hypertensive BPH2 and normotensive BPN3 (normal blood pressure) mice. Therefore this model has hypertensive mice with an control hypotensive line. Much is known about the blood pressure in these mice, however little is known about the nephron number. This model could therefore be used to examine relationship between nephron number and hypertension, because if Brenners hypothesis is true, then the BPH2 mice would have lower nephron number compared to the hypotensive mice [53] . The model could also be used to examine the relationship between nephron number and glomerular diameter and volume, and see if the kidney compensate for low nephron number through this mechanism. The model seems promising to get further evidence that support Brenners hypothesis.
Kidney stem cells
Adult kidney stem cells that are capable of regenerating a kidney-like structure from a single cell in vitro have been identified in rats. The stem cells are able to differentiate into a kidney-like structure in the absence of embryonic primordial cell types, such as metanephric mesenchyme and ureteric bud cells. This suggests that there may be different kidney organogenesis pathways, and that the organogenesis starting from adult kidney stem cells differs from that during embryonic development. It is also reported that instead of single stem cells, cell aggregates are needed to regenerate the kidney-like structure. It is suggested that stem cells possess intrinsic property that allows them to produce the three-dimensional structure of the organ from which they originate, which in this case, kidneys. Although the kidney-like structures lack vascularization and are unable to make urine, they are still useful for in vitro kidney regeneration research as well as for in vitro studies toward alternatives to animal experiments and tailor-made medicines. While further research is needed to investigate the physiological roles of these cells, it is hypothesized that analogous cells in the adult human kidney would be a valuable and potential resource for the regeneration of kidneys in vitro. [54]
Glossary of terms
| Term | Definition |
| Angiogenesis | The formation of new blood vessels from blood vessels that are pre-existing. |
| Fascia | A sheet of connective tissue beneath the skin that is responsible for the attachment, stabilization, enclosure and separation of muscles and other organs. It is primarily collagen. |
| Isthmus | A narrow organ, passage, or piece of tissue connecting two larger parts. |
| Metanephric blastema | Described as a cloud of mesenchymal tissue, its cells migrate and grow in the traditional kidney bean shape.The metanephric blastema gives rise to the glomeruli and Bowman's capsule, structures which interact in order to eventually form the nephron. |
| Mesenchyme/Mesenchymal tissue | A loosely organized, mainly mesodermal embryonic tissue containing unspecialised cells which develops into connective and skeletal tissues, including blood and lymph. |
| Parenchyma | The functional tissue of an organ as distinguished from the connective and supporting tissue. |
| Pharyngeal arch | Forms the structures of the head and neck. Five arches (1, 2, 3, 4 and 6) are formed in humans with four being visible on the embryo. |
| Retroperitoneum | The space between the peritoneum and the posterior abdominal wall that contains especially the kidneys and associated structures, the pancreas, and part of the aorta and inferior vena cava. |
| Ureteric bud | Dorsal outgrowth of the nephric/mesonephric/Wolffian duct which goes on to form the ureters and collecting tubules of the kidneys. |
| Vasculogenesis | The formation of new blood vessels via de novo endothelial cell production. |
References
- ↑ Bertram JF, Douglas-Denton RN, Diouf B, Hughson MD, Hoy WE (2011). Human nephron number: implications for health and disease. Pediatrics Nephrology 9: 1529-1533. doi: 10.1007/s00467-011-1843-8
- ↑ Song R, Yosypiv IV (2011). Genetics of congenital anomalies of the kidney and urinary tract. Pediatric Nephrology 26: 353-364. doi: 10.1007/s00467-010-1629-4
- ↑ Wallace MA (1998). Anatomy and physiology of the kidney. Association of preOperative Registered Nurses Journal 68: 803-820.
- ↑ Simeone C, Zanotelli T, Tralce L, Cancarini G, Tosana M, Da Pozzo GP (1993). Structure, physiology and physiopathology of the ureter. Arch Ital Urol Nefrol Androl 65: 21-25.
- ↑ Drake MJ (2007). The integrative physiology of the bladder. Annals of the Royal College of Surgeons of England 89: 580-585.
- ↑ deGroat WC, Booth AM (1980). Physiology of the urinary bladder and urethra. Annals of Internal Medicine 92: 312-315.
- ↑ Embryology of the Kidney (Easy to Understand). (2016). Retrieved October 26, 2017, from https://www.youtube.com/watch?v=Fir-v6EoZNE
- ↑ Costantini F, Kopan R (2010). Patterning a complex organ: branching morphogenesis and nephron segmentation in kidney development. Science Direct 18: 698-712. doi: 10.1016/j.devcel.2010.04.008
- ↑ Hatini V, Huh SO, Herzlinger D, Soares VC, Lai E (1996). Essential role of stromal mesenchyme in kidney morphogenesis revealed by targeted disruption of Winged Helix transcription factor BF-2. Genes & Development 10: 1467–1478.
- ↑ Stark K, Vainio S, Vassileva G, McMahon AP (1994). Epithelial transformation of metanephric mesenchyme in the developing kidney regulated by Wnt-4. Nature 372: 679–683.
- ↑ Barasch J, Yang J, Ware CB, Taga T, Yoshida K, Erdjument-Bromage H, Tempst P, Parravicini E, Malach S, Aranoff T, Oliver JA (1999) Mesenchymal to epithelial conversion in ratmetanephros is induced by LIF. Cell 99: 377–386.
- ↑ Pachnis V, Mankoo B, Costantini F (1993). Expression of the c-ret proto-oncogene during mouse embryogenesis. Development 119: 1005–1017.
- ↑ Krumlauf R (1994). Hox genes and vertebrate development. Cell 78: 191–201.
- ↑ Barasch J, Yang J, Ware CB, Taga T, Yoshida K, Erdjument-Bromage H, Tempst P, Parravicini E, Malach S, Aranoff T, Oliver JA (1999) Mesenchymal to epithelial conversion in ratmetanephros is induced by LIF. Cell 99: 377–386.
- ↑ Godley LA, Kop JB, Eckhaus M, Paglino JJ, Owens J, Varmus HE (1996). Wild-type-p53 transgenic mice exhibit altered differentiation of the ureteric bud and possess small kidneys. Genes Development 10: 836–850.
- ↑ Stark K, Vainio S, Vassileva G, McMahon AP (1994). Epithelial transformation of metanephric mesenchyme in the developing kidney regulated by Wnt-4. Nature 372: 679–683.
- ↑ Kreidberg JA, Sariola H, Loring JM, Maeda M, Pelletier J, Housman D, Jaenisch R (1993). WT-1 is required for early kidney development. Cell 74: 679–691
- ↑ Barker N, Bartfeld S, Clevers H (2010). Tissue-resident adult stem cell populations of rapidly self-renewing organs. Cell Stem Cell 7: 656–670.
- ↑ Cebrián C, Borodo K, Charles N, Herzlinger DA (2004). Morphometric index of the developing murine kidney. Developmental Dynamics 231: 601–608.
- ↑ Schell C, Wanner N, Huber TB (2014). Glomerular development – shaping the multi-cellular filtration unit. Seminars in Cell and Developmental Biology 36: 39-49. doi: 10.1016
- ↑ Barasch J, Yang J, Ware CB, Taga T, Yoshida K, Erdjument-Bromage H, Tempst P, Parravicini E, Malach S, Aranoff T, Oliver JA (1999) Mesenchymal to epithelial conversion in ratmetanephros is induced by LIF. Cell 99: 377–386.
- ↑ Kreidberg JA, Sariola H, Loring JM, Maeda M, Pelletier J, Housman D, Jaenisch R (1993). WT-1 is required for early kidney development. Cell 74: 679–691
- ↑ Blakemore C, Jennett S (2001). Kidneys. The Oxford Companion to the Body.
- ↑ Saeed K (2012). Renal infarction. International Journal of Nephrology and Renovascular Disease 5: 119-123.
- ↑ Del PF, Mariotti A, Ilardi M, Messina FR, Afeltra A, Amoroso A (1999). Kidney vasculogenesis and angiogenesis: role of vascular endothelial growth factor. European Review for Medical and Pharmacological Sciences 3: 149-153.
- ↑ Pollak MR, Quaggin SE, Hoenig MP, Dworkin LD (2014). The glomerulus: the sphere of influence. Clinical Journal of the American Society of Nephrology 9: 1461-1469. doi: 10.2215/CJN.09400913.
- ↑ Barajas L (1979). Anatomy of the juxtaglomerular apparatus. American Journal of Physiology 237: 333-343.
- ↑ Persson AE, Ollerstam A, Liu R, Brown R (2004). Mechanisms for macula densa cell release of renin. Acta Physiologica Scandinavica 181: 471-474.
- ↑ Roman RJ, Fan F, Zhuo JL (2016). Internal renin-angiotensin system. Journal of the American Heart Association 67: 831-833. doi: 10.1161/HYPERTENSIONAHA.116.07103
- ↑ Ito S (1997). Kidney and hypertension: role of the juxtaglomerular apparatus. The Tohoku Journal of Experimental Medicine 181: 411-429.
- ↑ Fyhrquist F, Saijonmaa O (2008). Renin-angiotensin system revisited. Journal of Internal Medicine 264: 224-236.
- ↑ Balbi APC, Marin ECS, Francescato HDC, Costa RS, Coimbra TM (2008). MAPK and angiotensin II receptor in kidney of newborn rats from losartan-treated dams. Pediatric Nephrology 23: 1433–1444. doi: 10.1007/s00467-008-0830-1.
- ↑ Lamouille S, Jian XJ, Derynck R (2014). Molecular mechanisms of epithelial–mesenchymal transition. Nature Reviews Molecular Cell Biology 3: 178–196. doi: 10.1038/nrm3758.
- ↑ Lima WR, Parreira KS, Devuyst O, Caplanusi A, N'kuli F, Marien B, Van DSP, Alves PM, Verroust P, Christensen EI, Terzi F, Matter K, Balda MS, Pierre CE, Courtoy PJ (2010). ZONAB promotes proliferation and represses differentiation of proximal tubule epithelial cells. Journal of the American Society of Nephrology 3: 478–488. doi: 10.1681/ASN.2009070698
- ↑ Kalluri R, Weinberg RA (2009). The basics of epithelial-mesenchymal transition. Journal of Clinical Investigation 6: 1420–1428. doi: 10.1172/JCI39104
- ↑ Chen Y, Lasaitiene D, Gabrielsson BG, Carlsson LM, Billig H, Carlsson B, Marcussen N, Sun XF, Friberg P (2004). Neonatal losartan treatment suppresses renal expression of molecules involved in cell-cell and cell-matrix interactions. Journals of the American Society of Nephrology 5:1232–1243. doi: 10.1097/01.ASN.0000123690.75029.3F
- ↑ Holick MF (2013). Vitamin D, sunlight and cancer connection. Anti-Cancer Agents in Medicinal Chemistry 13: 70–82. doi: 10.2174/187152013804487308
- ↑ DeLuca HF (2004). Overview of general physiologic features and functions of vitamin D. The American Journal of Clinical Nutrition 80: 1689–96
- ↑ Li YC, Kong J, Wei M, Chen ZF, Liu SQ, Cao LP (2002). 1,25-Dihydroxyvitamin D3 is a negative endocrine regulator of the renin-angiotensin system. Journal of Clinical Investigation 2: 229–238. doi: 10.1172/JCI0215219
- ↑ Moore KL (1992).Clinically Oriented Anatomy, ed 3: p223
- ↑ Yoshinaga K, Kodama K, Tanii İ, Toshimori K (2002). Morphological study of a horseshoe kidney with special reference to the vascular system. Anatomical Science International 77: 134–139
- ↑ Davidoff AM (2012). Wilms Tumor. Advances in Pediatrics 59: 247–267. doi: 10.1016/j.yapd.2012.04.001
- ↑ Call KM, Glaser T, Ito CY, Buckler AJ, Pelletier J, Haber DA, Rose EA, Kral A, Yeger H, Lewis WH (1990). Isolation and characterization of a zinc finger polypeptide gene at the human chromosome 11 Wilms' tumor locus. Cell 60: 509–520.
- ↑ Robson WL, Leung AK, Rogers RC (1995). Unilateral renal agencies. Advances in Pediatrics 42: 575-592.
- ↑ Kerecuk L, Schreuder MF, Woolf AS (2008). Renal tract malformations: perspectives for nephrologists. Nature Clinical Practice Nephrology 4: 312-325
- ↑ Dursun H, Bayazit AK, Buyukcelik M, Soran M, Noyan A, Anarat A (2005). Associated anomalies in children with congenital solitary functioning kidney. Pediatrics Surgery International 21: 456-459.
- ↑ Sauer B, Henderson N (1988). Site-specific DNA recombination in mammalian cells by the Cre recombinase of bacteriophage P1. Proceedings of the National Academy of Sciences 85: 5166-5170.
- ↑ Srinivas S, Goldberg MR, Watanabe T, D'Agati V, al-Awqati Q, Costantini F (1999). Expression of green fluorescent protein in the ureteric bud of transgenic mice: a new tool for the analysis of ureteric bud morphogenesis. Developmental Genetics 24: 241-251.
- ↑ <pubmed>2880702</pubmed>
- ↑ <pubmed> 12089378 </pubmed>
- ↑ <pubmed>15172686 </pubmed>
- ↑ <pubmed> 3063284 </pubmed>
- ↑ 53.0 53.1 53.2 53.3 <pubmed>28818273</pubmed>
- ↑ Kitamura S, Sakurai H, Makino H (2015). Single adult kidney stem/ progenitor cells reconstitute three-dimensional nephron structures in vitro. Stem Cells Journals 33: 774-784. doi: 10.1002/stem.1891