Paper - Observations on the neural crest of a ten-somite human embryo (1939)
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Baxter JS. and Boyd JD. Observations on the neural crest of a ten-somite human embryo. (1939) J Anat. 73: 318–326. PMID 17104759
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
Observations on The Neural Crest of a Ten-Somite Human Embryo
By J. S. Baxter and J. D. Boyd
In the course of a study of a ten-somite human embryo in a good state of preservation we observed an unusual arrangement of those neural crest cells which form the acoustico-facial primordium. This led us to investigate the arrangement and distribution of the whole of the neural crest, and, as there are few detailed accounts of the neural crest in human somite embryos, we present our results as a separate contribution.
The embryo was obtained by laparotomy from a case of tubal gestation. The clinical history suggests that, if ovulation occurred about the mid-point of the menstrual cycle, the conceptional age of the embryo is approximately 28 days.[1] When received in the laboratory the intact chorionic vesicle had been fixed in formalin (of unknown strength) for several days. In its longest axis the vesicle measured 14 mm., in its shortest 12 mm. Having been dissected from the main part of the chorion, the embryo, with the connecting stalk and the adjacent chorionic wall, was stained in bulk with alum cochineal, dehydrated by the drop method, imbedded in paraffin and cut transversely at Sp. into a complete series of sections. The sections were then stained on the slide with Orange G.
A number of reconstructions of this embryo have been made. Those of interest for the purpose of this description are a wax-plate model of the external form of the embryo, anda millboard-wax model (according to the method of Green, 1937), of the cranial portion of the nervous system and its associated neural crest primordia.
The sectioned embryo, and the reconstructions, show that the neural folds have united from a point somewhat cranial to the otic placodes to the level of the seventh somite. Here the closure of the neural tube is discontinuous, and the caudal extremity of the neural plate is a thickened, flattened area of columnar cells continuous laterally with the somatic ectoderm. The appearance of this caudal portion of the nervous system is very similar to that presented by other human embryos at a comparable stage of development. The cranial extremities of the neural folds, however, are not widely separated and thus contrast with the condition usually found at this stage. The anterior neuropore is, then, a narrow, deep cleft (Pl. I, figs. 1, 2); and, in our opinion, it is quite possible that this appearance may be ascribed to undue shrinkage of the embryo during fixation. There is a certain degree of asymmetry in the state of development of the two sides of the neural primordium, the right side of the central nervous system being more advanced than the left. This asymmetry has been described in a number of human somite embryos (cp. Bartelmez & Evans, 1926).
There are three regions in our embryo in which neural crest cells are found in relation with the developing nervous system (see Text-fig. 1). The most anterior of these regions is at the level where the cranial flexure is most pronounced, and here, on either side of the open neural folds, there is a discrete mass of neural crest cells. The right one is better developed and will be described first.

Text-fig. 1. Geometric projection of the right profile view of a reconstruction of the cephalic portion of the nervous system. The various primordia of the neural crest are shown. The hatched line indicates the margin of the left neural fold. The arrow shows the cranial limit of the first somite.
It is, obviously, the crista neumlis rostralis of Bartelmez & Evans, or the craniale Kopfganglienleiste of Veit (1919). Although, caudally, there is a small isolated portion of this neural crest material, we do not think that a distinct
subdivision into a “pars optica” and a “pars trigeminalis” is present. The neural crest cells do, however, extend forwards to the dorsal edge of the optic sulcus and, possibly, the aggregation of cells should be regarded as a compound of a crista neuralis rostmlis (associated with the trigeminal area and the Gasserian ganglion) and a crista neuralis prosencephali (from the dorsal edge of
the optic primordium). If this interpretation is correct then the term primordium opto-cristale of Bartelmez & Evans might be used for the anterior part of
this portion of the neural crest. We incline to the opinion, however, which has
been expressed by Adelmann (1925), that the crista neuralis prosencephali
should be regarded with extreme caution. Adelmann was unable to find any
trace of such a portion of the neural crest in a large number of rat embryos of
the somite stage; he states (p. 52), “ absence of such a neural crest from at least
a major portion of the forebrain seems to be a general condition”. Schulte &
Tilney (1915) were also unable to find W neural crest cells arising from the
prosencephalon during the early development of the central nervous system of
the cat. Indeed, these investigators were so impressed by the absence of neural
crest cells in the region of the forebrain that they suggested the retention of
these cells in the wall of the neural tube to form an elementiadditional to the
alar and basal laminae. We are unable to find any evidence for such a retention
of alleged neural crest cells in the anterior portions of the neural plates, but we
have the impression that the cephalic portion of our crista neuralis rostralis is
not of prosencephalic origin but is an extension forwards of the main part of
the mass which is related primarily to the anterior part of the rhombencephalon
and the mesencephalon. It is, probably, that portion of the crest which will be
concerned with the development of the ophthalmic division of the trigeminal
nerve.
The cells constituting this right anterior neural crest mass do not show
continuity with the neural plate or the somatic ectoderm. In this character
(which may be due to the same shrinkage process which has caused the approximation of the two halves of the open neural plate in this region) they
differ from the corresponding cells in the ten-somite embryo described by
Corner (1929), for he both described (p. 90), and figured, cells passing to this
mass from “the inner surface of the neural epithelium and the neuro-somatic
junction” of the entire midbrain region and the rostral part of the rhombencephalon. We have been unable to find a trigeminal placode in our embryo so the
question of a contribution from this source to the neural crest does not arise.
There are distinct differences between the neural crest cells and the
surrounding mesenchyme, but it is impossible to be definite in the identification
of the cells in the transition zone between the two groups. The bulk cochineal
stain is, as in Corner’s specimen,‘ not well adapted to show cytoplasmic
differences. We feel quite certain, however, that the neural crest material
extends towards, and merges with, the mesenchyme of the first branchial arch,
particularly with that portion of the arch which will later form the maxillary
process. The existence of this mingling of neural crest cells with the surrounding mesenchyme does not, of course, constitute evidence for the derivation of mesenchyme from neural crest cells. On this point our material does not allow of any definite statement and, indeed, we doubt the value of any contributions to this problem which are based on description alone, particularly on the study of a single embryo or a very small series of embryos.
On the left side of the presumptive midbrain region there is a mass of
neural crest cells which in position and relations is similar to the right crista
neuralis rostralis (Pl. I, figs. 1, 2). It is, however, very much smaller than the
corresponding mass of the right side. We correlate this difference in size with
the relatively retarded development of the left side of the central nervous
system in the head end of this embryo. As on the right side, there is no
evidence of the migration of neural crest cells from the edge of the neural plate.
The smaller size of this left crista neuralis rostralis probably accounts for its
better delimitation and the absence of an appearance, on this side of the
embryo, of an extension of the crest material into the branchial mesenchyme.
Caudal to these two masses of neural crest cells there is a region of the
developing nervous system which has no such cells related to it. A short
distance behind the anterior limit of fusion of the neural folds there is found
the next aggregation of neural crest cells (Pl. I, figs. 1, 2). This portion consists
of a large mass of cells situated dorsal to the neural tube and intimately connected with it (see Pl. I, fig. 3 and PI. II, fig. 7). The cephalic part of this mass
forms a continuum across the mid-line of the embryo with no trace of subdivision into right and left halves; but, when this mass of neural crest cells is
traced caudally it separates into two, bilaterally symmetrical, horn-like processes which come to lie on either side of the dorsal portion of the neural tube
(Pl. II, fig. 5). This group is the primordium acaustico-facialis (Bartelmez &
Evans), or Veit’s caudale Kopfganglienleiste, and is that portion of the neural
crest associated with the hyoid arch and the acoustico-facial nervous complex.
It is situated, as the figures show, in close relation with the otic placodes, but
there is no appearance which would suggest a contribution of placodal cells to
the crest material. The neural crest cells can easily be differentiated from the
cells of the adjacent neural tube, but most of the sections through this region
show an appearance as if crest cells are being budded off from the line of
apposition, in the median plane, of the two halves of the neural plate. The
appearance, in fact, suggests that complete dorsal fusion of the neural folds
has not yet taken place (see Pl. II, fig. 7), although they are completely separated from the overlying, continuous, somatic ectoderm by the relatively huge
mass of neural crest cells. This acoustico-facial primordium is so large that it
produces a definite protuberance upon the dorsal aspect of the embryo.
The form taken by the acoustico-facial neural crest mass in this embryo is
very different from that seen in Corner’s ten-somite human embryo (cp.
Text-fig. 2B). In his specimen the anterior limit of fusion of the neural folds
has not extended so far forward as in our embryo. It reaches only to the level
of the acoustico-facial neural crest primordium which, here, is represented by
bilateral and entirely separate cell aggregations; these, according to Corner’s
description and microphotographs, seem to stream out of the angle of junction of the neural and somatic ectoderm. In some regions he found a sharp demarcation of the neural crest cells from the adjacent mesenchyme; in other regions
it was impossible to draw a line between the lateral margin of the crest and the
mesodermal elements. In our embryo we find no difficulty in distinguishing
between the cells of the acoustico-facial primordium and the surrounding,
frankly mesodermal, elements. Hence there is no appearance of continuity
between the crest material and the hyoid, or other, branchial mesodermal condensations. Other human embryos of the somite period in which the neural
crests have been studied seem to correspond with Corner’s description of the
arrangement of the acoustico-facial neural crest elements. Thus in Payne’s
(1925) embryo of seven somites (Text-fig. 2A), and Veit’s embryo of eight
somites, there are two, quite separate, acoustico-facial primordia; and Heuser
(1930), in his fourteen-somite specimen, describes bilateral acoustico-facial
primordia in an embryo with complete closure of the neural tube in the
definitive hindbrain region (Text-fig. 2C).

Text-fig. 2. Diagram to show the arrangement of the acoustico-facial neural crest primordia in certain human embryos. A. Payne embryo of seven somites. B. Corner embryo (ten somites). C. Heuser embryo (fourteen somites). D. Baxter-Boyd embryo (ten somites). Ectoderm, neural folds and tube indicated in solid black; acoustico-facial neural crest material stippled; endoderm and dorsal aortae outlined. (For explanation see text.)
There is, then, a marked difference between the acoustico-facial neural crest primordium in our embryo and the same region in comparable human embryos. Our specimen may, of course, be pathological; tubal gestations are notoriously prone to show abnormalities of development. We are inclined, however, to regard it as showing no more than a variation of the usual neural crest cell pattern, for, if further development had occurred, it is quite possible that the mid-line mass would have separated into the typical, bilateral, acoustico-facial primordia. Indeed in its caudal portion there is already present such a separation (Pl. II, fig. 5) and the appearance here is very similar to that shown by other human somite embryos. The neural crest is primarily a derivative of the lateral part of the neural plate and whether it arises by proliferation from the lateral margins of the neural folds or from the roof of the closed neural tube is dependent upon the time at which the neural folds fuse dorsally. With delayed closure the tendency will be for the crest cells to arise from the edges of the neural folds and bilateral primordia will result. When closure is early the cells will take their origin from the dorsal portion of the fused folds and will, therefore, form a temporary mid-line mass of neural crest material. Lenhossek (1891) described both these modes of origin in the human embryo. Since the crest cells arise from the junctional zone between the somatic and neural ectoderm it is not surprising that, for a short time, immediately after closure of the neural tube, some crest cells are found dorsal to the line of closure. Raven (1932) has published figures of the development of the neural crests in the Axolotl which show a mid-line arrangement of a considerable extent of the crest material dorsal to the closed neural tube and a number of other workers (cp. Brachet, 1935) have drawn attention to this temporary arrangement in other vertebrates.[2] In our embryo, however, cells belonging to the neural crest are found in the dorsal mid-line only at the level of the acoustico-facial primordium; none are found at more caudal levels although closure of the neural tube has occurred as far back as the seventh somite. We conclude, therefore, that in this specimen the cause of the unusual arrangement of the acoustico-facial neural crest cells was either precocious closure of the neural tube at this level only, or a delay in the time of appearance of the neural crest cells. As other ten-somite human embryos show a less degree of closure of the neural tube than our embryo, we think the former explanation to be more correct; and if this be accepted, then the variation is easily understandable and is of no morphological significance, involving, merely, a time relationship.
We have been unable to find any evidence in our embryo for a contribution
to the acoustico-facial neural crest primordium from the otic placode. The
histological state of the material, however, and the fact that we are dealing
with a single embryo prevents us from making a significant statement" on this
possibility. We use the term acoustico-facial primordium as it is the customary
one employed in the literature to designate this portion of the neural crest.
Prof. J. P. Hill (personal communication) has expressed the opinion that it
shouldibe called the facial primordium, as he regards the acoustic ganglion to
be largely, if not entirely, of placodal origin. This opinion is supported by
Brachet (1907), Landacre (1921) and Knoulf (1927), all of whom worked on
amphibian material, and, more recently by van Campenhout (1935), who
worked on the pig embryo. On the other hand, Adelmann (1925) states that
the VIIIth ganglion is a derivative of the common acoustico-facial mass, owing its origin entirely to neural crest proliferation. Our embryo is too young to permit of a definite statement on this point, but obviously if the acoustico-facial complex is partly of crest and partly of placodal origin the term acoustico-facial primordium is still justifiable, whereas if the VIIIth ganglion is completely a derivative of the placode then facial primordium would be a more
suitable term for this part of the neural crest.
Caudal to the single mass of acoustico-facial neural crest cells there is a small portion of the neural tube which has no crest cells related to it. The next
subdivision of the neural crest takes the form of two masses of cells, one on
either side of the pre-somite region of the developing hindbrain (Text-fig. 1).
These masses are about equal in size; each is related to the dorso-lateral aspect
of its own half of the neural tube, but there is no appearance of continuity of
the crest cells across the middle line. Corner, in his description of his ten-somite embryo, uses the term “occipito-spinal neural crest” for all the crest
material found behind the acoustico-facial primordium. We think, however,
that this group of crest cells posterior to the acoustico-facial mass and anterior
to the region of the somites should be distinguished as a separate portion and should be called, as Bartelmez & Evans designate it, the crista neuralis post-otica. It is that portion of the crest which is related to the post-hyoidean
visceral arches and which is concerned in the development of the glossopharyngeal and vagus nerves. The surface ectoderm of our embryo at this level shows, on either side dorsally, definite thickenings, and although they are not so circumscribed as those described by Corner, we presume that they correspond to his “ Discus IX and X ”. We are unable to find any trace of a contribution, at this stage, to the crest cells from these areas of thickened ectoderm.
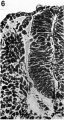
Pl. II, fig. 6
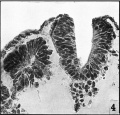
Pl. I, fig. 4


Behind the post-otic neural crest is found what we regard as the true occipito-spinal portion of the crest (Pl. II, fig. 6). This extends, on either side, from the region of the first somite, where it is more or less continuous with the crust post-otica, to about the level of the sixth somite. It is not a continuous column as it shows frequent small gaps. The gaps, however, do not have a segmental arrangement and there is no evidence of neural crest cells being more, or less, opposite to the somites, or the intersomitic spaces. At no point are the cells very numerous and, while there is no definite evidence for their derivation from the neural tube, the histological appearances are often very suggestive of a connexion between the crest cells and the dorsal portion of the closed tube. Caudal to the sixth somite there is no trace of a neural crest (Pl. I, fig. 4) the angle between the neural fold and the somatic ectoderm being completely free of cells. In this hinder region of the developing nervous system we are unable to find any cells which we can take to be forerunners of neural crest cells and there is no sign of activity in the adjacent ectoderm which would suggest that they may be derived from this source. .
That part of the occipito-spinal neural crest which lies opposite the first three somites is probably concerned with the development of the neurones and
sheath cells which, in later stages, are connected with the hypoglossal and
accessory nerves. Enough is not yet known about the developmental history
of this region for any statement to be made on the precise boundary of this
“occipital” portion of the crest. The importance of establishing an embryo-
logical basis for the interpretation of the accessory nerve and the dorsal root
ganglia of the hypoglossal nerve, however, makes some definition, even one
which is only of topographical significance, essential. In the absence of better
criteria the first three somites appear to be suitable as afirst approximation to
the establishment of precise boundaries for this portion of the neural crest. In
our sections this part of the crest and, indeed, the more caudal portion, shows
no evidence for a continuity with the mesenchyme, the crest cells being always
well-defined groups with no appearance of transition into the surrounding
mesodermal elements. The caudal part of the crest opposite to the fourth,
fifth and sixth somites is presumably the rudiment of the dorsal root ganglia
of the upper cervical region but, as has already been stated, it shows no
evidence of the segmentation so characteristic of later stages. In general
appearance and in the arrangement of its cells this “spina ” portion of the neural crest resembles very closely the “occipital” portion with which it is directly continuous.
Summary
The neural crest primordia of a ten-somite human embryo are described. It is shown that they form an interrupted column of cells on either side of the developing nervous system and reaching from the midbrain region to the level of the sixth somite. In cephalo-caudal sequence the primordia are (1) crista neuralis rostralis (concerned in the development of the trigeminal nerve); (2) crista neuralis acoustico-facialis (facial and, possibly, acoustic nerves); (3) crista neuralis post-otica (glossopharyngeal and vagus nerves); (4) crista neuralis occipitalis (related to the first three somites); (5) crista neuralis spinalis (related to the fourth, fifth and sixth somites). The last three groups of neural crest cells form an irregular continuum.
The crista acoustico-facialis of this embryo is peculiar in that it forms a midline mass which, anteriorly, shows no sign of separation into right and left components. It is suggested that this variation is within the normal range
and is the result of precocious closure of the neural tube in this region.
From a study of this single embryo no definite statement can be made on the precise mode of origin or the fate of the neural crest cells.
- ↑ We are indebted to Dr H. T. Laycock of Addenbrooke’s Hospital, Cambridge, for the opportunity of studying this specimen. Details of the clinical history of the patient from whom the embryo was obtained will be given in a subsequent contribution.
- ↑ When this paper was presented at the Summer Meeting of the Anatomical Society, 1938, Prof. J. P. Hill showed a section through the acoustico-facial crest of a sparrow embryo in which the distribution of the crest cells was very similar to that shown by our human embryo. He kindly drew our attention to the work of Belogolowy (Zur Entwiclclung der K opfnerven der Végel, Moscou, 1910), who figures sections through the neural crest of chick embryos in which there is a mid-line arrangement of the crest cells. Belogolowy writes: “Charakteristisch fiir diesen Nerv (acousticofacialis) ist eine erhalten bleibende Zellgruppe der dersalen Seite des Neuralrohres.” Unlike our specimen, however, the acoustico-facial primordium in the chick appears to be connected as much with the ectoderm as with the neural tube. It would seem that the time relations between closure of the neural folds and dilferentiation of the crest cells dilfer in bird and mammal. Our specimen in this respect resembles the avian pattern, but development has proceeded too far to show the con nexion between the acoustico-facial primordium and the overlying ectoderm which is completely separated from it.
References
ADELMANN, H. B. (1925). J. comp. Neurol. vol. xxxIx, p. 19.
BARTELMEZ, G. W. & EVANS, H. M. (1926). Ccmtr. Ernbryol. Carney. Instn, vol. XVII, no. 362, pp. 1-67.
BRACKET, A. (1907). Arch. Biol., Paris, t. XXIII, p. 165.
(1935). Traité d’Embryologie dea Vertébres. 2nd ed. (edited by Dalcq and Gérard). Masson et Cie, Paris.
CORNER, G. W. (1929). Contr. Embryol. Carnagie Instn vol. xx, no. 394, pp. 81-102.
GREEN, H. L. H. H. (1937). Nature, Lor1d., vol. CXXXIX, p. 759.
HEUSER, C. H. (1930). Contr. Embryol. Carnegie Instn vol. xxn, no. 414, pp. 135-54.
KNoU'1rr, R. A. (1927). J. Comp. Neural. vol. xmv, p. 259.
LANDACRE, F. L. (1921). J. Comp. Neural. vol. xxxIII, p. 1.
v. LENIIossEK, M. (1891). Arch. Anal. Phyeiol., Lpz., S. 1-25.
PAYNE, F. (1925). Contr. Ernbryol. Carnegie Instn . vol. XVI, no. 361, pp. 115-25.
RAVEN, CHR. P. (1932). Arch. Entwlllech. Organ. Bd. cxxv, S. 210.
SCHULTE, H. v. W. & TILNEY, F. (1915). Ann. N.Y. Acad. Sci. vol. XXIV, p. 319.
VAN CAMPENHOUT, E. (1935). Arch. Biol, Paris, t. XLVI, p. 273.
VEIT, O. (1919). Anal. Hefte, vol. 56, p. 305.,
Explanation Of Plates
Plate I

Fig. 1. Reconstruction of the future brain region of the embryo viewed from above and behind. In order better to show the dark masses of neural crest material against the background they have been outlined with white dots. The asymmetry of the neural folds and the corresponding asymmetry of the cristae neuralis rostralis is to be noted. The form of the crista neuralis acoustico-facialis is well seen.
Fig. 2. The same reconstruction viewed from in front; the neural crest primordia are again outlined with a white dotted line. The crista neuralis acoustico-facialis in this view overlaps somewhat the crista neuralis post-otica, but the boundary between the two is indicated.
Fig. 3. Microphotograph of a section through the cranial part of the acoustico-facial primordium of the embryo ( x 250). The single mass of cells of this primordium lying dorsal to the neural tube and fused with it in the midline is to be noted. There is no apparent connexion between this primordium and the overlying somatic ectoderm.
Fig. 4. Microphotograph of a section at the level of the eighth somite ( x 500). Here there are no neural crest cells in the re-entrant angle between the neural and somatic ectoderm. The neural folds at this level are quite separate.

Fig 1

Fig 2
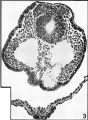
Fig 3
Fig 4



Plate II

Fig. 5. Microphotograph of a section passing through the caudal part of the acoustico-facial primordium ( x 500). Here the primordium is seen as bilaterally symmetrical masses of cells lying dorso-lateral to the neural tube (cp. figs. l and 7). The otic placodes are well shown.
Fig. 6. Microphotograph of section at the level of the first somite ( x 500). Neural crest cells of the occipito-spinal group are to be seen in the angle between the neural tube and the somatic ectoderm.
Fig. 7. Microphotograph of a section through the cranial part of the acoustico-facial neural crest primordium ( x 500). There is an appearance here as if cells are being budded olf dorsally from the region of apposition of the neural folds to join the mass of crest cells. The neural folds are seen not to be completely fused.
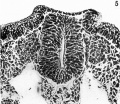
Fig 5
Fig 6
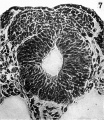
Fig 7


Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - Observations on the neural crest of a ten-somite human embryo (1939). Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_Observations_on_the_neural_crest_of_a_ten-somite_human_embryo_(1939)
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G