Book - Manual of Human Embryology 11C
| Embryology - 2 Aug 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Bardeen CR. XI. Development of the Skeleton and of the Connective Tissues in Keibel F. and Mall FP. Manual of Human Embryology I. (1910) J. B. Lippincott Company, Philadelphia.
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
XI. Development of the Skeleton and of the Connective Tissues

By Embryology History - Charles Bardeen|Charles R. Bardeen]], Madison, Wis.
- Skeleton and Connective Tissues: Connective Tissue Histogenesis | Skeletal Morphogenesis | Chorda Dorsalis | Vertebral Column and Thorax | Limb Skeleton | Skull Hyoid Bone Larynx
C. Vertebral Column and Thorax
The development of the vertebral column and thorax may be divided into three overlapping periods : a membranous or blastemal, a chondrogenous, and an osteogenous.
The Blastemal Period
The division of the axial mesenchyme into segments, sclerotomes, which correspond to the myotomes and spinal ganglia, is marked at an early stage by intersegmental arteries (Fig. 233, A. is.). The segmental differentiation extends into the region dorsal to the spinal cord, but ventrally it does not reach the chorda dorsalis. Each sclerotome becomes divided into two portions, a caudal half in which the tissue is condensed, and a cranial half in which the tissue is less dense (Fig. 234). In sections through hardened tissue a slight fissure, the intersegmental fissure (v. Ebner, 1888), may partially separate the two halves.
From the condensed tissue of the caudal half there arises a primitive vertebra of Remak, or scleromere, with dorsal (neural) and ventral (costal) processes and chordal processes which unite these to the perichordal sheath, a dense layer of tissue forming a continuous sheath about the chorda dorsalis (Figs. 234, 237, 238, 240, 241, 242). From the tissue of the anterior halves of the sclerotomes arise "interdorsal membranes" which unite the dorsal processes of the scleromeres (M. id., Figs. 236, 244, 245, 247), and *-interventral membranes" which unite the bases of the ventral processes (M. ii\, Figs. 235, 243, 244, 245). The chordal processes become hollowed out caudalwards by a loosening up of their tissue and strengthened cranialwards by a condensation of tissue immediately bounding the intervertebral fissure (Figs. 234, 235, 243, 244, 245). There is thus formed about the intervertebral fissure a primitive intervertebral disc. The tissue lying between each two discs now becomes completely surrounded by a membrane of condensed tissue, which may be termed an interdiscal membrane (Fig. 246, M. iv.). Meanwhile the perichordal sheath between each two discs becomes extended ventrodorsally, so that it gives rise to a "perichordal" septum which divides into two parts the space surrounded by the interdiscal membrane (Figs. 239, 246, 247, Pch.s.).
- Schultze (1896) has described in a corresponding position in selachians and reptiles a diverticulum which communicates with the myocoel. The fissure is apparently to be looked upon as an offshoot of the myocoel. In birds the fissure is said to arise independently of and to fuse secondarily with the myocoel. In mammals it appears after the myotome has become independent and the myoccel has disappeared.
|
Fig. 231. Diagrammatic outlines to represent the development of the skeleton during the blastemal period. Fig. 231. Embryo 2, length 7.5 mm. |
Fig. 232. — Fig. 232. Embryo 109, length 11 mm. o - occipital; c1 - first cervical; t1 - first thoracic; l1 - first lumbar; s1 - first sacral; co1 - first coccygeal vertebra. |


Figs. 231 and 232. — Diagrammatic outlines to represent the development of the skeleton during the blastemal period. Fig. 231. Embryo II, length 7.5 mm. Fig. 232. Embryo 109, length 11 mm. o - occipital; c1 - first cervical; t1 - first thoracic; l1 - first lumbar; s1 - first sacral; co1 - first coccygeal vertebra.
length 3.S mm. Fig. 234. Eir 335 and 236. Embryo CoXl.I. Isnglh 6 mm. Fig. 235 through the ( 23S through b more dons) plane. Fig*. 233. 236. 238 represent sections cut somewhat obliquely so that (h« right Bide of ihB Miction. » ventraJ to the left. In Fig. 234 and 236 on the right side the bodie. of several embryonic vertebne are reprewnled in outline. In Figa. 234 and 235, owing to artefncli, the myotomes ore pulled away from the iwlerolometi. ..4.i«., arteria interugmentalia; Cntt., ctelom; Chjl., chorda domlui; Der., dermis; F.v.E.. fissure of v. Ebner (intenertebntl Huure); M.id., membrans interdoTvalis; Af.tc.. membiaLimiiiteTventralia; M.ip.. spinal cord; Myo.. aiyatome: ^.fp.. nervua »pinalia; PiAS., perichontal nheath; Pr.c. processus eoslalis; PrjA., proM9»u« chord»li>; Pr.n.. proee8.«uB Dcuralis; Stl„ sclerotome; V^., vena cardioaLifl.
During the earlier stages of the blastemal period the scleromeres are essentially similar throughout the length of the vertebral column. The differentiation of the scleromeres begins in the cervical region and extends caudalwards. At the end of the first month of development the scleromeres present the appearance shown in Figs. 231, 240, 241, and 242, although their margins are less sharply marked than it is necessary to represent them in the model. At this period of development the interdorsal and interventral membranes have begun to appear in the cervical region, but are not represented in Fig. 231. Soon after this period the thoracic region of the spinal column becomes distinguishable from the neighboring regions through the great development of the costal process of the thoracic scleromeres (Figs. 232 and 239). Meanwhile centres of chondrification arise. These are described below.
The Occipital Region
In man, as pointed out above, the primitive axial mesenchjone in the head posterior to the otic region undergoes a partial segmentation. At the end of the first month of development there are three fairly well-marked occipital myotomes which afterwards disappear. The axial mesenchyme corresponding to these myotomes is not definitely divided into sclerotomes, although that opposite the last occipital myotome becomes divided like each of the spinal sclerotomes into a light anterior and a condensed posterior half (scleromere). The lighter half is continuous apicalwards with the slightly condensed, unsegmented mesenchyme which lies in the region of the more anterior occipital myotomes. This in turn Is continued into a thin layer of dense tissue which is closely applied to the back of the pharynx. The chorda dorsalis surrounded by a perichordal sheath is continued from the spinal region through the centre of the occipital sclerotome and the tissue in front of this into the dense tissue on the back of the pharynx, in which it may be followed to Seessel's pocket. Fig. 231 represents the sclerogenous tissue of the occipital region at the end of the first month. The posterior portion of this, corresponding in form with the first cervical scleromere, is composed of very dense tissue. Anterior to this the tissue is much looser in texture. The occipital scleromere has fairly well developed neural and chordal but has no clearly marked costal processes. No clearly defined interdorsal and interventral membranes are developed from the light half of the first cervical sclerotome. For the subsequent changes which take place in this region see p. 343.
- I have elsewhere (1905) called the united chordal processes of the scleromere a primitive intervertebral disc, but it seems better to restrict this term to the structure here described. according to Williams the primitive intervertebral discs are to be regarded as places in which the tissue remains dense while between them the differentiation of the bodies of the vertebras is marked by a loosening up of the tissue. according to Williams the scleromeres are not true morphological limits.
- Charlotte Miiller (1906) has described a transitory, longitudinal ridge of cells which extends between the mid-ventral surface of the spinal column and the aorta. Opposite the primitive discs this ridge is connected to the anlages of the corresponding ribs by bands of tissue (hypochordal Spangen) which are not fused to the discs. Opposite the vertebral bodies the lighter tissue of the bodies is continued into the lighter tissue of the centre of the ridge of cells. The ridge extended from the second to the ninth thoracic vertebra in a 13 mm. embryo. There is no segmentation visible in the tissue of the ridge.

Figs. 246-247. — (Afwr Banleen, Ainer, Jour. of Anftt., vol. Lv, 1905.) \ie»a of modcla repre
Fig9. 240-242. Embryo tl. length 7 mm. Ma(a.33):l. FiO243-245. Embn'O CLXIII, length . Magn. 23 ; 1. Figs. 246, 247. Embryo CIX, lengtli 11 mm. Magn. 2S : 1. Figa. 240. 243. 246. n from in front: Figi. 241, 244. 247. views from the nde: Figs. 242, 24S, \iewt! from briiind. A.U.. 'rii Inter^menlsUx: CA.(f.. rhiHila dorealis; Diar, mtervert«biAldi«: M.iif., manbrsasiDlenloraalia;
Chondrogenous Period
On each side of the blastemal vertebra three primary centres of chondrification appear at about the same time, one for the neural process, one for the costal process, and one for the vertebral body. Fig. 239 shows these centres as they appear in a cross section at an early period. Figs. 248 and 249 show, the early cartilages of an embryo slightly older (length 14 mm, age five and a half weeks).
The cartilages of the vertebral body are formed by a transformation of the loose tissue lying between the primitive intervertebral discs and surrounded by the interdiscal membrane. At first the cartilage of the left side is separated from that of the right by the perichordal septum. Soon this is broken through and the two cartilaginous anlages of the vertebral body become united about the chorda. In the thoracic region this union seems to take place at about the same time dorsally that it does ventrally. A sagittal section through the thoracic region of an embryo at this stage is shown in Fig. 250. The chorda dorsalis is surrounded by a perichordal sheath. The latter is encircled by dense intervertebral discs which alternate with light cartilaginous bodies, surrounded by perichondrium which is less condensed than the tissue of the discs. Ventrally and dorsally longitudinal ligaments have been differentiated from the surrounding mesenchyme.
according to Schultze (1896), the cartilages of the bodies lengthen at the expense of the discs and finally fuse to form for a brief period a continuous cartilaginous colunm. In human embryos between 20 and 40 mm. in length the discs become very thin near the chorda dorsalis, but the centre of the disc does not become so completely differentiated into embryonic cartilage that it is not possible to distinguish the boundaries between successive vertebral bodies. according to Charlotte Muller (1906), the intervertebral tissue near the chorda so far undergoes chondrification that capsules may be seen about the tissue cells. This stage is quite transitory. At the periphery of the discs the annulus fibrosus is meanwhile differentiated more and more into a condition resembling the adult (Figs. 250, 253, 256, 259).
The chorda dorsalis at the period shown in Fig. 250 is of about the same size at the level of the discs as at the centres of the bodies. It may become slightly swollen in the bodies, but as the bodies increase in size at the expense of the discs the chordal canal becomes enlarged in the intervertebral areas and constricted at the centres of the bodies (Figs. 253, 256, and 259). The chorda loses its continuity and the chordal cells become clumped in the vicinity of the discs, spread out there in the form of a flat disc (Fig. 258), increase in number, and give rise to the nucleus pulposus. Meanwhile the chorda cells lose their cell membranes and form a syncytium similar to that of the mesenchyme. About the cells mucin is formed in considerable amounts (Williams). The chordal canal long remains in the vertebral body (Figs. 256 and 259). The chordal sheath remains in the canal until the period of ossification.

Figs. 248-259. (After Bardeen. Anier. Journ. of Anal,, vol. iv. 1805.) Figs. 248. 249, 251, 252, 254. 255, 257, 258. Ventral, lateral, and doraal views of model made by vertebrae during the chandrogenoui" period. On the left side the cartllaginouii, on the right the enveloping fibrous liwue i* shown. The latter is also shown on the eighth vertebra in Fig. 252. Fign. 248. 249. Embrj'o CXIIV, lengtli 14 mm. M«gn. 17 : 1. Figs. 251. 252. Embryo 22. length 20 mm. Magn. 13:1. Figs. 254. 255. Embryo 145. length 33 mm. Magn.QM. Figs. 2S7, 258. Embryo LXXXIV, length 50 mm. Magn. 9:1. Fig. 257. dorsal view, left half. Fig. 258. median view. Figs. 250, 253, 250. 259. sagittal -enions in the mid-line through the niilh, seventh, and eighth thoracic segments of a series of embryos from 15 to 50 mm. long. Fig. 250. Embryo CXLI\'. length 14 mm. Fig. 25B. Embryo C\'1I1, length 22 mm Fig. 256. Embryo Template:CE97. length 33 mm. Fig. 259. Embryo 184. length 50mm, C.e.. eorpus vertebra; Co«o, rib; CA.rf., chorda dorsalis; Z>t«, inter vertebtal disc; I,., lamina; L.v.. lig. ventrale; pr.a.a.. proc. articulariaant. Isup,); Prji.p., proe. articularia post. {iaf.J; Pr.n.. proc. neumlis; Fr.rd., prae. radicularit; Pr.t.. prnc. spinaliK; PrJr., proc. transversus.
The cartilage of the bodies in embryos of the sixth week (Figs. 248, 249, and 250) is of an early, embryonic, hyaline type. At a slightly later stage (Fig. 253) two regions may be distinguished, a central and a peripheral. The peripheral cartilage resembles that of the preceding stage, while the central cartilage is more dense. Gradually the cartilage at the centre of the body undergoes further changes. The cells enlarge and become sharply set off against the matrix, and finally an invasion of vascular tissue takes place, chiefly from the posterior surface. These changes in the cartilage are preliminary to ossification.
During the development of the vertebral bodies changes have been active in the neural processes. At the period represented in Fig. 239 the neural cartilage is a small, flat body situated in the dorsal process of the blastemal scleromere; from this as a centre, radicular, transverse, cranial (superior) and caudal (inferior) articular, and laminar processes are rapidly developed. This structural differentiation may be followed in Figs. 248, 249, 251, 252, 254, 255, 257, 258.
The cartilaginous radicular processes are at first slender rods which grow out towards and finally fuse with the corresponding vertebral bodies (Figs. 249 and 251). Froriep (1883) has shown that in the chick these processes form a more essential element of the body than in mammals. In the atlas they form a part of the ventral arch, but in the thoracic region of mammals they fuse with the posterolateral portion of the corresponding vertebral bodies.
After their junction with these the radices of the arches increase in size but otherwise show no marked alterations of form. The transverse processes are at first short projections which lie at some distance from the corresponding ribs (Fig. 249). At the time tubercles are developed on the ribs the transverse processes grow outward and forward to meet them (Figs. 252 and 255). At first the developing cartilage of each rib and the corresponding vertebral transverse process are embedded in a continuous blastema, but before chondrification has proceeded far branches from successive intervertebral arteries become anastomosed in the region between the neck of the rib and the transverse process. Fig. 239, Aa, shows the loose tissue through which this arterial anastomosis will take place.
Between the extremity of the cartilaginous transverse process and the rib a joint cavity is developed, and the surrounding blastema is converted into the joint capsule and the costo-transverse ligaments. Similarly a joint is developed between the head of the rib and the corresponding intervertebral disc and vertebral bodies.
The articular processes develop slowly from the cartilage (Figs. 249, 252, and 255). Extension takes place anteriorly (Pr. a a.) and caudalwards {Pr. a.p.) in the interdorsal membranes. In an embryo of 14 mm (Fig. 249) these cartilaginous articular plates are separated by a distinct interval. In one of 17 mm they have approached each other very closely ; and in one of 20 mm. not only do the articular processes show distinctly more form (Fig. 252), but in addition the inferior articular process of one vertebra slightly overlaps the superior process of the vertebra next posterior. This overlapping of the articular processes is distinctly more advanced in an embryo of 28 mm and still more so in one of 33 mm (Fig. 257). In a fetus of 50 mm (Fig. 258) conditions essentially like the adult have been reached.
The laminar processes scarcely exist in an embryo 14 mm long (Fig. 249). In an embryo 20 mm. long (Fig. 252) they have begun to project dorsal to the region of the articular processes. The dense embryonic connective tissue covering the laminar processes at this stage gives attachment to a membrane surrounding the spinal cord, membrana reuniens dorsalis, and to another one covering the dorsal musculature. In a fetus of 33 mm the laminar processes extend well toward the mid-dorsal line (Fig. 255) ; in one of 50 mm. (Figs. 257 and 258) they completely encircle the spinal canal and from the region of fusion of each pair of laminar processes a spinous process extends distally, though not so far as in the adult. The completion of the spinal canal takes place earlier in the thoracic than in the cervical and lumbar regions.
Alterations in the cartilage of the neural processes preliminary to ossification begin at about the time that they take place in the vertebral bodies. They are first seen in centres which correspond to those in which the neural cartilage begins in the blastemal neural processes. In a fetus of 50 mm calcification may be seen in the arches of the first cervical to the sixth thoracic vertebrae. From the blastemal tissue surrounding the cartilaginous vertebrae are developed the various ligaments of the spinal column.
Summary
Each cartilaginous vertebra is developed from four centres of chondrification. In addition a separate centre appears for each rib. In comparing these centres with the blastemal formative centres, we find that each primitive centre of blastemal condensation enters into union with tissue derived from the anterior half of the body-segment next posterior and then gives rise to three centres of chondrification, one for the neural arch, one for the rib, and one for a lateral half of a vertebral body. When ossification first takes place the centres of ossification of the neural arches and the ribs correspond to the original chondrification centres in the blastema, but the centres of ossification of the bodies in most of the vertebrae show little trace of the bilateral origin of the centres of chondrification.
Regional Differentiation, the Thoracic Vertebrae and the Thorax
The chief steps in the development of the thoracic vertebrae have been described in the preceding section. In the blastemal stage these vertebrae became differentiated from the cervical and lumbar by the greater development of the thoracic costal processes. During the chondrogenous stage this difference becomes still more marked. The distal ends of the thoracic costal processes grow rapidly forward (Figs. 260 and 261). The cartilaginous ribs which are differentiated in these blastemal processes do not fuse with the vertebrae, but are connected with them at first by dense tissue and later by joints and ligaments. The blastemal distal ends of the ribs at first take a nearly horizontal direction (Fig. 260), but later their course of development becomes altered by the heart and liver. (See Figs. 261-263.) The distal blastemal ends of the ribs become united by a blastemal tissue to form on each side a sternal plate (Fig. 261). This sternal plate proximally is fused with the anlage of the clavicle and distally extends to the blastema of the seventh rib. The fusion of the tips of the more distal true ribs into the sternal plate does not always take place in regular sequence. The eighth rib is connected near the sternal aniage with the blastema of the seventh and the ninth with that of the eighth rib. the tenth rib becomes similarly attached to the ninth. The attachment of eaoh of the three distal ribs to the one next anterior is apical up to the latter half of the second month. After this it is marginal (Ch. Miiller, 1906).

Fig. 260-263. (After Charlotte Muller, Morpholog. Jahrb., 1906.) The development of the cartilaginous thorax. Fig. 260. Embryo 13 mm long. Fig. 261. Embryo 17 mm long. Fig. 262. Embryo 16 mm long. Fig. 263. Embryo 32 mm long.

Fig. 260. Embryo 13 mm long.
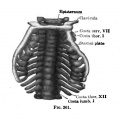
Fig. 261. Embryo 17 mm long.
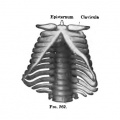
Fig. 262. Embryo 16 mm long.
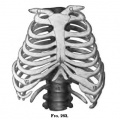
Fig. 263. Embryo 32 mm long.




The sternal ends of the clavicles become united with one another by a dense band of tissue which probably represents the episternum of lower forms (Fig. 261). Later, when the heart has descended into the tlioracic cavity, the cranial ends of the sternal plates become united with one another, and the epistemal band becomes united to them and loses its intimate connection with the clavicles (Fig. 262). It normally disappears before the end of the second month. The sternal bands gradually become fused throughout their length to form the sternal anlage (Fig. 263). Sometimes fusion takes place distally before it is completed in the middle. The ensiform process is formed by the fusion of bands of tissue which extend distally from each sternal plate. The eighth pair of ribs does not enter into its formation (Ch. Muller). Chondrification takes place in the sternal plates at the time these begin to fuse to form the unpaired sternum. according to Ch. Miiller, chondrification extends from the ends of the ribs into the sternal anlage, and the separation of the cartilaginous ribs from the sternum is a secondary process. The order of separation is not always regular. Some investigators assume that special centres of chondrification arise in the sternum.
Literature
The most important papers on the early development of the human sternum are those of G. Ruge, Morphol. Jahrbuch, Bd. 6, 1880; A. M. Paterson, Joum. of Anat. and Physiol., Vol. 35, 1901; and Ch. Muller, Morphol. Jahrb., 1906. Not infrequently the epistemal rudiments, instead of fusing completely with the sternum, become ossified either as separate bones or as bony projections from the upper margin of the sternum. (H. Eggeling, Verb. Anat. Gesellsch., 1903.) Paterson describes a sternal anlage independent of the ribs. The existence of such an anlage is disputed by Ch. Muller. Krawetz (1905) describes in the mouse two sternal anlages which have an origin independent of the ribs.
Cervical Vertebrae and the Base of the Occipital Bone
During the earlier stages of development the cervical vertebrae resemble those of the thoracic region. The two regions soon become differentiated from one another bv the much greater development of the costal processes in the thoracic region (Fig. 232). The seventh cervical vertebra alone, as a rule, has large costal processes and these do not extend far beyond the transverse processes of the neural arches.
In the costal processes of the seventh cervical vertebra centres of chondrification are found at the period when similar centres appear in the ribs. Centres of chondrification in the rest of the^ cervical vertebrae appear much later, usually not until the embryo has reached a length of from 16 to 18 mm. according to Valenti (1906), there are normally no separate centres of chondrification in the costal elements of the cervical vertebrae, but chondrification extends into them from the cartilage of the bodies. There seems, however, good evidence of separate costal centres which arise near and quickly fuse with those of the bodies.
As in the thoracic vertebrae, there are two bilaterally placed centres of chondrification for each of the vertebral bodies. These soon fuse with one another ventral and dorsal to the chorda dorsalis. Tn the first two vertebrae the ventral fusion takes place before the dorsal fusion.
There are separate centres of chondrification for the neural processes. In the more caudally situated cervical vertebrae these centres are similar to those of the thoracic vertebrje. In the more cranially situated cervical vertebrae each neural centre of chondrification appears as a basal plate lateral to the cranial end of the body of the vertebra. With this it soon fuses. From this plate-like base chondrification extends rapidly into the main part of the arch. From the neural arches are developed laminar, articular, and transverse processes. The cartilaginous costal centres become fused medially with the bodies of the vertebrae and laterally with the tips of the transverse processes. The dorsal growth of the laminar processes and the formation of the spinous processes of the cervical vertebrie take place in the main as in the thoracic. When fully formed, however, the cartilaginous cervical vertebrae have essentially the shape of the adult osseous cervical vertebrae. Even before the end of the second month of development distinct cervical characters may be distinguished.

Fig. 264-265. — o the cervinl vertebrae and theoccipiulcftrtilace of embryo


{kind=link}
Specific mention must be made of the mode of development of the epistropheus, of the atlas, and of the basioccipital.
Epistropheus
The general mode of development of the epistropheus is like that of the other cervical vertebras. The dens represents the body of the first cervical vertebra. Union of the body of the first vertebra with that of the second takes place through the transformation of the intervertebral disc into hyaline cartilage, first lateral to the midsagittal plane, then later in this plane. Fig. 265 represents a stage where the lateral fusion is complete while the medial fusion has not yet taken place. The articulations between the superior articular processes of the epistropheus and the lateral masses of the atlas apparently are developed rather in the interventral membranes than in the interdorsal membranes. (See Fig. 264; compare with Figs. 249, 252, 255, 258.)
Atlas
(Figs. 264 and 265) The base (radicular process) of each cartilaginous arch piece of the atlas becomes temporarily fused with the cartilage of the body (14 mm. embryo). This fusion is brought about by incompletely differentiated cartilage, and soon after it takes place the precartilaginous tissue between the arch and the body becomes transformed into a dense blastemal tissue in which ligaments and a joint cavity are later developed.
Meanwhile, during the period of chondrification in the arches and bodies of the cervical vertebrae, there takes place a condensation of tissue on the ventral margin of each of the more proximal cervical intervertebral discs near the cranial end of the vertebral body which lies caudalwards from it (Fig. 266). These condensed transverse bands of tissue connect the ventral ends of the blastemal neural processes with one another. They represent the hypochordal Spangen or braces of Froriep, and may appropriately be called hypochordal arches. In their intimate relations to the intervertebral discs they apparently differ from the hypochordal Spangen described by Charlotte Muller in the thoracic region of the human embryo (see note, p. 334). In man the hypochordal arches are transitory in all except the first cervical segment. In the more distal segments the tissue composing them seems to become merged in the intervertebral discs without going beyond the blastemal stage. In the first cervical segment the hypochordal arch becomes chondrified at the time of the separation of the arches from the body after the temporary fusion mentioned above. The cartilage of the hypochordal arch becomes united on each side to that of the neural hemiarch. There are evidences of two bilaterally placed centres of chondrification in the hypochordal arch but fusion of these centres with one another and with the cartilage of the neural hemiarches takes place as soon as ehondrification is well under way. according to Froriep (1883), in the cow there is a single median centre in the hypochordal arch. In the white rat, according to Weiss, there are two bilaterally placed centres of chondrification in the hypochordal arch of the atlas. Froriep reports in the cow temporary centres of ehondrification in the other cervical hypochordal arches, but no true cartilage formed, except very temporarily in that of the epistropheus. Weiss found no chondrification in any hypochordal arch in the white rat except that of the atlas.[1]

Fig. 266. Sagittal section through the head of an embryo 14 mm long. Arcui AvpaA-. ■rcua hypoehordsliB, bypochordiJ bi»c« or "Spatige"; A, 6o«(., arteri* bluularis; Calv. mrmbr., a^vKi* BtanbrsnMeft; Cart, occ.. oartilBgooocipitaliB; Cart. «pft.. cartilage iphenoidalii; C i. fl, corpus vertfbiw sextK; CA. rf., thorda doraaliB: Lintr., Uogua; (Ewp*., aaophagus; Si^ii. noji. »«ptum nam; Tr., trachea.
According to Sehauinsland (Hertwig's Handbuch, 1906), the neural arches of the mammalian vertebrae contain elements of both the ventral and dorsal arches found in the lower vertebrates, and the ribs belong primitively to the ventral arches. In the mammals and man, however, the presence of ventral arch elements is manifest merely in the caudal and cervical regions. In the caudal region temporary hsemal processes are developed (see p. 352). In the cervical region the ventral arches are represented by the hypochordal braces or arches. In reptiles and birds the hypochordal arches are more extensively developed than in man.
The costal processes of the atlas become fused medially to the basilar part of the neural arches {Fig. 264).
For a brief period (14 mm. embryo) the bases of the neural arches of the atlas and epistropheus together with the tissue intervening between the atlas and occipital bone become fused into a nearly continuous mass of precartilage {Fig. 267).*
Basioccipital
It has already been pointed out that opposite the last occipital myotome the axial mesenchj-me is differentiated, like that of the spinal sclerotomes, into a light anterior half and a dense posterior half. the dense posterior half is called a scleromere. In the spinal region each scleromere joins with the light half of the sclerotome next posterior in giving rise to the body and arch processes of a spinal vertebra. In man the occipital scleromere is not thus associated with the light half of the first spinal sclerotome. On the contrary, it becomes associated with the lighter tissue of its own segment and with the tissue into which this is continued cranialwards. Fig. 266 may serve to illustrate the anterior half of the first spinal sclerotome is composed of light mesenchjTnatous tissue, while the basioecipital and the bodies of the spinal vertebrse are composed of cartilage.
Fig. . Sagittai section through the uieni part ,1' Ti -111 Jill J of the cervical rapan of theepioal coJiuna of the embryo this. It will be noted that sbown in Fig. zee.
Chondrification of the base of the occipital begins in two bilaterally situated centres in the posterior portion of the occipital anlage. The union of these centres takes place caudalwards ventral to the notochord and apiealwards dorsal to the notochord. The neural processes of the caudal part of the occipital anlage seem to have separate centres of chondrification, but these centres fuse almost immediately with the centres of chondrification of the body. Figs. 264 and 265 show the appearance of the occipital cartilage toward the end of the second month of embryonic development. For further details see the subsection on the development of the skull.
"Hagen (1900) gives a aomewhal different account of the development of the atlsB and epistropheus in man. He concludes (1) that the dens epistrophei arisea from the region of the body of the epistropheus and a portion of the body of the atlas, (2) that the massie laterales of the definitave atlas arise from the rest of the primary anlag« of the body of the atlas, and (3) that the short piece which luiites them in front arises from the fusion of both neighboring septa. according to Weiss (1901), the liglit, cranial half of the first spinal sclerotome gives origin to a cartilaginous tip on the dens epistrophei. The caudal part of the occipital Weiss r^ards as arising from the neural processes of the last occipital scleromere. Robin (1864) gives several good pictures of early stages of the cartilaginous cervical vertebrae.
Ligaments and Joints
The atlanto-occipital like the lateral atlanto-epistropheal diarthroses are apparently formed rather in the interventral than in the interdorsal primitive membranes. From the interdorsal membranes between the atlas and the occipital bone arises the membrana atlanto-occipitalis.
From the periphery of the perichordal part of the light anterior half of the first spinal sclerotome are differentiated the cranial extremities of the anterior and posterior longitudinal ligaments, of the tectorial membrane, and of the alar and the crucial ligaments of the atlas. About the chorda dorsalis in this region the lig. apicis dentis is differentiated, probably chiefly from the perichordal tissue.
Cartilaginous and osseous nodules found occasionally in this ligament have been thought by some to represent remnants of the original tissue of the chorda (H. Muller, 1858). Albrecht (1880) advanced the view that these nodules represent the vestige of a supplementary vertebra (pro-atlas), but this view has been disputed by Comet (1888), Chiarugi (1890), and others. Weiss states that in the white rat the perichordal tissue of this region gives rise to the tip of the dens epistrophei, but this appears not to be the case in man. The tissue between the apical ligament of the dens, and the anterior, alar and crucial ligaments of the atlas becomes converted into a fibro-adipose tissue. In this there is ventrally a slight extension of the synovial cavity between the dens and the atlas and dorsally a greater extension of the cavity between the dens and the transverse ligament. The ligaments in the vicinity of the epistropheus are developed from the periphery of the perichordal tissue and from the interdorsal primitive membranes.
Lumbar, Sacral, and Coccygeal Vertebrae
In the earlier stages of development the lumbar, sacral, and coccygeal vertebrae resemble the thoracic. The blastemal vertebrae arise each from the contiguous halves of two primitive segments of the axial mesenchyme. Each vertebra exhibits a body from which neural and costal processes arise. The neural processes are connected by ** interdorsal" membranes. As the blastemal vertebrae become converted into cartilage specific differentiation becomes more and more manifest. The cartilaginous vertebral bodies and the intervertebral discs are all formed in a manner similar to that of the thoracic vertebrae and except for size manifest comparatively slight differences in form. The more distal coccygeal vertebrae are, however, irregular. But the chief specific differentiation is seen in the costal and neural processes.
The development of the vertebrae of the distal half of the vertebral column may be followed in Figs. 27^278 (p. 368).
In the lumbar vertehrce radicular, transverse, articular, and laminar processes arise from the neural cartilages. The radicular processes resemble the thoracic but are thicker; the transverse processes are shorter, much thicker at the base, and remain bound up with the costal processes; the superior articular processes develop in such way as to enfold the inferior articular processes of the vertebra next cranial wards ; the laminar processes are broad, grow more directly backward than do the thoracic, and on meeting their fellows in the mid-dorsal line fuse and give rise to the typical lumbar spines. The mammillary and accessory processes are developed in connection with the dorsal musculature and are not definitely formed in cartilage.
In the sacral vertebrae the neural cartilages give rise to very thick radicular processes; to articular processes, the most cranial of which develop like the lumbar, while the others long retain embryonic characteristics ; to transverse processes which in development are bound up with the costal processes; and to laminar processes which are very slow to develop and of which the last fail to extend far beyond the articular processes.
In the coccygeal vertebrae the neural processes of the first, and rarely of the second, give rise to cartilaginous plates. From these only radicular and incomplete articular and transverse processes arise. The comua of the adult coccyx represent fairly well the form of the embryonic neural semi-arches.
In the thoracic vertebrae cartilaginous ribs develop from separate centres in the blastemal costal processes.
In the lumbar vertebrae separate cartilaginous centres probably also always arise in these processes, but they are developed later than those of the thoracic vertebrae and quickly become fused with the cartilage of the transverse processes. The transverse processes of the adult lumbar vertebrae represent at the base a fusion of embryonic cartilaginous costal and transverse processes, but laterally an ossification of membranous costal processes.
In the sacral vertebrae separate cartilaginous costal centres are developed, but they soon become fused at the base with the transverse processes of the neural cartilages. Laterally by fusion of their extremities the costal processes give rise to that part of the sacrum which articulates with the ilium.
In the coccygeal vertebrae the costal processes of the first vertebra become fused with the transverse processes and develop into the transverse processes of the adult coccyx. It has not been determined whether a separate costal cartilage is developed in these processes or cartilage extends into them from the neural processes. The costal processes of the other coccygeal vertebrae have merely a very transitory blastemal existence.
For a brief period the more distal sacral and the coccygeal vertebrae have membranous hcemal processes. Schumacher (1906) describes a haemal arch on the first coccygeal vertebrae which he considers present in most human embryos 3-5 months old.
Centres of ossification correspond in general with centres of chondrification, but, as in the case of the vertebral bodies and the more distal sacral neuro-costal processes, a single centre of ossification may represent two centres of chondrification.
Period of Ossification
In the vertebrae, ribs, and sternum one may distinguish primary and secondary centres of ossification. Most of the primary centres appear early in intra-uterine life, while the cartilaginous vertebrae and thorax are assuming a definitive form. The secondary centres appear after birth. Vertebrae.
Primary Centres
There are three primary centres, one for the body of the vertebra and one for each hemi-arch (Fig. 268, A). These centres begin to appear at about the same time, but in the cervical region the centres of ossification in the arches appear before the centres in the bodies, while in the thoracic and lumbar regions and, as a rule, in the sacral region, the reverse is true. In the fetuses studied and tabulated by Mall (1906) the first centres found were in the second to the eighth neural arches of a fetus 33 mm. long and about 57 days old. In a fetus 34 mm long and about 58 days old centres of ossification were found in the arches of all the cervical and thoracic vertebrae, and in the bodies from the third thoracic to the first sacral. Centres of ossification in the bodies first appear in the more distal thoracic vertebrae and the first lumbar.
The centres of ossification in the arches extend from the cervical region distally in fairly regular sequence, although not with equal rapidity in different fetuses. In one fetus 53 mm long and about 72 days old they had extended to the third sacral vertebra (twenty-seventh spinal vertebra). On the other hand, no centres were found in the arches of this vertebra in several other fetuses, 73-105 days old. The centres in the arches of the more caudally situated vertebrae arise generally in the fifth or sixth month.
The centres of ossification in the bodies of the vertebrae extend cranialwards from the thoracic and caudalwards from the lumbar region. In a fetus 70 mm. long and about 83 days old they had extended on the one hand to the epistropheus and on the other to the last sacral vertebra (Mall). There is considerable variation, however, in the rapidity with which the centres of ossification appear in the bodies of the cervical and sacral vertebrae.
The type of ossification in the bodies is endochondral (see p. 309). The cells at the centre of the body of the vertebra enlarge and become sharply set off against the intercellular substance. Finally an invasion of blood-vessels takes place, chiefly from the dorsal periosteum. Calcium salts are deposited in the cartilage and this is followed by actual ossification in fetuses about three months of age. In the arches the process of ossification is likewise endochondral.

Fig. 268. (After R. Quain, Quain's Anatomy, 10th ed., vol. ii, Pt. 1, Figs. 19, 20, 21. and 22.) Diagrams to illustrate the development of various vertebra. A, fetal vertebra; B, thoracic vertebra of child of two years; C, thoracic vertebra in the seventeenth year; D and E, lumbar vertebra of about same age; F, atlas before birth; G, atlas in first year; H, epistropheus in fetus of seven months; I, epistropheus shortly after birth; K, sacrum before sixth month; L, saonmi at birth; M. sacrum at about twenty-third year; N (after Allen Thomson), first sacral vertebra at fourth or fifth year. C, centre of ossification in the body; C\ centre of ossification in the dens; Co, centre of ossification in costal element; Et epiphyisis; N, centre of ossification in neural arch; V, centre of ossification in ventral arch of atlas.
The centre of ossification in the body of the vertebra gives rise to the greater part of the body of the definitive vertebra. occasionally the centre may arise as or become divided into two bilaterally placed centres, one for each half body. This division may persist in the adult. The centres in the arch give rise to the posterolateral part of the body of the vertebra and to the greater part of the arch with its various processes. It is in the cervical region that the centres in the arch contribute most to the body. At birth the bones arising from each of the centres of ossification of a vertebra are separated from one another by cartilage. During the first year the centres of ossification in the neural arches in most of the vertebrae become united dorsally. This fusion takes place first in the lumbar region. Between the third and sixth years the bony arches become united to the body. This fusion takes place first in the thoracic region. The neurocentral suture lies in a nearly sagittal plane in the' cervical, in an oblique plane in the thoracic, and in a frontal plane in the lumbar region.
- In embryos cleared by the Schultze method the complementary primitive centres described by Rambaud and Renault do not appear. It is probable that they were artificially produced by the methods of preparation employed by Rambaud and Renault (Mall). The primary centres for the neural hemi-arches are single, not double. See, however, the account of the lumbar vertebrsB (p. 354).
Secondary Centres - Epiphyses
Toward the intervertebral discs the bodies of the vertebrae remain long covered by at layer of cartilage. About the seventeenth year a centre of ossification appears in the cartilage on each intervertebral surface. From each of these centres of ossification a thin epiphyseal disc of bone arises (Fig. 268, E). The discs fuse with the body about the twentieth year. The line of suture is visible usually for a year longer.
The tips of the spinous and transverse processes are covered during infancy by cartilage. In this cartilage epiphyseal centres of ossification appear between the sixteenth and the twentieth years and join the osseous arch after the twentieth year (Figs. 268, C and D). Similar secondary centres on the dorsal margins of the superior articular processes and on the costal facettes of the thoracic vertebrae have been described, but are not generally recognized. (See Poirier and Charpy, Traite d'Anatomie, vol. i, p. 342, 1899.)
Cervical Vertebrae
In most of the cervical vertebrae, according to Leboucq (1896), the ventral limb of the transverse process is ossified by ingrowth at one end from the radix, on the other from the tip of the transverse process. In the seventh cervical vertebra frequently, in the sixth occasionally, and in the fifth and fourth rarely, there may arise during the second to the fifth month a separate centre of ossification for the costal element. While this costal element may remain free as a cervical rib, it usually becomes fused with the osseous projections from the radix and the transverse process (Figs. 269, A and B). Except in the seventh cervical vertebra the epiphyses of the spines are usually double.
Atlas
The posterior arch and the lateral masses of the atlas are ossified from two bilaterally placed centres which correspond to the centres of the neural arches of the other vertebrae (Fig. 268, F). In the anterior arch one, or sometimes apparently two, centres of ossification appear during the first year after birth (Fig. 268, G). The dorsal union of the osseous neural arch pieces occurs between the third and fifth years. Often a separate centre of ossification appears in the spinous process before the neural arch pieces are united (Quain). The union of the posterior with the anterior arch occurs between the fifth and ninth years.
Epistropheus
The neural arch and the body are ossified essentially as in the other cervical vertebras except that occasionally there are two bilaterally placed centres in the body. The odontoid process becomes ossified from two bilaterally placed centres which appear in the fourth or fifth fetal month and soon fuse together (Fig. 268, H and I). These centres furnish material for a part of the superior articular processes. Between the fourth and sixth years the odontoid process becomes joined to the body and the radices of the arch, first laterally and then ventrally and dorsally. Between the centre of the base of the odontoid process and the body of the epistropheus a disc of cartilage remains till late in life. The apex of the odontoid process is formed from a separate centre of ossification, which appears in the second year and is joined to the main part of the process about the twelfth year. This apical piece probably represents an epiphysis. There are also said to be rudiments of a cranial epiphysis of the body of the epistropheus, but this statement is not generally accepted. A caudal epiphysis of the body is constant.

Fig. 269. (After T. Dwight, Piereors Human Anatomy, 1907, Fig. 168.) Diagrams illustrating the homology of the costal elements. A, sixth cervical vertebra; B, seventh cervical vertebra; C, fifth thoracic vertebra; D, second lumbar vertebra; E, fifth lumbar vertebra; F, transverse section through sacrum. The costal elements are stippled.
Lumbar Vertebrae
The mammillary processes of the lumbar vertebrae, of the first sacral vertebra (Fawcett, 1907), and of the twelfth thoracic vertebra have special epiphyses, which appear about the time of puberty or a little later and join the rest of the vertebrae after the eighteenth year (Fig. 268, D). Somewhat rarely the costal element of the first lumbar vertebra has a separate centre of ossification, which appears early in fetal life. It may remain free as a lumbar rib. Sometimes in the fifth lumbar vertebra, and very rarely in some of the others, there are found two centres for the arch on each side; one for the radix, transverse process, and superior articular process, the other for the lamina, inferior articular process, and spine. according to Poirier and Charpy (Traite d'Anatomie, 1899), the fifth lumbar vertebra has a special epiphysis for the anterior tubercle of the transverse process.
Sacrum
The usual primary centres are found for each of the five sacral vertebrae, one for the body and one for each neural hemi-arch. In addition there are separate centres for the costal elements of the first three or four vertebrae (Fig. 268, K and L). Sometimes, apparently, costal centres are found merely in the first two sacral vertebrae, sometimes in all five. Changes preliminary to ossification occur both in the bodies and in the neural processes of the sacral vertebrae at a period quickly following their appearance in the lumbar region. Actual ossification in these centres in the more distal vertebrae, as a rule, does not take place until a considerably later period, usually not until the fourth month in the bodies and the fifth or sixth in the arches. The centres in the arches join those of the bodies between the second and sixth years. The more caudally -situated join before the more cranially situated. Union of the laminae with one another takes place from the seventh to the fifteenth years. It takes place first in the more cranially situated vertebrae, frequently does not occur in the fourth and seldom in the fifth. The centres for the costal elements of the first three vertebrae arise usually, according to Posth, in the fifth, sixth, and seventh fetal months respectively. That for the fourth vertebra does not usually arise until tiie third month after birth. There are considerable variations in the time of origin. The costal centres unite with those of the neural arches between the second and fifth years. They unite with the bodies slightly later than with the arches. Rambaud and Renault (1864) describe special centres which are said to arise in the sixth month in the transverse processes. Posth could not confirm this.
In addition to these primary centres there are epiphyseal plates for each body and two for each lateral sacral margin, one for the auricular surface and one for the rough edge distal to this (Fig. 268, M). according to Poirier (Traite, 1899), the auricular epiphyseal plates arise from the fusion of the epiphyses of the transverse processes. Fawcett (1907) describes them as arising from four costal epiphyses belonging to the first two sacral vertebrae. The tuberosities he describes as arising from the costal epiphyses of the third and fourth sacral vertebrae and transverse epiphyses of the fourth and fifth. The epiphyses of the bodies begin to arise about the fifteenth year and those of the auricular plate between the eighteenth and the twentieth years. Epiphyses for each of the tubercles of the spinous processes are described by Rambaud and Renault, 1864, and Fawcett, 1907. Fawcett describes twelve costal epiphyses, eight epiphyses belonging to the transverse processes, two to the mammillary processes and three to the spinous processes.
The sacrum begins to be consolidated about the time of puberty. The costal processes on each side fuse with one another. This is followed by union of the epiphyses with the bodies and by ossification in the intervertebral discs. The process begins caudally and extends in a cranial direction. The bodies of the first and second sacral vertebrae usually become united about the twentyfifth year but the centres of some of the intervertebral discs may persist longer than this. The lateral epiphyseal plates unite about the twenty-fifth year.
For the recent literature on the development of the sacrum see Posth (1897) and Fawcett (1907).
Coccygeal Vertebrae
Ossification in the coccygeal vertebrae usually takes place after birth. Each is ossified from a single centre. The centre for the first vertebra usually appears in the first year but may appear much later, that of the second appears from the fifth to the tenth year, that for the third just before and that for the fourth just after puberty. The three more distal vertebrae usually become united with one another before being joined to the first. This latter union may not occur until the thirtieth year. The first coccygeal vertebrae not infrequently becomes united to the sacrum. In old individuals the whole coccyx is often united by bone to the sacrum, more often in men than in women.
According to some authors, there are two epiphyseal plates for each of the bodies of the first four coccygeal vertebrae and in addition separate centres of ossification for each of the comua of the first vertebrae.* Two centres of ossification for the fifth coccygeal vertebra, one for the body and one for an epiphysis, are also described as arising in the tenth year (Poirier and Charpy, 1899).
Ribs
Ossification begins in the ribs before it does in the vertebrae. Centres appear in the bodies of the sixth and seventh ribs toward the end of the second month and then rapidly come to view in the other ribs. The centre in the first rib usually appears before that in the twelfth. All are usually present by the end of the second month, but that in the twelfth may not appear until later. In two specimens out of 29 fetuses with an estimated age of 55 to 110 days, Mall (1906) found a centre of ossification in the costal element of the seventh cervical vertebra.
The osseous nucleus arises near the angle of each rib and extends rapidly toward the head. At the end of the fourth month the osseous shaft of the rib bears about the same proportional relation to the costal cartilage which it has in the adult. About puberty epiphyseal centres arise, one for the articular surface of the head, one for the articular surface of the tubercle, and one for the non-articular surface of the tubercle. Frequently only one epiphysis seems to arise on the tubercle (Fig. 270). Usually no tubercular epiphysis is found on the eleventh and twelfth ribs. The union of the epiphyses with the shaft takes place after the twentieth year. The epiphysis of the head does not usually join before the twenty-fourth year.
- According to Poirier, the primary centres appear in the fourth or fifth year in the first vertebra, in the sixth to the ninth year in the second, third and fourth. The epiphyseal plates appear from the sixth to the twelfth year.

Fig. 270. (After R. Quain, Quain's Anatomy, 10th ed., vol. ii, Pt. I, Fig. 31.) Diagram to illustrate the epiphyHes of the head and tubercle of one of the mid-thoracic ribs at about the twentieth year. 1 , body ; 2, epiphysis of the head ; 3, that of the tubercle.
The centres of ossification of the ribs are subperiosteal in character.
In the adult the first costal cartilage may become partially or completely covered by a superficial layer of bone. Late in life the other costal cartilages may become thus covered, especially on the superficial surface. This process is more frequent in men than in women (Quain).
Sternum
Ossification in the sternum begins considerably later than in the ribs. The centres of ossification are variable in the time and place of their appearance (Fig. 271, B and D). About the middle of the sixth fetal month a centre usually appears in the manubrium. Often other accessory centres appear (Fig. 271, D). Thus Mayet (1895) in fourteen sternums out of eighteen found one or more accessory centres in the manubrium; in ten instances one extra centre situated caudalwards from the main centre; in four instances two or more accessory centres. In addition there are two epiphyseal centres next the sternoclavicular joints. These fuse with the manubrium between the twenty-fifth and twenty-eighth years. The body of the sternum is usually ossified from five or from seven centres. The segment next the manubrium is usually ossified from a single centre which appears in the seventh month. The next segment may be ossified from a single centre or from a pair of centres. The last two segments most frequently are each ossified from a pair of centres, but may be ossified from a single centre. As a rule, all the centres of ossification except those in the last segment are present at birth and these last appear during the first year after birth. By the sixth year the centres of each pair usually have become fused with one another. Generally the various osseous segments of the body of the sternum become united in the 12-25 years, but lines indicating the boundaries between them remain till late in life. The manubrium and body rarely fuse; according to Gray, in about 6 or 7 per cent, of cases after 60 years of age. There may be a foramen in the sternum due to lack of fusion of a pair of centres of ossification or to failure of a centre of ossification to develop (Fig. 271, E).

Fig. 271. (After Quain. Qudii.
Four times out of twelve Mayet found the bilaterally placed centres of the body fused vertically with those of their own side before the fusion of pairs across the median line had taken place.[2] The ensiform process is ossified from a single centre which appears late, usually not before the sixth year, and rarely transforms the whole process into bone. The centre of ossification arises at the base of the process. The osseous ensiform process is usually united to the body in middle life.
Relative Length of the Different Regions of the Spine during Development
In 1879 Aeby contributed an important paper dealing with the length of the various regions of the spinal column at different ages, with the height of the constituent vertebrae and with the thickness of the intervertebral discs in man. He showed that in young embryos the cervical region is relatively much longer, the limabar much shorter than in the adult. These results have been confirmed by Moser (1889), Ballantyne (1892), and others. It has been shown by Bardeen (1905) that, in embryos during the second and third months of development, if the length of the thoracic region be taken as 100 the length of the cervical region is about 60, the lumbar from 40 to 50, the sacral from 33 to 42.5. In the adult the cervical region has been estimated at from 41.7 to 47.5, the lumbar from about 56.3 to 71.6, the sacroccCoygeal from 61 to 68. (See Ravenel, 1877, Aeby, 1879, Tenchini, 1894, Dwight, 1894 and 1901.)
Curvature of the Spinal Column during Developmen
During the first month of embryonic development the spine acquires a marked ventral flexion (see 2, Fig. 272). From this period until the time of birth the cervico-thoracc-lumbar region of the spine, at first rapidly and then more gradually, becomes straighter (109, 144, 108, 145, 184, Fig. 272). The sacral region also becomes much straightened during the second and third month of embryonic development (109, 144, 108, 145, 184, Fig. 272), but subsequently acquires a second ventral flexion (Ad, Fig. 272). During the latter half of embryonic development there takes place a marked dorsal flexion at the lumbosacral border (184, Fig. 272). After birth and the assimiption of the erect position dorsal flexion takes place in the cervical and the lumbar regions (I and Ad, Fig. 272).
Number of Vertebrae and Regional Differentiation
At the period of greatest development of the caudal extremity of the human embiyo thirty-six vertebne usually are present. This stage is reached in embryos from 8-16 mm in length. occasionally the number of vertebrsB may reach thirtyseven. Beyond the last vertebra the chorda dorsalis extends for some distance distally (Fig. 273).
Regional differentiation, as already pointed out, becomes well marked toward the latter part of the blastemal period. The thoracic region is clearly demarcated by the great development of the costal processes of the thoracic vertebrsB. The sacral region becomes definitely marked when the blastema of the sacrum comes into contact with that of the ilium. according to Rosenberg (1877, 1899, 1906), the costal processes of the seventh cervical and the first lumbar vertebra at the period of chondrification are to be regarded as ribs, so that in subsequent development there is a reduction in the number of thoracic vertebrae. While each costal element of the seventh cervical vertebra has a centre of chondrification like a true rib, and near the body of the vertebra appears enough like a rib to be called a " rudimentary rib," one is not more likely to mistake it for the first rib than one would be to mistake the costal element of the seventh cervical vertebra in the adult for a true rib. This is even more true of the costal element of the first lumbar vertebra. Although this normally probably has a separate centre of chondrification, it has distinct characteristics which sharply demarcate it from the twelfth thoracic rib, characteristics of form as well as of size. (See Figs. 261 and 262, from an article which has been cited by Rosenberg in support of his hypothesis.)
According to Rosenberg, the saemm is composed at first of a more distal set of vertebra than those belonging to it in the normal adult condition; in other words, the iliac attachment of the skeleton of the limb is supposed to advance cranialwards along the spinal column during ontogeny. The studies of Holl (1882), Paterson (1893), Bardeen (1904), and others have shown that the views of Rosenberg do not correspond with the conditions found in the majority of the human embryos and fetuses, at the period under discussion, which have been carefully studied.

Fig. 272. — Diagram to show the ourvature of the spinal oolumn, the proportional lengths of the various regions, the relations of the acetabula to the sacral region, and the direction of the long axis of the femur in a series of embryos and fetuses 7 to 50 mm in length, in an infant and in an adult. Each curved line represents the choida dorsalis of an individual. The cervical, lumbar, and coccygeal regions are represented by the heavy, the thoracic and sacral by the light portions of the line. The approximate position where a line joining the coitres of the two acetabula would cut the median plane is represented at a. For Embryo II, in which the skeleton of the inferior extremity is not yet differentiated, the position of the future acetabula is deduced from Embryo 163, length 9 mm The line passing in each instance from a and terminating in an arrow point represents the long axis of the femur. For Embryo 2 this line is pointed toward the centre of the tip of the limb-bud. From a in each instance a perpendicular is dropped to a line connecting the two extrmnities of the sacral region. The numbers refer to the following embryos and fetuses in the collection of Professor Mall: 2, length 7 mm.; 109. length 11 mm.; 144, length. 14 mm.; 108, length 20 mm.; 145, length 33 mm.; 184, length 50 mm.; I new-born infant; Ad, Adult.
Variation in the number of vertebraB belonging to each of the regions of the spinal column occurs in the embryo as well as in the adult. Bardeen (1904) reaches the conclusion that the frequency of variation in the embryo is probably the same as that in the adult. Before this is definitely decided a much greater number of embryos must be studied than are at present on record. Regional variation in the embryo and fetus must not be confounded with the normal changes taking place in the development of the costal elements of the vertebrae of the cervical and lumbar regions. A separate centre of chondrification in the costal element of the first lumbar vertebra does not indicate a lumbar rib unless the costal process and the centre of chondrification resemble morphologically the twelfth thoracic rib so that there is no sudden change of form from the one to the other.
Comparative Development of the Vertebrae
For an account of the embryological development of the vertebne in the tower vertebrates and a summary of the literature relating to the subject the exeellent article by Schauinstand in Hertwig's Handbuch der Entwickelungsgesehichte der Wirbeltiere should be cousulted. In the anamniotes, the chorda dorsalis plays a relatively much greater part than in the amniotee, and in the sauropsida a mueh greater part than in the mammals. Primitively there are apparently four arch pieces developed on each side in each sclerotome, two dorsal and two ventral. As a rule, the cranially situated dorsal and ventral arch pieces of each sclerotome are incomplete while the caudally situated arches are firmly united to the sheath of the notochord. In the amniotes the cranial arch pieces of one sclerotome unite with the caudal of the sclerotome next cranialwards to form the definitive vertebral arches and arch bases. The bodies of the vertebra develop between the regions of attachment of the arches to the chorda. The neural and articular processes come from the dorsal arches, the transverse, costal, hiemal, and hypochordal (see p. 345) processes from the ventral arches.
In the development of the human vertebne the caudal dorsal and ventral arch aniages of each sclerotonie arise simultaneously and are soon united by a common base or cbordal process to the mesenchymal sheath about the notochord. The cranial dorsal and ventral arch aniages arise later, the dorsal becoming the interdorsal membranes, the ventral the interventral membranes. The cartilages of the bodies develop about the notochord between the bases of the cranial and caudal ventral arch anlages. But one centre of chondrification appears to give rise to the definitive vertebral hemi-arch, although a separate centre arises for the costal process.

Fig. 273.
The ossification of the definitive vertebrae varies so in different classes of vertebrates that no comparison of the process will be attempted here.
References
Bibliography
(On the development of the spinal column and of the thorax.)
Adolphi, H. : Uber die Variationen des Brustkorbes und der Wirbelsaule des Menschen. Morph. Jahrb. Bd. 33. 1905.
Aeby: Seltene Rippenanomalie des Menschen. Reichert's und du Bois-Reymond's Arch. f. Anat. und Physiol. 1868. Die Altersverschiedenheiten der menschlichen Wirbelsaule. Arch. f. Anat. und Physiol. Anat. Abt. S. 77. 1879.
Albrecht, p.: Uber den Pro-atlas, einen zwischen dem occipitale und dem Atlas der amnio ten Wirbeltiere gelegenen Wirbel und den N. spinalis I. s. proatlanticus. Zool. Anz. Bd. 3. 1880.
Alexander, Bela: Beitrage zur Kenntnis der Ossifizieurung der kartilaginosen Wirbelsaule. Orvosi HetUap. Nr. 51. 1903. Die Entwicklung der knochemen Wirbelsaule. Arch, und Atlas der normalen und patholog. Anat. in Rontgenbildem. Hamburg 1906.
Allen: On the Varieties of the Atlas in the Human Subject. Journ. of Anat. and Physiol. Vol. 14. 1879.
Ancel et Sencert: Variation numerique de la colonne vertebrale. Comptes rend. Assoc, des Anat., p. 158-165. Lyon 1901.
- Les variations des segments vert^bro-costaux. Bibliogr. Anat. t. 10, p. 214 239. 1902.
- De quelques variations dans le nombre des vertebres chez Thomme. Journ. de FAnat. et de la Physiol. Annee 38, p. 217-258. 1902.
Anderson: Eight True Ribs in Man. Anat. Anz. Bd. 4. 1889.
Aron, E. : Zur Kasuistik der Halsrippen. Berlin klin. Wochenschr. Jahrg. 29. 1892.
Ballantyne: Spinal Column in Infants. Edinburgh Medical Journal. 1892.
Barchielli, Alberto : Variazione del margine superiore dello stemo umano e loro significato. Monit. Zool. Ital. Vol. 15. 1904.
Bardeen: Costo-vertebral Variation in Man. Anat. Anz. Bd. 18, p. 377. 1900.
- Vertebral Variation in the Human Adult and Embryo. Anat. Anz. Bd. 25, p. 497. 1904.
- Development of the Thoracic Vertebrae in Man. Amer. Journ. of Anat. Vol. 4, p. 163. 1905.
- Studies of the Development of the Human Skeleton. The Development of the Lumbar, Sacral and Coccygeal Vertebrae. The Curves and the Proportionate Lengths of the Spinal Column during the First Three Months of Embryonic Development. Amer. Journ. of Anat. Vol. 4, p. 265-302. 1905.
Bardeleben, K. v.: Beitrage zur Anatomic der Wirbelsaule. Jena 1874. Uber das epistemum des Menschen. Sitzb. med. naturw. Ges. Jena. Bd. 146. 1879.
- Uber Verbindungen zwischen dem fiinften und sechsten sowie zwischen dem sechsten und siebenten Rippenknorpel. Anat. Anz. Bd. 15. 1898.
Bartels, M. : Uber Menschenschwanze. Arch, f . Anthrop. Bd. 13. 1881.
Bernhardt, M. : Uber das Vorkommen und die klinische Bedeutung der Halsrippen beim Menschen. Berlin, klin. Wochenschr. Jahrg. 32. 1894.
BiANCHi, S. : Sull interpretazione morf ologica della prima vertebra eoceigea nell' uomo. Atti della R. accad. del Fisiocritici in Siena. Ser. 4. Vol. 7. 1895.
Black, J. : Report of a Case of Cervical Ribs. Joum. Anat. and Physiol. Vol. 33. 1898.
BoCK£NH£iM£R : Zur Kenntnis der Spina bifida. Arch. klin. Chir. Bd. 65. 1902.
BoGUSAT, H. : Anomalien und Varietaten des Brustbeins. Diss. med. Konigsberg 1902.
BOLK : Ueber eine Wirbelsaule mit nur sechs Halswirbeln. Morph. Jahrb. Bd. 29, S. 84-^3. 1901.
- Zur Frage der Assimilation des Atlas am Schadel beim Menschen. Anat. Anz. Bd. 28. 1906.
Brugsch, Th. : Die Entwicklung des Ligamentmn caudale beim M^ischen. Diss. med. Leipzig 1903.
Chiarugi, G. : Per la storia dell' articolazione occipito-atlo assoidea. Monit. Zool. Ital. 1890.
Cornet, Y. : Note sur le pretendu pro-atlas des mammif^res et de Hatteria punctata. Bull. Acad. Roy. des Sciences de Belg. 1888.
Cunningham, J. : Tbe Neural Spines of tbe Cervical Vertebra as a Race-Character. Joum. of Anat. and Physiol. Vol. 20. 1886.
- Proportion of Bone and Cartilage in the Lumbar Section of the Vertebral Column of Apes and Several Races of Man. Joum. of Anat. and Physiol., Vol. 24, p. 117. 1889.
- The occasional Eighth True Rib in Man. Joum. Anat. and Physiol. Vol. 24. 1889.
Dukes, L. : Anomalies in the Cervical and Upper Thoracic Region, involving the Cervical Vertebrae, First Rib and Brachial Plexus. Joum. of Anat. and Physiol. Vol. 36. 1902.
Dwight, T. : The Sternum as an Index of Sex and Age. Jonm. of Anat. and Physiol. Vol. 15. 1881.
- Methods of Estimating the Height from Parts of the Skeleton. Medical Record. 1894.
- Description of Human Spines. Memoirs Boston Society of Natural History. Vol. 5, p. 237-312. 1901.
- A Transverse Foramen in the Last Lumbar Vertebra. Anat. Anz. Bd. 20, p. 571 bis 572. 1902.
v. Ebner: Urwirbel und Neugliederung der Wirbelsaule. Wiener Sitzungsber. Bd. 97. 3 Abt. 1888.
- Uber die Beziehungen des Wirbels zu den Urwirbeln. Wiener Sitzungsberichte. Bd. 101. Abt. 3. 1892.
Eggeling, H. : Uber den oberen Rand des menschlichen Brustbeinhandgriffes. Verb. Anat Ges. p. 41^8. Heidelberg 1903.
- Zur Morphologie des manubrium stemi. Festschrift zum 70. Oeburtstag von E. Haeckel. Jena 1904.
Falcone, C. : Sopra una particolaritk di sviluppo della colonna vertebrale nell' embrione umano. Giom. Intemaz. Sc. Med. Anno 23, p. 543-549. 1901.
Favaro, G. : II canale caudale nelF uomo. Verb. Anat. Qes, Padua 1906, p. 638.
Fawcett, Ed.: On the Completion of Ossification of the Human Sacrum. 6 Fig. Anat. Anz. Bd. 30, p. 414. 1907.
FiCK, A. G. : Zur Entwicklungsgeschichte der Rip pen und Querfortsatze. Arch, f . Anat. und Physiol. Anat. Abtl. p. 30-42. 1879.
FiscHEL, A.: Untersuchungen iiber die Wirbelsaule und den Brustkorb des Menschen. Anat. Hefte. Bd. 31. 1906.
Fischer, H. : Fissura stemi congenita mit partieller Bauchspalte. Deutsche Zeitschrift f. Chimrgie. Bd. 12. 1880.
FoL : Sur la queue de I'embryon humain. C. R. de PAcad. de Sc, Paris. Vol. 100, p. 1469. 1885.
Froriep, a.: Kopfteil der Chorda dorsalis bei mensch. Embryonen. Festschr. f. Henle. 1882.
- Zur Entwieklungsgeschichte der Wirbelsaule. Arch. f. Anat. und Physiol. Anat. Abt. S. 177-184. 1883; S. 69-150. 1886.
FuNKEy E. : Uber einen Processus ondontoideus atlantis hominis. Anat. Anz. Bd. 14. 1898.
Ganpini : Sopra alcune f acette articolari del basi-occipitale in rapporto ai processi basilari. Monit. ZooL Ital. Vol. 17, p. 60-68. 1906.
Gaupp, E. : Die Entwicklung der Wirbelsaule. Zusammenfassende Ubersicht. Zool. Gentralblatt. Bd. 3 u. 4. 1896-1897.
GERARD, G. : Anomalies osseuses. Sur la presence d'une cote eervicale articul^ avec la premiere cote formee elle-meme de la fusion des deux premieres cotes thoraciques. Bibliogr. Anat. Vol. 8. 1900.
Grotme, H. : Anomalien der Halswirbelsaule, nach den dem anatomischen Institute in Gottingen gesammelten Praparaten. Inaug. Dissert. Gottingen 1904.
Gruber: Uber die Halsrippen des Menschen mit vergleich. anatomischen Bemerkungen. Memoires de TAcad. des Sciences de St. Petersbourg. Vol. 13, Nr. 2. 1869.
Hagemann, Nil. : Selten vorkommende Abnormitat des Brustkorbes. Jahrb. f. Kinderheilk. 1880.
Hannover: Primordialbrusken og dens Forbening: Truncus og Extremiteme hos Menneskel for Fodselen. Vidensk. Selskabs. Skr. Vol. 4. 1887.
Harrison, R. G.: On the occurrence of Tails in Man, with a Description of the Case reported by Dr. Watson. The Johns Hopkins Bulletin. Vol. 12, p. 121-129. 190L
Hasse, C. : Die Entwicklung des Atlas und Epistropheus des Menschen und der Saugetiere. Anat. Studien. Bd. I., S. 542-568. 1873.
Hasse, C, und Schwarck, W. : Studien zur vergleichenden Anatomic der Wirbelsaule insbesondere des Menschen und der Saugetiere. Anat. Studien, Bd. 1, S. 21-171. 1873.
Herber: Uber Halsrippen. Med. Diss. Bonn 1903. Hoffmann : Zur Morphologic des Schultergiirtels und des Brustbeins bei Reptillien, Vogeln, Saugetieren und dem Menschen. Niederl. Arch. f. Zool. Bd. 5. 1879.
HoLL, M. : Uber die richtige Deutung der Querf ortsatze der Lendenwirbel und die Entwicklung der Wirbelsaule des Menschen. Sitzungsb. d. K. Akad. d. Wiss. Math.-naturw. Klasse. Wien. Bd. 85, S. 181-232. 1882.
V. Jherino, H. : Uber den Begriff der Segmente bei Wirbeltieren und Wirbellosen, nebst Beobachtungen iiber die Wirbelsaule des Menschen. Centralbl. f. d. med. Wiss. Nr. 9. 1878.
Keibel, Fr. : Uber den Schwanz des menschlichen Embryo. Arch. f. Anat. und Physiol. Anat. Abtl. 1891.
Keith, A.: The Extent to which the Posterior Segments of the Body have been transmutated and oppressed in the Evolution of Man and allied Primates. Joum. of Anat. and Physiol. Vol. 37. 1903.
Klein, C. : Uber die Konfiguration der lumbalen Intervertebralraume. Mitt, aus der Grenzgeb. d. Med. und Chir. Bd. 12, Heft 5. 1903.
Koch, W. : Beitrage zur Lehre von der Spina bifida. Kassel 1881.
KoLLMANN, J.: Die Entwicklung der Chorda dorsalis bei dem Menschen. Anat. Anz. Bd. 5, p. 308-321. 1890.
Krausse, 0. : Uber Halsrippen des Menschen. Med. Diss. Leipzig 1902.
Kravetz, L. p.: Entwieklungsgeschichte des Sternum und des Epistemalapparats der Saugetiere. Bull. Soc. Impr. Natur. Moscou. Ann^e 1905. Moscow 1906. Entwieklungsgeschichte des Sacrum und des Epistemalapparats der Saugetiere. Bull. Soc. Imp. Nat. Ebenda.
Lammers : Zur Frage der Entstehung des Promontoriums wahrend der Fetalperiode. Diss. Strassburg 1904.
Leboucq, H. : Recherches sur le mode de disparition de la chorde dorsale chez les vertebres superieurs. Arch, de Biol. T. 1. 1880.
- De quelques anomalies des cotes chez Thomme. Extr. des Ann. de la Soc. de Med. de Gand. 1885.
- 1. Zur Frage nach der Herkunft iiberzahliger Wirbel. 2. Die Querfortsatze der Halswirbel in ihrer Beziehung zu Halsrippen. Verhandlg. der Anat. Gesellsch. 8 Vers. Anat. Anz. Bd. 9. Erg. Heft. 1894.
- Recherches sur les variations anatomiques de la premiere cote chez riiomme. Arch, de Biol. Vol. 15, p. 125. 1898.
Le Damany, p. : L'adaptation de Thomme a la station debout. Joum. de FAnat. et Physiol. Vol. 41, p. 133. 1905.
Lovett, R. W., Davis, S. R. et Montgomery, A. M. : Curve fisiologiche della spina umana, loro origine e sviluppo. Arch, ortoped. Anno 23, p. 372-389. 1906,
Low: Description of a Specimen in which there is a Rudimentary First Rib, with Thirteen Pairs of Ribs and Twenty-five Presacral Vertebrae. Jour. of Anat. and Physiol. Vol. 34, p. 451-457. 1901.
Luschka, H.: Die Halsrippen und die Ossa suprastemalia des Mensohen. Denkschrift d. Kais. Akad. d. Wiss. Bd. 16, Abt. 2. Wien 1859.
Macauster: Notes on the Development and Variation of the Atlas. Joum. Anat. and Physiol. Vol. 27. 1893.
- The Development and Varieties of the Second Cervical Vertebra. Journ. Anat. and Physiol. Vol. 28. 1894.
Mall, F. P. : On Centers of Ossification in Hmnan Embryos less than 100 Days old. Amer. Joum. Anat. Vol. 5, p. 433. 1896.
Marchand: Article on Spina Bifida in Eulenburg's Realencyclopiidie. Bd. 22, S. 578-601. 1899.
Markowski, J.: Uber die Varietaten der Ossification des Brustbeins usw. Poln. Arch. f. Biol, und Med. Wiss. Bd. 1. 1902.
- Sollte der Verknocherungsprozess des Brustbeins von keiner morphologischen Bedeutung sein? Anat. Anz. Bd. 26, S. 248-269. 1905.
- Ma yet: Recherches sur Tossification du sternum chez les sujets normaux et chez les rachitiques. Bull, de la societe anat. de Paris. Vol. 70, p. 381-384. 1895.
- Note sur un nouveau procede de recherche et d'etude des points d'ossification epiphysaire. Bull. Soc. Anat. Vol. 70, p. 375. Paris 1895.
Meyer, R. : LTer einige Abnormitaten am Schwanzende menschlicher Feten. Virchow's Arch. Bd. 180, S. 334. 1906.
Moser: Das Wachstum der menschlichen Wirbelsaule. Diss. Strassburg 1889.
MuLLER, Ch. : Zur Entwicklung des menschlichen Brustkorbes. Morph. Jahrb. Bd. 35, S. 591-696. 1906.
Naunyn: Spina bifida occulta. Deutsch. med. Wochenschrift. 1898.
Papillault, G. : Variations numeriques des vertebres lumbaires chez Phomme. Bull, de la Soc. d'Anthropologie de Paris. T. 9, p. 198-222. 1900.
Paterson, a. M. : The Human Sacrum. Scientific Transactions of the Royal Dublin Society. Vol. 5, p. 123. 1893.
Paterson The sternum - its early development and ossification in man and mammals. (1900) J Anat Physiol. 35(1): 21-32 PMID 17232454
- The Sternum; Its Early Development and Ossification in Man and Mammals. Joum. of Anat. and Physiol. Vol. 35, p. 31-32. 1900.
- The Human Sternum — Three Lectures delivered at the R. College of Surgeons. London 1904.
Pauusch, 0.: Das vordere Ende der chorda dorsalis und der Frank*sche Nasenkamm. Arch. f. Anat und Phvsiol. Anat. Abtl. S. 187-215. 1887.
Pilling: Uber die Halsrippen des Menschen. Diss. Rostock 1894.
Pjatnizky, J. J. : Uber den Bau des menschlichen Schwanzes und iiber menschliche Schwanze im allgemeinen. Diss. St. Petersburg 1893. (Russian.)
Planteau, H. : Developpement de la oolonne vertebrale. Th^ de Paris 1883.
PoSTH, M.: Le Sacrum. Th^se de Paris 1897.
Rabaud, E. : Gen^e des spina bifida. Arch. gen. de med. N. s. Vol. 5, p. 283-309. 1901.
Rawzi: Zur Kasuistik der Halsrippen. Wien. Klin. Wochenschr. 1903.
Ravenel: Die Massen verbal tnisse der Wirbelsaule und des Riickenmarks beim Menschen. Zeitschr. f. Anat. und Entwicklungsg. Bd. 2, S. 334. 1877.
Recklinghausen, F. v.: Untersuchungen iiber spina bifida. Virchow's Arch. Bd. 105. 1886.
Retzius, a.: Tiber die richtige Deutung der Seitenfortsatze an den Riickenu. Lendenwirbeln beim Menschen und bei den Saugetieren. Arch. f. Anat. u. Physiol. 1849.
Robin: Memoire sur le developpement des vertebres, atlas et axis. Joum. de PAnat. et de la Physiol. Vol. 1, p. 274-209.
- 1864. Memoire sur revolution de la notoccrde. Paris. 212 pp., 12 pis., 63 Figs. 1868.
RoDENACKER, G. : Uber den Saugetierschwanz mit besonderer Beruckachtigung der kaudalen Anhange des Menschen. Diss. Freiburg-Breisgau 1898.
Rosenberg, E. : Uber die Entwicklung der Wirbelsaule und das cent rale carpi des Menschen. Morphol. Jahrb. Bd. I, S. 83. 1876.
- Uber die Entwicklung der Wirbelsaule. Sitzungsber. des Naturforschergesellschaf t bei der Universitat Dorpat. Bd. 7. Sitzung vom 17. Febr. 1883.
- Uber eine primitive Form der Wirbelsaule des Menschen. Morphol. Jahrb. Bd. 27, S. 1-118. 1899.
- Bemerkungen iiber den Modus des Zustandekommens der Regionen an der Wirbelsaule des Menschen. Morph. Jahrb. Bd. 36. 1907.
RUGE, G. : Untersuchungen uber Entwicklungsvorgiinge am Brustbein und der Stemoclavicularverbindung der Menschen. Morphol. Jahrb. Bd. 6, S. 362-414. 1880.
ScHAUiNSLAND, H. : Die Entwicklung der Wirbelsaule nebst Rippen imd Brustbein. O. Hertwig's Handbuch der Entw.-Lehre. Bd. 3. 1906.
Schultze: Uber embryonale und bleibende Segmentierung. Verhandl. der Anat. Gesellschaft. 10 Vers. Berlin, S. 87-92. 1896.
Schumacher, S. v.: Uber Haemalbogen bei menschlichen Embryonen. Verb. Anat. Ges. Rostock. Erg. Heft. Anat. Anz. Bd. 29, S. 173-178. 1906.
Schwegel: Die Entwicklungsgeschichte der Knochen des Stammes und der Extremitiiten. Sitz. Ber. Akad. d. Wiss. Wien. Bd. 30. 1858.
Spanje, N. p. van : Een angeboren defect van het sternum. Weekblad van bet Nederl. Tydschr. voor Geneesk., Nr. 6. 1904.
Staderini, R. : Richerche statistiche suUa frequenza della varietk numeriche delle vertebre neir uomo e considerazioni sulla loro genesi. Monit. Zool. Ital. Vol. 5. 1894.
Steinbach: Die Zahl der Caudalwirbel beim Menschen. Diss. Berlin 1889.
Stieda, L. : Uber Halsrippen. Virchow's Arch. Bd. 36. 1866.
Strauch, M. : Anatoniische Untersuchungen des Brustbeins des Menschen. Diss. Dorpat 1881.
Struthers, J.: On Variations of the Vertebrae and Ribs in Man. Joum. of Anat. and Physiol. Vol. 9, p. 17-96. 1875.
Swjetschnikow : Uber die Assimilation des Atlas und die Manifestation des occipital-wirbels beim Menschen. Arch. f. Anat. u. Physiol. Anat. Abtl. S. 155-194. 1906.
Szawlowski : Uber einige seltene Variationen an der Wirbelsaule beim Menschen. Anat. Anz. Bd. 20, S. 305-320. 1901.
Tenchini : Di una nuova maniera di compenso nelle anomalie numeriche vertebrali deir uomo. Archixio per I'Anthropologia. Vol. 24. Firenze 1894.
Thaler, H. A.: Atypische Verhaltnisse in der Steissgegend menschlicher Feten und eines Neugeborenen. Deutsche Zeitschr. f. Chir. Bd. 79, S. 112. 1906 und Jahresbericht u. Arb. d. zweiten chir. Klinik Wein, 1906, p. 140-153.
ToPiNARD, P. : Anomalies de nombre de la colonne vertebrale chez Thomme. Revue d'Anthropologie. T. 6, p. 577. 1877.
TouBNEUX, F. ET Herbmann, G. I SuF la persistance de vestiges medullaires coccygiennes pendant toute la periode foetale chez Phamme. Joum. de FAnat. et de la Physiol. Ann^e 23, p. 498. 1887.
ToUBNEUX, F. ET Mabtin, E. : Contribution k Fhistoire du spina bifida. Joum. de lAnat. et de la Physiol. Annee 17. 1881.
Tbedgold, A. F. : Variations of Ribs in Primates with Especial Reference to the Number of Sternal Ribs in Man. Joum. of Anat. and Physiol. Vol. 31. 1897.
TuRNEB, W. : On the So-called Two-headed Ribs in Whales and in Man. Joum. of Anat. and Physiol. Vol. 5. 1871.
- Ungeb und Bbugsch: Zur Kenntnis der Fovea und Fistula saero-coccygea s. caudalis etc. Arch. f. mikr. Anat. Bd. 61, S. 151-219. 1903.
Valenti, G. : Sopra il significato dell' apofisi laterale delle vertebre lombari e delle massi lateral! del sacro. Rend. accad. Sc. Istit. Bologna, Anno accad. 1902-1903 ; Bull. Sc. Med. Anno 74. Mem. accad. Sc. Istit. Bologna, Anno accad. 1902-1903; Bull. Sc. Med. Anno 74. Mem. accad. Sc. Istit. Vol. 10. Bologna, 1903. Sopra un caso di costa raddoppiata osservato nelF uomo. Mem. accad. Sc. 1st. Ser. 5. Vol. 9. Bologna 1903. Sopra il significato delle Apofisi lateral! delle vertebre cervical! nelP uomo. Ricerche embryologiche. accad. Sc. Istit. Bologna 1906.
VoLTZ, W. : Tiber kongenitale volkommene Synostose der Wirbelsaule in Verbindung mit Wachstumsanomalien der Extremitatenknochen. Mitt. a. d. Grenzgeb. d. Med. u. Chir. Bd. 16, S. 61-75. 1906.
Walter, F.: Uber Halsrippen. Med. Diss. Halle 1906. Weiss, A.: Die Entwicklung der Wirbelsaule der weissen Ratte, besonders der vordersten Halswirbel. Zeitschr. f. wiss. Zool. Bd. 66, S. 492. 1901.
WiRTH, A. : Zur Anatomic und Entwicklungsgeschichte des Atlas und Epistropheus. Leipzig 1884.
- Skeleton and Connective Tissues: Connective Tissue Histogenesis | Skeletal Morphogenesis | Chorda Dorsalis | Vertebral Column and Thorax | Limb Skeleton | Skull Hyoid Bone Larynx
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2026, August 2) Embryology Book - Manual of Human Embryology 11C. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Book_-_Manual_of_Human_Embryology_11C
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G