Paper - Development of the region of the isthmus rhombencephali (1928)
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Frazer JE. Development of the region of the isthmus rhombencephali. (1928) J Anat. 63: 7-18. PMID 17104212
| Online Editor |
|---|

|
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Development of the Region of the Isthmus Rhombencephali

{kind=link}
J. Ernest Frazer, F.R.C.S.
Professor of Anatomy in the University of London
IT does not appear that much attention has been paid to the ontogenetic development of the region of the isthmus rhombencephali, in spite of the interest attaching to it from many points of view. Some years ago I was struck by the appearance of invagination of the hind-brain in this part, when I was making observations on relative positions of the fourth nerve, and I have recently confirmed and extended these earlier findings. The interest and importance which might be connected with them seem to justify their production in this paper, and it will simplify the matter if the nerve and some of the problems associated with it are considered first.
A. The Embryonic Isthmus and the Fourth Nerve
In Plate I are shown drawings, from models, of the region in embryos of 12, 16, 21, and 26 mm.; these are seen in sagittal section, and in the cases of the 12 and 16 mm. specimens a view is also given of the cranial slope of the hind-brain in each. Looking at this cranial slope one is struck by the fact that alar and basal laminae are prolonged upwards and forwards above the opening into the mid-brain. In the 12 mm. embryo the laminae, although evident, are not very sharply defined, but in the older one they are very clearly cut, the upper part of the alar lamina on each side showing already the beginning of its slow cerebellar change. Turning now to the sagittal section of the 12 mm. embryo, the basal lamina is seen passing up beside the passage into the midbrain, forming its lateral boundary, to reach the upper part of the hind-brain above this. The mid-brain shows a thick “ floor lamina,” above which the thin side wall of the large cavity is made by the tectum. The floor lamina is continued in a backward and downward direction into the basal lamina of the hind-brain at the lower part of the side wall of the opening between the two brain regions, and in doing this meets the basal lamina at an acute angle, opening upwards and forwards: the deep sulcus made in this way between the two laminae, and running into the angle, can be referred to for the purposes of this paper as the intralaminar sulcus, this name covering its present appearance and its later fate.
By the 16 mm. stage the basal lamina has evidently grown in thickness, leading, with its fellow, to more marked narrowing of the mid-brain opening, and the sagittal section shows that, in its growth, it bulges to some extent into the mid-brain. In doing this it is applied to the upper surface of the floor lamina, making a longer and well-marked intralaminar sulcus, the terminal angle of which is moved a little forward. There is a definite forward movement of this part of the hind-brain with reference to the dorsal notch which marks the isthmus, for the dorsal decussation of the fourth nerve is on the hin(l-brai11 in the 12 1n1n. embryo, but has moved forward in the later specimen, in which it occupies the angle of the notch (see Plate 1, fig. 1). It is possible, perhaps probable, that some of this change in position may be (1110 to the backward push of the growing mid-brain tectum, but this, judging from the condition of the teetum in the neighbourhood of the lamina, cannot be considered to be a factor of much importance, whereas a forward growth of the hind-brain structure is only in keeping with the undoubted nature of the process as seen in subsequent stages.
This displacement of the nerve from an original situation in the hind-brain seems to be fairly constant~so far, at least, as my own observations extend on other forms. In the human embryo the decussation is always in the hind-brain up to the stage of 15 or 16 1mn., its position in the earlier stages being rather further back than shown in the embryo of 12 mm. This may perhaps explain the statement, occasionally made, that the nerve appears later than the other cranial nerves: it seems to me that it may be found as soon as the other cranial nerves, although it must be admitted that its minute size renders its recognition dillieult in young embryos.
The next stage seen in Plate I, that of 21 mm., shows the appearance of a new factor in the modelling of the region. The alar lamina now presents the appearance of being carried forward into the mid-brain, lying above the original basal projection, which is thus pressed down against the floor lamina. The intralaminar sulcus, as a result of this, undergoes a fusion of its opposed walls, the fusion progressing from behind forward, so that on surface View the deep sulcus is now represented by a shallower groove leading to the angular open region. Behind and above this angle is the dislocated basal lamina, and the displaced alar structure above and behind this again. Sections show the fused walls of the intralaminar sulcus as a thick shelf of darkly staining ependymal nuclei projecting into the laminar structure for a considerable distance.
The last specimen drawn, one of 26 mm., shows a little advance; the basal and floor laminae are more closely united, with a less marked groove between them, the alar portion is perhaps more evident, and some new structures associated with the tectal growth are beginning to come into view above these alar and basal parts. It is to be noted that the nerve decussates still at the bottom of the dorsal angle. This is really its definitive position (to which it may therefore be said to attain at 16 mm.) and it is not affected by the appearance of the alar lamina here: the explanation of this will come later, but it may be said here-that the appearances, as seen from within, are in a sense a little deceptive so far as the alar lamina is concerned.
It remains to call attention to the recess in the middle line of the floor, below the junction of floor and basal laminae. The interpeduncular ganglion is formed from the thick ependyma at the angle of this depression, which makes it useful as a topical point, and the recess may be referred to when necessary as the interpeduncular recess.
The developmental stage reached so far has, for all practical purposes, put the nerve in its final position. Its decussation has been shifted forward to the hinder limit of the mid-brain, ,and its nucleus of origin has been carried into this region of the brain by the dislocation forwardof the basal lamina. At 12 mm. the nucleus is in the front part of the basal lamina, about halfway up the height of the side wallof the mid-brain opening.
- Of course only an approximate position can be given for the “nucleus,” the ependymal thickening around the apparent termination of the nerve fibres being taken for this purpose.
As this lamina is carried forward and downward against the floor lamina, the nucleus is carried in the same direction, and the original front face of the basal lamina being now the upper wall of the intralaminar sulcus, the nucleus is now in contact with this. As the walls of the sulcus disappear the nucleus is no longer held back by it, and there is some reason to think that it is displaced further downwards and inwards at more advanced stages as a result of pressure from other formations. There would seem to be in these changes a very simple explanation of the angled intracerebral course pursued by the nerve.
There are two problems of greater interest, however, connected with this nerve, which up to the present have not received any adequate explanation— its dorsal exit and its decussation after its origin.
In dealing with the dorsal situation of the superficial origin of the nerve, it is necessary to refer to that part of the hind-brain which has been seen to be above the opening into the mid-brain. In this part are two laminae, basal and alar, on each side, directly and evidently continuous with the same undoubted structures below the level of the opening. Examination of sections shows that the structure of the upper parts, the disposition of neuroblast nuclei, etc., are exactly as in the lower portions, and there does not, in fact, seem to be any reason at all for regarding them as anything other than the continuations of these parts. In other words, the floor of the hind-brain is continued past the opening into the mid-brain, as is really at once evident on looking at the examples in Plate I. Comparing this aspect with the embryonic structure of the spinal cord, as a standard, it would seem better to restrict the term “floor” to the basal laminae, for it is customary to consider the cord to be “roofed” by the dorsal laminae and the roof-plate joining them.
The basal laminae, then, of the upper part of the hind-brain, above the level of the mid-brain opening, make a true floor to the neural tube here, and the thin layer joining them across the middle line is evidently an ordinary floor-plate: they form here the ventral moiety of the neural tube, just as they do lower down, and in the spinal cord. Hence the fourth nerve, when it first appears, makes its exit from the ventral surface of the tube, as is the case with all efferent roots. This ventral surface is then moved toward the mid-brain, to the isthmus. As the original situation was above the mid-brain, it follows that the ventral surface when it moves must reach the dorsal aspect of the isthmus, just as it would reach the ventral aspect if it moved from below the level of the opening. The attainment of the isthmus, then, is the result of the forward move, and the dorsal position here is the necessary registration or expression of the originally ventral situation of the nerve. The nerve is essentially ventral although actually dorsal.
But although we can refer the dorsal origin in this way to the existence of an upper portion of the hind—brain, from which the nerve originally arises, no light is thrown on the meaning of the presence of such a part of the brain. I have not so far been fortunate enough to meet with any adequate suggestion in developmental occurrences, and must perforce be content to leave the matter there for the present.

Text-fig. 1. Schemes to show the essentially similar nature of a dorsal efferent root and of the fourth nerve (right). C., H. contralateral and homolateral origins. The arrow indicates that the contralateral origin of the fourth nerve is secondarily displaced further away, as is mentioned in the text.
The question of the meaning of the decussation after origin does not admit of solution on mechanical lines similar to the previous one, but there is, I think, good reason for saying that it may be explained on general grounds connected with the origin of many, if not all, efferent roots. It is known that the anterior roots of the spinal nerves, in addition to the fibres derived from ventral horn cells of their own side, possess also fibres which arise from cells situated on the other side of the cord. These contralateral fibres cross in the White ventral commissure and pass from there directly into the issuing root. The cells giving origin to them are described by Winkler as being in what he terms the “pars intermedia” of the cord, and the fibres are destined for the “sympathetic” system. Presumably, also, there is some cerebellar influence exercised on these cells, for Winkler describes cerebello-fugal fibres as reaching this same zone. We may say, then, that (in the dorsal, dorso-lumbar, and probably sacral regions at any rate) the spinal efferent roots have homolateral and contralateral origins: this is shown in the left-hand diagram in text-fig. 1, where C is the relatively small contralateral element in the root, and H the large and overshadowing homolateral portion. The condition which, it seemed to me, might be represented by the fourth nerve is shown on the right side, where the homolateral element is reduced to a minimum, while the contralateral portion is forming practically the whole of the nerve. This puts this nerve quite in line with the other, only differing in the proportions existing between its constituent parts, while its contralateral fibres cross in the ventral commissure, which is of course superficial here owing to the absence of the ventral projections which are so marked in the spinal cord. Taking this idea as a working hypothesis, a search was made for some representative of the homolateral origin, if perchance it might still be in evidence, and I think I have been able to find such a remnant: this has so far only been found in the older specimens, the younger ones being indeterminate in all their nuclear structures, as already pointed out. The small bundle of fibres to which I refer has been found in all stages examined, after the middle of the third month and up to the time of birth (see Plate III, C). The cells from which they seem to arise form a group about halfway down the side wall of the cavity, in what would, probably, have been the upper part of the (now invaginated) basal lamina. My sections, however, are ordinary serial sections, stained in the ordinary routine way, and I cannot be quite certain that the bundle actually arises in this way, although I believe that the observation is correct. Under these circumstances I was glad to find and fall back upon the experimental results of van Gehuchten, who described, in the rabbit, the existence of homolateral degeneration occurring after destruction of the fourth nerve. This was an example of secondary degeneration, some weeks after the ablation of the nerve, and the result seems to have been quite clear.
The explanation offered, then, for the problem of the decussation is little more than a suggestion, having value only in so far as van Gehuchten’s experiment and the observations on the human embryo may be considered to have weight. The suggestion, simply, is that the nerve is like other efferent roots, possessing homolateral and contralateral origins, but differs from them in that its contralateral part completely overshadows the other, so that practically the whole nerve decussates.
Sagittal sections of the region in embryos of 35 mm. and 48 mm. are shown in Plate II, fig. 1. They exhibit progressive changes on the lines already indi. cated. The intralaminar groove has disappeared completely, and the interlaminar sulcus is very deep. The tectal thickening is progressing slowly, and two rounded masses, already faintly indicated in the 26 mm. stage, are now seen to be advancing inwards in the floor of the caudal projection of the midbrain, above the region occupied by the basal lamina.
B. The Deeper Structure of the Region
The method of transverse section can be taken as a convenient way of illustrating the changes taking place during the building up of the region of the isthmus. The line of section which has been adopted passes through the dorsal notch above and the interpeduncular recess or ganglion below, these being easily defined: thus the sections (reconstructed) are approximately similar and comparable.
The first figure in text-fig. 2 is a diagram intended to show the disposition of the several parts entering into the formation of the wall of the cavity in the 12 mm. embryo. In the hind-brain (H) the basal and alar areas are represented respectively by cross-hatching and small circles, while the tectal region of the mid-brain (M) is shown by dots. The floor lamina of the mid-brain is marked like the basal lamina, with which it is continuous. It must be noted here that this floor lamina contains a Well-developed zone of short association fibres lying against its thick ependymal layer, extending back to the basal lamina and forward to the region of the third nerve nucleus.

Text-fig. 2. On the left, a scheme showing the component parts in the regions of hind-brain, H ., and mid-brain, M. Further explanation in text. The arrow gives direction of section (on right). BL. basal lamina, FL. floor lamina, 221.3. intralaminar sulcus, a. f. area. of short association fibres.
These fibres, although best marked in this stage in the mid-brain, are at any rate indicated to some degree in the hind-brain. Later, as will be mentioned, they extend, and it becomes evident that they are the primary foundation of the posterior longitudinal bundle. They do not reach the region of the posterior commissure at the 12 mm. stage, nor for some little time after this. The fibres in the commissure are forming, however, but are quite distinct from those now being described. I hope to be able to enter more fully into these matters in a future communication.
The transverse section (text-fig. 2) thus shows a lower part, the floor lamina, marked by the association fibres and separated above from the upper part by a deep cleft, which is the intralaminar sulcus. The part above this sulcus is the basal lamina, containing no association fibres. The trochlear nerve, issuing from the hind-brain at this stage, is not cut at the top of the section, but is divided lower down near its nucleus, which is just in front of the line of section. Thus there are nothing but basal and floor structures in the section of the isthmus region, and the thickness of the floor lamina seems to be due almost entirely to the large formation of association fibres.
Coming to the 16 mm. stage, there is decided growth of the basal lamina, and also to some extent of the floor lamina. The association fibres have increased considerably, and are now definitely continued into the hind-brain. In addition they are involving the portion of the basal lamina which is laid against the floor lamina, turning round the hinder and shallower part of the intralaminar sulcus in doing so.
The section shows (text—fig. 3) the early result of the growth of the basal lamina. The intralaminar sulcus is being pushed down in the side wall and is being invaded by the groups of association fibres. The nerve is now cut in decussation in the roof.

Text-fig. 3. 16 mm. stage. Position of structures, indicated as in last figure. Section through isthmus: fourth nerve decussating in roof, t.s. translaminar groove, other references as before.
The section at this stage, then, shows considerable modification in the form of the cavity, with lowering and closing of the intralaminar sulcus and the beginning of a new secondary “translaminar” groove higher up, dividing the basal lamina into upper and lower parts. The whole wall, however, is still composed only of basal tissues, in which the nerve decussates in the roof and an increased area of association fibres lies in the lower part.
The 21 mm. stage is of interest, because in it the dorsal lamina appears to have reached, essentially if not actually, the small degree of forward movement as a whole which seems to have been necessary to admit of further extension of its parts.
Two sections are shown in Plate II, fig. 2, on the right through the hindbrain of an embryo of 19 mm., just below the interpeduncular angle, on the left through an embryo of 26 mm. at a lower level. The neuroblastic groups are shown as shaded areas. It is not necessary to go into the matter of their derivations in this paper, and the sections are introduced to show the low prominence a on the outer surface of the hind-brain, more or less opposite the junction of alar and basal laminae. This is only the sectional appearance of a ridge running up this part of the hind-brain, evidently caused by a condensation or collection of neuroblasts in this situation. The ridge is visible, though ill-defined, in the 16 mm. specimen (Plate II, fig. 3) and passes, of course, up towards and on to the upper part of the hind—brain, above the level of the isthmus. In the 21 mm. embryo (on the right in Plate II, fig. 3) the growth of the alar lamina has moved the ridge forward, so that its front part reaches and runs on to the caudal end of the mid-brain, beyond the dorsal notch of the isthmus: in the figure the projecting part of the mid—brain is represented as raised up, to give a better View of the upper end of the ridge. The important thing connected with this ridge is that—when they are first found——the fibres of Gowers’ tract lie in the clear zone behind the neuroblastic condensation, while those belonging to other tracts and reaching the mid—brain ultimately are placed in the clear areas in front of it: these latter areas, then, as the result of the changes reached in the 21 mm. embryo, are brought into relation and continuity with the mid-brain, whereas the spino-cerebellar fibres are left further back, behind the isthmus.

Text-fig. 4. Schematic analysis of 21 mm. stage (left), with section through isthmus (right).
The growth of the alar lamina, which has led to the changes on the outer aspect which have just been considered, has its effect also on the internal appearance, as has been shown in Plate I. As already mentioned, however, this appearance is deceptive. The schematic analysis of the part is given in text-fig. 4, where it can be seen that the alar region, although carried forward and separated by what is apparently an interlaminar sulcus from the (lower part of the) basal lamina, is occupying here a situation in which the marking of the basal structure is still represented. The explanation is that the basal lamina remains, and alone forms the side wall of the cavity here, while the derivatives of the alar lamina, in growing forward, pass on its outer side. The appearance of an interlaminar sulcus is produced by a modification of a condition faintly foreshadowed in the 16 mm embryo.
In this embryo (Plate I) it can be seen that the upper part of the basal lamina, for a little distance below the nerve, projects somewhat into the cavity. This can be seen also on section (text-fig. 3) where’ the upper projection is separated from the rest of the basal lamina by a shallow depression: this has already been noticed as a “translaminar” groove, crossing the basal lamina (Plate I) in a U-shaped manner. The hinder limb of the U-groove is lost in the upper part of the true interlaminar sulcus. When the alar lamina begins to grow in thickness the interlaminar sulcus is pushed forward, so that it runs into the U-groove nearer its lowest part. This is seen in text-fig. 5, in which a sagittal section of the region in an embryo of 19 mm. is shown.
- An embryo of 18 mm. provided comparable results. As many of the sections however had been injured, I was very glad to have the opportunity of examining this perfect specimen, for the loan of which I am much indebted to Dr R. Gladstone.
The other diagrams in the figure show schematically how the change is effected: the forward displacement of the interlaminar sulcus makes a groove system of a Y-shape, and there is a rapid deepening of the front limb of the Y, with an equally rapid shallowing and practical disappearance of the hinder limb, whence it comes about that the stem and front limb remain as a continuous sulcus. It is evident, however, that although the groove made in this way has all the appearance of an interlaminar sulcus, its upper and front part, in the region of the isthmus, is actually translaminar.

Text-fig. 5. Sagittal section, 19 mm., on left, showing the intermediate stage in the changing aspect of the wall "of the cavity. On the right, schemes to represent the method in which the change is brought about. A, B, F, alar, basal, and floor laminae.
In this way, then, the section across the isthmus (text-fig. 4) of the 21 mm. specimen shows the basal lamina lining the cavity, with the deep translaminar sulcus cutting into it, while a small ependymal thickening, low down, marks all that is left of the originally deep intralaminar groove. The association fibres (or posterior longitudinal bundle, as one might now venture to term the collection) extend now well above the intralaminar remnant, and the lamina is thickening outside this, with a scattered mantle layer of nuclei. The alar lamina is now represented by a. small area of the section outside and above the translaminar sulcus. This addition to the section has included the issuing nerve in part, burying it in its course to a small extent.
Text-fig. 6 gives the schematic analysis of the 35 mm. stage, with a half section (A). It can be seen that the basal wall of the cavity is practically as before, save that there is now no sign of the intralaminar remnant, the trans-laminar being represented alone as a deep groove. The lower part of the basal area has thickened considerably, with migration of neuroblasts. The main feature, however, is the great increase in size of the alar area, which now extends down well over the outer part of the basal formation.
The second half-section, B, represents the conditions in the 48 mm. specimen, at a much smaller magnification. The alar growth is now larger and a ventro-lateral extension of it is shown: whether this is really alar in formation, or a secondary involvement of the basal surface by fibres running to the alar structures, cannot be decided, although the latter seems to me to be the more likely explanation. Fibres extending (superior peduncle) into the basal region from the alar, are clearly seen: these, as a matter of fact, are also suggested to some extent in the 35 mm. embryo. The ventro-lateral fibres seem to belong to the lateral lemniscus formation.

Text-fig. 6. Schematic analysis, 35 mm., on left. Sections A 35 mm., B 48 mm.
A more interesting set of fibres, however, is found in this specimen, which could not be picked up with any certainty in the earlier embryo. Plate III, fig. 1, A, is a section running through the roof of the fourth ventricle dorsally, a little distance behind the isthmus, while its oblique direction carries it ventrally through the group of nuclei connected with the fifth nerve. Certain fibres are clearly seen extending between these dorsal and ventral regions which are assumed to be connected with the nuclei and nerve. It is impossible, of course, to assert positively that they are so connected, but, to my mind at any rate, there can be little doubt about the matter, and the ultimate distribution of the fibres, when traced cranially, seems to decide it. At the level of the section the fibres can be seen to reach small nuclear masses in the ventricular roof, and also, by the inner fibres, to be connected with large cells or small cell groups near the interlaminar sulcus. As they are followed upwards in successive sections the fibres going to the interlaminar region seem at first to be associated more with cells on the basal side of the groove than with alar cells, but there soon comes a change: the fibres npw pass round the groove and reach cell groups on its alar side. Then these groups leave their near relation to the ependyma and become relatively deep, and in this way the isthmus is reached.
The position of the groups or cells is now shown in the section (text—fig. 6) by large black dots. This clearly represents the mesencephalic root of the nerve, and it extends forward at this stage only as far as the alar invasion seems to have extended, that is, to the deep hollow in front of the invaginated parts.
The level at which the cells associated with these fibres sink to a deeper situation seems to correspond with the passage of the true interlaminar sulcus into its translaminar portion, in other words with the forward growth of the alar lamina outside the basal structure. It is of much interest to observe that this extension of the connections of the fifth nerve into the mid-brain is associated with the alar lamina. I have made no observations on its further extension forward.
Models of the region in embryos of 75 mm. and 100 mm. show no essential difference in the region of the isthmus. The cavity seems smaller, partly owing to the relative size of the surrounding parts, but also perhaps because it is actually smaller. The translaminar sulcus is very deep, the wall above it being flattened down. Otherwise, the cavity is the same as in the earlier stage, and the solid structures outside it are further differentiations in .s'it12 of the parts placed in position in this earlier stage. This general description seems to be applicable to the conditions throughout the rest of foetal life, and the state in the fifth month (Plate III, B) illustrates this very well. The next figure (C) gives an illustration of the homolateral origin of the fourth nerve, such as may be found, apparently, in most specimens in the foetal stages.
Little or nothing would be gained by further description or discussion of what has been brought forward in this paper. I venture to think that the various changes which have been shown, and can be grouped together as associated with the process of “ dislocation ” in the region of the isthmus, must be looked on as facts, whatever explanation of their occurrence may be adopted ultimately. The same, perhaps, may be said for the change in position of the nerve-decussation in its relation to the explanation of the dorsal exit of the fourth nerve. The explanation of the decussation after origin has been put forward in the form of a suggestion based on comparison with other origins, and fortified by experimental result and observation: it seems to me, I confess, to be in keeping with what is known about the nerve. So far as the other observations are concerned, it must be remembered that they were carried out on ordinary serial sections not made with any special object nor stained in any special way: as a rule iron haematoxylin was the stain used, occasionally counterstained with eosin. Under these circumstances I imagine that the observations err on the minimal side, that is, for instance, the fibres described were probably there at an earlier stage than that in which one could certainly recognise them, and their field of extent was possibly larger than it seemed to be, and so on. These, after all, are faults on the right side, if they exist, so no more need be said about them. Beyond indicating its position and extent, nothing has been said about the tectal growth in the mid-brain, because it is not really concerned in building up the region of the isthmus. In the later stages figured, the slow ingrowths of this formation are to be seen above those described, and it is only necessary here to add that this slow growth leads gradually to the closure of that portion of the cavity contained in the projection of the mid-brain caudally: the cavity is closed in a specimen of 115 mm.
Summary
- The floor of the cranial slope of the hind-brain is continued upwards past the opening into the mid-brain.
- The fourth nerve has its superficial origin at first from this upward prolongation of the floor.
- The basal lamina beside the mid-brain opening is displaced forward into this, and folded down against the floor lamina of the mid-brain, producing an intralaminar sulcus. Thus the nucleus of origin of the fourth nerve is carried forward and put in immediate relation with the intralaminar sulcus.
- At the same time, with the forward displacement of the upper part of the basal lamina, the decussation of the nerve is carried on to the dorsal surface of the isthmus. This dorsal position is therefore only the necessary result of the displacement of the original ventral origin on that part of the hind-brain above the region of the isthmus.
- The alar lamina is displaced forward, passing on the outer side of the basal lamina in the isthmus, and thus leaving this lamina to form the wall of the cavity here. The displacement of the alar lamina enables formations associated with it to reach the mid-brain.
- A deceptive appearance of an interlaminar sulcus is produced in the region of the isthmus by the deepening of a translaminar groove across the basal lamina. The intralaminar sulcus disappears by fusion and breaking up of its ependymal walls, after it has been pushed down to a low level in the wall of the isthmus, and the fourth nucleus is thus left low down in the wall.
- The translaminar sulcus remains as a deep sulcus running through the isthmus, on each side, but the wall, both above and below it, is only made by basal lamina.
- A short description is given of the general structure of the walls of the isthmus at different stages, the mesencephalic root of the fifth nerve being shown to belong to the alar lamina here.
- A suggestion is made that the decussation after origin of the fourth nerve is comparable with the decussation of contralateral fibres of origin of spinal nerves: in these the homolateral origin obscures the contralateral, but in the fourth nerve the homolateral origin is reduced to a minimum, and the contralateral becomes the main origin. Some support for this is advanced.
References
A. VAN GEHUCHTEN (1906). Anal. du Syst. Nerv. de l’Homme. Iv. Louvain.
C. WINKLER (1918). Opera Omnia. Tom. VI, &c. Haarlem.
Plates
Plate I

Fig. 1. Above, cranial slope of hind-brain and sagittal section of hind- and mid-brain in 12 mm. embryo. Middle row, similar views of parts in 16 mm. embryo. Lower row, sagittal sections in 21 mm. (left) and 26 mm. (right). N. decussation of fourth nerve; a.l. alar lamina; b.l. basal lamina; f.l. floor lamina; t. tectum of mid-brain; s. interlaminar sulcus; i.l.s. intralaminar sulcus; i.p.r. interpeduncular recess. Magnifications decrease with advancing age.
Plate II

Fig.1.Sagittal sections through the region in embryos of 35 mm. and 48 mm.
Fig. 2. Two sections through the hind-brain, from embryos of 19 mm. and 26 mm. To show the low ridge, a, visible on the outer surface of the hind-brain.
Fig. 3. Outer aspects of the region in embryos of 16 mm. (left) and 21 mm. (right), showing the‘ ridge, a. This passes up behind the isthmus in the younger embryo, but runs towards it in the other, so that its front part reaches the mid-brain. The teotum has been raised to show this.
Plate III

Fig. 1. A, section through upper part of fourth ventricle, 48 mm.: V, place where fifth nerve appears, immediately above this: nuclear masses associated with the nerve are seen dorsal to this: f. fibres extending between the nerve and nuclei ventrally and the roof of the ventricle dorsally, the innermost fibres reaching l.c., which probably represents the locus caeruleus. B, section passing in the neighbourhood of the isthmus, 115 mm., to show the flattening down of the root, leaving the translaminar sulcus as the upper limb in a T-shaped cavity. 0, produced by adding several sections from an embryo of 100 mm., to show h. l., the (1?) homolateral roof fibres of the trochlear nerve, joining a separate arched bundle of (contralateral) fibres.
The plane of section is very oblique, and runs dorsally into the cavity of the mid-brain; t.l.s. translaminar sulcus.
Text-Figures
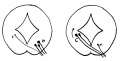
Fig 1
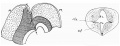
Fig 2

Fig 3
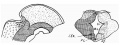
Fig 4

Fig 5
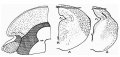
Fig 6
Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - Development of the region of the isthmus rhombencephali (1928). Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_Development_of_the_region_of_the_isthmus_rhombencephali_(1928)
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G