Paper - A morphological study of testicular descent
| Embryology - 30 Jun 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Wyndham NR. A morphological study of testicular descent. (1943) J Anat., 77(2):179-188.3. PMID 17104926
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
A Morphological Study of Testicular Descent
BY N. R. Wyndham, Department of Anatomy, University of Sydney
Much attention has been paid during recent years to the problem of testicular descent with special reference to hormonal influences. It was hoped that studies in this field would have thrown some light on the causation of the changes in position of the testis occurring during human development. But there are two obstacles to the fulfilment of this hope: first, there are gaps in our knowledge of the general topography of the
structures concerned with testicular descent and, secondly, though the same biological laws govern man and other mammals, anatomical studies of the latter are rather inconclusive since the structures are not identical in all types. An endeavour has therefore been made to trace the development from their first appearance of the structures concerned with the descent of the testis, using human material only.
Historical
Similar surveys have been made from time to time for the last one hundred and fifty years. Galen referred to the occasional abdominal position of the testis, and Vesalius described the processus vaginalis, but no work of value was published on testicular descent till that of several men working independently at about the same time. Of these investigators the most important were Haller (c. 1750), Camper (1756), Paletta (1777), Vicq d’Azyr (1784), and John Hunter (1786). Hunter’s contribution, in particular, is of great value and few of his observations have needed correction. Amongst those who have contributed more recently to our knowledge of the subject are Bramann (1884), Weil (1885), Lockwood (1888), Frankl (1895, 1900), Klaatsch (1896), Felix (1905, 1912), Berry, Hart (1909), and Moszkowicz (1935).
Material and Methods
Fifty specimens of human embryos were examined. The methods of examination varied according to the stage of development and the nature of the anatomical detail to be investigated. The younger specimens were stained in bulk with haematoxylin and eosin, and were studied in transverse and longitudinal sections cut at a thickness of 10 µm. Older specimens were mounted on celloidin and dissected, unstained, with or without the aid of a dissecting microscope. The findings obtained by dissection were confirmed by means of serial sections.
The subject is considered in four parts. The first deals with the position of the testis in relation to the abdominal wall and the early stages of development of the place inguinalis, and includes a study of embryos from 12 to 20 mm. C.R. length; the second part deals with the early development of the gubernaculum and processus vaginalis and other structures which finally take a share in testicular descent, and is concerned with embryos ranging from 20 to 50 mm. C.R. length; the third part deals with the appearance of the structures in specimens from 50 mm. to 23 cm. C.R. length; the fourth part deals with the final stages of descent as seen in specimens of 24 cm. C.R. length and over.
It is realized that the methods outlined above have the limitations that must always exist when vital processes are studied by the use of dead material, and that the mechanical forces at work in bringing about the changes in the relative positions of the various structures concerned can be only suggested and not proved by these methods. It may be presumed, however, that the whole development of this region is organized by the testis under the control of the pituitary gland, though this aspect of testicular descent cannot be considered in a study such as this.
Position of Testis and Early Development of Plica Inguinalis
During the greater part of embryonic life the testis lies at the caudal end of the abdominal cavity. Its position is associated with the development of the infra-umbilical part of the abdominal wall and is determined by the rearrangement of the constituents of the urogenital fold consequent upon the atrophy of the mesonephros.
In an embryo of 14 mm. it is represented by a thickening of germinal epithelium, 1.04 mm. long, lying in the long axis of the body between the suprarenal gland medially and the mesonephros, Wollfian and Miillerian ducts, and abdominal wall laterally.
In 20 mm. embryos the gonads can be displayed by dissection. In one specimen at this stage the right gonad was found to be 2.3 mm. long and the left 2.5 mm. They were situated between the large suprarenal gland and the abdominal wall and ventral to the metanephros. The shortest distance from the gonad to the definitive groin was 0.7 mm. on the right side and 1.0 mm. on the left. These measurements were confirmed by the dissection of other specimens of an approximately similar age.
In a 30 mm. embryo the right gonad was found to be 3.0 mm. in length and the left 2.8 mm. The shortest distance from the groin was 1.6 mm. At this stage the gonads were lying almost horizontally and were separated from the suprarenal gland by the metanephros.
Some writers, e.g. Hunter, Cooper (1845), Cleland ( 1856), Peters (1927), Keith (1933), and Hamilton Bailey (1936), have suggested that the testis is developed high in the abdomen in a position that would correspond with that of the kidney in the adult and that it then descends to the groin. Thus Peters writes of an ‘inneren Descensus’; Keith states that ‘The testis descends from the loins to the iliac fossa in the third month’; and Hamilton Bailey embodies in a figure these errors with regard to the early position of the testis.
Although its upper pole does correspond at first with the last thoracic segment, the gonad is never far from the groin. Bramann was the first to point this out, showing that the maximum distance between the testis and the groin in early stages is no more than 1.0 mm. Felix also has denied that there is any abdominal descent.
Though there is certainly a rapid increase in the distance between testis and kidney, this is not due to any downward movement of the testis but is accounted for almost completely by the rapid growth of the lumbar part of the vertebral column and the adjacent structures. This growth carries the metanephros upwards since it is free to slide up behind the peritoneum, while the testis, rendered relatively immovable owing to being suspended by its mesentery, is left attached to the groin. This fact was emphasized by Lockwood, who marked the relation of the testis to the acetabulum as a fixed point and noted that for some time their relative positions remained constant.
If there is any alteration at all in the distance between the testis and the groin it is an increase, as can be seen from the measurements of the embryos in this series given in Table 1. This increase in the distance between the testis and the groin has been referred to by Weil as a ‘preliminary ascent’.
The long axis of the gonad is at first parallel to that of the embryo, but quickly assumes an oblique and almost horizontal position; this is due partly to the atrophy of the mesonephros and partly to the increase in size of the suprarenal gland and kidney which apparently pushes the cranial pole of the gonad laterally. Pl. 1, fig. 1 shows the gonad lying parallel to the groin in a 49 mm. embryo.
The urogenital ridge, comprising the gonad, the mesonephros, and the Wolffian and Müllerian ducts, has four peritoneal folds developed in association with it; they are the Wolfiian mesentery, the mesorchium, the plica diaphragmatica, and the plica inguinalis.
The Wolflian mesentery, or urogenital mesentery, at first slings all the components of the urogenital ridge to the dorsal wall of the abdomen, and when the mesonephros atrophies it suspends the gonad and the ducts from the dorsal wall, and later becomes the mesentery of the testis and epididymis. The mesorchium slings the gonad to the mesonephros at first and later forms a connexion between testis and epididymis. The cephalic end of the mesonephros extends at first as far as the lower cervical segments. As its caudal part develops the cranial part atrophies but leaves behind it a thin strand that later on extends from the diaphragm to the cranial end of the Wolffian duct; it is known as the plica diaphragmatica. This fold was named by Kolliker (1849) the ‘Zwerchfellsband der Urniere’. Lockwood referred to it as the ‘plica vascularis’, since he believed that it subsequently contains the spermatic vessels.
Table 1 - Embryo Measurements
| Embryo | Crown Rump Length (mm) | Gonad (mm) | Groin distance (mm) |
|---|---|---|---|
| H 125 | 20 | 2.4 | 1.0 |
| H 181 | 20.6 | 2.8 | 1.1 |
| H 271 | 21 | 3.0 | 0.7 |
| H 408 | 22 | 2.3 | 1.0 |
| H 283 | 30 | 29 | 1.6 |
| H 434 | 38 | 3.4 | 1.5 |
| H 385 | 41 | 2.5 | 1.5 |
| H 35 | 42 | 3.0 | 1.8 |
| H 97 | 47 | 3.8 | 1.8 |
| H 226 | 49 | 4.0 | 1.8 |
| H 65 | 56 | 3.3 | 0.9 |
At its caudal end the mesonephros is attached to the caudal wall of the abdomen (for in early stages there is no true ventral wall caudal to the attachment of the body stalk) by a mesodermal fold which is probably a continuation of the nephrogenic cord, the unsegmented mesoderm in which the mesonephric units develop at a more cranial level. It cannot, however, be said for certain whether this mesodermal fold is derived from the mesoderm of the intermediate cell mass, i.e. the nephrogenic cord, or from the parietal mesoderm, for the attachment is just where these two mesodermal areas become continuous.
When atrophy of the caudal, as of the cranial, end of the mesonephros occurs this mesodermal connexion becomes thinned out and stretches not from the mesonephros but from the Wolffian duct to the caudal wall of the abdomen. Its point of attachment to the duct is where that structure bends forwards towards the cloaca in the lateral part of the genital cord, or, as Bramann describes it, at ‘der Stelle an welcher das Vas deferens aus dem Nebenhoden entspringt’.
This mesodermal fold is the plica inguinalis. It can be seen in P1. 1, figs. 2, 3 and 4, as a fold with a concave upper border lying medial to the umbilical artery and containing the Wolffian duct as that structure passes to join the urogenital sinus. This is the condition in a 12 mm. embryo. At the 14 mm. stage changes have taken place in the lower end of the urogenital ridge; the mesonephros is much smaller; the gonad is well defined and has its own mesentery; the Wolffian duct enters the urogenital sinus posteriorly rather than from the side and thus turns medially out of the plica inguinalis to reach the sinus; the plica inguinalis reaches the body wall to the lateral side of the umbilical artery while the Wolflian duct remains on its medial side, which means that the artery must cross beneath the plica from the lateral to the medial side. Moszkowicz has described an intermediate stage, in a 13.5 mm. embryo, in which the artery ran parallel to the fold and below it. In a 20 mm. embryo (Pl. 2, fig. 5) the plica inguinalis passes from the caudal end of the urogenital fold to the ventral wall of the abdomen. It is 0.42 mm. long and is attached proximally close to the Wolffian and Müllerian ducts at the caudal end of the fold. In a dissection of a 20 mm. embryo the umbilical artery was seen crossing beneath the inguinal fold from the lateral to the medial side, and the connective tissue round the artery was intimately fused with that of the inguinal fold.
As embryos from 30 to 50 mm. are examined the inguinal fold is found to become more cord—like and isolated from surrounding structures and it acquires a mesentery. Its upper end is attached to the Wolffian duct, as has been stated by Curling (1841), Weil, Lockwood, Frankl, Eberth (1904), Felix, and Moszkowicz. On the contrary, it has been described by Meckel (1820), Cooper, Bramann, and Klaatsch as being attached to the lower pole of the testis.
Gubernaculum Testis and Processus Vaginalis
The gubernaculum testis has been described by many previous workers as arising from the cells contained within the plica inguinalis. This fold has been referred to above as passing to the ventral abdominal wall, and it may be noted here that it reaches the body wall at a place where there are no muscle fibres, that its cells seem to pass in both a cranial and caudal direction in the abdominal wall, and that some of them pass to the base of the genital swelling and to the inner aspect of the thigh. The appearance of the inguinal fold and its contained cells, or, as it may now be called, the gubernaculum, is seen well in a section of a 20 mm. embryo (Pl. 2, fig. 6). The cells of which it is composed can be distinguished by their irregular arrangement and by being more condensed than those of the body wall. As they pass into the abdominal wall they lie between the rudiments of the rectus and oblique muscles of the abdomen, but in some specimens they seem to bring about a disordered arrangement of the oblique muscles, particularly of the transversus and internal oblique. But the cells of which the gubernaculum is composed are present in situ before the abdominal wall and its musculature are formed, and there is no evidence of its cells invading the lymphatic spaces of the body wall, as was described by Berry Hart (1909).
As the gubernaculum of a 20 mm. embryo is traced more caudally it is found to end indefinitely in the region of the genital swelling, not as a single strand but in a cone-shaped manner, the base being the insertion, i.e. it is attached to an area and not to a point. The dissection of the distal end of the gubernaculum in embryos of 12 or 14 mm. is very difficult owing to the loose friable nature of the strands of which it is composed and to the fact that they have no definite borders but merge into the surrounding musculature.
In older embryos dissection of the gubernaculum is easier, for it becomes- a more cord-like structure lying in the free edge of the plica inguinalis. When a 20 mm. embryo (Pl. 2, figs. 5, 6) is compared with an older specimen, for example of 42 mm. (Pl. 2, figs. 7, 8), it is found that in the latter the distal end of the gubernaculum appears to invaginate the abdominal wall and thus lies in a small pocket, to the dorsal wall of which it is attached by a short, thick ‘mesentery’. This little crescentic pocket is the beginning of the processus vaginalis.
The time at which this processus vaginalis makes its appearance is variable; Weil gives the end of the second month as the usual time; Bramann, and Lockwood, first noted it in the 14th week; Frankl, and Eberth, describe its presence in embryos of 28 mm.; in the present series it can be seen on both sides in an embryo of 40 mm., on one side in one specimen of 42 mm. and absent in another, absent in a female embryo of 47 mm., and present on the right side only in a male embryo of 49 mm. As serial sections of the 42 mm. embryo (Pl. 2, figs. 7, 8) are studied, it is found that though the processus vaginalis appears as a crescentic cleft it is in reality a blind pouch, open at its cranial end into the abdominal cavity, and lined with coelomic epithelium. It appears to surround the gubernaculum on all sides except dorsally, and it separates that structure from the fibres of the cremaster muscle laterally. In this embryo the vaginal process is 0.21 mm. long.
At this stage the gubernaculum contains non-striped muscle fibres which form a large part of its bulk, and as it is traced distally its outline becomes less distinct owing to the intermingling of its fibres with the constituents of the abdominal wall.
The above description differs from that of several previous workers, especially with regard to the relation of the gubernaculum to the processus vaginalis. The latter is said by Weil, Weber (1847), Meckel, Klaatsch, Frankl (1900), and Kirk (1936) to sink into and hollow out the gubernaculum, so that the gubernaculum is divided into two parts, a central core and a superficial part which becomes the cremaster muscle. There is no evidence to support this in the present series. On the other hand, the findings do support the descriptions of Bramann, Hunter, Cooper, and Lockwood, namely, that the processus vaginalis is a sac of peritoneum which develops on the ventral aspect of the gubernaculum and becomes covered with a layer of striated muscle from the internal oblique and transversus muscles, which layer is called the cremaster. The formation of the processus vaginalis is not due to the activity or proliferation of any of its parts, but is secondary to growth changes of the abdominal wall, as described by Felix.
Further Development of Gubernaculum Prior to Testicular Descent - Embryos of 50 mm to 23 cm
The position and relationships of the testis at the commencement of this period differ little from what is found at earlier stages.
The distance from the groin is approximately the same, though the marked angularity at the point of junction of epididymis and vas deferens gives the impression of downward traction.
The processus vaginalis keeps pace with the growth of the whole embryo; it is 1.8 mm. long in a 56 mm. embryo and 3.0 mm. long in an embryo of 120 mm. The inguinal canal is well defined in a 66 mm. embryo; it contains the processus vaginalis, the gubernaculum and the cremaster; its relations are as in the adult, but the deep inguinal ring is not well defined. Keith has denied the existence of an inguinal canal at this stage. There is no evidence of an ascending portion of the cremaster muscle as was described by Hunter, Lockwood, Eberth, and Kolliker.
The testis lies anterior to the epididymis to which it is joined by the mesorchium which gradually becomes shorter. The junction of epididymis and vas deferens is sometimes found at the mouth of the processus vaginalis. The gubernaculum increases only slightly in length during the period under discussion, but it attainsia much greater bulk particularly in the later stages, e.g. in embryosof 20 cm. The inguinal canal also increases in width, due almost entirely to enlargement of the cremaster.
In all embryos examined from the 6 to the 12 cm. stage the gubernaculum could be traced to the scrotum and in some cases to the pubic region and to the inguinal ligament also, but no fibres were found going to the perineum. During this period there are practically no striated muscle fibres in the gubernaculum, but they become prominent in later stages, at about 22 cm.
Towards the completion of the development of the parts concerned with testicular descent, i.e. in embryos of about 20 cm., one of the most outstanding features is the great bulk of the gubernaculum so that the testis has the appearance of a little knob perched on its summit; in fact, the growth of the gubernaculum is so great at this stage that it pushes the testis for several millimetres into the abdomen. This growth of the gubernaculum begins at the cephalic end and finally involves the whole structure; it is an intrinsic property of the gubernaculum and is independent of the growth of the cremaster. In none of the present specimens was there found an appearance such as Keith describes of the gubernaculum growing down as a solid cellular mass, with an actively growing cellular cap, into the subcutaneous tissue of the scrotum.
All observers agree that the gubernaculum does come to contain striated muscle fibres in later stages, and these are usually described as being situated peripherally in the gubernaculum and as being cremasteric in origin. Bramann has described a central core of connective tissue as being present in the gubernaculum, while van den Broek (1906) and others have compared-the whole structure with the conus inguinalis of some lower forms.
In the present series sections of a 22 cm. foetus show that below the level of the processus vaginalis there is an intermingling of gubernaculum and cremaster, but there is no evidence of any ascending cremasteric fibres contributing to the more proximal part of the gubernaculum. In these sections of the 22 cm. specimen, shown in P1. 3, figs. 9, 10 and 11, three histological entities are found, striated muscle fibres, unstriated muscle fibres, and blood vessels. The striated muscle fibres, which are well seen in P1. 3, fig. 11, lie with their long axis parallel to that of the gubernaculum and in the section they are cut obliquely. They grow from the scattered spindle-shaped cells which are found throughout the gubernaculum prior to this stage. The unstriated muscle fibres can be seen lying between the striated fibres. The blood vessels are numerous and tortuous (P1. 3, figs. 9, 10) and the great vascularity of the gubernaculum is characteristic of this stage, as noted by Weil.
As in the earlier stages, so towards the end of this period, the processus vaginalis keeps pace with the general growth of the foetus. It measures 9-9 mm. in length in a foetus of 20 cm. It is a flattened sac lying between the gubernaculum dorsally and the cremaster ventrally, and it is only at its cephalic end that it surrounds the gubernaculum completely, thus giving it a sort of mesentery. At this stage the vaginal process is found to be 10 mm. in advance of the testis in the inguinal canal, but this distance seems to be a very variable one. As can be seen from Table 2, the, testis doubles its size between the 12 and 20 cm. stages. Its distance from the groin varies.
Table 2. Statistics of the Dissected Embryos
| Embryo | Crown Rump Length (mm) |
Gonad (mm) |
Groin distance (mm) |
Gubernaculum (mm) |
Infra-umbilical belly wall (mm) |
Processus vaginalis (mm) |
|---|---|---|---|---|---|---|
| H 125 | 20 | 2.4 | 1.0 | 1.8 | 1.4 | — |
| H 181 | 20.6 | 2.8 | 1.1 | 1.9 | 1.6 | — |
| H 271 | 21 | 3.0 | 0.7 | 1.4 | 1.6 | — |
| H 408 | 22 | 2.3 | 1.0 | 1.8 | 2.0 | — |
| H 283 | 30 | 29 | 1.6 | 3.2 | 3.1 | — |
| H 434 | 38 | 3.4 | 1.5 | 3.5 | 5.2 | — |
| H 385 | 41 | 2.5 | 1.5 | 3.5 | 6.2 | 0.4 |
| H 35 | 42 | 3.0 | 1.8 | 4.0 | 6.8 | — |
| H 97 | 47 | 3.8 | 1.8 | 4.4 | 7.1 | — |
| H 226 | 49 | 4.0 | 1.8 | 3.8 | 7.2 | 0.9 |
| H 65 | 56 | 3.3 | 0.9 | 4.2 | 8.0 | 1.8 |
| H 219 | 66 | 4.0 | 1.2 | 6.6 | 11.0 | 2.4 |
| H 134 | 90 | 4.4 | 1.3 | 6.6 | 15.6 | 3.3 |
| H 388 | 94 | 3.8 | 1.5 | 5.8 | 16.0 | — |
| H 472 | 118 | 3.2 | 2.8 | 5.9 | 18.0 | 2.6 |
| H 310 | 137 | 7.0 | — | 7.5 | 20.0 | — |
| H 150 | 145 | 6.6 | 3.4 | 13.2 | 26.0 | 8.9 x 3.0 |
| H 528 | 195 | 6.4 | — | 16.0 | 28.0 | 9.9 x 5.1 |
| — | 220 | 7.0 | 6.5 | 21.0 | 34.5 | 11.5 x 4.5 |
| — | 230 | 9.0 | 2.0 | 27.5 | 36.0 | 12.5 x 5.0 |
| — | 240 | 10.9 | — | 4.9 | 46.0 | 25.0 x 8.5 |
| — | 250 | 10.0 | — | 1.9 | — | 24.5 |
| — | 260 | 11.0 | — | — | 44.0 | 21.2 |
| — | 270 | 12.3 | — | 0.7 | 42.0 | 28.8 x 5.6 |
The Anatomy of Final Descent - Foetuses of Over 24 cm
In no specimen of 23 cm. or less had descent begun, yet it was completed in every specimen, except one, of 24 cm. and over. Thus in the present series descent of the testis from the groin to the scrotum occurs at the beginning of the seventh month of intra-uterine life. Only normal material was used, and the scarcity of material in which intermediate stages of descent could be studied suggests that the process of descent is a rapid one.
The time at which the mouth of the processus vaginalis becomes closed is a very variable one. In twelve cases where both testes were in the scrotum the processus
vaginalis was patent on each side. Seiler (1817), Klaatsch, Frankl, and Moszkowicz have expressed the opinion that the structure which fills the inguinal canal prior to
descent is the conus inguinalis which, by the contraction of its own musculature, shortens and turns itself inside out like the finger of a glove, thus forming a cavity
which is lined with peritoneum. This conus inguinalis supplants the gubernaculum in the plica inguinalis, leaving only a core of embryonic connective tissue and striated muscle. As has been stated above, the structure that fills the inguinal canal is not the equivalent of the conus inguinalis.
Frankl (1900) and Broman (1911) have shown that descent is accompanied by a
shortening of the gubernaculum. This finding is supported by measurements in the present specimens, in which it was found that the distance between the testis and the bottom of the processus vaginalis remains approximately constant. In every case the
main attachment of the gubernaculum was to the bottom of the scrotum but was rather
loose, so that it was easily torn in dissection. In one case there was an additional
attachment to the pubis. By the time that the testis has reached the bottom of the
scrotum there is no sign of the gubernaculum.
Factors causing descent. It is outside the scope of this paper to discuss the reasons
why testicular descent does occur in practically all placental mammals. Hunter suggested that the testis was in some way responsible for organizing its own descent, and
modern endocrinological research, according to Engle (1932) and others dealing with
the subject of cryptorchidism, supports this idea. At the same time Hunter admitted
that the causes were probably mechanical. It seems likely that atrophy of the mesonephros, the rearrangement of the structures on the posterior abdominal wall, the de-
velopment of the infra-umbilical portion of the abdominal wall and the descent of
the testis are all closely related.
Cleland, Kolliker, and Berry Hart suggested that descent could be accounted for by differences in the rate of growth of structures related to the testis. Berry Hart
considered also that the inguinal canal grew up to envelop the testis. Measurements made on the present series do not at all support this conception. Descent seems to
take only a short time, and just before it occurs the testis is 3 cm. from its final
position, a distance too great to be covered rapidly by any upward growth of the
inguinal canal. Meckel suggested that the testis is pulled down by scar contraction
of the gubernacular connective tissue. But there is very little connective tissue in
the gubernaculum.
Furthermore, an objection to any theory of gubernacular contraction is the fact
that the attachment of the distal end of the gubernaculum is a very slender one and
is made to a mobile area of skin. The attachment of its proximal end in the abdomen
is much more secure, and so any contraction would tend to pull the scrotum up rather
than the testis down.
The intense vascularity, the increase in size prior to descent, and the fact that the
striated muscle fibres of the gubernaculum lie in the plane of the inguinal canal must
all have some significance. It should also be noted that the testis becomes more
mobile just before descent. It was ‘Bell (1826) who pointed out that the vessels of the
testis lengthen so that they become ‘elegantly tortuous’. Failure of such lengthening
would prevent descent.
Finally, it should be emphasized that the testis has a mesentery and that it is not a retroperitoneal organ like the kidney free to slide down the abdominal wall behind the peritoneum.
Summary
- An account is given of the development of the structures associated with descent of the testis in embryos ranging in size from 12 mm. to 27 cm. C.R. length.
- The testis is never far from the groin. Up to the 90 mm. stage the average distance is 1-3 mm. Between the 20 and 50 mm. stage the testis shows a preliminary ascent. It doubles in size between 12 and 20 cm. The final descent is rapid and occurs between the 23 and 24 cm. stages, i.e. early in the seventh month.
- In a 20 mm. embryo the testis lies vertically, but by the 30 mm. stage it is almost horizontal, and it is parallel to the groin at 50 mm. These changes are due partly to growth of the suprarenal gland and partly to atrophy of the mesonephros.
- Atrophy of the mesonephros causes a thinning out of the mesentery which originally slings it. The cranial part becomes the plica diaphragmatica, the middle part forms a part of the mesorchium, and the caudal part forms the plica inguinalis. The Wolffian duct at first runs in the plica inguinalis to the cloaca.
- The gubernaculum develops in the substance of the plica inguinalis. Its cranial end is attached to the Wollfian duct and not to the testis; its caudal end flares out and is attached to the ventral wall of the abdomen where it spreads up and down, to the groin, to the region of the genital swelling, and to the thigh.
- The gubernaculum becomes greatly enlarged about the 20 cm. stage. It contains striated and unstriated muscle fibres and is very vascular. The muscle fibres are an intrinsic development, and there is no evidence that the muscles of the abdominal wall, particularly the cremaster, contribute to its formation.
- At about the 20 cm. stage the gubernaculum becomes so large that it pushes the testis for some distance into the abdomen. The gubernaculum does shorten during testicular descent, but there is no evidence that it pushes down as a cap into the scrotum.
- The processus vaginalis can be seen at the 42 mm. stage as a small crescentic slit 0-21 mm. in length. It passes down the inguinal canal, which is well defined at 66 mm., between the gubernaculum dorsally and the cremaster ventrally.
- The present study does not support the view that the inguinal canal grows back towards the testis. The flimsy attachment of the gubernaculum to the scrotum is an obstacle to it exerting much traction on the testis.
The material has been provided through the courtesy of Professor C. W. Stump, of the Department of Embryology and Histology, the University, Sydney, from a large collection of human embryos possessed by that department.
References
BAILEY, H. (1936). Diseases of the Testicle. London.
BELL, J. & C. (1826). Anatomy and Physiology of the Human Body. London.
BRAMANN, F. (1884). Arch. Anat. EntwGesch.
BROEK, A. J. P. VAN DEN (1906). Petrus Camper. Dect. 4-.
BROMAN, I. (1911). Normale und Almorme Entwiclclung des M enschen. Wiesbaden.
CLELAND, J. (1856). The Mechanism of the Gubernaculum Testis. Edinburgh.
COOPER, A. (1845). Observations on the Structure and Disease of the Testis. London.
CURLING (1841). Land. Med.‘ Gaz. g
EBERTH (1904). Die M dnnlichen Geschlechtsorgane.
ECCLES, W. (1903). The Imperfectly Descended Testis.
ENGLE, E. T. (1932). Endocrinology, 16, 506.
FELIX, W. (1905). Handb. d. vergl. u. exp. Entwicklungslehre d. Wirbeltiere, 5. Jena.
Felix W. The development of the urinogenital organs. In Keibel F. and Mall FP. Manual of Human Embryology II. (1912) J. B. Lippincott Company, Philadelphia. pp 752-979.
FRANKL, O. (1895). Arch. Anat. Physiol., Lpz.,
FRANKL, O. (1900). S.B. Alcad. Wiss. Wien, 109, 3.
GALENUS, CLAUDIUS (1833). Medicorum Graecorum Opera. Kuhn.
HART, D. BERRY (1909). J. Anat., Lond., 4-3 and 44-.
HUNTER, J. (1786). Observations on Certain Parts of the Animal Economy. On the Situation of the Testis in the Foetus.
Keith A. Human Embryology and Morphology. (1902) London: Edward Arnold.
KIRK, J. (1936). J. Anat., Lond., 71.
KLAATSCH, H. (1896). Morph. Jb. 16.
KOLLIKEN, A. (1849). Z. gee Anat. 1. Z. Amt. EntwG’e.sch.
Locxwoon, C. B. (1888). J. Anat. Physiol. 22.
MECKEL, FR. (1820). Handbuch der menschlichen Anatomic.
Moszxowrcz, L. (1935). Z. gas. Amt. 1. Z. Amt. EntwG’e.sch.
PETERS, K. (1927). Hundbuch der Anatomie des Kindes. Miinchen.
SEILER. (1817). Observationes nonnullae de testiculorum ex abdomine in serotum descensu. Leipzig.
VICQ D’AzYR, F. (1784). Recherche sur la, Structure et la Position ales Testicles. Paris.
WEBER, E. H. (1847). Arch. Anat. Phyeiol., Lpz.,
WEIL, C. (1885). Uber den Descenaus testiculorum.
Explanation of Plates
Plate 1

Fig. 1. Dissection of embryo H 226 of 49 mm. x6. A ‘well-formed pars abdominalis gubernaculi is evident. The interstitial part has also been dissected, the outline being less clearly defined. l, gonad; 5, gubemaculum; 6, bladder.
Figs. 2-4. A series of transverse sections through the caudal end of the mesonephric ridge of a 12 mm. embryo H 323. x 35. A, Wolflian duct; B, umbilical artery.
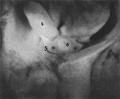
Fig. 1. Dissection of embryo H 226
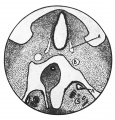
Fig. 2. Mesonephric ridge embryo H 323.
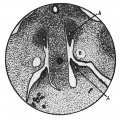
Fig. 3. Mesonephric ridge embryo H 323.
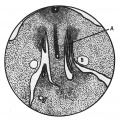
Fig. 4. Mesonephric ridge embryo H 323.




Plate 2

Fig. 5. Transverse section through caudal region of a 20 mm. embryo H 304. x 20. b, plica. inguinalis; g, pubic anlage.
Fig. 6. The same section as above. X120. Showing the detailed relations of the cells of the definitive gubemaculum to the belly wall.
Fig. 7. Transverse section of a 42 mm. embryo H 42. x 11. a, testis; b, gubernaculum.
Fig. 8. Transverse section of a 42 mm. embryo H 42. x 100. c, cremaster muscle; d, processus vaginalis.
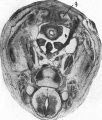
Fig. 5. Transverse section through caudal region of a 20 mm embryo H 304

Fig. 6. Gubernaculum detail 20 mm embryo H 304
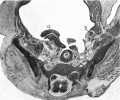
Fig. 7. Transverse section of a 42 mm embryo H 42
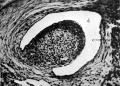
Fig. 8. Transverse section of a 42 mm embryo H 42




Plate 3

Figs. 9-11. Sections through the gubernaculum of a 22 cm. foetus. The magnifications are 120, 250 and 550 respectively.

Fig. 9.
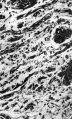
Fig. 10.

Fig. 11.



Wyndham NR. A morphological study of testicular descent. J Anat. 1943 Jan;77(Pt 2):179-188.3. No abstract available. PMID 17104926
Cite this page: Hill, M.A. (2026, Haziran 30) Embryology Paper - A morphological study of testicular descent. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_A_morphological_study_of_testicular_descent
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G