Paper - A histological investigation of the development and structure of the human lung
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Cooper ERA. A histological investigation of the development and structure of the human lung. (1938) J Pathology 47: 105-114.
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
A Histological Investigation of the Development and Structure of the Human Lung
Eugenia R. A. Cooper, M.D., M.Sc., Lecturer in histology in the University of Manchester.
Plates XXV.-XXXI.
Introduction
The diffculties encountered in describing the histological structure of the lung to students and the variable accounts occurring in the literature were responsible for this investigation. It was hoped that by studying the development during its pre— and post—natal stages, some light might be thrown on the controversy regarding the structure of the ultimate elements of the lung—the alveoli— and the relationship between them and the vascular supply. In his recent book, Miller (1937, p. 56) has given a critical review of the literature to date. As he states, the main difference of opinion lies in the structure of the alveolar walls. This has varied from descriptions which maintain that there is a complete epithelial lining to the alveoli, to accounts which question the presence of a continuous cellular lining, maintaining that the air-containing alveoli are limited by capillary plexuses (vide Maximow and Bloom, 1934; The latter view does not appear to be compatible with histological, pathological or embryological evidence. An arrangement of free spaces communicating with the exterior and in contact with thin—walled capillaries does not accord with histological findings in general. On the other hand the disposal of free blood in “ open ” circulation around and between alveoli possessing a complete wall of epithelial cells has a parallel in other situations. For example, an open circulation occurs in the placenta, where gaseous exchange is permitted in foetal life and a similar vascular arrangement is described in the marrow and the liver.
If the development of the lung is compared with that of the liver, it will be observed that both organs arise as entodermic buds from the primitive gut; moreover, both grow out into vascular mesoderm as “ glands ” with a duct system. In addition, in both cases, the duct system becomes subordinated to the terminal cellular “elements ” which, in the lung, form the alveoli, and in the liver, the liver lobules. finally, in both viscera, the purpose of these terminal elements is to effect exchange of substances in the blood, gaseous in the lung, products of digestion in the liver. Many histologists, including the author, believe that the blood in the liver is in free and open circulation between and around the hepatic cells and it is conceivable that a similar disposal of the blood occurs in relation to the alveolar cells in the lung.
Material studied. The human lungs studied ranged from those of a 10 mm. embryo to those of a child of 5 years. The details are given in the accompanying list.
List of embryos examined

As the investigation was entirely histological and confined to cellular detail, paraffin sections of uniform 6 ,1, thickness were prepared and general histological stains were used.
In order to corroborate the findings in human material, a series of mouse lungs was also examined. This included the last seven days of gestation and the first seven days after birth. In addition, lungs of four full term foetal cats were investigated.
In a section through the centre of the thoracic region in a 10 mm. human embryo, a circumscribed mass of simple mesoderm projects on either side from that surrounding the oesophagus and dorsal aorta into the posterior portion of the pleuro-peritoneal cavity. The thin limiting membrane lining this cavity is reflected over the mesodermic mass (called hereafter the pulmonary mesoderm), and represents the visceral pleura.
The mesoderm itself consists of angular branching cells, the processes of which join to produce a simple meshwork. Here and there a few cells have become narrow and elongated, mapping out a small cavity believed to represent the differentiation of a blood vessel in situ. Within some of these blood vessels nucleated erythrocytes are present.
At its entrance into the pulmonary mesoderm, the bronchus possesses a Well formed cavity surrounded by tall columnar cells primary bronchial branchings. X40. in two or three layers. The cytoplasm is pale and vacuolated and its peripheral aspect is in contact with a hyaline basement membrane around which the mesodermic cells are concentrated. A curious feature of the cells lining the bronchus is the internal position of the nuclei which appear adjacent to the lumen (Fig. 1).
Plate XXV

Fig. 1. Left lung 10 mm embryo. X40.
Fig. 2. Portion of lung of 55 mm embryo.
Fig. 3. Primary bronchus of first generation A wedge-shaped area extends inwards from on right, primary of second generation on left the margin. This indicates grouping of 55 mm. embryo. X 300.
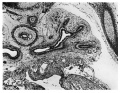
Fig. 1. Left lung. 10 mm embryo.
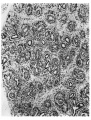
Fig. 2. Portion of lung of 55 mm embryo.
![Fig. 3]]. Primary bronchus of 55 mm embryo.](/embryology/images/thumb/9/9d/Cooper1938_fig03.jpg/96px-Cooper1938_fig03.jpg)
Fig. 3]]. Primary bronchus of 55 mm embryo.


![Fig. 3]]. Primary bronchus of 55 mm embryo.](/embryology/index.php?title=File:Cooper1938_fig03.jpg)
The main entering bronchus has already branched and the divisions are slightly smaller than the entering bronchus. They will be referred to as the primary bronchial branches, indicating that they have arisen from the main bronchus and are identical in structure.
In a 30 mm. embryo, differentiation of the cartilage plates is apparent in the two main bronchi, which present the characteristic columnar cell lining.
At 55 mm. the lungs measure 10><3 mm. in section. Two distinct lobes comprise each lung and the extent of the bronchial branching is much greater than in the earlier embryos. All the branches are regarded as primary branches since they are direct outgrowths of the original bronchus. They will persist as bronchial passages and will not become “ respiratory.” A recognisable grouping of the primary bronchial branches suggests that each group arises from one of the original divisions of the main bronchus (Fig. 2).
In some of the primary branchings the tall vacuolated lining cells occur in two or three layers and appear heavily stained ; others possess only a single layer of columnar cells. It is believed that the former represent the first generation of branches derived directly from the main bronchus While the latter belong to a second generation which hasarisen from the first generation (Fig. 3).
In a specimen of 60 mm. the distinction between the first and second generations of primary bronchial branches is even more clearly demarcated. In many positions sharply defined capillaries are recognisable, consisting of a minute ring with a crescentic nucleus at one aspect and containing a single red blood corpuscle. Throughout the section isolated erythrocytes are seen within spaces bounded by processes of mesodermic cells (Fig. 4). It is believed that these blood cells have developed in situ and will stimulate the mesodermic cells in the vicinity to form a channel for them.
Another example at this stage presents a clearer definition of the first generation of primary bronchial branches, namely the folding or pleating of the lining epithelial cells. Furthermore, the pulmonary arteries associated with the first generation of primary bronchi have acquired a definite wall of concentric elongated mesodermal cells (Fig. 5). Lobulation of the lung is clearly recognisable. A primary bronchial branch of the first generation surrounded by many primary branches of the second generation comprises a “ lobule ” which is separated from adjoining lobules by loose stroma. Many isolated erythrocytes are present in the latter.
In the lungs of 7-9 cm. embryos a further distinction of the primary bronchial branches is discernible, namely the “ interlobular ” bronchial branches. These are the initial branches derived from the main bronchus entering the lung at the hilum. It is from these that the primary branches of the first generation arise, and these in their turn give origin to primary branches of the second generation. The latter are of smaller calibre and disseminated throughout the mesoderm, and many of the ultimate ones are not as yet cavitated. Fig. 6 illustrates a cavitated and a non—cavitated primary bronchial branch of the second generation. In all cases the lining cells are columnar with their nuclei adjacent to the lumen.
Lungs from 11 and 11-5 cm. embryos exhibit an advance in development. The lumina of the bronchial branchings are much wider and the nuclei of the columnar cells lining them seem to have acquired a more central position. Many well formed capillaries are seen, but free erythrocytes are present in the mesodermic meshwork.
In lungs from 12-14 cm. embryos, a change occurs in the second generation of “primaries.” They lose their round or oval shape and become more widely opened up. In addition, further extensions are thrust out, giving the structure an irregular shape. This metamorphosis is accompanied by a change in the lining epithelial cells, which are seen to be pale and cubical, with central nuclei. These extensions derived from the primaries of the second generation will be designated “secondary branchings,” and are recognisable by the change from a columnar to a cubical type of lining cell (Fig. 7). Each bronchial branch is encircled by capillaries, but many free blood cells are present. Lymphatic channels are seen alongside the pulmonary blood vessels.
The modification of the second generation of primaries into “secondaries” is almost complete in the lung of an embryo of '15-4 cm. length, 73.e. the primaries of the second generation are mostly dilated, possess extensions, and are lined by pale vacuolated cubical cells. In addition, the outline of the secondaries is irregular, showing further diverticula instead of the neat appearance seen at a somewhat earlier stage. These additional structures are a new generation created by the secondaries and are designated “ tertiary ” bronchial branchings; they arise on all aspects of the walls of the secondaries (Fig. 8). Like the latter, the tertiaries are lined by pale cubical cells with central round nuclei. At this stage it is necessary to point out that the lining of all bronchial branches, whether primary, secondary or tertiary, is continuous and complete.
Plate XXVI

Fig. 4. Single erythrocyte (A) surrounded by mesodermic cells. 60 mm embryo. X 700.
Fig. 5. On right is a “primaries ” of the first generation, with cartilage. On the left are “ primaries ” of the second generation. 68 mm embryo. X 125.
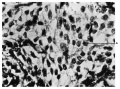
Fig. 4. 60 mm embryo.
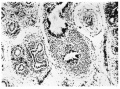
Fig. 5. 68 mm embryo.


‘ whorled ” pulmonary artery. In the centre are the tertiary bulgings approach each other, but they are separated from their neighbours by mesodermal stroma. Capillaries are numerous at this age and are situated in contact with the cells lining the tertiary bulgings, but many free corpuscles are visible. In some situations they are seen amongst the cells lining the bronchial branches. This is no doubt due to the section having passed in a position where a capillary or a free corpuscle lies between two tertiary diverticula.
At 16 cm. the lumina of the primary and secondary bronchial branches are much larger than in previous sections, but the lining cells form a complete and continuous single layer. There is an increase in the formation of tertiary branches. As yet there is no change in the vascular arrangement, many capillaries and free blood cells occurring in the same field.
The histological appearance of lungs from foetuses of 17-18 cm. length much more nearly resembles that of the post—foetal lung than heretofore. There is greater dilatation of the secondary and tertiary portions, so that the walls of adjacent tertiaries are almost in contact with each other (Fig. 9). A minimum of mesodermic stroma intervenes between the tertiary branchings and contains capillaries and free blood corpuscles, both of which indent the walls and further distort their shape. flattening of the cells lining the tertiaries is visible, especially where a capillary or corpuscle is in contact with them. Thus a rectangular, flattened or attenuated condition of the cells is recognisable in foetal life before the lung is inflated by air. An important histological point is that the lining of all branches, Whether primary, secondary or tertiary, is continuous and complete ; however much distortion is created by capillaries or free erythrocytes abutting on the walls, the cellular lining of the latter is never interrupted (Fig. 10). As two adjacent tertiaries dilate, the apposed portions of their walls are brought into closer contact, with only a capillary or a few free blood cells interposed. In this Way, “ septa ” are produced which demarcate one tertiary branching from another (Fig. 11).
Judging from the available sections, there does not seem to be any doubt about the presence of free erythrocytes in the mesodermic meshwork. It is suggested that their presence is not accidental, but that they belong to a free “ open ” circulation between the capillaries and the venous return.
Lungs from foetuses measuring 18-20 cm. exhibit increase in size owing to the extensive formation of tertiary buddings. In addition, many more of the secondary branchings are widely dilated, and in sections they are visible to the naked eye. A complete continuous lining of epithelial cells throughout the branchings is maintained. The cells in the primaries are columnar, in the secondaries and tertiaries cubical or rectangular. Lungs from foetuses of 20-31 cm. length consist of masses of closely packed tertiary branchings lined by cubical or rectangular cells. The connection between the primary branchings and the secondaries and their numerous tertiaries cannot now be recognised so easily, owing to the formation of successive generations of tertiaries and their consequent removal from the parent bronchial branch. The Vascularity is very rich, and both larger blood vessels and capillaries are engorged with blood. The capillaries and innumerable free corpuscles occupy the mesoderm between the tertiary branchings and come into contact with their lining. The mesoderm is rendered less obvious by the plexus of capillaries and free corpuscles. The primary branchings, however, still remain characteristic with their tall columnar lining. A secondary branching can be identified with certainty only when its continuity with a primary branching is visible. In such cases the change from the columnar cells of the primary to the cubical or rectangular type in the secondary is still noticeable (Fig. 12).
The opening up of the tertiary branchings approximates their walls and obscures the mesoderm. In addition there is a flattening of the lining cells. It is not difficult to find examples of these flattened “alveolar plates,” as many detached ones occur in all parts. When seen on the flat they are round, cubical or rectangular in shape, with a pale, round, central nucleus with dark nucleolar spots. If viewed laterally they appear as long spind1e—shaped cells with similarly shaped nuclei (Fig. 13).
The vascularity is intense at term, and capillaries with their crescentic nuclei are conspicuous around the tertiary branchings or alveoli; in addition, large numbers of free red corpuscles occur between adjacent alveolar walls. Fig. 14 illustrates a surface view of the flattened cells forming an alveolus. Blood cells within a capillary as well as free corpuscles are seen, from which it is suggested that beyond the capillary plexus there is a free circulation of the blood prior to its return into the venous system. The fact is borne out that the walls of the tertiary branchings or alveoli remain intact, with a continuous and complete lining of rectangular and flattened cells, despite the indentation of many of them by capillaries and free blood cells (Fig. 15).
An interesting appearance was presented in the lungs of four of the full term still—born foetuses. The central parts of the section were entirely filled with blood and it was impossible to distinguish the alveoli. In fact the intralobular bronchi and bronchioles also contained blood. At the periphery, however, the alveoli were empty and exhibited irregular walls formed by cubical or flattened cells indented by capillaries or free red blood cells (Fig. 15).
This engorgement of the lungs is no doubt associated with the still birth, but whether a contributory cause or a result cannot be surmised. Judging from histological evidence, one fact is suggested with regard to the pulmonary circulation, and that is the establishment of a free and open circulation beyond the capillary field. It appears as though the blood permeates around and between the alveolar walls, and is by them guided into the thin—walled venous channels.
Plate XXVII

Fig. 6. Cavitated and non-cavitated
Fig. 7. Primary and secondary bronchial branchings showing change in type of cells from columnar to cubical. P = primary branching; S = secondary branching. 12-6 cm. embryo. X 200.
Fig. 8. Primary (P). secondary (S) and tertiary primaries of the second generation. (T) bronchial branchings. 15-4 0111. embryo. 85 mm. embryo. X 450. X 100.
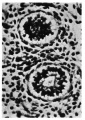
Fig. 6.
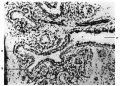
Fig. 7.
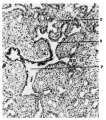
Fig. 8.



Lungs from an anencephalic foetus which had breathed were obtained and represented the earliest post-natal stage. They were much larger in size than those of the still—born foetuses, and to the naked eye the sections presented the characteristic honeycomb appearance. The extensive distension of the alveolar ducts (secondary branchings), alveolar sacs and alveoli at once distinguishes a lung which has breathed from one which has not. As to the continuity of the cells lining the alveoli there appears to be no doubt, in spite of the additional attenuation occurring when air distends the lungs (Fig. 16).
The next post-foetal stage is represented by the lungs of infants during the first post—nata1 year who died from bronchopneumonia. Although pathological, they are included, since the continuous cellular lining of the alveoli is unquestionable (Fig. 17). As pointed out by Miller (pp. 59-61) an effusion of fluid causes the cells to become swollen and enlarged and they are rendered much .more easily visible. Lung units are discernible and typical in their architecture. As regards the blood supply, the capillaries are not nearly so much in evidence as they are at full term, but free corpuscles in direct contact with the cells lining the alveoli are readily perceived throughout the section (Fig. 18). Other examples of the child’s lung corroborated these observations.
To substantiate the findings in the human lung, tissue from mice was examined during the last seven days of gestation and the first seven days after birth. The appearances in the foetal lungs are similar to those found in the human. The lungs of a foetal mouse killed just after birth are particularly interesting since there is an extravasation of blood into the alveoli similar to that seen in some still-born human foetuses. The lining of the alveoli is seen to be continuous in the alveoli not choked with blood and the presence of an open circulation is recognised. At two days after birth the extravasated blood has disappeared from the alveoli, the intact and continuous lining of the alveolar walls is visible and a free circulation of blood is apparent. In the mouse of three days, many lung units are discernible and all alveoli possess a complete lining of flattened cells (Fig. 19). The open circulation of blood is visible in the sections. Similar conclusions Were drawn from the examination of the lungs of full—time foetal cats.
Discussion
There is a good deal of confusion in the nomenclature of tlie constituent parts of the lung unit. On tracing the development, each unit is seen to develop in three stages, primary, secondary and tertiary. What appears to be the simplest and most convenient series of terms is one that will bring to mind Which portions arise from the original primary, secondary and tertiary branchings.
It is believed that the non—respiratory bronchial air passages end in the terminal bronchioles, each of which maintains its columnar lining cells. Beyond this the passages are “ respiratory ” in function and are beset with bulgings or alveoli. Thus a descriptive term is required which will imply that this is the case, and the selected one is “alveolar.” Since these alveolar passages arise as secondary and then tertiary branchings, the most appropriate designations are alveolar ducts (secondaries) and alveolar sacs (tertiaries) respectively. From both these passages, which of course are continuous With each other, the final bulgings or alveoli arise as further generations of tertiaries. Miller (p. 40) implies that the terminal bronchiole continues into the respiratory bronchioles, which in their turn communicate with alveolar ducts. The latter lead into spherical cavities, the atria, beyond which lie the alveolar sacs and alveoli. In the available sections, the spherical atria cannot be recognised.
It is not possible in a histological investigation to estimate the number of branchings accurately, but it is clearly elicited that increase in the amount of lung depends on the tertiary branchings. So far as can be judged, it seems that the non—respiratory bronchial passages (from the main entering bronchus to the terminal bronchioles) arise during the first half of foetal life (Le. to about 15 cm. in length). Subsequent development depends on the formation of respiratory alveoli. These originate in relation to secondary and tertiary bronchial branchings derived from the non—respiratory passages. It is the generations of the final tertiary branchings which add to the amount of lung tissue. If this be correct, then here is an explanation of the increase in the capacity of the lung which occurs in athletes, singers, Wind instrumentalists, etc. If, during pre—natal life, infancy and childhood, the lung increases by the formation of generations of new alveoli (tertiaries), there is every reason to believe that a similar increase can be stimulated as the necessity arises for greater air capacity. Possibly a limit is reached beyond which no further development of alveoli is possible, either on account of the restriction created by the chest Walls or the inability of the alveolar cells to divide. After that there is a breaking down of the septa between adjacent alveoli of the same lung unit, leading to the condition recognised as emphysema.
Plate XXVIII

Fig. 9. Prima.ry (P), secondary (S) and tertiary (T) bronchial branchings. 18 (am. embryo. X 100.
Fig. 10. Loose erythrocytes and capillaries in the walls of tertiary bronchial branchings. Cap. : capillary ; R.B.C. 2 erythrocyte. 17'5 cm. embryo. X450. PLATE XXIX.

Fig. 9.

Fig. 10.


Plate XXIX

Fig. 11. “ Septum " between two tertiary bronchial branchings. Cap. = capillary; R.B.C. = erythrocyte. 18 cm. embryo. X650.
Fig. 12. Primary, secondaries and tertiaries. Approaching full term. X 80.
Fig. 13. Tertiary branchings showing continuous lining. A detached cell is seen at A. Cap. = capillary; R.B.C. = erythrocyte. Approaching full term. X450.
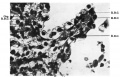
Fig. 11.

Fig. 12.
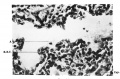
Fig. 13.



Plate XXX

Fig. 14. Surface view of alveolar cells. Cap. = capillary; R.B.C. : erythrocyte. Full term embryo. X IVU.
Fig. 15. Flattened and cubical cells lining alveoli. A : flattened cells; B : cubical cells. Full term embryo. X 700.
Fig. 16. Cells lining the alveoli in the lung of an anencephalic full - term foetus which had breathed. Note continuity of cellular lining of alveolus marked X. X300.
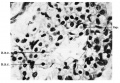
Fig. 17. alveolar cells Full term embryo
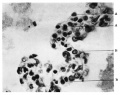
Fig. 18. Flattened and cubical cells Full term embryo
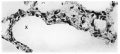
Fig. 16. alveoli term foetus



Plate XXXI

Fig. 17. Swollen cells lining an alveolus in the bronchopneumonic lung of an infant aged 12 weeks. X 630.
Fig. 18. Cells lining alveoli in the lung of an infant aged 8 months. R.B.C. = erythrocyte. X700.
Fig. 19. Lung of a three-days-old mouse showing continuous lining of cells in the alveoli. R.B.C. = erythrocyte. X625.

Fig. 17. infant aged 12 weeks
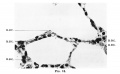
Fig. 18. infant aged 8 months

Fig. 19. three-days-old mouse



The most controversial point in regard to the structure of the lung is the presence or absence of a complete epithelial wall in the alveoli. The present study has shown that a continuous lining occurs throughout the bronchial branching in pre— and early postnatal life. This is in agreement with the recent findings of Gazayerli (1936), but not with those of Barnard and Day (1937). The recognition of a continuous wall is most difficult in still-born foetuses at full term when the lungs are engorged with blood. Even then, however, by washing out the blood and examining the alveoli under the pleura, the continuity is seen to be maintained. As pointediout and illustrated by Grazayerli (1936) and Miller (1937, pp. 59-61), the continuity of the alveolar lining is undeniable in pathological lungs where a small amount of exudate has caused a swelling of the cells and has raised them from the surrounding supporting tissue. The difficulty encountered in recognising a complete alveolar lining in adult lungs may be due to the partial destruction of the cells consequent on the chronic or subacute pulmonary conditions which affect so many individuals.
Kolliker and Eberth (both quoted by Miller) describe the alveolar walls as consisting of islands of epithelial cells alternating with non—nucleated plates (Kolliker) and meshes of capillaries (Eberth). The distortion of the alveolar walls by the capillaries or blood chains and the attenuation of the alveolar epithelium over these structures gives the appearance of islands of cells alternating with capillaries. The present research, however, reveals the continuity of the cells over the capillaries. Colberg (quoted by Miller) observed the elongation of the alveolar cells without interruption of their continuity, and he described the lining as a “ complete membrana epithelica.” The final problem in relation to the lungs is the nature of the pulmonary circulation through them. All observers are agreed that the bronchial blood flow is nutrient to the “ non-respiratory ” passages and does not communicate freely with the pulmonary system. As far as can be discovered in the available literature, observers are unanimous in describing a rich plexus around the alveoli. On embryological and histological evidence afforded by the present research, it is suggested that a free circulation of blood occurs between the pulmonary capillaries and the venous radicles. The alveolar lining cells constitute the walls of the minute interalveolar channels through which the blood flows. Therefore only one layer of cells is interposed between the alveolar air and the blood. These cells have been shown to be primarily “alveolar ” but it is suggested that they establish a continuity with endothelial cells lining the capillaries on the one hand and the venous radicles on the other.
Conclusions
1. Three generations of branches arise in succession from the main bronchus entering the pulmonary mesoderm. These are described as primaries, secondaries and tertiaries. The primaries become the non—respiratory bronchi, bronchioles and terminal bronchioles. The secondaries mark the commencement of the respiratory portion of the lung and constitute the alveolar ducts. The initial tertiaries form the alveolar sacs and further generations of tertiaries become the alveoli.
2. A change from columnar to cubical cells lining the bronchial branches demarcates the non—respiratory from the respiratory portion of the developing lung.
3. On histological evidence it is shown that the lining of all bronchial branchings persists as a continuous layer of cells albeit they become attenuated.
4. In all pre— and post—natal lungs, free red blood corpuscles in addition to capillaries appear to be in Contact with the outer surface of the lining of the bronchial branches and alveoli. The appearance of these red cells is not accidental, and it is suggested that a free circulation occurs between the arterial capillaries and the venous radicles. The cells lining the tertiary branchings or alveoli constitute also the lining of the interalveolar spaces through which the blood passes from capillary to venule.
The author wishes to express her thanks to Professor Lucas Keene for the loan of her remarkable series of foetal lungs, to Professor Dougal, Dr Evans and Dr Stent for material, to Professor Stopford and Professor Wood Jones for advice and criticism and to Mr Gooding for the kind and skilful manner in which he has prepared the sections and photographs.
References
BARNARD, VV. G., AND DAY, T. D. this Journal, 1937, xlv. 67.
GAZAYERLI, M. E. . . . . . this Journal, 1936, xliii. 357.
MAXIMOW, A. A., AND BLOOM, W. A textbook of histology, 2nd ed., 1934, Philadelphia and London, p. 458.
MILLER, VV. S. . . . . . . The lung, London, 1937, pp. 40, 56 and 59-61.
Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - A histological investigation of the development and structure of the human lung. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_A_histological_investigation_of_the_development_and_structure_of_the_human_lung
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G