Paper - A description of 34 human ova within the first 17 days of development
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Hertig AT. Rock J. and Adams EC. A description of 34 human ova within the first 17 days of development. (1956) Amer. J Anat., 98:435-493.
| Online Editor |
|---|

See also:
Hertig AT. and Rock J. On a normal human ovum not over 7.5 days of age. (1945) Anat. Rec. 91: 281. Hertig AT. and Rock J. On a normal ovum of approximately 9 to 10 days of age. (1945) Anat. Rec. 91: 281. Hertig AT. and Rock J. On a human blastula recovered from the uterine cavity 4 days after ovulation. (1946) J Gerontol. 1(1): 96-117.
|
| Historic Disclaimer - information about historic embryology pages |
|---|
|
A Description of 34 Human Ova within the First 17 Days of Development


Arthur T . Hertig, John Rock and Eleanor C. Adams
Departments of Pathology and Gynecology, Harvard Medical School, Boston, Massachusetts, The Pathology Laboratory of the Free Hospital
for Women, Brookline, Massachusetts and the Department of Embryology, Carnegie Institution of Washington, Baltimore
- This study has been aided over the years by grants from the Carnegie Corporation, the Milton Fund of Harvard University and the Carnegie Institution of Washington. During the past year it has also been supported by a grant, PathC-2451, of the National Institutes of Health of the United States Public Health Service.
Ninety Figures
Introduction
The purpose of this paper is to describe the early morphologic development of the normal human ovum and to compare it with corresponding phases in the development of the abnormal ovum. The observations recorded encompass a period from the second to the 17th days of development. The 34 specimens were found in the tubes or uteri from 211 patients who came to hysterectomy salpingectomy and unior bilateral oophorectomy for various therapeutic reasons.
The stimulus for this study was received in the years 1933-34 when the senior author had the good fortune to be a National Research Fellow under the inspiring guidance of the late Dr. George L. Streeter, then Director of the Department of Embryology of the Carnegie Institution of Washington at Baltimore. During this time many discussions were held with Drs. Streeter and Chester H. Heuser and Mr. O. O. Heard, all of whom emphasized the lack of material covering the first two weeks of human development. Opportunity to search for early human ova came when one of us (ATH) returned to Boston to become in succession pathologist at the Boston Lying-in Hospital (’34) and the Free Hospital for Women (’38). He found at the latter institution that the other of us (JR), a surgeon in search of ripe ova, was already dating in relation to the menstrual cycle laparotomies, considered advisable by others on the staff of the hospital. The senior author’s affiliation at the latter hospital made possible the closer association of the authors and the utilization of the material from that clinic. Thus began our happy association with the Department of Embryology of the Carnegie Institution of Washington in the joint enterprise of search, discovery, preparation and study of 34 early human ova. The first 4 were discovered and the first two (the 11- and 12-day stages) were prepared for publication during Dr. Streeter’s directorship which terminated May 1, 1940.
In the years since 1940, Dr. George W. Corner, to whom this volume is dedicated, ha.s enthusiastically encouraged the continuation of this joint study. He has been instrumental in securing financial assistance through his own budget to help defray the expenses of the Boston pole of the Boston Baltimore axis. The authors are ever mindful of Dr. Corner’s contributions to this joint study: his scholarly interest, his friendly encouragement, his expert embryologic interpretation of the specimens and his unique editorial help in the publication of these stages of human reproduction.
The authors are grateful to their other Baltimore colleagues without whose great and varied skills the study would have been impossible: Dr. Chester H. Heuser for his incomparable preparation and interpretation; Chester F. Reather and his successor, Richard D. Grill, for their superb photography; J amcs F. Didusch for his unique embryologic illustrations; Osborn 0. Heard for his inventive skills and Harriet Caspari for her varied technical assistance.
To our Boston colleagues the authors are grateful: Drs. F. O. Irving, F. A. Pemberton, G. Van S. Smith, past or present chiefs of staff; Lotte Lee Sichel, Susan Hedge and Jean Nauss, our successive clinic secretaries, and the resident and visting staff, notably Dr. W. J. Mulligan, who has aided in the clinical handling of these patients and who has operated on the later ones in the series.
Material and Methods
This paper presents the purely morphologic aspects of the 34 specimens. The senior author, because of his background in obstetric and gynecologic pathology with its necessary relationship to placental and endometrial pathology, has emphasized primarily the trophoblast and implantation site rather than the embryo in these early specimens.
A second paper is in preparation by Dr. John Rock and Mrs. Miriam Menkin. That paper deals entirely with the physiology and pathologic physiology of reproduction as learned from these 211 patients. Deductions will be drawn from the 34 patients whose uteri or tubes contained fertilized ova, as well as from the 177 in whose uteri no ova were found.
The heavy clinical load of the Free Hospital for Women has been admirably adapted to our purpose. Patients seen in the Out-Patent Department who require surgical relief for conditions not in1mediately harmful must await hospitalization for variable lengths of time (during the early years of the project frequently for several months). From women who are put on the list for operation by the examining surgeons in the Out-Patient Department we selected those to whom the following statements apply:
- They are incapacitated by symptoms caused by gross uterine displacement or minor uterine pathology, which can best be relieved by hysterectomy.
- Their menstrual habit is such as to indicate probable regular ovulation.
- They are living with their husbands and are willing and intelligent enough to co-operate.
The selected patients were asked to report each menstrual period during the months of waiting for admission and to record the dates of coitus during the cycle in which operation was to take place. In some cases urine for assay of hormones has also been collected prior to operation. (Rock and Hertig, ’42).
From 1938 to the end of 1954 the uteri and oviducts with their contents from this series of 211 patients have been examined by the following technique (Hertig and Rock, ’44a) provided menstruation had not occurred before the specimen was removed (even though menstruation had occurred such patients were kept in the series because they represented true failures) :
- “The uterus is opened laterally under saline or Locke’s solution and the uterine cavity is closely inspected with a four—power binocular dissecting microscope in an effort to recover a free blastocyst. The fluid is pipetted off and examined under the microscope. Also, in the event that the ovum may not yet have entered the uterus, the Fallopian tube corresponding to the ovary containing the most recent corpus luteum is routinely flushed, and the collected fluid is carefully examined in every case. The endometrium is then carefully examined under fluid prior to fixation by means of the low-power binocular dissecting microscope. If an implanted ovum is not found, Bouin’s fluid 3 is gently pipetted over the endometrial surface while the specimen is still immersed in salt solution and the endometrium is reexamined with low-power binocular vision.”
- Locke's solution - An aqueous solution of sodium chloride, calcium chloride, potassium chloride, sodium bicarbonate, and glucose adjusted to pH 7.4.
- Bouin's solution - Compound fixative solution of picric acid, acetic acid and formaldehyde.
Using this technique 34 early ova were recovered. Eight were free-lying, 1 in the tube and 7 in the uterine cavity. Twenty-six were implanted. Of the 34 specimens, 21 were normal and 13 were abnormal to one degree or another.
Once an implanted ovum was identified, the uterus was fixed in Bouin’s fluid for 36 hours. The endo- and myometrium surrounding the ovum was then blocked out into a cube approximately 15 mm in all diameters which was gradually dehydrated to 70% alcohol. The block of tissue containing the ovum was transported to Baltimore in 70% alcohol where it was photographed as an intact specimen and faultlessly sectioned by Doctor Heuser using the double embedding method of celloidin followed by paraffin. The serial sections were then stained with hcmatoxylin and eosin.
The free-lying ova stuck to the bottom of the preparation dish when Bouin’s fluid was added to the salt solution. They continued to stick to the glass throughout the gradual dehydration to 70% alcohol and this feature was utilized for the safe transportation of these tiny objects to Baltimore where they were photographed and later sectioned by Doctor Heuser.
Since July 1, 1953, cold acetone has been used to fix material for histochemical studies.
All but 6 of the 34 specimens have been described or illustrated in varying degrees of detail. All references to these prior publications are individually listed in tables 1 and 2 and will, therefore, not be referred to in the text. The essential clinical data from all 34 specimens together with their probable ages are summarized in these two tables.
The embryological measurements are in tables 3 and 4. Table 5 gives the location of the implantation sites within the uterus. Any discrepancies between these tabulated data and those previously published occur because this paper represents the first complete survey of the entire series of ova. The present data therefore are based on the interpretation of each specimen and its relation to all the others in the entire series.
Table 1
| Table 1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ovum Numbers | Streeter Horizon | Estimated Age of Ovum (days) | Probable Day of Cycle on which ovulation occurred | Probable Day of cycle on which fertile coitus occurred | Range of EOT | Range in onset of Catamenia | Day of Cycle on which Operation occurred | Histologic "Dating" or Pattern of Endometrium | Age of Patient | No. of Labor | No. Abortions | Previous Publication References |
| 8698 | II Segmenting egg 2 cells | 1.5-2.5 | 12-13 | 12* | 9-15 | 25-27 | 13 | 17 | 42 | |||
| 8904 | II 12 ± cells | 3 | 16 | 15 or 17 | 7-18 | 23-30 | 20 | 18 | 35 | |||
| 8794 | III Free blastocyst 58 cells | 4 | 13 | 12 | 8-14 | 24-26 | 17 | 19 | 40 | |||
| 8663 | III 107 cells | 4.5 | 22 | 22 | 18-25 | 34-37 | 27 | 22 | 38 | |||
| 8225 | Va Ovum implanted but avillous. Solid trophoblast | 7 | 14 | 14* | 8-18 | 24-30 | 22 | 21 | 30 | |||
| 8020 | Va | 7 | 17 | 16* | 12-20 | 28-32 | 24 | Earliest progestational change | 38 | |||
| 8155 | Va | 8 | 13 | 12 or 13 | 12-16 | 28 | 22 | Earliest progestational hyperplasia | 29 | |||
| 8215 | Vb Beginning lacunae | 9 | 46 | 45 or 46 | 11-43 | 27-55 | 56 | Earliest progestational hyperplasia | 30 | |||
| 8171 | Vb | 9 | 13 | 12 | 11-18 | 27-30 | 23 | Earliest progestational hyperplasia | 38 | |||
| 8004 | Vb | 9 | 15 | 15 | 15-22 | 31-34 | 25 | Moderate progestational hyperplasia | 29 | |||
| 7699 | Vc Lacunae well-formed, extra-coelomic cavity present | 11 | 14 | 12 | 10-17 | 26-29 | 25 | Moderate progestational hyperplasia | 34 | |||
| 7950 | Vc | 12 | 14 | 13* | 11-21 | 27-33 | 26 | Moderate progestational hyperplasia | 31 | |||
| 7700 | Vc | 12 | 17 | 15* | 17-22 | 33-34 | 29 | Advanced progestational hyperplasia | 34 | |||
| 8558 | Vc | 12 | 13 | 12 or 13 | 11-23 | 27-33 | 25 | Advanced progestational hyperplasia | 38 | |||
| 8330 | Vc | 12 | 11 | 10 | 7-14 | 23-26 | 23 | Advanced progestational hyperplasia | 31 | |||
| Carnegie Embryo | Vc | 12 | 16 | 15 | 13-17 | 29 | 29 | Advanced progestational hyperplasia | 32 | |||
| 8672 | VI Primitive villi, distinct yolk-sac | 13 | 12 | 12* | 11-23 | 27-35 | 25 | Advanced progestational hyperplasia | 33 | |||
| 8360 | VI | 13 | 15 | 14 | 10-18 | 26-30 | 28 | Early decidua | 33 | |||
| 7801 | VI | 13 | 14 | 14 | 8-16 | 24-28 | 28 | Early decidua | 33 | |||
| 7802 | VII Branching villi, axis of germ disc defined | 16 | 17 | 16 | 10-21 | 26-33 | 33 | Early decidua | 37 | |||
| 8602 | VII | 17 | 11 | 11 | 13-17 | 29 | 28 | Early decidua | 34 | |||
| ||||||||||||
|
Reference: Hertig AT. Rock J. and Adams EC. A description of 34 human ova within the first 17 days of development. (1956) Amer. J Anat., 98:435-493. | ||||||||||||
Description of Horizons
(Streeter, ’42)
Horizon I
One-cell fertilized egg. None was found in this series although an ovum with pronuclei was reported by Hamilton in 1949.
Horizon II
Segmenting ovum. Two were found; the twocell tubal specimen (8698) and the 12—cell intra-uterine morula. The latter (8904) unfortunately was lost before it could be photographed or sectioned. However under the dissecting binocular it was a spherical morula surrounded by a clear zona. Twelve blastomeres were counted, including one large one, centrally situated and presumably embryonic in potential, and 11 smaller peripheral ones. The two-cell stage is illustrated in figures 1 and 2. Its estimated age is 1.5—2% days; that of the 12—cell ovum is 3 days.
Horizon III
Free blastocyst. The two ova found in this stage are described and illustrated in figures 3—6. The segmentation cavity is just forming in the younger 58-cell one (8794) but is Well formed in the older 107-cell ovum (8663). In this stage the primitive trophoblastic cells are multiplying at a much more rapid rate than the larger formative or embryonic cells of the inner cell mass. A zona is present in the younger of the two specimens but has disappeared in the older specimen. The estimated age of the 58-cell stage is 4 days; that of the 107—cell stage is 4% days.
Horizon IV
Implanting ovum. None was found in the series. This stage would be comparable to the 6 implanting macaque blastocysts described by Heuser and Streeter (’41). Heuser, Rock and Hertig (’45) and Hertig and Rock (’45c) interpreted a 7-day (8020) and a 9-day specimen (8004) with solid and lacunae-riddled trophoblast respectively as belonging to Streeter Horizon IV, i.e., implanting ova. Actually these two specimens (included now in Horizon Va and b respectively) represent two significantly different stages of development. Therefore, we now believe that Horizon IV should be reserved for the attaching blastocyst whose age would probably be midway between our 4%; and 7-day stages (8663 and 8020 respectively).
Horizon V
Ovum implanted but still avillous. Eleven normal ova were found in this developmental stage. We propose to subdivide Horizon V into three phases as follows.
Va
Solid trophoblast: three ova (figs. 9-14). At 7 and 8 days of age the ovum has become shallowly implanted on solid endometrium approximately equidistant from several gland mouths. (Implantation probably occurs at about 5% to 6 days.) Presumably due to augmented corpus luteum function, the endometrium begins to show evidence of progestational hyperplasia in the form of dilated but non-congested capillary sinusoids. (See discussion of Endometrium, page 455.) The ovum is discoid in shape with solid mixed syncytio- and cytotrophoblast at its basal and lateral margins (embryonic or implantation pole) but with the unaltered th-in-walled primitive trophoblast of the blastocyst wall at its superior margin (abembryonic pole). This portion of the ovum is thus exposed to the uterine cavity. The blastocyst wall has formed definitive trophoblast only in the area of endometrial contact, presumably as the result of an induction factor of endometrial origin. Thus the remainder of the blastocyst wall is relatively unchanged and is comparable to the trophoblastic blastomeres of the 4%—day specimen (8663). An amniotic cavity and/or amnion is associated with a simple bilaminar germ disc but no exocoelomic (Heuser’s) membrane has formed. The presence of the germ disc is apparently responsible for the induction of amniogenesis from the adjacent trophoblast. The formerly distended blastocyst cavity has collapsed in the process of implantation to become a flattened chorionic cavity.
Vb
Formation of trophoblastic lacunae with amniotic and exocoelomic cavities: three ova (figs. 15—23). At 9 days of age lacunar spaces are forming in the trophoblast which is becoming more organized into definitive cytotrophoblast, usually centrally located, and syncytiotrophoblast, usually peripherally located. It is not well understood whether each type of trophoblast forms independently from the trophoblastic blastomeres of the blastocyst wall or whether the syncytium forms from the primitive cytotrophoblast. At any rate mitoses are found in the latter but not in the former which possibly may divide by amitosis. At the abembryonic pole the trophoblast is thicker than in the 7- and 8-day specimens because of contact with endometrium as the ovum embeds more deeply and as healing of the endometrial penetration defect begins.
Although the vast majority of the lacunar spaces have coalesced they contain relatively little maternal blood, and that mostly plasma, since few adjacent capillary sinusoids of the endometrial stroma communicate directly with the lacunar network as yet. The foundation, however, is being laid for a utero-placental circulation of relatively large volume although at present its capacity is, for the most part, unused.
The amnion is continuing to be derived from the cytotrophoblast adjacent to the embryo. The amniogenic cells show variable degrees of differentiation and cytotrophoblastic attachment. The amniotic cavity is well formed, either cleftor lens-shaped and is bounded below by the concave germ disc and above by the level or arched amnion, the latter continuous with the periphery of the germ disc.
The simple bilaminar germ disc, without embryonic axis, is composed of tall columnar vacuolated ectoderm and flattened or polyhedral endoderm with relatively large nuclei. The mesoblast of the exocoelomic (Heuser’s) membrane is continuous with the endoderm and derived from but focally still attached to the cytotrophoblast lining the chorionic cavity. The membrane shows variable degrees of development which is not absolutely correlated with the size and development of the specimen.
Vc
Intercommunicating lacunae with beginning merolacunar circulation: five ova (figs. 24-38). At 11 to 12 days of age, the ovum has become still more deeply embedded in the maternal stroma, and, following progressive healing of the penetration defect by the endometrium, the trophoblast has proliferated over most of the surface of the ovum. The lacunar spaces now intercommunicate and contain enough maternal blood to make ova in this stage easily identified on careful gross examination of the endometrial surface prior to fixation. Such lacunar blood appears as a discontinuous red circle about 1mm in diameter. Hemorrhage and congestion about the implanted ovum are also prominent grossly and, in some cases, there is an associated area of similar hemorrhage and congestion on the opposing endometrial surface where there had been contact with the implantation site prior to the opening of the uterus. This suggests the elaboration by the trophoblast of a substance capable of dilating blood vessels and allowing the escape of erythrocytes into the surrounding tissues.
The future utero-placental circulation is developing from the confluence of maternal sinusoids with the lacunar space. These capillary sinusoids, originating from anastomoses between spiral arterioles and endometrial veins, dilate and undergo actual growth adjacent to the proliferating trophoblast. This growth is evidenced by frequent endothelial mitoses. The sinusoids are in the process of connecting with the lacunar spaces by new secondary capillaries or venules. At this developmental stage, some connections are complete and other capillaries are still separated from the lacunar space by their intact endothelium. These intact and hence untapped vessels often are surrounded for a considerable distance by syncytiotrophoblast.
Organization of the trophoblast has progressed so that the chorionic cavity is surrounded by the small uniform cells of the cytotrophoblast. As the ovum ages, small masses of these cells covered by a thin layer of syncytium project peripherally into the rapidly expanding area of lacunae and syncytiotrophoblast. These primordial villi set the pattern for the constant relationship of the two types of trophoblasts; the inner cyto— and outer syncytiotrophoblast. The syncytium thus will always form the lining of the intervillous lacunar space or, conversely, the covering of the villi. In many of these specimens the syncytiotrophoblast appears to have a brush border. Mitoses frequently are seen in the cytotrophoblast but not in the syncytiotrophoblast whose nuclei are irregular in shape and size and are frequently clumped. There is occasional evidence of ingestion by this tissue of maternal stromal cells and/or leucocytes.
The chorionic cavity has become spherically distended causing the ovum to protrude above the surrounding endometrial surface. Mesoblasts are delaminating from the cytotrophoblasts lining the chorionic cavity to form a fine loose meshwork lying between it and the exocoelomic membrane. The latter is formed by a single layer of mature flattened loosely joined mesoblasts and serves to delineate the exocoelom from the rest of the chorionic cavity. Angioblasts also delaminate from the cytotrophoblast and are seen as vacuolated cells and strands intermingling with the mesoblasts at the periphery of the chorionic cavity. This exocoelomic cavity is but transitory and reaches its maximum size during the latter part of this phase. The fluid in the cavity appears to be of somewhat different composition from that outside, where there is a greater amount of finely granular, eosinophilic precipitate surrounding the mesoblasts.
The bilaminar germ disc continues to grow at a relatively slow rate with respect to the rest of the ovum. It flattens somewhat as it ages, perhaps due to the expansion of the amniotic cavity. Amnioblasts are still delaminating and arising in situ from the overlying cytotrophoblast to enclose the dome—shaped amniotic cavity.
A 6th specimen in this general stage of development was fixed in acetone for histochemical study and will be described and illustrated in a separate publication. Drawings from reconstructions of selected stages of previllous ova are shown in figures 54-58.
Horizon VI
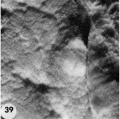
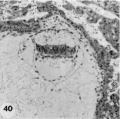
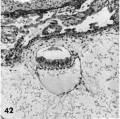
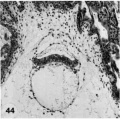
Primitive villi, distinct yolk sac: three ova (Plate 7 and Plate 8 - figs. 39-47).
At 13 days of age the ovum is evident upon gross examination by virtue of its size and elevation. It is not completely embedded within the endometrium. Bleeding, which may occur through the unhealed implantation site, arises from the increased flow of maternal blood into the lacunar spaces resulting in rupture of the thin-walled abembryonie pole of the trophoblast. There is also bleeding into an occasional endometrial gland whose Walls have been eroded by invading trophoblast thereby allowing the maternal blood iii the lacunar spaces to flow into the gland lumen. Since this bleeding occurs at about the time of the first missed menstrual period it is an occasional source of clinical inaccuracy in foretelling the date of expected delivery.
In the trophoblastic shell of the ovum, the cytotrophoblastic cells are rapidly proliferating so that their total mass now exceeds that of the syncytium. The latter now lines the intervillous space which contains occasional streamers of projecting syncytiotrophoblast. Some syncytium detaches as it invades the endometrial stroma and is seen as giant cells within the decidua. Primitive villi are now forming. They have a shallow core of mesoblasts and angioblasts arising from and projecting into otherwise solid cytotrophoblast which in turn is covered by syncytium. The peripheral tips of the villi are coalescing to form the cytotrophoblastic placental. floor which is perforated by capillary sinusoids supplying the intervillous space with maternal blood.
The mesoblast which lines the chorion is more abundant than in earlier stages particularly in the space between the amnion and the chorion. Angioblastic tissue, also delaminating from cytotrophoblast and lying in this mesoblast, is in various stages of early differentiation.
The definitive yolk-sac is present in all three of our specimens in this developmental stage. The mechanism of its formation is not well understood. The yolk-sac as it appears in the youngest specimen of this group is considerably smaller than the exocoelomic cavity found in our ova one day younger. Apparently the abembryonic portion of the exocoelomic membrane ruptures in several places to form small vesicles or strands of mesoblast now seen lying within the chorionic cavity. The free ends of the exocoelomic membrane which are still continuous with the endodermal plate join to enclose the smaller definitive yolk-sac cavity whose mesoblastic membrane soon becomes double-layered. The extraembryonic mesoblast of the inner layer becomes transformed into extraembryonic “yolk-sac endoderm” continuous with but not derived from the embryonic endoderm of the germ disc.
- This is essentially Doctor Streeter’s concept of the formation of the yolk sac. It is a larger part of his thesis that all auxiliary structures are derived from the trophoblast and its descendants and not from the inner cell mass and its resulting germ disc.
One of our specimens is in the single-layered stage, the other two have a double-layered yolk-sac membrane. The domeshaped amniotic cavity has increased in size and its membrane in some places has become double-layered also.
In our oldest embryo of this age group the germ disc shows evidence of an axial differentiation and beginning embryonic mesoderm. The embryo of this specimen, 7801, has been described in detail (Heuser, Rock and Hertig, ’45). The embryos of the two younger specimens await detailed study.
Horizon VII
Branching villi, axis of the germ disc defiiiczl: two ova (figs. 48-53). By 16 days of age the grossly visible ovum has branched villi around its entire surface. Their mesoblastic cores contain vascular primordia derived from (1) cells arising by delamination from the cytotrophoblast lining the chorionic cavity and then migrating into the mesoblastic core of the villus, and (2) angioblasts differentiating from the cytotrophoblastic cells of the trophoblastic columns during the formation of primitive villi (Hertig, ’35). [The isolated vascular primordia thus formed show independent power of growth and, later (by the 21st to 23rd day), the fetal circulation in the villi will begin to function as these isolated primordia coalesce.] The flow of maternal blood through the intervillous space is increased as additional newly-formed capillaries and Venules link this space with many large blood-filled sinusoids in the endometrium surrounding the ovum.
Once the architecture and active maternal blood supply of the placenta have been established, the germ disc, whose rate of growth has been relatively slow, rapidly begins to differentiate. Our 16-day specimen not only has an axial differentiation but in addition a definite primitive streak, the beginnings of a head process and a cloacal membrane.
Within the chorionic cavity there is the double-layered yolksac consisting of a thin inner extraembryonic “endodermal” layer separated by a variable distance from a wavy festooned outer mesoblastic layer. Early angiogenesis is evident in cells clumped together between these layers. The allantois is also forming in our 16-day specimen of this group (7802).
The amniotic cavity has expanded and has an amniotic duct which extends dorsally into the condensed mesoblast lining the chorionic cavity.
Our specimen, 7802, has been described as a late example of Horizon VII and 8602 appears to be somewhat older. The embryo of the latter awaits more detailed study.
These two ova are older than those found at about the time of the first missed menstrual period. The patient producing the 16-day specimen had cycles of variable length and in this instance ovulated early in the cycle. The patient producing the 17-day specimen, said to have regular 28-day cycles, ovulated earlier than was to be expected. (See table 1.)
The measurements of the 20 normal specimens just described are given in table 3.
Abnormal Ova
These 13 specimens encompass a range of development from early segmentation to the formation of early villi. (See table 4 for measurements.) Although the 4 segmenting ova have from 5 to 12 blastomeres and were found in the uterine cavity they may have started to degenerate in the tube. They are characterized and illustrated in figures 59-66. Their chief interest lies in the fact that they Were recovered from patients of normal fertility (19 full term labors and only three abortions from the group of 4 patients, table 2). Moreover their fertility is essentially the same as the group of 4 patients (20 full term labors and three abortions) from whom were recovered the 4 unquestionably normal segmenting ova illustrated in figures 1-6. Thus 50% of the free-lying ova were abnormal in a series of patients of high fertility.
The 9 implanted ova show variable types and degrees of abnormality including an ovum with syncytiotrophoblast only (figs. 67, 68); an ovum with poor trophoblast and no embryo (figs. 69, 70); an ovum with relatively good trophoblast but no chorionic cavity (figs. 71, 72, 73); three ova with variable degrees of trophoblastic hypoplasia (figs. 74—80); two ova showing shallow implantation, one of which also had a maloriented germ disc; and a final specimen whose only abnormality was a maloriented germ disc (figs. 81-90). The significance of these implanted but abnormal specimens, also occurring in patients of high fertility (see table 2), lies in their probable relationship to the pathogenesis of spontaneous abortion. Although 34.6% of the implanted ova in this series (9 out of 26) are abnormal, one cannot be sure that they all would have aborted. By the same token the more abnormal ones might not have caused even clinical evidence of pregnancy perhaps they would have been merely swept away with the unsuppressed menstrual flow. It is impossible to say how many of these abnormal pregnancies would have terminated in spontaneous abortion. It is significant that about 12% of clinically recognized pregnancies do terminate in spontaneous abortion.
Table 2

Table 3

Table 4
| Carnegie Number | Horizon | Age (days) | Overall Size | Chorionic Cavity | Trophoblast | Embryonic Disc | Amniotic Cavity | Exocoelomic Cavity | Yolk-Sac Cavity | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 8630 | II | 4 | 0.095 x 0.070 | |||||||||
| 8450 | II | 4 | 0.088 x 0.056 | |||||||||
| 8190 | II | 4 | 0.116 x 0.073 | |||||||||
| 8452 | II | 5 | 0.090 x 0.072 | |||||||||
| 8370 | Vc | 9 | 0.320 x 0.440 | Absent | 0.03 to 0.24 | 0.06 x 0.04 | 0.04 x 0.026 | Absent | ||||
| 7770 | Vc | 12 | 0.580 x 0.510 | 0.245 x 0.28 | 0.04 to 0.180 | 0.11 x 0.06 | 0.08 x 0.02 | 0.16 x 0.22 | ||||
| 8299 | Vc | 12 | 0.610 x 0.860 | 0.32 x 0.45 | 0.07 to 0.285 | 0.12 x 0.07 | 0.10 x 0.02 | 0.08 x 0.21 | ||||
| 8329 | Vc | 11 | 0.400 x 0.640 | Absent | 0.400 to 0.640 | Absent | Absent | Absent | ||||
| 8000 | Vc | 12 | 0.740 x 0.640 | 0.43 x 0.32 | 0.005 to 0.42 | 0.11 x 0.05 | 0.10 x 0.04 | 0.28 x 0.25 | ||||
| 7771 | Vc | 12 | 0.77 x 0.49 | 0.08 x 0.06 | 0.21 to 0.38 | Absent | Absent | Questionably present | ||||
| 7850 | VI | 13 | 0.66 x 0.860 | 0.46 x 0.43 | 0.07 to 0.19 | 0.12 x 0.06 | 0.08 x 0.025 | Involuted | 0.16 x 0.20 | |||
| 8290 | VI | 13 | 1.33 x 1.2 | 1.2 x 0.75 | 0.06 to 0.38 | 0.20 x 0.10 | 0.15 x 0.07 | Involuted | 0.07 x 0.15 | |||
| 7800 | VII | 14 | 1.08 x 1.4 | 0.8 x 1.14 | 0.1 to 0.48 | 0.20 x 0.06 | 0.14 x 0.14 | Involuted | 0.40 x 0.25 | |||
|
Tables as Images: table 1 | table 2 | table 3 | table 4 | 1956 Hertig Rock Reference: Hertig AT. Rock J. and Adams EC. A description of 34 human ova within the first 17 days of development. (1956) Amer. J Anat., 98:435-493. | ||||||||||||
Location of Implantation Site
There was no correlation between the position of the implanted ovum and the side on which the most recent corpus luteum was found. On the other hand, in our limited series of 26 implanted ova there appears to be a correlation between the wall on which the ovum is implanted and its normality: the normal ova were twice as frequently recovered on the posterior wall, while the abnormal ones were twice as frequently found on the anterior wall. Regardless of whether normal or abnormal, all the ova were implanted in the upper half of the uterine cavity. These data are summarized in table 5.
Table 5
| Table 5 | ||||||
|---|---|---|---|---|---|---|
| Ova | No. | |||||
| Anterior | Posterior | Same | Opposite | Center | ||
| Normal | 17 | 5 | 12 | 10 | 6 | 1 |
| Abnormal | 9 | 6 | 3 | 5 | 4 | 0 |
|
Reference: Hertig AT. Rock J. and Adams EC. A description of 34 human ova within the first 17 days of development. (1956) Amer. J Anat., 98:435-493. | ||||||
The Endometrium
Within a day or two after implantation of the fertilized ovum, the endometrium throughout the uterus undergoes a series of changes designated by us as progestational hyperplasia. The initiation of this process is seen in one of our 7-day specimens. This histologic sequence of events undoubtedly reflects the effect of continually increasing stimulation of the endometrium by the corpus luteum which in itself begins to show morphologic evidence of heightened activity (White, Hertig, Rock and Adams, ’51). This sequence begins as an increased blood flow manifested as a slight dilatation and congestion of the capillary sinusoidal network of the compact zone. It is soon followed by increasingly evident dilatation and congestion of the spiral arterioles. Such increased blood flow is presumably responsible for the marked stromal edema which not only persists but increases (beyond the peak reached at about 22 days in the non-pregnant cycle) only to decline when early decidua has been formed at the time of the first missed menstrual period. Glandular secretion, which had reached its peak at about the 21st day (7th postovulatory) of the menstrual cycle, is reactivated in endometria associated with the 9-day ovum and increases more or less progressively to the stage of early decidual formation. The predecidual reaction in the stroma about the spiral arterioles and beneath the surface epithelium, ordinarily beginning during the 23rd day (9th postovulatory) of the normal non-pregnant cycle, develops at its usual gradual pace but with fewer infiltrating leucocytes " until the time of the missed period when it morphologically resembles early decidua.
Therefore, the first diagnostic morphologic evidence of pregnancy, aside from the ovum itself, is vascular prominence with edema as seen in endometrium associated with the 8-day specimen. From that time on variable degrees of persisting edema, increased vascularity and renewed glandular secretion are seen up to the stage when obvious early decidua is present — about the time of the first missed menstrual period.
The endometria of the 9 implanted but abnormal ova are not only morphologically normal but show progestational hyperplasia of a degree consistent with the interval between the probable day of ovulation and the day of operation. Although there is a wide spectrum of abnormality in the ova themselves, the endometria give no morphologic evidence of, or reason for, such abnormalities. The corpora lutea, however, show a variety of morphologic changes which deviate from those associated with normal ova 8 or more days of age. Such morphologic deviations, all indicating lessened activity, include poor theca interna, lessened vascularization, relative ischemia, reduction of K cells and reduced peripheral vacuolization of granulosa lutein cells (White et al., ’51). Judging from the progestational hyperplasia seen in the endometria, the initial quality of these corpora lutea was presumably within normal limits.
Although as preclecidua develops into decidua the infiltrating polymorphonuclearand mononuclear leucocytes are somewhat reduced in numbers, these leucocytes may be present in variable, and often, considerable numbers about the ovum without their presence being of pathological significance.
Therefore, the failure of the corpora lutea to undergo secondary hyperplasia, as in a normal pregnancy, must necessarily be due to lack of adequate direct or indirect stimulation by the variously defective trophoblast.
Summary
During the 16-year period from July 1938 to July 1954, surgically removed uteri and tubes from 211 patients of known high fertility were searched for the presence of early ova of a developmental age up to 14 days. Thirty-four specimens were found ranging from a two-day, two-cell tubal egg to a 17-day well implanted ovum with branching chorionic villi and a Well defined embryonic axis. Twenty-one specimens were normal and 13 were abnormal in one or more ways.
The essential clinical and embryological data on each specimen have been tabulated. The gross and microscopic features of each ovum have been illustrated.
The progestational changes in the endometrium incident to pregnancy are of corpus luteal origin and are directly, or via the hypophysis, indirectly induced by the presence of the ovum. Such changes are first apparent in the 8-day specimen and increase progressively up to the formation of early decidua at the time of the missed menstrual period.
The endometria associated with the implanted but abnormal ova are normal, although the poor secretory quality of the corpus luteum, due presumably to inadequate trophoblastic stimulation, would suggest the probability of an inevitable abortion of most of those specimens.
Literature Cited
Hamilton WJ. Early stages of human development. (1949) Ann R Coll Surg Engl. 4(5): 281-94. PMID 18121228 PMC2238331
Hertig AT. Angiogenesis in the early human chorion and in the primary placenta of the macaque monkey. (1935) Carnegie Instn. Wash. Publ. 459, Contrib. Embryol. 25: 37-81.
Hertig AT. On the development of the amnion and exocoelomic membrane in the previllous human ovum. (1945) Yale J Biol Med. 18:107-15. PubMed 21007544
Hertig AT. and Rock J. On a complete normal 12-day human ovum of the pre-villous stage. (1939) (Abst.) Anat. Rec. 73: 26.
1941 Two human ova of the pre-villous stage, having an ovulation age of about 11 and 12 days respectively. Contrib. to Embryol., 2.9: 127-156.
1942a On the eleven day pre-villous human ovum with special reference to the variations in its implantation site. (Abst.) Anat. Rec., 82: 420.
1942b On a series of 9 human fertilized ova (5 normal and 4 pathological) obtained in surgically removed uteri prior to the first missed menstrual period. (Abst.) Fed. Proc. Part II p. 178.
1943a. On the seven and one-half day human ovum (Carnegie 8020). (Abst.) Anat. Rec., 85: 317.
1943b On the nine and one-half day human ovum (Carnegie 8004). Ibid., 8.5: 317-318.
1944a On the development of the early human ovum with special reference to the trophoblast of the previllous stage: a description of 7 normal and 5 pathologic human ova. Am. J. Obst. and Gynec., 47: 149-184.
1944b On a normal human ovum of approximately 795 to 8 days of age (Carnegie 8155). (Ahstr.) Anat. Rec., 88: 437-438.
1944c On a normalhuman ovum of approximately 8 days of age (Carnegie 8171). (Abst.) Ibid., 88: 438.
1945a On a normal human ovum not over 7-}: days of age. (Abst.) Ibid., 91: 281.
1945b On a normal ovum of approximately 9 to 10 days of age. (Abst.) Ibid., 91: 281.
1945c Two human ova of the pre-villous stage, having a developmental age of about seven and nine days, respectively. Contrib. to Embryol., 31: 67-84.
1946 On a human blastula recoverd from the uterine cavity 4 days after ovulation. (Abst.) Anat. Rec., 94: 469.
Hertig AT. and Rock J. Two human ova of the pre-villous stage, having a developmental age of about eight and nine days respectively. (1949) Contrib. Embryol., Carnegie Inst. Wash. Publ. 583, 33: 169-186.
Hertig AT. and Rock J. A series of potentially abortive ova recovered from fertile women prior to the first missed menstrual period. (1949) Am. J. Obst. and Gynec. 58: 986-993.
1950 Abortive human ova and associated endometria. Menstruation and its Disorders. Charles C Thomas, Springfield, Ill., pp. 96-137.
1951 The implantation and early development of the human ovum. Am. J. Obst. and Gynec., 61A: 8-14.
HERTIG, A. T., J. ROCK, W. J. MULLIGAN, 1*‘. A. PEMBERTON AND E. C. ADAMS 1952 On three segmenting human ova, one recovered from the tube and two from the uterine cavity (Carnegie 8698, 8794 and 8663). (Abst.) Anat. Rec., 112: 342-343.
Hertig AT. Rock J. Adams EC. and Mulligan W.J. On the preimplantation stages of the human ovum: a description of four normal and four abnormal specimens ranging from the second to the fifth day of development. (1954) Carnegie Instn. Wash. Publ. 603, Contrib. Embryol., 35: 199-220.
Heuser CH. and Streeter GL. Development of the macaque embryo. (1941) Contrib. Embryol., Carnegie Inst. Wash. Publ. 525, 29: 15-55.
HEUSER, C. H., J. ROCK AND A. T. HERTIG 1945 Two human embryos showing early stages of the definitive yolk sac. Ibid., 3]: 85-99.
Rock, J., AND A. T. HERTIG, 1942 Some aspects of early human development. Am. J. Obst. and (}ynec., 4-4: 973-983.
1944 Information regarding the time of ovulation derived from 1 stud tof three unfertilized and 11 fertilized ova. Ibid., 4'7: 343-356.
1948 The human conceptus «luring the first two weeks of gestation. Ibid., 5.}: (5-14.
Streeter GL. Developmental horizons in human embryos. Description of age group XI, 13 to 20 somites, and age group XII, 21 to 29 somites. (1942) Contrib. Embryol., Carnegie Inst. Wash. Publ. 541, 30: 211-245.
Wn1'rE, R., A. T.HERTIG, J. ROCK: AND E. C. ADAMS 1951 Histologic mnl. histoehemical observations on the corpus luteum of human pregnancy with special reference to corpora lutea associated with early normal and abnormal ova. Contrib. to Embryo1., 34': 55-74.
Plates
Plate 1
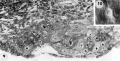
Normal preimplantation stages in human development; one segmenting egg found in the tube, Streeter Horizon II, and two free intrauterine blastoeysts, Streeter Horizon Ill.

1 Intact two-cell segmenting egg. Note smaller upper polar body between the two blastomeres. The lower large one, however, is not in focus. The noun is defective, possibly caused during fixation and partial dehydration or during removal of the ovum from the dish to which it was adherent. See figure 2 for section. Carnegie 8698, Sequence 12. X 500.
2 Mid-serial section of segmenting tubal egg shown in figure 1. Note nuclear detail, difference in size and shape of blastomeres and loss of zona. Carnegie 8698, Section 4. X 500.
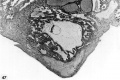
3 An intact 58-cell intrauterine blastocyst. Note eccentrically situated segmentation cavity, the faintly seen inner cell mass and the peripherally arranged blastomeres. The zona and one polar body (lower right) are faintly seen. Carnegie 8794, Sequence 4. X 500.
4 Mid-serial section of 58-cell intrauterine blastocyst shown as intact specimen in figure 3. Note absence of zona, about two-thirds of the periphery, probably of artifactitious origin. The two polar bodies are evident at opposite poles of the ovum. The inner cell mass of 5 blastomeres, only 4 of which are present in this section, is lying eccentrically within the segmentation cavity and surrounded by primitive troplioblastic blastomcres. The primitive segmentation cavity, composed of several coalescing spaces, is most clearly delineated as a single unit in this and the two contiguous sections. Note that the nuclei of the embryonic blastomeres, although of equal size, are less vesicular and contain denser clumps of chromatin than those of the trophoblastic blastomeres. Carnegie 8794, Section 7. X 500.
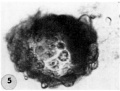
5 An intact 107-cell intrauterine blastocyst. Note segmentation cavity, cccontrically located inner cell mass at right and pebbly-appearing well formed by trophoblastic blastomerea. There is no zona. Carnegie 8663, Sequence ‘I. X 500.
6 Mid-serial section of 107-cell blastocyst shown as intact. specimen in figure 5. There are 8 definite large vacuolated formative (embryonic) cells, 69 mural and 30 polar trophoblastic cells. Possibly -1 of the latter are actually formative endodermal cells. Note parts of 4 formative (embryonic) cells at right of picture. The. elongated cell to the upper left of the embryonic disc, interpreted originally as a polar trophoblast, may actually be one of the 4 potential endodermal cells. Note marked irregularity in form and some variation in size of mural trophoblastic cells. The polar trophoblastic cells encircle the. inner cell mass as a corona and vary in shape but tend to be polyhedral. Carnegie 8663, Section 9. X 500.
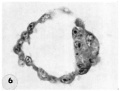
7 Endometritun, associated with Carnegie No. 8663 (figs. 5, 6), showing the typical morphologic features of the 19th day of a classic 28-day menstrual cycle. Presuma.bly ovulation occurred approximately 108 hours before the surgical removal of the uterus. Note tortuous actively secreting glands, with regularly arranged basal vacuoles and moderately edematous stroma without predecidual reaction. l*‘IlW, S-48-5000. X 91.5.
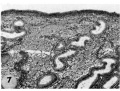
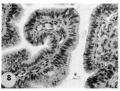
8 Mucosa of fallopian tube, associated with Carnegie 8663 (figs. 5. 6). showing the normal tall columnar ciliated epithelium of the mid-interval phase.
- Streeter Horizon ll
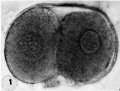
1 Two-cell segmenting egg
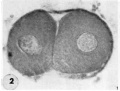
2 Mid-serial section two-cell


- Streeter Horizon Ill
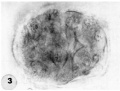
3 Intact 58-cell blastocyst
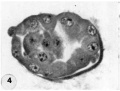
4 Mid-serial section 58-cell blastocyst
5 An intact 107-cell blastocyst
6 Mid-serial section 107-cell blastocyst
7 Endometrium
8 Mucosa fallopian tube






Plate 2
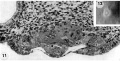
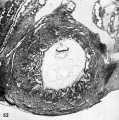
Three normal, previllous ova, Streeter Horizon Va, possessing solid syncytioand cytotrophoblast without. lacunar development, an amniotic cavity and/or an early amnion, a simple hilaminar germ disc and early mcsoblast formation.

9 A recently implanted ovum of not more than 7.5 days of age, based upon a critical coital date. Note solid trophoblast. composed of paler discrete mononuelear cells and darker less discrete multinucleated cells. The solid disc of trophoblast at the embryonic (implantation) pole is continuous with the membranous unaltered blastocyst wall at the abembryonic pole. Note also the amniogenic cells, tiny cleft-like amniotic cavity, large columnar, non-vacuolated ectodermal cells and smaller tlattcned or polyhedral endodcrmal cells. The ehorionic cavity is irregularly flattened due to collapse of the hlastocyst. Carnegie 8225, Section 20-5-5. X 300.
10 Surface view of the normal 7-day ovum seen in figure 9 photographed under fluid after fixation and partial dehydration. Note embryonic mass within chorionic cavity and the relatively large proportion of the ovum exposed to the uterine lumen. Note prominent gland mouths and surface wrinkling consistent with the 22nd day of the menstrual cycle. Carnegie 8225, Sequence 5. X 22.
11 Another recently implanted ovum, also not more than 7; days of age in view of critical dating of coitus, hut interpreted as being slightly older than 8225 hecause of germ-disc. development. and greater trophoblastic maturity. Note greatest concentration of dark syncytiotrophoblast in the center of the implantation pole dorsal to the embryo. The lighter cytotrophoblast is most prominent peripherally but also lines the inner surface of the proliferating trophoblast. It is amniogenic at its area of contact with the embryo. The nuclei of the surface endomctrial epithelium at its contact with the trophoblast show the abnormality of hydropic degeneration. These might be mistaken for nuclei of syneytiotrophoblastic origin. Actually some of the smaller denser nuclei of the syncytiotrophoblast appear to he of cndometrial stromal origin through the process of ingestion. Carnegie 8020, Section 6-5-9. X 300.
12 Surface view of the same 7-day ovum seen in figure 1]. Note clearly visible embryonic mass within dark chorionic cavity which is surrounded by an opaque zone of trophoblast folded at the equatorial plane of the ovum. Carnegie 8020, Sequence 6. X 22.
13 An ovum, probably about 8 days of age. Although coital data for this specimen are not critical, its development places it between the 7- and 9-day stages on which critical coital data are available. This inconspicuous stage of implantation is characterized by nearly complete nidation, solid but festooned trophoblast, a small ehorionic. cavity containing a few mesoblasts and a dorsally concave germ disc. Although the primordium of the amniotic cavity is prominent there appears, as yet, to be no amniogencsis. Note actively invading prongs of syncytiotrophoblast surrounding an area of endomctrial stroma and containing two distinct types of nuclei. The large ones divide amitotically but at least some of the smaller ones appear to be of cndometrial stromal origin suggested by the indistinct zone of demarcation between the trophoblast and the maternal stroma. Carnegie 8155, Section 4-4-8. X 300.
14 Surface view of intact ovum shown in figure 13. Note the inconspicuous nature of the slightly raised but opaque implantation site. Carnegie 8155, Sequence 8. X 22.
- Streeter Horizon Va
9-10 implanted ovum 7.5 days of age
11-12 implanted ovum 7.5 days of age

13-14 8 implanted ovum 8 days of age



Plate 3
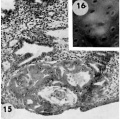
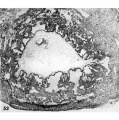
Three 9-day specimens of Horizon Vb, characterized by syncytiotrophoblastic lacunae with early utero-placental circulation, amniogenesis, a simple bilaminar germ disc and only variable degrees of exocoelomie (Heuser’s) membrane formation.

15 Because the decidual reaction is not yet apparent, this ovum is considered to be slightly younger than the other two 9-day specimens illustrated on this plate. The trophoblastic lacunae contain few cellular elements in the plasma. indicating
that at this stage a sluggish circulation begins Within the trophoblastic shell of the ovum. Carnegie 8215, Section 12-5-1-. X100.
16 Surface view of intact specimen. This endomctrium, both grossly and microscopically is slightly younger than that of the following specimens. Carnegie 8215, Sequence 3. X 22.
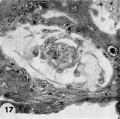
17 Detail of the embryo, chorionie cavity and adjacent trophoblast. Note lensshaped germ disc, amniogenesis from adjacent eytotrophoblast, and delaminating mesoblasts at the abembryonic pole which are continuous with the endoderm and thus together form the exocoelomic membrane. Carnegie 8215, Section 12-54. X 300.
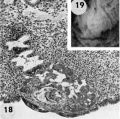
18 Note early predecidual reactions with infiltrating leucocytes about the ovum. Early utero-placental circulation is visible in the form of an empty maternal blood space at right communicating with the coalescing lacunae of the future intervillous space. Carnegie 8171, Section 3-2-11. X 100.
19 The significant feature of this gross implantation site is its inconspicuous nature due to small size of ovum and lack of endomctrial ulceration or congestion. Carnegie 8171, Sequence 3. X 22.
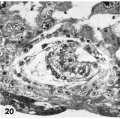
20 Detail of embryo and surrounding trophoblast. Note bilaminar germ disc with vacuolated ectoderm, early amniogenesis and an early exoeoelomic membrane. delaminating from the cytotrophoblast, best seen at the abembryonic pole. Note small shallow surface ulcer with alteration of adjacent nuclei. The paler cytotrophoblast lines the chorionie cavity and varies in thickness whereas the darker synrcytiotrophoblast lies peripherally and contains many intracytoplasmic lacunae which are coalescing to form the future intcrvillous space. Note mitotic figure in cytotrophoblast. Carnegie 8171, Section 3-:2-11. X 300.
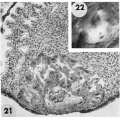
21 The endomctrium has slightly more early decidual reaction than 8171 in figure 18. This specimen is similar in general development to 8171 except that its exocoelomic membrane is not yet formed, possibly because of hemorrhage into the chorionic cavity. See figure 23 for details. Carnegie 8004, Section 11-4-7. X 100.
22 Surface view of intact specimen after fixation and partial dehydration. This was prominent prior to fixation because of size, ulceration of endomctrial surface over the ovum and the unusual feature of hemorrhage into the chorionic cavity. This is the youngest human implantation site that was visible without fixation. Carnegie 8004, Sequence 7. X 22.
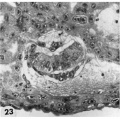
23 Detail of embryo, chorionic cavity and surrounding trophoblast. The maternal hemorrhage in the chorionic and amniotic cavities is abnormal and the result of communication of the trophoblastic lacnmae and chorionic. cavity through a. defective chorionic membrane. A few mesoblnsts are delaminating from the cytotrophoblast but in general this process appears to be delayed perhaps due to the above mentioned hemorrhage. Note mitotic figure in cytotrophoblast. Carnegie 8004, Section 11-4-4. X 300.
- Streeter Horizon Vb
15-16 9-day specimens
17 Detail embryo, chorionie cavity and adjacent trophoblast
18-19 predecidual reaction
20 Detail embryo and surrounding trophoblast
21-22 endometrium and intact specimen
23 Detail of embryo, chorionic cavity and surrounding trophoblast






Plate 4
Three normal, late previllous ova 11 to 12 days old, of Horizon Vc. characterized by intereommunieating laeunar spaces filled with maternal plasma containing still increasing numbers of blood cells, a distended chorionic. cavity, an execoelomic cavity of maximum size, a bilaminar germ disc and well~defined amnion and amniotic cavity. Details of the germ discs of these ova are shown on plate 6.

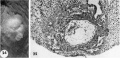
24 Surface view of an intact. ll-day implantation site showing wrinkling of endomctrium between and at the gland mouths. From the endometrial defect created by the ovum arises a. coagulum which communicates with maternal blood within the lacunae. Carnegie 7699, Sequence 8. X 22.
25 A medium-power view of a mid-cross section of the ovum shown in figure 2-}. Note that the endometrial epithelium is proliferating to repair the penetration defect. The cytotrophoblast, immediately surrounding the chorionic cavity, gives rise, by peripheral proliferation and (lifferentiation, to irregular masses which grow into and become invested by syneytiotrophoblast, thus constituting the primordia of chorionic villi. Note syncytium surrounding an endometrial gland and maternal white blood cells in the lacunae. Carnegie 7699, Section 8-5-3. X 100.
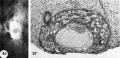
26 Surface view of an intact 12-day-ovum, transilluminated after clearing, showing ehorionic cavity as a dark central area with surrounding opaque but vaeuolated trophoblast. Note wrinkled endometrial surface with gland months evident as dark pits. Carnegie 7950, Sequence 10. X 22.
27 A medium-power view of a mid-cross section of the ovum shown in figure 26. The edema of the endometrinm, though variable, is characteristic of specimens of this general age. Note the distended exocoelomic cavity, the coalesced lacunar spaces showing evidence of an early utero~placental circulation and the budding of the cytotrophoblast to form primordia. of villi. The penetration defect has not been completely healed. Note the cross-section of a gland at the left margin of the ovum. Carnegie 7950, Section ll-5-2. X 100.
28 A surface view of an intact 12-day ovum after fixation, dehydration and clearing to show the appearance of maternal blood in the lacunar spaces. This is the most distinctive gross feature of, and first. appears at, this stage of develop ment. It is due to flooding of the trophoblastic. lacunae with maternal blood. Carnegie 7700, Sequence 9. X 22.
29 A medilun-power view of a. mid-cross section of the ovum shown in figure 28. Early decidua is developing in the area iimnediately surrounding the ovum. Note three irregular masses of cytotrophohlast-—primordial villi—projccting into the syncytium. Note fully developed cxocoelomic cavity and membrane with a loose. mesh-work of mesohlasts between it and the cytotroplioblast lining the «ho:-ionic cavity. The trophoblast at the abembryonic pole is poorly differentiated, as contrasted to that at the embryonic or implantation pole. Carnegie 7700, Section 6-1-5. X100.
- Streeter Horizon Vc
24-25 surface and mid-cross section view 11-day ovum
26-27 surface and mid-cross section view 12-day ovum

28-29 surface and mid-cross section view 12-day ovum



Plate 5
Two normal 12-day ova also in Horizon Vc, characterized by intereommunicating lacunar spaces with evidence of early utero-placental circulation, a distended chorionic cavity, an exocoelomie cavity of maximum size, a bilaminar germ disc and a well-defined amnion and amniotic cavity. Details of their germ discs are shown on plate 6.

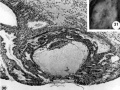
30 A medium-power view of a mid-cross section of the ovum shown in figure 31. A coagulum is lying over the uuhealed penetration defect as evidence of seepage of maternal blood and plasma from laennar space into the uterine lumen. Note the persistent edema of the endometrium and the early decidua developing immediately around the ovum. Note primordial villi and also the gland on the left which has been surrounded by syncytimn. A dilated blood vessel is seen at the margin of the ovum at the right. Carnegie 8558, Section 10-1-4. X 100.
31 A surface view of an intact 12-day implantation site to show the endonietrial detect, the source of maternal bleeding into the uterine lumen at. this and slightly later stages. Note the small tabs of coagalum in the ulcer. A hemorrhage, which became detached and hence is not shown, arose from this defect. Carnegie 8558, Sequence 5. X22.
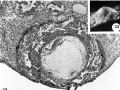
32 A medium-power view of a mid-cross section of the ovum shown in figure 33. Note the early deeidual reaction about the ovum, the primordiuni of a villus on the left, the few blood cells within the lacnnar spaces and the large exocoelomic cavity. Carnegie 8330, Section 9-2-6. X 100.
33 A profile View of a. 12-day ovum to show its elevation from the surrounding endometrium. Carnegie 8330, Sequence 3. X 12.
- Streeter Horizon Vc
30-31 12-day ovum
32-33 12-day ovum


Plate 6
High-power views of the germ discs of the five 11- to 12-day ova of Horizon Vc illustrated previously on plates 4 and 5.

34 The germ disc and anmion of an 11-day ovum. Note the orderly arrangement of cylindrical vacuolated ectodermal cells, beneath which is the irregularly arranged layer of vacuolated and foam_\'-appearing polyhedral cells of the endoderm. Dorsal to the germ disc is the amnion, delaminated in situ from the overlying cytotrophoblast and attached to the peripheral margin of the ectoderm. Note a. few mesoblasts also of cytotrophoblastic origin external to the amnion and merging with those at the junction of the endodermal plate and the exocoelomic membrane. Carnegie 7699, Section 8-5-3. X 300.
35 The germ disc and amnion of a 12-day ovum. Occasional irregularly distributed vacuoles are seen in the ectoderm. Note the thin layer of polyhedral cells of the endodcrmal plate which joins laterally with the elongated mesohlasts of the exocoelomie membrane. Note the fiat amniotic cavity with amniogenesis from the overlying cytotrophoblasts. Carnegie 7950, Section 11-5-2. X 300.
36 The germ disc and amnion of a 12-day ovum. Note the mitotic figure of the amniotic cell delaminating from the cytotrophoblast. Note also the ventral orientation of the vacuoles in the eetoderm and the less evident dorsal orientation of similar structures in the endoderm. Carnegie 7700, Section 5-7-7. X 300.
37 The germ disc. of a 12-day ovum. Note that in this specimen the vacuolated ectodermal cells appear to be located peripherally. Carnegie 8558, Section 10-1-4. X 300.
38 The germ disc of a 12-day ovum. Note that vacuolated ectodermal cells appear to be concentrated toward the left and that the cxtraembryonic. mesoblast is more condensed in this area perhaps indicating the beginning of axis formation in this embryo, the oldest in our Horizon Vc. Carnegie 8330, Section 9-2-5. X 300.
- Streeter Horizon Vc

34 germ disc and anmion 11-day ovum

35 germ disc and amnion 12-day ovum

36 germ disc and amnion 12-day ovum

37 germ disc 12-day ovum

38 germ disc 12-day ovum
Plate 7
Three ova of Horizon VI about 13 days of age cliaraeterized by primitive villi and a distinct yolk-sac. Low power views of mid sections of each of these ova are shown on plate 8.

39 Surface view of an intact 13-day ovum found on the posterior uterine wall adjacent to the left lateral sulcns. Note that the endometrial surface has an undulating wrinkled appearance and that the gland mouths are prominent only around the ovum. Carnegie 8672, Sequence 2. X 8.
40 Medium-power view of one of the middle sections of the ovum seen in figure 39 to show detail of the embryo and adjacent trophoblast-. Note small double-layer yolk-sac. No villi are seen in this photograph but they are evident at tho ahembryonic pole as shown in figure 45. Note increase in size of amniotic cavity. Its continuity with the trophoblast may be evidence of continuing mnniogenesis from this tissue. Vacuoles are not apparent in the ectoderm but are present in the endoderm. Carnegie 8672, Section 11-3-2. X 100.
41 Surface view of an intact implantation site of about 13 days. Note smooth dome-shaped elevation of ovum and the marked undnlation of cndometrial surface with fine and coarse wrinkling. '1‘he stellate crevices represent closed gland mouths. Carnegie Embryo 8360, Sequence 3. X 8.
42 A medium-power view of one of the middle sections of the ovum seen in figure 41. Note that the yolk-sac is single-layered, the ectoderm non-vacuolated (a usual feature from now on), and the extrnembryonie mesoblast is prominent in the area lateral and dorsal to the amnion. Carnegie Embryo 8360, Section 27-4-2. X 100.
43 Surface view of a 13-day ovum. Note large tab of coagulated maternal blood (a variable but normal feature at this stage) which has escaped from the underlying trophoblastic lacunae through the endomctrial defect. The undulnted and wrinkled surface endometrium is now beginning to resemble decidua. Carnegie 7801, Sequence 2. X 8.
44 A medium-power view of a mid~cross section of the ovum shown in figure 43. Note that the eetoderm laterally has a dorsally curved margin and that the yolk-sac is double-layered. The stulJb_v, unbranched primitive villi have formed about the entire periphery of the chorion and contain extraembryonic mesoblast or primitive fibroblastic tissue. The peripheral tips of these villi will extend outwardly to the endometrium and, as they coalesce at the placental floor, form the anchorage for the developing ovum. Axial differentiation is just appearing in this embryo. The primordium of the primitive streak and a few cells of body mcsoderm are found in sections more cnudad than the one illustrated. Carnegie 7801, Section 12-1-1. X 100.
- Streeter Horizon VI
39 surface view 13-day ovum
40 middle section 13-day ovum

41 surface view 13-day ovum
42 middle section 13-day ovum

43 surface view 13-day ovum
44 mid~cross section 13-day ovum






Plate 8
Sections at low-power magnification of the three 13-day ova of Horizon VI shown on plate 7. They are characterized by primitive unbranched villi and a distinct yolk-sac.

45 A 13-day ovum with a yolk-sac and beginning villi. Note undulating margin of chorionic cavity as the extraembryonic mesoblast forms shallow, broad cores of the villi. Also note yolk-sac formzition with remnants of the disrupted transitory exocoelomic membrane lying in the chorionic cavity ventral to the embryo. Note further the early decidual reaction imniedititely around the ovum. Carnegie 8672, Section 11-3-2. X 35.
46 A 13-day ovum with yolk-sac and beginning villi. Note decidual reaction and diluted sinusoids. Also note blood-filled gland lumen at the base of the ovum. This is a frequent finding at this stage of development and is due to leakage of blood from the lncunur spaces into the gland lumen following erosion of the gland epithelium by the invading trophohlast. It is often mistaken for a thrombosed blood vessel. Carnegie Embryo 8360, Section 27-4-1. X 35.
47 A 13-day ovum with unbranched villi and yolk~s:ie. Note remnant of exocoelomic cavity at abembryonic pole of chorionic cavity, many villi and evidence of more active circulation within the lacunar (now properly termed. intervillous) space. Note also the distended and blood-filled gland at the base of the ovum and the coagulated plasma (“Schlussconguluni” of Peters) at the embryonic pole. This maternal blood and plasma, leaking out from the intervillous space, is continuous with the gross blood clot seen in figure 43. Numerous detached darkly staining syncytial giant cells are scattered among the decidual cells immediately surrounding the ovum. Carnegie 7801, Section 12-1-1. X 35.
- Streeter Horizon VI

45 13-day ovum with a yolk-sac

46 13-day ovum
47 13-day ovum


Plate 9
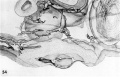
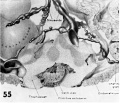
Two villous ova of 16 to 17 days of age. Streeter Horizon VII encompasses that developmental stage characterized by branching villi and an axis of the germ disc, whereas Horizon VIII includes that stage characterized by Hensen’s node and primitive groove. The younger of these two ova has not only branching villi and :1 well-defined axis of the germ disc, but also a definite primitive streak (although not a very distinct primitive groove) and the first beginnings of a head process. Heuser et. al. when reporting this specimen in detail (K15) observed the apparent overlapping of the criteria listed for Horizon VII. and VIII, and considered the younger specimen (7802) an advanced member of Horizon VII. 'l‘he embryo of the older specimen as yet has not been studied in detail and ultimately may well be placed at least. in Horizon VIII. in this publication our primary interest is in the extra-embryonic structures of these ova.

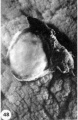
48 A surface view of an intact 16-day ovum. Note coarsely wrinkled and undulating appearance. of endometrial surface, the dome-shaped elevation of ovum and the blood clot of maternal origin. Carnegie 7802, Sequence 5. X 8.
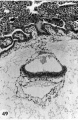
49 A section to show the embryo and adjacent trophoblast. Note the dorsally concave ectoderm, thin endoderm and intervening head process. The double-layered yolk-sac shows angiogcnesis. Angioblasts and mesoblasts individually derived from cytotrophoblast are found also in the condensed mesoblast at the periphery of the. chorionic cavity and in the mesoblastic, cores of the villi. Note also the double-layered anmion with a condensation of mesoblast between it and the periphery of the chorionic cavity. Carnegie 7802, Section 44-3-5. X 100.
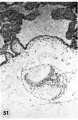
50 A surface view of the anterior wall of a uterus containing a 17-day ovum. The endometrial surface was pale, edematous and thickened and grossly resembled decidua. Note velvety undulations and stellate gland mouths. The ovum lies near the right. lateral sulcus, the fundus being the myometrium whose cut curface is shown on the left. Carnegie 8602, Sequence 1. X 1:}.
51 A medium-power view of a section of the ovum illustrated in figure 50 to show detail of the embryo. Note that the amnion has increased in size and is enfolding the germ disc and that. there is increased differentiation of the ectoderm and cndoderm at the right. The stems of two villi seen at the upper left have a mesoblastic core covered by inner Langhans epithelium (cytotrophoblast) and an outer thin layer of syncytium. Early angiogenesis is seen in the mesoblast of these villi t'oresha«lowing the development of the future fetal circulation which will be completed and functioning in 4 or 5 days (7-17 somites). During this period of time the amount of blood circulating through the spiral artcriolar sinusoids, the intervillous space and the venous sinusoids of the utero-placental circulation will also increase. Thus are establishcd the two contiguous circulations, separated only by the trophoblast, fibrous tissue and endothelium — the placental barrier. By this mechanism there results an interchange of nutritive, waste and other substances between the embryo and mother. Carnegie 8602, Section 25-3-2. X 100.
- Streeter Horizon VII
48 surface view 16-day ovum
49 section 16-day ovum

50 surface view anterior wall uterus 17-day ovum
51 section 17-day ovum




Plate 10
Low-power views of sevtions from the two villous ova illustrated on plate 9.

52 A 16-day ovum showing branched villi on the entire external surface‘ of the chorion. Note that the cytotrophoblast at the peripheral tips of the villi is beginning to coalesce to form the plat-o11t:1l floor which is peiietrated by nmternal blood vessels of the utero-placental circulation. Note large sinusoid at upper right. The decidual reaction around the entire surface of ovum is prominent, thus foreshadowing the formtion of the decidual basalis and capsularis. Carnegie 7802, Section 4-4~3—5. X 30.
53 A 17-day ovum showing the further development of the branching villi with their anchorage to the eytotrophoblust of the placental floor. Note expanded intervillous space connecting with numerous prominent thin-walled maternal sinusoids thus constituting the utero-placental circulation. This is the beginning of :1 more active flow of innternzil blood through the placenta although small but increasing amounts of such blood have been present since the 10th (lay. Note also the decidua around the ovum with many darkly staining tlettielied syncytiotrophoblastic giant cells. Carnegie 8602, Section 25—3—2. X 30.
- Streeter Horizon VII
52 A 16-day ovum
53 A 17-day ovum


Plate 11
Drawings made from transparent three-dimensional plastic reconstructions of ova of 7, 8, 9, 11 and 12 days of age.

54 Reconstruction of half a 7-day ovum, which presents an uneven surface of its solid trophohlast (stippled) to the adjacent endometrium. The trophoblast has alrezuly surrounded a. capillary sinusoid (left). The cut surface of the hilaminar germ disc is evident (endoderm cellular, ectoderm solid). Note primitive amniogenic cells above tiny amniotic cavity. The maternal sinusoidal network is merely the arteriovenous capillary network of the endometrium that has begun to undergo hypertrophy and dilation. See figures I1 and 12. Carnegie 8020. X 260.
55 Reconstruction of half an 8-day ovum viewed from the level of the. greatest diameter of the embryo. The top section of the reconstruction coincides with the photomicrograph of this ovum in figure 13. The trophoblast (stippled) is still solid and is encircling the tips of maternal siuusoids which, although prominent, actually contain very little blood. The large sinusoid toward the right is interpreted as venous in type, whereas the coiled smaller one to the left is a spiral arteriole. The existing capillary anastomoses between this endometrial arteriole and vcnule are the structures that actually grow and dilate under the stimulus of the immediately adjacent trophoblast. Note amniotic cavity but lack of amnioblasts. dorsally concave hilaminar germ disc and mesoblasts within the chorionic cavity. See figures 13 and 14. Carnegie 8155. X 200.
56 Reconstruction of half a 9-day ovum and surrounding endometrium, viewed from the level of the section illustrated in figure :31. Note bilamiuar germ disc and amnion still attached at one point to the overlying‘ trophoblast. Although the trophohlastic lacunae appear discontinuous on the cut surface they actually are a continuous laeunar network in the depths of the trophoblast. Note several points at which the surrounding endometrial sinasoids communicate with this laeunar network. The latter is still ischemie presumably due to spasm of the arteriolar blood supply. See figures 21-23. Carnegie 8004. X 100.
57 A non-schematic line drawing of an 11-day ovum to illustrate the uterolacanar circulation at this developmental stage. In contrast to the cross-sectional views of the other reconstructions on this plate, this is 9. view of the external aspect of the ovum. Note the vein and spiral arteriole with arteriovenoas capillary anastomoses. The growth and dilatation of the large sinusoid continuous with and derived from such an anustomosis are presumably due to the presence of the adjacent trophoblast. Note that the glands are represented merely as stumps to allow uninterrupted view of the utero-laeunar circulation. See figures 24, :35 and 34. Carnegie 7699. X 50.
58 Reconstruction of half a 12-day ovum to illustrate the details of the embryo and chorionic cavity. The top surface of the reconstruction corresponds to the section in figure 29. Note intercommunicating nature of laeunar space which is confluent with capillaries of the large arteriovenous plexus in the surrounding cmlomctrium. The filmy trabeculae of the cxtraembryonic mesoblast. support the exocoelomic membrane (thus accounting for the shape of its enclosed cavity), germ disc and cleft-like amniotic cavity. See figures 28, 29 and 36. Carnegie 7700. X 80.
- 7, 8, 9, 11 and 12 days
54 Reconstruction half a 7-day ovum
55 Reconstruction half an 8-day ovum
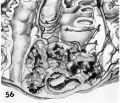
56 Reconstruction half a 9-day ovum
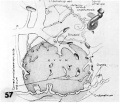
57 A non-schematic line drawing 11-day ovum
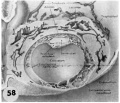
58 Reconstruction half a 12-day ovum





Plate 12
Four abnormal ova of the pre-implantation stage. They all appear to be 3 to 5 days of age on the basis of coital data, endometrial findings and ovular development. They have, in common, the abnormality of nulltinucleated blastomeres. In addition some show variable degrees of cellular degeneration or necrobiosis and it appears doubtful that they would have retained the ability to implant. even though surgery had not intervened.

59 An intact abnormal 5-cell morula showing variation in the size and shape of the blastomeres each of which contains a number of small rounded nuclear fragments. Note intact zona closely applied to the blastomeres, an artifact of fixation. Carnegie Embryo 8630, Sequence 7. X 500.
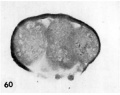
60 A section of the ovum shown in figure 59 to sho\v the variation of nuclear form. The blastomere at the left contains numerous small rounded nuclei or nuclear fragments. The middle blastomere contains a large indistinct nucleolus and the one on the right contains three pairs of chromatin masses that strongly suggest, a disintegrating mitotic figure of triaster type. Note two polar bodies below and artifactitiously ruptured zona. Carnegie Embryo 8630, Section 9. X 500.
61 An intact abnormal 8-cell morula to show its flattened shape. When viewed from above as an intact object it was disc-like with some blastomeres obviously multinucleated and others indistinct and foamy. Carnegie Embryo 8450, Sequence ll. X 500.
62 A section through the greatest diameter of the ovum shown in figure 61. Note the two nearly normal blastomeres on the right contrasted with the two paler ones whose nuclei are undergoing degeneration. Other blastomeres not shown in this section were. multinucleate and/or indistinct with nuclear masses or dense nuclear fragments. Carnegie Embryo 8450, Section 8. X 500.
63 A drawing of a glass-plate reconstruction of a 9-cell abnormal morula. Note artifactitious rupture of zona, unexplained cellular fragments and nuclear detail in the various blastomeres. Carnegie Embryo 8190. X 500.
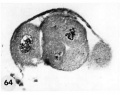
64 A section of the ovum shown in figure 63. Note variation in size and shape of the blastomeres (many in other sections were multinucleated), the cleft. which may indicate early formation of a segmentation cavity and the mitotic figure. Carnegie Embryo 8190, Section 5. X 500.
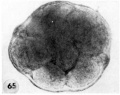
65 An intact abnormal 11- to 12-cell morula showing the intact zona, the position of the two polar bodies and the size, shape and arrangement of the blastomeres.Carnegie 8452 Sequence 3. X 500.
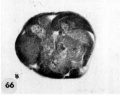
66 A section through the greatest diameter of the ovum shown in figure 65. Note intact zona, the polar body and the variation in size, shape and staining reaction of the blastomeres. The pale, larger cells may be of potential embryonic nature, whereas the darker irregular multinucleated ones may be of future trophoblastic type. Carnegie 8452, Section 8. X 500.
- Abnormal 3 to 5 days
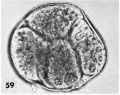
59
60
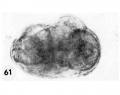
61
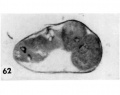
62
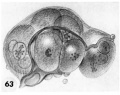
63
64
65
66








Plate 13
Three abnormal implanted ova, one consisting of trophoblast only, one with trophoblast and a small chorionic cavity but no embryo and a third with trophoblast and embryo but no chorionic cavity. It is, perhaps, conceivable that abnormal morulae similar to those illustrated in figures 59456 could have produced this sort of abnormal implantation but, in any event, this type of pregnancy is inevitably destined to abort. Clinical evidence of pregnancy in these patients would have been problematic, depending upon whether the trophoblast were of such quality as to influence the persistence of the corpus luteum.

67 An intact implantation site of an ovum of about. 11 days which was grossly abnormal because of the flattening and radial wrinkling of its endometrial surface. Carnegie 8329, Sequence 2. X 22.
68 A mid-cross section of the ovum shown in figure 67. Note the absence of germ disc, chorionic. cavity and cytotrophoblast. Although syncytiotrophoblast is present, its lacunar development. is abnormal with evidence of circulation only in the equatorial region of the. ovum. The lack of cytotrophoblast and its asso» ciated chorionic gonadotropic. hormone probably would have resulted in early abortion. Carnegie 8329, Section ]342»3. X 100.
69 A polypoid hemorrhagic implantation site of an ovum of about 12 days of age. The type of implantation site is in itself abnormal although it gave no clue as to the fundamental nature of the ovular abnormality shown in figure 70. It is impossible to prove whether this gross appearance resulted from the ovum implanting on an endometrial polyp even though there were many other polyps present. Carnegie 7771, Sequence 1. X 1-1.
70 Section through the greatest diameter of the specimen shown in figure 69. Note prominence of surrounding endometrium (suggesting previous polyp), defective trophoblast with pathologically large lacunae and small chorionic, cavity. The latter contains no recognizable germ disc although there is a mesoblastic membrane faintly reminiscent of that of the cxocoelom. This ovum would undoubtedly have aborted and is an example of the commonest cause of spontaneous abortion — that due to lack of or early death of the embryo. Carnegie 7771, Section 34-]. X 100.
71 Implantation site from a pathologic ovum of about 9 days of age. Note irregular contour and flattened appearance with relatively large ulceration. Although this is not typical of a normal implantation site of a late previllous ovum, the type and degree of abnormality could not have been foretold. Carnegie 8370, Sequence 5. X 22.
72 High-power detail through germ disc and amniotic cavity of specimen seen in figure 73. Note close apposition of trophoblast to germ disc. Carnegie 8370, Section 6-3-5. X 300.
73 Section through largest diameter of specimen seen in figure 71. Although there is a germ disc and amniotic cavity (fig. 72), there is essentially no development of the chorionic cavity. This defect, together with the laminar rather than the circmnfcrential arrangement of syncytio- and cytotroplioblast, makes this a pathologic ovum. It is inconceivable that it could have developed much further although it is impossible to predict the precise type of pathologic abortns that would have been produced by this ovum. Carnegie 8370, Section 6-3-5. X 100.
- Abnormal 9 to 12 days
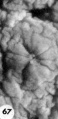
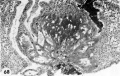
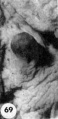
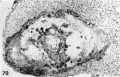
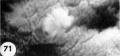

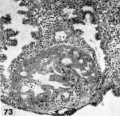







Plate 14
Two abnormal previllous ova, of about 12 to 13 days of age, with hypoplastic trophoblast and moderate to extreme dilatation of the lacunar spaces. These ova are probably older than their apparent trophoblastic development as suggested by the advanced decidual reaction and hemorrhage into a gland, features usually associated with normal early villous ova (figs. 39-47).

74 A medium-power view of the section through the greatest diameter of the younger of these abnormal previllous ova showing thin trabeculae of syncytium with dilated lacunae and a relative lack of extraembryonic mesoblast within the ehorionic cavity. Note formation of primordial villus, amniogenesis aml normal appearing germ disc with distended exocoelom. It is doubtful how much longer this deficient placenta would have maintained the integrity of corpus luteum function. It would appear that early abortion might well follow the resulting hormonal deficiency. In any event spontaneous abortion is caused by or associated with variable degrees of placental hypoplasia. Carnegie 7770, Section 12-3-5. X 100.
75 Surface view of the implantation site of the abnormal ovum shown in figure 74 to show its grossly normal appearance. Note rounded contour and ulcer ation of implantation site with surrounding normal endometrium. Carnegie 7770, Sequence 5. X 22.
76 Surface view of the normal appearing implantation site of the abnormal ovum shown in figure 77. Carnegie 7850, Sequence 4. X 20.
77 A medium-power view of the section through the greatest diameter of an abnormal previllous ovum to show the extreme dilatation of the trophoblastie lacunae. Note the marked hypoplasia of the syneytiotrophoblast which is reduced to fine irregular strands and the relative paucity of cytotrophoblast and the primordial villi resulting therefrom. Note well-formed amnion but relatively small germ disc. The development of the yolk-sac cavity beneath the latter is inconsistent with that of the trophoblast. The primary defect probably lies in the trophoblast with corresponding deficiencies in the structures derived therefrom. Note also the significant deeidual reaction with pronounced leucoeytic infiltration. Inevitable early abortion can be predicted for such a specimen on the basis of its lack of ability to maintain function of the corpus luteum. Such an abortion might take the form of a profuse flow not much later than the next expected period and thus give no undisputed evidence of its true nature. Carnegie 7850, Section 14-3-4. X 100.
- Abnormal 12 to 13 days
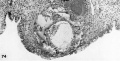
74
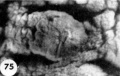
75
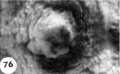
76
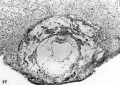
77




Plate 15
An ovum of about 14 days developmental age which should have chorionic villi but shows only extreme hypoplasia of the trophoblast.

78 Surface View of the implantation site to show a hemorrhagic mass attached to the abembryonic pole of the implanted abnormal ovum. Similar hemorrhage may however be associated with normal specimens of the same developmental age. Carnegie 7800, Sequence 2. X 8.
79 Profile view of implantation site to show the elevation of the hemorrhagic mass over the implanted abnormal ovum. Carnegie 7800, Sequence 3. X 8.
80 Low-power view of the section through the largest diameter of the abnormal ovum shown in figures 78 and 79. The trophoblast around three-quarters of the ovum is extremely hypoplastic and is markedly abnormal in the other one-quarter where there is some attempt to form villi. Note that the germ disc and the formation of amnion and yolk-sac appear morphologically to be unremarkable. This stage of ovular development would coincide with the time of the patient’s next expected period. The deeidual reaction and hemorrhage within the endometrial gland are normal. Abortion might be imminent on the basis of the deficiency of the ehorionic gonadotropin produced by the placenta which maintains corpus luteum function. Actually it is impossible to predict the precise time of such an inevitable abortion. Carnegie 7800, Section 15-1-3. X
- Abnormal 14 days
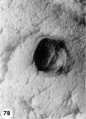
78
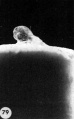
79
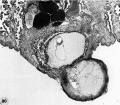
80



Plate 16
Two abnormal ova showing malposition of the germ disc and, in one of the specimens, shallow implantation. Whether these ova would ultimately have aborted it is impossible to say.

81 A surface view of the normal appearing implantation site of the abnormal ovum shown in figure 82. Note endometrial defect. over the ovum, normal for this stage of development. Carnegie 8299, Sequence 4. X 22.
82 A medium-power view of the section through the greatest diameter of a 12-day abnormal ovum shown in figures 81 and 83. Note the malposition of the embryonic disc. The normal amniotic development indicates that the malorientation is of recent development since the amnion has been shown to delaminate in situ from adjacent trophoblast at about 7 days of age (Hertig, ’-15). Hence it appears that this normal appearing embryonic. disc has only recently shifted its position due to some unknown factor. It is possible that this maloricntation would have been corrected as the ovum grew and that tile germ disc would have become reoriented. Whether this patient would have aborted ultimately it is impossible to predict, though probably not. It is impossible to say what effect, if any, such an abnormality would have upon the development of the body stalk and the embryo. Carnegie 8299, Section 11-4-5. X 100.
83 A surface view of the posterior wall of a uterus showing the position and sim of the implantation site shown in figures 81 and 82. Carnegie 8299
84 A low-power view of a section of an abnormal 13-day villous ovum showing shallow implantation and malorientation of the germ disc. Note polypoid type of implantation which may have resulted from failure of proper trophoblastic development at the embryonic pole. The polypoid protrusion may have caused mechanical pressure from the opposite wall of the uterus resulting in distortion of the chorionic. cavity with consequent buckling of the germ disc. This feature is shown in detail in figure 86. Carnegie 8290, Section 25-2-4. X 20.
85 Profile view of implantation site of ovum shown in figures 8-1 and 86. Note polypoid structure due to shallow implantation of ovum rather than to intrinsic endomctrial polyp formation. The superficial hemorrhage is normal. Carnegie 8290, Sequence 7. X 5.
86 A medium-power view of a section of the abnormal ovum shown in figures 84. and 85. Note the malorientation of the embryonic disc with a portion of it still attached to the poorly developed avillous trophoblast at the embryonic pole. The fate of an ovum such as this is unpredictable. This shallowly implanted ovum. with its excellent nbembryonic trophoblast, potentially capable of covering the internal 0a, is possibly related to llofl‘meier’s concept of the etiology of placenta previa. It is interesting to note that 8 months prior to h,vsterectom_v this patient (a para X), delivered a slightly premature stillborn baby whose death was caused by placenta previa, a condition which tends to be somewhat repetitive. Carnegie 8290, Section 25-2-4. X 100.
- Abnormal Ova
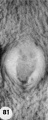
81
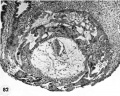
82
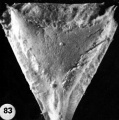
83
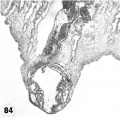
84
85

86





Plate 17
The implantation site of a 12-day previllous ovum, normal except for a shallow type of implantation.

The implantation site of a 12-day previllous ovum, normal except for a shallow type of implantation.
87 Surface view of the intact implantation site showing the excessive hemorrhage above the ovum, best appreciated in the profile view shown in figure 88. Carnegie 8000, Sequence 2. X 5.
88 Profile view of the intact implantation site to show that the excessive hemorrhage completely conceals the shallowly implanted ovum seen in figures 89 and 90. Carnegie 8000, Sequence 3. X 5.
89 A low—power View of a section of the ovum to show the excessive hemorrhage from its normally defective decidua capsularis giving the ovum a more polypoicl appearance than its depth of implantation would actually warrant. Note progestational hyperplasia of endometrium normal for this stage of pregnancy. Carnegie 8000, Section 11-3-3. X 20?“,
90 A medium-power view of a section of the ovum in figures 87-89 to show the essential normality of the specimen, aside from its shallow implantation. It is doubtful whether this pregnancy would have resulted in abortion, although such a shallow implantation might possibly result in the formation of a circumvallate placenta. This anomaly of placental development may in turn cause abortion although many pregnancies whose placentas are of circumxrallate type do go on to term. Carnegie 8000, Section 11-3-2. X 100.
- Abnormal Implantation 12 days

87 Surface view of the intact implantation site

88 Profile view of the intact implantation site

89 A low-power view of a section of the ovum
90 A medium-power view of a section of the ovum



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
All Figures
- Hertig1956 Figures”
1 Two-cell segmenting egg
2 Mid-serial section two-cell
3 Intact 58-cell blastocyst
4 Mid-serial section 58-cell blastocyst
5 An intact 107-cell blastocyst
6 Mid-serial section 107-cell blastocyst
7 Endometrium
8 Mucosa fallopian tube
9-10 7.5 days
11-12 7.5 days
13-14 8 days
15-16 9-day specimens
17 Embryo, chorionic cavity and adjacent trophoblast
18-19 pre-decidual reaction
20 Embryo and surrounding trophoblast
21-22 endometrium and intact specimen
23 Embryo, chorionic cavity and surrounding trophoblast
24-25 surface and mid-cross section 11 day
26-27 surface and mid-cross section 12 day
28-29 surface and mid-cross section 12 day
30-31 12 day
32-33 12 day
34 germ disc and anmion 11 day
35 germ disc and amnion 12 day
36 germ disc and amnion 12 day
37 germ disc 12 day
38 germ disc 12 day
39 surface view 13 day
40 middle section 13 day
41 surface view 13 day
42 middle section 13 day
43 surface view 13 day
44 mid~cross section 13 day
45 13 day with a yolk-sac
46 13 day
47 13 day
48 surface view 16 day
49 section 16 day
50 surface view anterior wall uterus 17 day
51 section 17 day
52 16 day
53 17 day
54 Reconstruction 7 day
55 Reconstruction 8 day
56 Reconstruction 9 day
57 line drawing 11 day
58 Reconstruction 12 day
59 abnormal 5-cell morula
60 section abnormal 5-cell morula
61 abnormal 8-cell morula
62 section abnormal 8-cell morula
63 abnormal 9-cell morula
64 section abnormal 9-cell morula
65 abnormal 11- to 12-cell morula
66 section abnormal 11- to 12-cell morula
67 implantation site 11 days
68 cross section 11 days
69 polypoid hemorrhagic implantation site 12 days
70 cross section polypoid hemorrhagic implantation site 12 days
71 implantation site abnormal 9 days
72 cross section abnormal 9 days
73 cross section abnormal 9 days
74 abnormal previllous 12 to 13 days
75 implantation site abnormal previllous 12 to 13 days
76 implantation site abnormal previllous 12 to 13 days
77 abnormal previllous 12 to 13 days
78 surface view abnormal 14 days
79 implantation site abnormal 14 days
80 section abnormal 14 days
81 surface view abnormal 12 day
82 section abnormal 12 day
83 surface view abnormal 12 day
84 abnormal villous 13 day
85 implantation site abnormal villous 13 day
86 section abnormal villous 13 day
87 Surface view of the intact implantation site
88 Profile view of the intact implantation site
89 A low-power view of a section of the ovum
90 A medium-power view of a section of the ovum
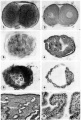
plate 1 fig 1-8

plate 2 fig 9-14
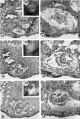
plate 3 fig 15-23
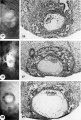
plate 4 fig 24-32
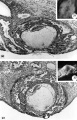
plate 5 fig 30-33

plate 6 fig 34-38
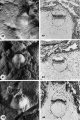
plate 7 fig 39-44
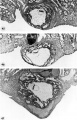
plate 8 fig 45-50
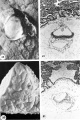
plate 9 fig 48-51
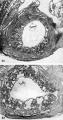
plate 10 fig 52-53
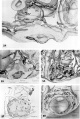
plate 11 fig 54-58
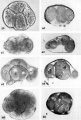
plate 12 fig 59-66
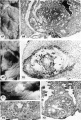
plate 13 fig 67-73
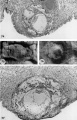
plate 14 fig 74-77

plate 15 fig 78-80
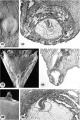
plate 16 fig 81-86
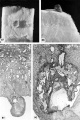
plate 17 fig 87-90
Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - A description of 34 human ova within the first 17 days of development. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_A_description_of_34_human_ova_within_the_first_17_days_of_development
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098GCarnegie Collection[[Category:Carnegie Embryo 7699]
- Draft
- Arthur Hertig
- Carnegie Embryo 8698
- Carnegie Embryo 8794
- Carnegie Embryo 8663
- Carnegie Embryo 8225
- Carnegie Embryo 8020
- Carnegie Embryo 8155
- Carnegie Embryo 8215
- Carnegie Embryo 8171
- Carnegie Embryo 8004
- Carnegie Embryo 7950
- Carnegie Embryo 7700
- Carnegie Embryo 8558
- Carnegie Embryo 8330
- Carnegie Embryo 8672
- Carnegie Embryo 8360
- Carnegie Embryo 7801
- Carnegie Embryo 7802
- Carnegie Embryo 8602
- Carnegie Embryo 7699
- Carnegie Embryo 8630
- Carnegie Embryo 8450
- Carnegie Embryo 8190
- Carnegie Embryo 8452
- Carnegie Embryo 8329
- Carnegie Embryo 7771
- Carnegie Embryo 8370
- Carnegie Embryo 7850
- Carnegie Embryo 7800
- Carnegie Embryo 8299
- Carnegie Embryo 8290
- Carnegie Embryo 8000