Introduction
Monito del monte (Dromiciops gliroides) are small nocturnal marsupials representatives of the ancient order Microbiotheria and also the only known hibernating mammal in South America (Southern Chile and Argentina). Thomas, 1894 original designation Monito del monte (Spanish = "little mountain monkey") or colocolo opossum. Their distribution is within Chile and adjacent Argentina from about 36S to near 43S.
- Links: Category:Marsupial
Some Recent Findings

Distribution of dromiciops gliroides
- Morphology and evolution of the oral shield in marsupial neonates including the newborn monito del monte[1] "Newborn marsupials can be arranged into three grades of developmental complexity based on their external form, as well as based on their organ systems and their cytology. The dasyurids are considered the least developed marsupials at birth, while didelphids and peramelids are intermediate, and macropods are the most developed. Currently there is still little information on caenolestid and microbiotherid development at birth. Developmental stages can be graded as G1, G2 and G3, with G1 being the least developed at birth, and G3 the most developed. Marsupials are also characterized by having an extremely developed craniofacial region at birth compared with placentals. However, the facial region is also observed to vary in development between different marsupial groups at birth. The oral shield is a morphological structure observed in the oral region of the head during late embryological development, which will diminish shortly after birth. ...We find that a simple to develop oral shield structure (type 2-3) may have been present in marsupial ancestors as well as in early therians, even though this structure is not present in the extant monotremes. This in turn may suggest that early marsupials may have had a very simple pouch or lacked a pouch as seen in some living marsupials, such as some dasyurids, didelphids and caenolestids. The study's results also suggest that different morphological stages of the oral shield and hindlimb development may be influenced by species size and reproductive strategy, and possibly by yet unknown species-specific adaptations."
- On the development of the chondrocranium and the histological anatomy of the head in perinatal stages of marsupial mammals[2] "An overview of the literature on the chondrocranium of marsupial mammals reveals a relative conservatism in shape and structures. We document the histological cranial anatomy of individuals representing Monodelphis domestica, Dromiciops gliroides, Perameles sp. and Macropus eugenii. The marsupial chondrocranium is generally characterized by the great breadth of the lamina basalis, absence of pila metoptica and large otic capsules."
- The hibernating South American marsupial, Dromiciops gliroides, displays torpor-sensitive microRNA expression patterns[3] "When faced with adverse environmental conditions, the marsupial Dromiciops gliroides uses either daily or seasonal torpor to support survival and is the only known hibernating mammal in South America. As the sole living representative of the ancient Order Microbiotheria, this species can provide crucial information about the evolutionary origins and biochemical mechanisms of hibernation. Hibernation is a complex energy-saving strategy that involves changes in gene expression that are elicited in part by microRNAs. To better elucidate the role of microRNAs in orchestrating hypometabolism, a modified stem-loop technique and quantitative PCR were used to characterize the relative expression levels of 85 microRNAs in liver and skeletal muscle of control and torpid D. gliroides. Thirty-nine microRNAs were differentially regulated during torpor; of these, 35 were downregulated in liver and 11 were differentially expressed in skeletal muscle. Bioinformatic analysis predicted that the downregulated liver microRNAs were associated with activation of MAPK, PI3K-Akt and mTOR pathways, suggesting their importance in facilitating marsupial torpor. In skeletal muscle, hibernation-responsive microRNAs were predicted to regulate focal adhesion, ErbB, and mTOR pathways, indicating a promotion of muscle maintenance mechanisms. These tissue-specific responses suggest that microRNAs regulate key molecular pathways that facilitate hibernation, thermoregulation, and prevention of muscle disuse atrophy."
- The ancestral chromosomes of Dromiciops gliroides (Microbiotheridae), and its bearings on the karyotypic evolution of American marsupials[4] "The low-numbered 14-chromosome karyotype of marsupials has falsified the fusion hypothesis claiming ancestrality from a 22-chromosome karyotype. Since the 14-chromosome condition of the relict Dromiciops gliroides is reminecent of ancestrality, its interstitial traces of past putative fusions and heterochromatin banding patterns were studied and added to available marsupials' cytogenetic data. Fluorescent in situ hybridization (FISH) and self-genomic in situ hybridization (self-GISH) were used to detect telomeric and repetitive sequences, respectively. These were complemented with C-, fluorescent banding, and centromere immunodetection over mitotic spreads. The presence of interstitial telomeric sequences (ITS) and diploid numbers were reconstructed and mapped onto the marsupial phylogenetic tree. No interstitial, fluorescent signals, but clearly stained telomeric regions were detected by FISH and self-GISH. Heterochromatin distribution was sparse in the telomeric/subtelomeric regions of large submetacentric chromosomes. Large AT-rich blocks were detected in the long arm of four submetacentrics and CG-rich block in the telomeric regions of all chromosomes. The ancestral reconstructions both ITS presence and diploid numbers suggested that ITS are unrelated to fusion events. Although the lack of interstitial signals in D. gliroides' karyotype does not prove absence of past fusions, our data suggests its non-rearranged plesiomorphic condition."
- Comparative gene expression analyses reveal heterochrony for Sox9 expression in the cranial neural crest during marsupial development[5] "Compared to placental mammals, marsupials have short gestation period, and their neonates are relatively immature. Despite these features, marsupial neonates must travel from the birth canal to the teat, suckle and digest milk to complete development. Thus, certain organs and tissues of marsupial neonates, such as forelimbs to crawl and jaw elements to suckle, must develop early. Previous reports showed that cranial neural crest (CNC) cells, as the source of ectomesenchyme of jaw elements, are generated significantly early in gray short-tailed opossum (Monodelphis domestica) compared to other amniote models, such as mouse. In this study, we examined the expression of genes known to be important for neural crest formation, such as BMP2/BMP4 (neural crest inducer), Pax7 (neural border specifier), Snail1 and Sox9/Sox10 (neural crest specifier) in Monodelphis domestica, and compared the expression patterns with those in mouse, chicken, and gecko embryos. Among those genes, the expression of Sox9 was turned on early and broadly in the premigratory CNC cells, and persisted in the ectomesenchyme of the cranial anlagen in opossum embryos. In contrast, Sox9 expression diminished in the CNC cells of other animals at the early phase of migration. Comparison of the onset of Pax7 and Sox9 expression revealed that Sox9 expression in the prospective CNC was earlier and broader than Pax7 expression in opossum, suggesting that the sequence of border specification and neural crest specification is altered."
|
| More recent papers
|
|
This table allows an automated computer search of the external PubMed database using the listed "Search term" text link.
- This search now requires a manual link as the original PubMed extension has been disabled.
- The displayed list of references do not reflect any editorial selection of material based on content or relevance.
- References also appear on this list based upon the date of the actual page viewing.
References listed on the rest of the content page and the associated discussion page (listed under the publication year sub-headings) do include some editorial selection based upon both relevance and availability.
More? References | Discussion Page | Journal Searches | 2019 References | 2020 References
Search term: Monito del monte
<pubmed limit=5>Monito del monte</pubmed>
<pubmed limit=5>Dromiciops gliroides</pubmed>
|
Taxon
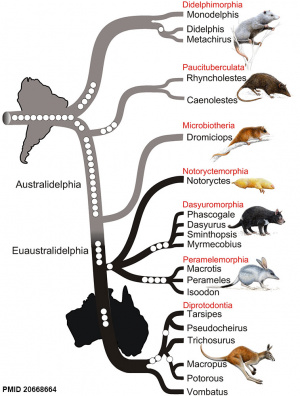
Phylogenetic tree of marsupials derived from retroposon data
[6]Taxonomy ID:
Genbank common name: opossum
Rank: species
Genetic code: Translation table 1 (Standard)
Mitochondrial genetic code: Translation table 2 (Vertebrate Mitochondrial)
Lineage ( full )
cellular organisms; Eukaryota; Opisthokonta; Metazoa; Eumetazoa; Bilateria; Deuterostomia; Chordata; Craniata; Vertebrata; Gnathostomata; Teleostomi; Euteleostomi; Sarcopterygii; Dipnotetrapodomorpha; Tetrapoda; Amniota; Mammalia; Theria; Metatheria; Microbiotheria; Microbiotheriidae; Dromiciops
- Links: Taxonomy Browser - Dromiciops gliroides
Development Overview
Adult female can produce one litter of a maximum of 4 infants per year. Data below from a 2005 study.[7]
- Intra-uterine development - (3-4 weeks) blastocyst implantation, marsupium development is marked by teat enlargement and formation of lactating tissue.
- parturition - neonates migrate from the vagina to the marsupium.
- intra-marsupium - (2 months) pouched young seem to remain within the marsupium and firmly affixed to teats.
- extra-marsupium - (juvenile) Increased motor capacities allow pouched young to leave the marsupium for short exploratory excursions.
Placenta
Choriovitelline placenta - chorion is formed by the fusion of yolk-sac mesoderm and vessels to the inner face of the serosa.
- Links: Placenta Development
System Development
The marsupial neonate at birth has a variation between the degree of development of different systems.[8]
- well-developed - digestive, respiratory and circulatory system.
- not well-developed - retains fetal excretory system with a fully functional mesonephric kidney and undifferentiated gonads and genitalia.
Oocyte
- Links: Oocyte Development
Spermatozoa
Spermatozoa morphology:
Head cuneiform, acrosome restricted to anterior third of dorsal surface
of nucleus, midpiece not preserved; spermatozoan pairing not determined; the single head said to be similar to that of Australian Trichosurus vulpecula (Phalangeridae).
[9]
Sperm pairing is found in all South American marsupials except Dromiciops gliroides [9] and has never been observed in any Australian marsupials. Sperm pairing was first described for the Virginian opossum (Didelphis virginiana) [10] and described as a post-testicular event (Biggers and Creed, 1962) that occurs as spermatozoa passes through the epididymis. It is a feature present in all American marsupials except Dromiciops gliroides.
- Links: Spermatozoa Development
Movies
References
- ↑ <pubmed>28620997</pubmed>
- ↑ <pubmed>28203388 </pubmed>
- ↑ <pubmed>27090740</pubmed>
- ↑ <pubmed>27489568</pubmed>
- ↑ <pubmed>24934187</pubmed>
- ↑ 20668664</pubmed>
- ↑ Muñoz-Pedreros A, Lang, BK Marta Bretos, M. and Meserve, PL. Reproduction and Development of the Monito del Monte, Dromiciops gliroides Thoma (marsupialia: Microbiotheriidae) in Temperate Rainforests of Southern Chile. (2005) Gayana 69(2):225-233 DOI: 10.4067/S0717-65382005000200002 PDF
- ↑ <pubmed>11999321</pubmed>
- ↑ 9.0 9.1 Hershkovitz, P. (1999). Dromiciops gliroides Thomas, 1894, Last of the Microbiotheria (Marsupialia), with a Review of the Family Microbiotheriidae. Fieldiana. Zoology, (93), 1-60.
- ↑ Selenka, E. (1887). Part 4. Das Opossum (Didelphis virginiana); in: Studien über die
Entwickelungsgeschichte der Thiere. Wiesbaden, Kreidels
Reviews
<pubmed></pubmed>
<pubmed></pubmed>
<pubmed></pubmed>
Articles
<pubmed></pubmed>
<pubmed></pubmed>
<pubmed>20668664</pubmed>
Muñoz-Pedreros A, Lang, BK Marta Bretos, M. and Meserve, PL. Reproduction and Development of the Monito del Monte, Dromiciops gliroides Thoma (marsupialia: Microbiotheriidae) in Temperate Rainforests of Southern Chile. (2005) Gayana 69(2):225-233 DOI: 10.4067/S0717-65382005000200002 PDF
Books
Brown BE. Atlas of New World marsupials (2004) Field Museum of Natural History.
Search PubMed
Search PubMed: Monito del monte development | Dromiciops gliroides
External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2026, Haziran 25) Embryology Monito del Monte Development. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Monito_del_Monte_Development
- What Links Here?
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G



