Paper - The Maturation of the Human Ovum
| Embryology - 18 Apr 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Thomson A. The maturation of the human ovum. (1919) J Anat. 53(2-3): 172-208. PMID 17103860
| Online Editor | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

|
| Historic Disclaimer - information about historic embryology pages |
|---|
|
The Maturation of the Human Ovum

{kind=link}
Professor of Human Anatomy, University of Oxford
- The expenses of illustrating this paper were defrayed by the Medical Research Committee.
The human anatomist is handicapped by the nature of the material with which he has to deal. More particularly is this the case when the problem attacked involves the recognition of delicate changes in cellular structure.It is only exceptionally, as in operation cases, that he can get his tissues fresh. Under ordinary circumstances a considerable period must have elapsed, amounting in some cases to days, before the organ or tissue desired can be obtained post-mortem.
Further, the time and manner of death may exercise important influences on the condition of the parts to be examined, for it will be obvious that if death occur during the warm summer months the putrefactive changes that ensue Will be more rapid than those occurring in a cold season. Similarly the processes involved in death after prolonged illness a.re likely to produce changes in the tissues of an abnormal kind, differing in this respect from those met with in sudden death either by accident or acute disease.
From all, or most of these conditions, the zoologist is happily free. Engaged in a like or similar enquiry, he can kill the animal selected whilst still in a normal condition, and thus obtain its tissues and organs fresh. Subjected alike to similar processes of fixation and staining, the results obtained from these two different sources of supply, human, as distinct from animal, must necessarily vary iii the refinement and delicacy of the appearances observed though what the resulting differences may be when comparing the “fresh” with the “stale” tissue, is still a matter of doubt.
In such an enquiry as that here involved, we must be content with What We can get. In the present instance the specimens examined have all been obtained from women who died suddenly or were accidentally killed, but the necessary delay incurred in obtaining and performing a post-mortem examination must militate against the freshness of the material examined. Of the possible changes that may intervene during the period of death and fixation we are ignorant, and Whilst admitting that some such changes may affect the more delicate structures involved, such as the fibrillar appearance of the asters and-the arrangement of the chromosomes, there is sufficient evidence to justify us in assuming that the grosser phenomena involved in the division of the egg, such as the position of the nucleus, the presence or absence of a nuclear membrane, the occurrence of centrospheres and the presence of polar bodies can all be identified, though possibly in a form differing somewhat from those met with in the fresher material.
The utmost care has been exercised in the fixing, cutting, mounting and staining of the tissues examined. Where possible complete serial sections of the eggs examined have been reproduced, on a reduced scale, so that in the plates provided a complete conspectus of the contents of the egg is available.
Thanks to the great industry and skill of my Laboratory Assistant, Mr W. Chesterman, to whose ability and enthusiasm I am greatly indebted, the sections obtained are as good as the conditions permit.
A word or two as to the method of illustration adopted. As the result of our united efforts, Mr Chesterman has acquired a high degree of skill in the production of photomicrographs, these I have utilised without exception in the present paper, for, though laying claim to some experience as a draughtsman, I have come to the conclusion that well executed microphotographs are of much more value than elaborately executed drawings; in the latter the draughtsman is unwittingly led to emphasise those details which he thinks of importance, whilst at the same time he may omit features which for the ' time being he considers of little moment. In numerous instances a structure which at first appeared of no account has been found subsequently to form a link in the chain of evidence.
The production of such microphotographs is tedious and tiresome, and demands an intimate knowledge of the technique required, b11t the results, in my opinion, justify the amount of labour and expense involved. In the present instance the illustrations may be relied upon as affording an accurate record of the appearances described. They are a record of the facts, the reader may deduce from them such conclusions as he thinks justifiable—I ask no more.
In work of this kind, where delicate measurements are involved, I have departed somewhat from the approved method, as I found it so irksome and in some instances so difficult of application——I mean the use of the eyepiece micrometer in conjunction with a stage micrometer. W hat has been done in every case where a photomicrograph was taken was to introduce at the same time and under similar conditions of the optical arrangement a stage micrometer, so that the degree of magnification could be accurately gauged o11 the ground glass of the camera. In this way the amplification of the image on the positive or negative photograph was accurately known, and by a simple calculation in reduction, the magnification being known, the precise meas11re— ments of the various details exhibited in the sections were readily deduced from their measurement on the print.
In regard to the present enquiry — the maturation stages of the human egg — no observations, so far as I know, have yet been published. It may be that like or similar appearances have been seen, but hitherto no attempt has been made to arrange them in their order of sequence.
Our conception of what occurs in the human female has hitherto been based on our knowledge of what takes place in lower forms. So extensive is 174 Arthur Thomson the literature of that subject that no reference need here be made to it, other than to accept the general conclusions based upon it as set forth in the various text-books on Human Anatomy, in which a general description of the changes presumably involved, is set forth. It is assumed that the reader is familiar with one or other of these accounts, and consequently space has been economised by taking such information for granted. It may be stated broadly that the results obtained conform to the general plan of the descriptions given as applicable to the mammalian type, the only point of difference, as will be subsequently explained, depending on the solution of the question asto whether or no the oocyte in man is mature, i.e. has passed through the stages of its first and second division, before it leaves the ovary. It appears on the authority of numerous observers that this is not the rule in’ Vertebrates, and that the sperm enters the ovum during the stage of the second maturation division. If this be so, it will appear on the evidence adduced that man is an exception to the rule. The details, however, on which this and other conclusions are based, had better be postponed until the eggs have been fully examined. We will therefore proceed forthwith with their description.
Here let me say how much I am indebted to my friends holding pathological appointments for their ever ready help in providing me with suitable material.

Fig. 1. Human ovum in discus proligerus in early part of prophase of first division. 1x 600. |
The first figure is that of a section, 0.007 mm thick, stained with Mallory’s Connective Tissue Stain, of an ovum in the discus proligerus of a Graafian follicle from a woman aged 22 who died from a femoral hernia.
|
At one point just opposite the centrosphere there is evidence of cellular structure. The cytoplasm, fairly uniform in character, is separated from the inner wall of the Zona by what may be possibly a retraction cavity, some of the cytoplasm remaining adherent to the inner surface of the Zona.
The nucleus, of size approximately 0.03 x 0.022 mm, is fairly central in position, and has a faint and ill-defined nuclear membrane. The arrangement of the chromatin is best realized by reference to the figure.
What appears to be the nucleolus breaking up is seen towards one end of the nucleus. The nucleus is in part separated from the surrounding cytoplasm by a narrow cleft probably a retraction cavity though possibly it may be a space filled with fluid derived from the karyoplasm.
At one extremity of the nucleus lying within the cytoplasm just outside the faint nuclear membrane is a small granule.
Lying to one side of the nucleus and in contact with the nuclear membrane, which is here only faintly indicated, is a small sphere, 0.013 x 0.011 mm, consisting of a material so clear that it might be mistaken for a vacuole; it has, however, an obvious structure and contains near its poles two more darkly stained granules. This object I take to be the Centrosphere, Attraction Sphere (Quain), Archoplasm Sphere (McMurrich), Centrosome (Robinson), Microcentrum (Heidenhain), Microsphere (Siedlecki), in the young oocyte sometimes called the Idiosome (corresponding to the attraction sphere of other cells). Van der Stricht identifies this with the Yolk nucleus or Body of Balbiani, as does Jenkinson,[1] see also Wilson.[2]
Its contents are variously termed the Centroplasm or Archoplasm, whilst the granules are known as the Centrioles.
Other sections of this egg a.re available, but they exhibit nothing worthy of note. The appearance indicated in the figure, however, justifies us in assuming that the specimen represents the prophase condition in the Human Ooeyte prior to the formation of the first polar body.

Fig. 2. Human ovum in discus proligerus in prophase of first division. x 600. |
Fig. 2 shows an ovum in discus proligerus in Graafian follicle. Section 0.007 mm thick. Stained with Ehrlich’s acid haematoxylin and eosin.
Size including Zona 0.117 x 0.1 mm; inside Zona 0.1 x 0.08 mm. From a woman aged 22. Died of a femoral hernia. Same ovary as fig. 1. The figure shows that the egg is not spherical.
The Zona pellucida consists clearly of two layers, the combined thickness of which amounts to from 0.008 to 0.O11 mm. The three deeply stained bodies seen overlying the Zona pellucida are nuclei which have got floated over it in the process of mounting. |
The inner zone is more homogeneous than the outer, it exhibits here and there evidence of a lamellar arrangement and at one or two points there seems to be distinct proof of the presence of cellular elements in its substance. At one pole it appears to be faintly striated radially.
This layer of the Zona pellucida I have reason to believe is the ovular layer.
The outer layer, continuous with the processes which pass to the cells of the Corona radiata is more open and fibrillar in structure. This I believe to be the ovarian layer of the Zona pellucida, the Zona thus‘ being a compound structure made up of ovular and ovarian elements.
The cytoplasm, of granular appearance, is fairly uniform in density throughout. It is retracted from the inner surface of the Zona pellucida, the retraction cavity being slit-like for the most part, but increased in width towards one end, where it forms a considerable cavity, which is in part occupied by a ‘faintly granular translucent material capping the substance of the cytoplasm at this end of the egg. This is possibly of the nature of a coagulum.
The nucleus measures roughly 0.02 x 0.017 mm and occupies a position nearly central in the section of the cytoplasm. The nuclear membrane has disappeared, and the arrangement of the chromatin in scattered granules is best realised by reference to the figure. Some of these granules appear to fall in line parallel to the equatorof the nucleus.
There is no evidence of a nucleolus, unless the presence of a larger chromatin granule denotes its presence.
The contents of the nucleus (karyoplasm) are everywhere confluent with the cytoplasm.
On either side of, and near, the nucleus are two small clear spots, each containing a highly refractive granule. In the specimen, these lie in line with the greatest diameter of the egg, but it is diflicult to determine whether this corresponds with the true polar axis of the oocyte, since the example is taken from aspecimen in which the serial sections are not complete.
These bodies, I take it, represent the divided centrosphere shown in fig. 1, each half containing its accompanying centriole. The divided centrosphere has now shifted its position so that its two halves come to lie on either side of thenucleus.
Other cytoplasmic inclusions. With the , exception of a deeply stained granule lying some little distance to one side of the nucleus there is no apparent feature in the cytoplasm which calls for comment. I must leave it to those better versed in cytology to determine what the nature of this particle may be.
From the evidence before us we may, I think, assume that fig. 2 represents a further stage in the prophasc condition of the formation of the first polar body exhibited in fig. 1, the centrosphere there displayed having now divided into the two clear spheres each with a eentriole and placed so as to lie on opposite sides of the nucleus, which now devoid of its membrane is approaching the stage when it divides.

Fig. 3. Human ovum in discus proligerus in anaphase of first division and commencement of metaphase of second division. x 600.
Ovum in discus proligerus in Graafian follicle. Thickness of section 0.01 mm, stained with Weigert’s Iron Haematoxylin and Van Giesen. Size including Zona 0.097 x 0.088 mm; inside Zona 0.083 x 0.08 mm.
From a woman whose ovary was forwarded without any history, in the same case a morula and an ovarian blastula were found; these will be described in a subsequent paper. In this case the Fallopian tubes were much twisted and bound down by adhesions—presumably impervious to the descent of an ovum though apparently permitting the passage of spermatozoa.
The section here displays a more spherical appearance.
The cells of the Corona radiata are neither so discrete nor so clearly seen as in the previous figures (1 and 2). They are separated from the outer surface of the Zona pellucida by a zone about 0.001 mm in width, which exhibits but faint indications of structure, the fibrillar arcades being conspicuous by their absence and the tissue intervening, between the outer surface of the Zona pellucida and the innermost row of the cells of the Corona radiata on one side of the egg, appears but faintly striated and granular, whereas on the opposite side of the ovum the interval appears clearer as if occupied with fluid—further, there is a breach in the continuity of the girdle of surrounding coronal cells, as if by the penetration of the liquor folliculi, a stage, possibly, in the liberation of the oocyte from the cells of the discus proligerus immediately around it. It may reasonably be assumed that the arcading of the tissue connecting the cells of the Corona radiata with the Zona pellucida exhibited in the previous figures is part of the same process, brought about by the permeation of fluid (liquor folliculi?) in between.
The Zona pellucida on the two sides of the egg exhibits a marked difference in section, varying from 0.007 mm on one side to 0.003 mm on the opposite side of the ovum, a difference due no doubt to the obliquity of the section.
The innermost, more homogeneous and ovular (‘?) layer of the Zona pellucida alone remains, the external fibrillar or ovarian layer having now all but disappeared on one side, whereas on the other side it is reduced to a faint reticulum. The Zona pellucida left exhibits traces of concentric lamination with a suggestion at one point of cellular structure. In this specimen I can see no evidence of radial striation.
Within the Zona the cytoplasm has shrunk excentrically from the inner wall of the Zona pellucida, leaving on one hemisphere a considerable interval between the two, whereas on the opposite side it forms a linear cleft. This space is occupied by a fine clear granular material (coagulum?) exhibiting three large vacuoles. In the substance of this material at the point where the cavity is widest is a collection of chromatin grains arranged in nuclear form and associated on one side with a small sphere of granular matter, undoubtedly both combined being the first (?) polar body.
If this be so, and there can be little doubt of it, then the question arises whether this perivitelline space is an artefact or is a normal condition during life, in which a fluid liberated from the cytoplasm fills the space. If this be so, then the polar body, as seen in the section, will lie free in this fluid, and as it is presumably of greater weight than the liquid by which it is surrounded, this offers an explanation of how it sinks to one or other pole of the egg. As to which, there is some difference of opinion, for whilst most observers assign to it a position near the animal pole, Van der Strieht (see Quain[3], p. 11) on the other hand states that in the bat the polar bodies occupy a position which afterwards becomes the vegetative pole. If, as we are led to believe, the position of the vegetative pole is determined by the greater weight of the deutoplasm rotating the egg in that direction, then, if the explanation of the position of the polar bodies now offered be accepted, we have to some extent a confirmation of Van der Stricht’s views, for the rotation of the egg and the position of the polar bodies may both result from the same cause, viz. the action of gravity. There seems no evidence to show that the polar body always remains in the position in which it is extruded from the cytoplasm, and there is ground for supposing that the extrusion in an egg which is not truly spherical may take place at some considerable distance from the polar axis, the‘ subsequent disposal of the polar body towards one or other pole of this axis being possibly determined in the manner suggested.
In this specimen the cytoplasm exhibits remarkable characters. We may assume that the appearance presented corresponds to the period immediately after the expulsion of the first (?) polar body, and that the contents of the oocyte display the phenomena associated with the anaphase condition of the first division and the commencement of the metaphase condition of the second division.
Three spheres are seen (the section was 0.01 mm thick), lying, as may be seen from the overlap of their outlines, one in front of the other.
The anterior one, if such a term may be used in this connexion, is a relatively large vesicle (0.03 x 0.026 mm) containing a clear I faintly granular material, the only appearance of definite structure being two faint thread-like filaments of. granular appearance lying close to and parallel with each other. The circumference of the vesicle is clearly defined from the surrounding cytoplasm. Behind this, and in part overlapped by it, is another spherical mass of approximately the same size. It consists of a mass of granular matter of darker tint than that contained in the vesicle just described, yet lighter than that of the surrounding cytoplasm. Scattered throughout its substance are numerous chromatin granules, not, however, so far as one can-see, exhibiting any definite arrangement.
If this be the nuclear material of the daughter cell or Oocyte II, and there is every reason to believe it is, then it appears at this stage to be absolutely larger than the nuclei seen in figs. 1 and 2. The circumference of the nucleus, though clearly defined from the surrounding cytoplasm, exhibits no evidence of a nuclear membrane.
To the further side of the nuclear sphere, and in part covered by it, is a third spherical body of approximately half the size. The difference in size is misleading, for here it is cut unequally, as may be seen on examining fig. 4, which is the next section in series with it. In the present section this vesicle is only faintly differentiated from the surrounding cytoplasm, and A little if any details of structure are observable.
There can be little doubt but that the structures here described are the nuclear material of the daughter cell in the middle with the two centrospheres on either side of it, though, as will have been gathered from the description given, these latter bodies exhibit no evidence of the fibrillation which one would expect to see at this stage; that, however, may be a very transient feature and one which does not militate against this assumption. The fact that the first (?) polar body is seen immediately outside these active changes The M aturation of the Human Ovum 181 Within the cytoplasm and nucleus, justifies us in thinking that the specimen has been sectioned immediately after the extrusion of the first polar body, and prior to the formation of the second polar body, the nuclear mass not having meanwhile undergone the changes associated with the resting condition which will succeed the extrusion of the second polar body.
Unfortunately the serial sections of this specimen are incomplete, so that it is well nigh impossible to form any opinion as to the position or direction of the plane of section, consequently it is difficult to determine whether the section passes through the ovum close to the equator or nearer one or other pole. For this reason the amount of the cytoplasm displayed in the section may consist of but a proportion of that contained in the widest diameter of the cell. As displayed in the figure it will be seen that the cytoplasm on the two sides of the egg, at right angles to the position of the extruded polar body, differs in character and appearance, this may be due possibly to the unequal thickness of the section. On one side it appears darker, the granules appear more deeply stained, particularly along the circumference of the nuclear mass, whereas on the other side, the cytoplasm is less dense, though still darker than the substance of ‘the nuclear mass, and exhibits a cloudy appearance.
In the substance of the cytoplasm between the smaller centrosphere and the periphery of the cytoplasm there is a small mass of more deeply stained substance at the other end of the axis of which the polar body forms one extremity. As to whether this has any significance I must leave an open question.
Fig. 4, thickness of section 0.01 mm, stained with Weigert’s Iron Haematoxylin and Van Giesen. This represents the appearance displayed by the contiguous section to fig. 3; it calls for less detailed description since it represents many of the same details of structure already referred to in fig. 3.
The essential points of difference are: first, that the section has passed above or below the position of the polar body, thus giving us a clue to its size. As the sections are 0.01 mm thick it follows that the largest diameter of the polar body cannot exceed that measure; second, the material (coagulum?) filling the space between the cytoplasm and the Zona pellucida is uniform in structure, and the large vacuoles have disappeared; third, the size of the three spherical vesicles within the cytoplasm is more nearly equal, suggesting that they have been cut in the axis of the amphiaster. Associated with each lateral sphere, which, as we have already noted, is to be regarded as a centrosphere, may be seen a darkly stained granule, occupying a position, if not quite in the centre, at least near it. These centrospheres are not so precisely defined from the surrounding cytoplasm as in fig. 3, nor are their contents so clear; but these are appearances which may be due to the density of the staining, or to the slight variation in the thickness of the sections.
The nuclear material of the daughter cell, which lies intermediate in position between the two centrospheres, exhibits a granular appearance due to the presence of chromatin grains, some of which are arranged in thread-like fashion.
At one point in this mass there is however a definite clearly defined shieldshaped body, containing some highly refractive material, in the substance of which are a few more deeply stained granules. What the significance of this structure is I am not in a position to suggest. A The cytoplasm around the centrospheres and nuclear mass appears darker on one side than the other, the darkest part lying in immediate relation to the nuclear mass on that side where the clear body within the nuclear substance is situated. On the opposite side of the nuclear mass there is a fairly large deeply stained granule. Around the centrospheres the cytoplasm appears more deeply stained, thus helping to define clearly the circumference of the spheres. In the lighter hemisphere of the cytoplasm, three or four more distinctly stained granules may be seen towards the circumference of the cytoplasm.

Fig. 4. Human ovum in discus proligerus in anaphase of first division and commencement of metaphase of second division. x600.
The appearance of this section confirms in every respect the opinion set forth in respect of fig. 3, that the specimen represents the anaphase condition of Oocyte II and the commencement of the metaphase condition and the formation of the second polar body.
Plate X provides a conspectus of a complete series of sections of an oocyte within the discus proligerus iii a Graafian follicle from the ovary of a woman, aged 38, who died of heart failure, one day after admission to hospital.
There are 15 sections, each of 0.007 mm thick, stained with Kernschwarz. In these the egg or its covering appears in 13 sections, so that from this the “ thickness” of the egg at right angles to the plane of section may be estimated at 0.091 mm. The greatest diameter of the egg is met with in sect. 8, where including the Zona pelluci(la it measures 0.11 x 0.1 mm. In this section the diameters taken from the inner surface of the Zona pellucida measure 0.093 x 0.083 mm.
The cells of the Corona radiata are separated from the outer surface of the Zona pellucida by a clear space averaging about 0.006 mm in width. Stretching across this space, numerous groups of fine fibres, with clear intervals in between them, are seen connecting the bases of the inner row of cells of the Corona radiata with the external surface of the Zona pellucida. The intervals between the bundles of fibres are apparently caused by the permeation of fluid (liquor fo1liculi?), and as will be seen in Plate X, the cells of the discus proligerus are being so separated up by the infiltration of the liquor folliculi, that the Corona radiata, so-called, is reduced to but a single row of cells in many parts of the sections shown.
The Zona pellucida, of an average thickness of about 0.008 mm, is seen to consist mainly of the inner or ovular layer, the superficial layer, that formed by the felting of the fibrils derived from the deep row of cells of the Corona radiata, and consequently ovarian in origin, is now much reduced in thickness, and is apparently undergoing disintegration owing to fluid infiltration throughout it.
The inner layer of the Zona pellucida does not new exhibit any evidence of concentric lamination, but, as seen in sections 7, 8, 9 a.nd 10, shows marked indications of radial striation. The change in the appearance of the Zona pellucida is so marked in this specimen, as contrasted with the earlier figure, as to suggest that some change i11 its structure is now taking place. It is worthy of note that the innermost layer of it is more dense or deeply stained, and in sections 7, 8 and 9, exhibits a clearly defined double outline. This appears to be due to no optical effect, but seems to be clearly structural. If this be so, we have to consider whether this is to be regarded as a distinct egg envelope—a vitelline membrane.
There is much confusion regarding the description given of the envelopes surrounding the egg. Bryce in Quain[3] mentions only the Zona pellucida. Robinson[4] in his diagram of the ovum and its coverings on p. 13, there indicates the vitelline membrane, as distinct from the Oolemma or Zona pellucida, but he makes no mention of a vitelline membrane in the text, and consequently we are at a loss to know exactly What his attitude is regarding this layer. McMurrich[5] makes no mention of any layer other than the Zona pellucida, whilst Jenkinson[6] (p. 45) speaks of the ovum of vertebrates as “always immediately surrounded by a vitelline membrane, frequently termed a Zona pellucida,” thereby implying that the two membranes are one and the same thing. Wilson[7] evidently regards the vitelline membrane as the equivalent’ of the Zona pellucida, Whilst Paterson[8] defines the vitelline membrane as the proper cell Wall of the ovum — “It is a clear, structureless membrane enveloping the protoplasm of the ovum, underneath the Zona pellucida.”

Fig. 5. Human ovum in discus proligerus exhibiting process of extrusion of the second polar body. x 600.
The appearance here represented would seem to confirm the separate identity of these coverings of the egg. As we see it in this series of sections, in some positions the vitelline membrane, if we reserve that name for the innermost covering of the egg as distinct from the Zona pellucida, is revealed by a double outline. In some sections its independent nature is clearly shown by its being stripped off from the inner surface of the Zona, whilst in places there seems evidence of cellular structure associated with it. It is from the inner surface of this layer that the cytoplasm retracts, forming, as we have suggested, a space occupied by fluid, and leaving on the inner surface of this membrane some small fragments of cytoplasm. This perivitelline space is of varying width, in places quite clear as if it had contained a fluid, and occupied here and there by what appear as extrusions from the cytoplasm. These points will of course be better seen in figs. 5, 6 and 7, where the structures are exhibited on a scale of magnification of 600 diameters.
The most remarkable feature exhibited within the body of the ovum is the excentric position of the nucleus or nuclear mass, as seen in section 8, of which a separate figure (fig. 5) is given.
In fig. 5, thickness of section 0.007 mm, stained with Kernschwarz, the nuclear mass abuts against theinner surface of the Zona pellucida, the vitelline membrane, if this be regarded as a separate structure, alone intervening. In this section the nucleus appears as if possessed of a distinct membrane, but this appearance is most probably an optical effect due to the difference in the refractive index of the material lying within the nucleus (the karyoplasm) and the cytoplasm. The karyoplasm, which is paler and less granular than the surrounding cytoplasm, is retracted, leaving a narrow space around it, this may be an artefact (the specimen was hardened in formol) or it may contain what has been described as the karyo-lymph or nuclear juice. Within the nuclear vesicle is a deeply stained granule measuring about 0.008 mm in diameter, this exhibits a definite structure, having within it a paler sphere, to the side of which nearest the Zona pellucida is a more highly refractive granule. The whole nuclear vesicle measures 0.024 x 0.020 mm. The cytoplasm of the egg exhibits a remarkable appearance. Around its periphery it appears darker and more compact. Centrally, however, it becomes looser in texture, and the mitochondrial grains stain deeper and stand out more clearly. Near its centre is a spherical granule possibly the centriole, and what seems like vacuoles occur Within the substance of this less compact cytoplasm. The significance of this arrangement will be better understood, if now we examine the sections before and after this example.
Fig. 6 is a photomicrograph of section 7, of a thickness of 0.007 mm, stained with Kernschwarz. The central cytoplasm here exhibits a radial arrangement of its constituent elements which appear to converge towards the nuclear mass or vesicle, this appearance is emphasised by the disposition of the granules in filamentous threads, the extremities of these threads often seem to end peripherally in one or more deeply stained grains, frequently surrounded by a vacuole-like space, or clear interval, as if they were centres of some fluid secretion.
In this figure, these collections of clearer material are also seen to invade the more compact peripheral cytoplasm. As already stated, the filamentous granules converge towards the nuclear mass, in approaching which they form a denser or more compact zone in contact with the nuclear Vesicle.
The nuclear mass consists of a paler, less granular material than the cytoplasm, within it are two deeply stained granules, whilst a third, of nearly similar size, lies on the circumference of the nuclear mass where it blends with the cytoplasm. There is no surrounding membrane, but the nuclear material is pretty clearly defined from the surrounding cytoplasm. As seen in the figure the nuclear mass is placed on one side close to the inner surface of the Zona pellucida in contact with what I take to be the Vitelline membrane; the surface of the nuclear material, so placed, appears clearly limited by a thin interrupted dark line.

Fig. 6.’’’ Human ovum in discus proligerus exhibiting appearances associated with the extrusion of the second polar body. x 600.
There are one or two other details associated with the cytoplasm which may be noted. The circumference of the cytoplasm is here and there emphasised by what are evidently flattened layers of denser material which on section show up as linear thickenings. That they are not optical in their origin is proved by their recurrence in the same situation in several sections. One of these is seen in fig. 6 a short distance above the n11clear mass. It recurs again in fig. 5 and again in fig. 7. A similar condensation of the cytoplasm on its periphery is similarly seen in these three sections on the side of the cytoplasm opposite the nuclear mass.
In fig. 6 there appear two rounded bodies lying in the perivitelline space, one of which, the left, as is proved by an inspection of the section before fig. 6 (not here represented but seen in the plate showing the whole series) is evidently a part of the cytoplasm which has become separated off from the main mass. The other, the right, on localising it by the dial of the watch placed between 12 and 1, I am not so clear about. I am inclined to think that it is a cell of the Corona radiata which has got floated out of position in the mounting of the section.

Fig. 7. Human ovum in discus proligerus showing the second polar body in process of extrusion. x600.
Fig. 7 thickness of section 0.007 mm stained with Kernschwarz is a representation of the appearance of the section‘ following fig. 5. I have placed them in this order for the sake of clearness and convenience of description. Here, the nuclear mass is seen in part pushing its way into the perivitelline space, it presents the same pale granular appearance already described, and contains in its substance two darkly stained granules, the one larger and more compact than the other. The part of the nuclear mass which is being forced beyond the periphery of the cytoplasm is clearer and paler in character, and exhibits distinct evidence of structure, there being a number of faint granules surrounded by clear spaces. Just as it passes the line of what we may regard as the circumference of the cytoplasm, this clearer mass is constricted so that the part of it pushed against the egg envelope is dumb-bell shaped in appearance, the remainder of this material, of somewhat spherical form, is bedded in the substance of the nuclear mass. The extruded part is accompanied by a small amount of cytoplasm. There can be little doubt that the appearance here observed corresponds to a stage in the extrusion of the second polar body. I say this with some confidence, for, as I shall presently show, there is evidence of the existence of the first polar body undergoing a state of division in this same oocyte.
The cytoplasm, as in the other figures, shows evidence of the same activity in the arrangement of its substance centrally, though here the radial arrangement is not so distinct, probably owing to the fact that it is cut tangentially, there does, however, appear to be some evidence that the darker granules are arranged in spherical form. About the position of 1 on the watch dial there is, near the periphery of the cytoplasm, a vacuole with a fairly large, deeply stained granule within it. The same vacuole is seen in the preceding and succeeding section; whether this has any significance, I know not, I note its presence, for now-a-days so many cytoplasmic inclusions are de scribed that possibly it is well that the presence of this particular structure 4 should be recorded.
As will be seen, the Zona pellucida in this figure has apparently been slightly damaged in the sectionising, but this is not without its advantages, for in the position of 7-8 on the watch dial there seems distinct evidence that, though there is a breach in the continuity of the Zona pellucida, its innermost layer is stripped off undamaged, a circumstance which would seem to confirm the belief in a vitelline membrane as distinct from the Zona pellucida.
From the description given — the figures must speak for themselves — there can be little doubt but that the appearances displayed correspond to the metaphase condition in the second division of the oocyte, and the phenomena associated with the extrusion of the second polar body.
The central mass within the cytoplasm corresponds to the mother centrosphere and spindle, the clearer material with its accompanying small amount of cytoplasm forced outwards against the egg envelopes is the daughter centrosphere, whilst, in between, we have the nuclear mass in process of undergoing division, though it must be confessed that the arrangement The Maturation of the Human Ovum 189 displayed by the chromatin gives little clue to any definite arrangement of the chromosomes.
I have not hesitated to assume that the conditions here described are the changes associated with the extrusion of the second polar body, for, if now we turn to look at the plate of the entire series of sections we will there be able to note the presence in sections 3 and 4 of certain structures which we have every confidence in believing is the first polar body undergoing division.

Fig. 8. Human ovum in discus proligerus, shows first polar body undergoing division in the ovum in which the second polar body is in process of being extruded. x 900.
In order to render this clearer I have introduced figs. 8 and 9 showing the details under a higher power of magnification. Before describing these, however, let me call attention to a point of some importance. Be it noted that the point of extrusion of the second polar body does not correspond to the position occupied by the first polar body. If we assume that the first polar body is polar in position (be it animal or vegetative pole) then the point of extrusion of the second polar body’ corresponds fairly closely to the equator of the egg, which fact helps to confirm the belief that the polar bodies, 190 Arthur Thomson once extruded, sink, by their own weight, in the perivitelline fluid to occupy the most dependent position within the egg envelope.
In fig. 8, which is a photomicrograph of section 4 of the entire series, 0.007 mm thick, stained with Kernschwarz, magnified 900 diameters, there will be seen lying in the perivitelline space, which is here cut very obliquely, since it lies close to one of the poles of the egg, two distinct bodies in the position of 11 on the watch dial. One of these, the more external, is oval in shape, rests on the "egg envelope, and consists of a granular material containing fairly well marked deeper stained grains; at either extremity of the oval may be seen some of the chromatin aggregated together to form little spots of more compact granules. In this section, the oval body is separate and distinct from a darker mass lying between it and the cytoplasm, which in part overlaps it. This collection of deeply stained matter exhibits denser granules Within it. If now we examine fig. 9, which is a photomicrograph of section 3 of the entire series, 0.007 mm thick, stained with Kernschwarz, the section immediately preceding that which is shown in fig. 8, we will see, in exact correspondence with the structures previously described in fig. 8, two deeply stained bodies containing chromatin granules arranged in such a way as to suggest that these are chromosomes; further the two bodies so described are seen to be connected by a bridge, which indicates their incomplete separation at this point. If these facts are admitted, and no other explanation seems possible, then I hold we have clear evidence that the structure we are dealing with is the first polar body undergoing division, coincident with the extrusion of the second polar body.

Fig. 9.’’’ Human ovum in discus proligerus showing first polar body undergoing division in the ovum in which the second polar body is in process of extrusion. x 900.
If this be so, we have in the series of sections of this oocyte a complete demonstration of the stages effected in the egg at the time of the extrusion of the second polar body. Further we have clear proof that in this oocyte, at least, the maturation stage associated with the discharge of the first and second polar bodies is completed within the ovary whilst the egg still lies within the discus proligerus of the Graafian follicle.
In Plate XI is reproduced the complete series of 16 sections (0.008 mm thick, stained with Ehrlich’s acid haematoxylin and eosin) of an oocyte within the discus proligerus, inside a. Graafian follicle, obtained from the ovary of a woman aged 38, who died of heart failure twenty-four hours after admission to hospital. It was from the same subject that the examples already described under Plate X were obtained.
What strikes one at first is the size and shape of the egg.
The largest diameters are displayed in section 9 of the series, and are estimated as 0.113 x 0.1 mm. The corresponding diameters measured inside the Zona pellucida are as follows: 0.1 x 0.084 mm.
The egg and its envelopes 11111 through 14 out of the 16 sections figured. As each section was 0.008 mm thick, the total “thickness ” of the egg sectionised amounts to 0.112 m1n., so that the oocyte has evidently been cut transverse to its long axis, the two extremities of the egg as seen in the serial sections will correspond therefore to the poles, if these are determined by its longest diameter, a point of some importance in regard to the disposition of the polar bodies, and one concerning which Jenkinson[9] makes the definite statement that “ In cases where the egg is ovoid or ellipsoid (Myxinoids, Amia) the egg axis is the major axis.” The diameters given for this oocyte, which is in a very perfect state of preservation, preclude the possibility of it being described as spherical. A matter worth drawing attention to is that this oocyte exceeds by an appreciable amount the diameters given for those already described.
The cells of the discus proligerus or cumulus, which immediately surrounds the egg to form the Corona radiata, are closely packed, only at one point is there a breach in their continuity shown in sections 6, 7, 8 and 9. Here the liquor folliculi, shown in the photomicrographs as a pale granularlooking coagulum, is free to enter, and has evidently bathed the external 1 Vertebrate Embryology, Oxford, 1913, p. 40. 192 ~ . Arthur Thomson surface of the Zona pellucida, though as yet in this specimen it does not appear to have extended much by infiltration between the outer surface of the Zona pellucida and the deeper row of cells of the Corona radiata. In consequence, the fibrillar substance, cementing the innermost cells of the Corona radiata to the outer surface of the Zona pellucida, and forming by its “felting”. the outer or ovarian layer of that membrane, is more compact, and not so broken up by fluid infiltration as that described in the previous examples.
The Zona pellucida, of an average thickness of from 0.O06 to O.OO8. mm is very well seen in many of the sections, particularly in Nos. 7 and 8, where its separation into an inner and an outer layer is very clearly seen, the outer being obviously of a different character from the inner and being formed by the felting of the basal fibres of the innermost cells of the Corona radiata and therefore ovarian in derivation, whilst the inner layer, clearer and more homo ‘ geneous, and derived, as I have reason to believe from a study of the growth period of the oogonium, from the ovum itself (I hope to publish this series of observations later), is limited internally by a well defined line, which on careful examination is seen to be double. This, I take it, is confirmatory of the View that internal to the Zona pellucida, and in contact with it, there is another egg membrane, the true vitelline membrane of which I have already spoken. The ovular or inner layer of the Zona pellucida proper exhibits appearances here and there which suggest concentric lamination; in other situations, in some’ of the sections, there is an appearance of radial striation, but I have not been able to observe any of the fibrillae derived from the inner cells of the Corona radiata traversing the substance of the Zona pellucida. In one or two instances, notably in section 7, there are evidences of cellular formation or cell inclusions within the substance of the inner or ovular layer of the Zona pellucida.
As has been already suggested, there appears to be evidence of a layer within the Zona and adherent to it, which is possibly a true vitelline membrane, but in these sections it must be admitted that the evidence is not very convincing, for it is extremely difficult to make out whether the appearance is due to an optical effect or no.
The cytoplasm in all the sections exhibits an evenly granular appearance, increasing in density in the centre and around the nucleus. This central darker mass is separated, here and there, from the surrounding lighter cytoplasm by a more open grain in the granular material, amounting in places to definite irregular clefts, whether this appearance is due to the influence of the hardening agent (formol) or is a natural condition induced by excess of fluid is a moot point.
In most of the sections the cytoplasm appears to fill completely the egg envelope, but if the middle sections of the series, viz. those near the equator of the egg, be carefully examined there is definite evidence of a narrow interspace between what Waldeyer has called the ooplasm cortex, i.e. a narrow The Maturation of the Human Ovum 193 zone of finely granular material surrounding the cytoplasm, and the inner side of the egg envelope. This space, the perivitelline space, for such it would appear to be, is probably filled with a fluid extrusion from the cytoplasm, for in it appear two or three bodies which will now be described.
In sections 13 and 14 of the series, close to the pole at that end of the egg, are seen to be certain conspicuous features. In section 14, here shown in fig. 10, there are two distinct nuclear granules surrounded by an irregular mass of slightly darker material than the adjacent cytoplasm. This substance is also faintly indicated in the previous section (not here figured).
{kind=link}
Fig. 10. Section of a. mature human ovum in the discus proligerus showing near one of its poles the now divided first polar body. x 600.
From a careful study of this object I think there can be little doubt but that we have the first polar body in a divided condition. It will be noticed that so far as its site is concerned it is distinctly polar in position, that is to say it lies at one extremity of the greatest diameter of the egg.
Having thus accounted for the first polar body it remains to determine the existence of the second polar body. In doing this considerable difficulty was experienced, as there were sundry objects which might be taken for it, Anatomy LIII 13 194 Arthur Thomson but after careful examination I am inclined to believe that the structure exhibited in section 7 of the series, opposite 11 of" the watch dia.l, is the object we arein search of——the second polar body (fig. 11). Here is seen a darkly stained granule of chromatin which on high magnification resolves itself into more or less distinct groups of chromatin particles apparently connected together, the whole being surrounded an envelope of a substance identical with that of the adjacent cytoplasm. If this be the second polar body, it should be noticed that its position does not correspond to that of the first polar body. As to whether it may ultimately sink to the same position as that occupied by the first polar body is a matter of surmise, evidently, like that described in the last figure, it is extruded from the cytoplasm at a very different level from that now occupied by the first polar body.
{kind=link}
Fig. 11. Section of a. mature human ovum in the discus proligerus showing in the direction of 11 of the Watch dial the second polar body within the zona. pellucida. x 600.
All the other deeply stained bodies, which one might have associated with a polar body, have been eliminated, for the reason that they are evidently Washed granules in the process of mounting. There still remains the fact, however, that, here and there, along the inner surface of the envelope covering the left hemisphere of the egg, there are bulla-like extrusions from the substance of the cytoplasm, forcing their way into the Zona pellucida across the perivitelline space. The contents of these blister-like bodies seems less solid and granular than the material of the cytoplasm.
That this appearance is the result of a vital act is suggested by the evident approach of vacuoles towards the surface of the cytoplasm seen at 10 and 11 of the dial face in section 8, where one of them contains a deeply stained granule. It would seem as if these vacuoles were on their way to reach the perivitelline space, there to discharge their contents.
{kind=link}
Fig. 12.’’’ Section through a. mature human ovum in the discus proljgerus: the polar bodies of this ovum have already been shown in figs. 10 and 11. x 600.
The nucleus, 0.03 x 0.0244 mm in size, has returned to a nearly central position in the egg, it is surrounded by a faint and delicate membrane, somewhat unduly emphasised in the photograph shown in fig. 12. A These facts, taken in conjunction with the undoubted presence of the first polar body, afford convincing proof that the oocyte has passed to the resting stage of the mature egg, and that already the phenomena associated with the extrusion of the second polar body have been passed, since we are led to believe, on the evidence of those familiar with the changes occurring in other mammalian ova, that there is no resting stage between the first and second maturation divisions, hence the necessity for a search for the second polar body. If the present assumption be true, all these facts seem to harmonise and amply justify us in establishing the identity of the second polar body already referred to. Herein we have therefore to deal with a mature oocyte provided with what is now the female pronucleus. The nucleus, so described, as it appears in the figure (fig. 12) has a deeply stained spherical nucleolus 0.007 mm in diameter, lodging a more refractive granule within it. The nucleus contains, besides, in the finely granular karyoplasm a number (about 15 in this figure) of small chromatin granules mainly disposed around the nucleolus, and lying, too, along the inner side of the nuclear membrane.
The karyoplasm is traversed by irregular clefts, whether artefacts or vital features it is impossible to determine.
In the preceding and succeeding sections in which the nucleus is cut and appears, the nuclear membrane in the photographs is faint but distinctly traceable.
In the section just before fig. 12, the nucleolus is also represented by a thin slice, and scattered through the karyoplasm of the three sections are chromatin granules to the approximate total number of 45 to 50. Some of these appear to be isolated grains, some linked together in threads, and others arranged in groups.
Around the nucleus, externally, and lying either near, or in close contact With, its membrane, are sundry vacuole-like bodies. Three of these," of varying size, each with a faint granule in it, lie opposite to the upper right quadrant of the nucleus. Another, not so clear, lies in the direction of 5 of the dial face.
In section 9 of the Series Plate XI (shown in fig. 13) a larger vesicle, 0.O12 x 0.008 mm in size, lies in the direction of 3 of the Watch dial, in contact with the nuclear membrane. Its contents are clearer than the surrounding cytoplasm, and more finely granular, possibly this may be the remains of a centrosphere. What the significance of the other structures may be, I know not, but their constant recurrence in other ova which I have studied suggest that they are normal features associated in some way with the nutrition or activity of the egg.
There can be little doubt, I think, that the oocyte above described has reached its mature condition. All the facts seem to justify this assumption.
The presence of a divided first polar body, the withdrawal of the nucleus to a central position, the reappearance of the nuclear membrane and the nucleolus, the probable identity of the second polar body, and the disappearance of all activity on the part of the centrospheres, all point to the correctness of this conclusion. It may further be added that judging by the thinness of the overlying ovarian layer, this oocyte was found in a Graafian follicle at a stage which must have been close to the period of rupture.
Plate XII represents a complete series of 12 sections (0.008 mm thick, stained with Weigert’s Iron Haematoxylin and Van Giesen) of an oocyte in the discus proligerus, inside a Graafian follicle, from the ovary of a woman aged 38 who died of heart failure one day after admission to hospital. This is the same subject as that from which the specimen figured in Plate XI was obtained.
The largest diameters, 0.111 x 0.09 mm, are seen in section 5 of the series. Measured from Within the inner surface of the Zona pellucida the corresponding diameters are 0.10 x 0.08 mm.
{kind=link}
Fig. 13. Section through a mature human ovum in the discus proligerus, the vesicle to the right of the nucleus may possibly be the remains of a centrosphere. x 600.
The egg and its envelopes run through 10 of the 12 sections,‘and as each section was cut to a thickness of 0.008 mm, the total diameter of the egg as estimated by taking the total thickness of the 10 sections amounts to 0.080 mm, so that the inference is that the sections displayed in the plate have been cut in a plane parallel to the long axis of the egg.
In this instance, also, the egg, which is in a good state of preservation, can in no sense be described as typically spherical.
This specimen is much of a size with that described in connexion with Plate XI.
The cells of the Corona radiata towards the middle of the series are seen to consist of a single layer augmented here and there by cells forming a second row, but it is evident from an inspection of the figures that this combined layer is separated from the surrounding loosely arranged cells of the discus proligerus by intervals of varying width occupied doubtless by liquor folliculi. It is interesting to note that the coagulum formed by the liquor folliculi seen towards the edges of the lower sections is of an open fibrillar nature as contrasted with that seen in the last series of sections (Plate XI). The significance of this difference is difficult to determine, for both specimens were fixed with the same reagent (formol); presumably it is due to some change in the constitution of the fluid. There are occasional breaches in the continuity of the inner row of coronal cells, so that ample channels are available for the free permeation of the fluid between the inner layer of cells of the Corona radiata and the outer surface of the Zona pellucida. In consequence we see that the fibrillar matter connecting the cells of the Corona radiata with the outer surface of the Zona pellucida is much broken up, the bundles of fibrils being separated by interspaces of varying width, thus providing free access for the liquor folliculi. In consequence only a feeble connexion is established between the Zona pellucida and the cells of the Corona radiata — so also the outermost layer of the Zona pellucida formed by the felting of the fibres derived from the coronal cells is considerably reduced in thickness.
The Zona pellucida, of an average thickness of from 0.004 to 0.005 mm in the sections where it is cut transverse to its surface, is mainly composed of the inner homogeneous or ovular layer, the outer or ovarian layer as just described is relatively thin. The Zona pellucida has been variously described as a firm, tough, and elastic membrane. I confess that in my observations on the human ovum, limited it is true to hardened specimens alone, this account ill accords with what one sees, for the frequency with which various objects in contact with it indent its surface, rather belies this description; it would seem to me to be of a more plastic nature than is usually described. It may of course be that the erosions in its substance are caused by the vital activity of the material in contact with it. In the sections which are meridional there is evidence of a clearer layer in contact with the inner surface of the Zona pellucida as indicated by quite definite outlines and paler tint. This, if our surmise be correct, is the vitelline membrane proper, though in this specimen there is no indication of its independent nature by being separated or torn away from the Zona pellucida. In the sections close to the meridian the inner layer of the Zona pellucida occasionally presents faint indications of a concentric laminated«?arrangement, it is only in the region of the lower pole of sections 5 and 6 that a faint radial striation is reproduced in the photographs. The cytoplasm is retracted from the vitelline membrane, to which some few particles of its substance still adhere, to form a perivitelline space, well seen in many of the sections.
In sections 4, 5 and 6, in the position of 1 of the watch dial a considerable mass of the cytoplasm, clearly fragmented, has been pulled away from the general substance of the cytoplasm as if here its attachment to the vitelline membrane had been more intimate, a circumstance which suggests that the cavity is due rather to an artefact than to a Vital process. The reader may be reminded, however, that in previous examples (figs. 2, 3, 4) we have seen this space occupied by a coagulum, a condition which presupposes its existence during life.
{kind=link}
Fig. 14. Section through a mature human ovum in the discus proligerus. x 600.
The cytoplasm displays an evenly granular appearance, gradually increasing in density and tint towards the central area of the cell and around the nucleus. The darker zone, however, is not of uniform appearance, but has a mottled look due to the alternation of lighter and darker material. It is noticeable too that in sections 5 and 6 the darker material seems to radiate out into the peripheral cytoplasm by pointed strands. Here and there throughout the cytoplasm in general may be seen a few more deeply stained granules. Towards the circumference in section 6 there is a suggestion of the granular material being arranged in radial threads.
{kind=link}
Fig. 15. Section through a mature human ovum in the discus proligerus showing vesicular ‘appearance within the nucleus. x 600.
The nucleus, O.028 x 0.O22 mm, best seen in section 5 (fig. 14), is there shown all but central in position. It by no means follows, however, that it is central in the egg, for whilst there are three sections antecedent to it, there are six sections following it. In section 5 it exhibits a distinct membrane, possibly emphasised in the. process of reproduction by photomicrography, but undoubtedly revealed on microscopic examination as a distinct clearly defined covering. In the serial sections on either side of section 5 the nucleus is again cut, but there, in the photographs, there is but slight indication of a nuclear membrane. This does not necessarily imply that the membrane is not there, for experience has taught me that so delicate a structure is very difficult to define, particularly when it is cut obliquely. Careful examination, however, in both these sections reveals a faint indication of its presence. Within its membrane, as shown in section 5 (fig. 14), the karyoplasm of the nucleus has shrunk from its envelope, leaving a clear space presumably occupied by karyolymph. The mass of the contained karyoplasm is faintly granular and contains numerous chromatin granules, some single, some in pairs, and some arranged in filaments; whilst one -mass, thicker and of elongated form, near the centre of the nucleus displayed in section 5 may be described as a chromosome. Whilst many of these chromatin particles appear in the substance of the karyoplasm, a number are shown closely applied to the inner surface of the nuclear membrane.
{kind=link}
Fig. 16. Section through a. mature human ovum in the discus proligerus. Note the occurrence of the clear vesicle containing darkly stained granules to the left of the nucleus. x 600.
In the three sections dividing the nucleus seven filamentous threads, seven paired granules, and ten isolated grains may be noted besides the chromosome referred to, and in addition a more or less compact mass of granules shown at the lower end of the nucleus in section 6.
In connexion with some of these granules there appear little vesicles which would seem to be filled with the products of their activity, of these three or four may be seen in section 4 and one in section 5 (section 4 is shown in fig. 15).
In section 6, reproduced in fig. 16, to all appearance outside the nucleus, but in contact with it, is a remarkably clear Vesicle 0.01 x 0.006 mm, lying in the direction of 9 of the watch dial ; within it peripherally and also directed to 9 of the dial is a single chromatin granule, to the 5 dial side of which is a curved wreath of /\-shaped chromatin particles. What this is is matter for speculation. It will be discussed later.
Near the nucleus in section 6, in the direction of 2 of the dial there is a group of small pale vesicles, three in number, embedded in the substance of the central cytoplasm; each exhibits an appearance of structure within, and each is fairly clearly defined from the surrounding structure.
{kind=link}
Fig. 17. Section through a mature human ovum, showing, in the direction of 12 of the watch dial, the first polar bodies. x 600. .
The recurrence, at the pole of the egg (see section 10 of the series, reproduced in fig. 17), of a feature closely resembling that which in corresponding position in Plate XI we have identified as the first polar body, suggests a similar explanation of the appearance here seen. In the section lying between the cytoplasm and the egg envelope in the direction of 12 of the dial is an object composed of granular substance and more deeply stained material evidently composed of chromatin particles. If now section 9 be examined in the corresponding position two (or it would almost appear three) fairly clearly defined oval-shaped bodies are seen. The appearance exhibited in these two sections, though not so definite and typical as those displayed in the figures already given (fig. 10), is yet sufficient to justify us in assuming that the appearance here described is none other than the first polar body in a divided condition. If this be not admitted we are forced, in face of the confirmatory evidence, to supply a better explanation of its presence, that I confess I cannot do, since it is obviously not due to any defect in the cutting or mounting, we are left with the alternative of accepting the only feasible explanation, viz. that it is the first polar body.
Accepting this as a fact, then we have to take in connexion with it the obvious detail that the nucleus is more or less central, that there is a nuclear membrane, and that the centrospheres have disappeared, in other words that the nucleus is now returning to the resting stage. If so, we are in duty bound to look for the second polar body. At first sight one was inclined to assume that the fragmented mass of cytoplasm already referred to in sections 4, 5 and 6, as occupying the perivitelline space was the body in question, but careful microscopic examination, emphasised by the microphotographs themselves, puts it beyond a doubt that this appearance is to be explained as an artefact. The other alternative presented is shown in section 8 (fig. 18), where apparently lying on the surface of the Zona pellucida there is a collection of granules in the direction of 8 of the watch dial. It must be remembered that here the egg envelope is being cut obliquely, so that the appearance of the object here represented as more or less embedded in the substance of the Zona pellucida is misleading. It rests rather on the inner sloping surface of the egg envelope, and lies appropriately in what is the perivitelline space, no longer visible owing to the oblique manner in which the structures are cut. Again, though not so typical as that previously described in fig. 11, it would appear, for lack of a better explanation, that the object referred to is none other than the second polar body. In point of structure it appears as four fairly large spherical granules, two darkly stained and two clearer, the latter lying between and partly overlapped by the former. The position occupied by this body is separated by one clear section 0.008 mm thick from the plane of section of that containing the first polar body. Further, be it noticed, that it lies almost at the opposite side of ‘the egg, occupying a position corresponding to 8 of the dial, whilst the first polar body lies in the direction of 12 of the dial. There is no evidence to show the point where the second polar body was extruded from the cytoplasm, it only appears, that at the present stage, its position does not coincide with that of the first polar body, as we have been led so frequently to believe.
If we accept the above description as satisfactory, in so far as it establishes the identity of the first and second polar bodies, then we have to explain the occurrence of the remarkable body seen to the 9 dial side of the nucleus in 204 ' Arthur Thomson section 6 (fig. 15) to which attention has been already directed (p. 52). The chromosome-like characters of the contained chromatin, and their arrangement in wreath-like fashion in the equatorial zone of the vesicle, appear to suggest that we are dealing with some nuclear structure in an active division phase, but what it may be, if this be granted, is a mystery, for there is evidence that it lies outside the membrane of the nucleus of the egg, and there is no appearance of any centrosphere activity around it. For the present I must leave it at that, frankly admitting that in the present state of my knowledge of the subject its, presence is inexplicable. It may be a coincidence, but the fact is worth recording that in section 9 of series Plate XI, there is in the direction of 3 of the dial a vesicle of much the same size lying in contact with the nuclear membrane. In that case, however, there are no chromatin contents. Is it possible that these two structures are similar and represent a phase in the disappearance of a centrosphere? From the facts observed, this oocyte, like the last one described, is most probably in the mature condition. Its nucleus is now the female pronucleus, and since the identity of the first polar‘ body has been fairly convincingly established We are justified in assuming that taking into account the more or less central position of the nucleus, and the fact that it has now resumed its nuclear membrane, the egg has passed beyond the phase in which the second polar body was extruded and is now in the resting condition of the mature oocyte. This is an additional reason for assuming that the object identified as the second polar body is in fact that structure.
{kind=link}
Fig. 18. Section through mature human ovum in the discus proligerus, showing in the direction of 8 of the dial, what is regarded as the second polar body. x 600.
On carefully tracing the Graafian follicle, in Which the ovum was contained, its Wall was found of a thinness which would suggest that its early rupture could not be long delayed.
Resumé
Reviewing the facts and their interpretation as expressed in the previous pages, we may summarise the results as follows:
1. The human egg is not a sphere but an ovoid structure.
2. Size of the Human Oocyte. The diameters of the various examples described are here tabulated.
| Oocyte | Including Zona pellucida (mm) |
|---|---|
| Fig. 1 | 0.1 x 0.09 |
| Fig. 2 | 0.117 x 0.1 |
| Fig. 3 | 0.097 x 0.88 thickness. |
| Pl. I | 0.111 x 0.1 x 0.091 |
| Pl. II | 0.113 x 0.1 x 0.112 |
| Pl. III | 0.111 x 0.09 x 0.08 |
The variation in size is very slight, and apparently there is no evidence to indicate that a change in size occurs during the maturation period. My measurements are considerably less than those usually quoted.
3. The character of the Zona pellucida appears to undergo a process of thinning, by the dissolution of its external ovarian layer, by means of which it is intimately connected with the inner cells of the Corona radiata. The reduction in the thickness of this layer is evidently effected by the permeation of the liquor folliculi through its substance, thereby leading to a separation of its elements into strands or small groups of fibrils which at the period of expulsion of the oocyte may be easily ruptured.
4. There seem grounds for believing that lining the inner surface of the Zona pellucida there is a structurally distinct layer, the vitelline membrane proper. The evidence in support of this view is that in a case (fig. 7') in which the Zona pellucida was torn across, this fine inner layer remained intact, further the occurrence of a faint double outline exhibiting a paler structure Within seems to point to the independent nature of this membrane.
5. The Zona pellucida, whilst generally speaking homogeneous in character, yet exhibits in some instances a faint appearance of concentric lamination. From the observations on the specimens here recorded it would seem as if the radial striation only occurred in the later stages of the maturation of the oocyte, but I hesitate, on such slender evidence, to lay stress on this. Within the substance of the zona there appears from time to time the suggestion of fine cellular elements. I have not yet been able to trace the fibrils from the cells of the Corona radiata through the substance of the Zona pellucida as some have described. The thickness of the Zona pellucida ranges from 0.003 to 0.011 mm, yielding as an average a little over 0.006 mm.
6. The perivitelline space. The existence of this space has been a matter of much controversy. The strongest evidence I can produce in its favour is the presence of a distinct coagulum within its cavity (see figs. 2, 3, 4, and pp. 17 7 , 179, 181). Its greatest development appears to coincide with the extrusion of ' the polar bodies which eventually come to lie within it. After the expulsion of the polar bodies the cytoplasm appears to fill the egg envelope more completely. 7. In the cytoplasm the appearance of a single centrosphere has been recorded. This is again shown divided and placed on either side of the nucleus in fig. 2, the two parts resulting from this division are again represented in fig. 3 as well defined spherical masses. In a later stage the central part of the cytoplasm is evidently affected by the centrosphere (fig. 4). Later, after the extrusion of the second polar body, the centrosphere seems to be associated with the regrouping of the denser central cytoplasm. finally, the clear vesicle lying in each instance against the membrane of the nucleus in the last two plates has been provisionally interpreted as the centrosphere (of which there is now only one, the other having been expelled with the second polar body), undergoing a state of resorption.
8. Other cytoplasmic inclusions. These include vacuoles of various sizes often exhibiting a darkly stained granule within, and evidently containing some fluid secretion. There is evidence to suggest that some of these are making their way through the cytoplasm towards the perivitelline space, there to discharge‘ their contents. Darkly stained isolated granules are occasionally met with.
9. The nucleus ranges in size from 0.020 X 0.017 to 0.030 x 0.024 mm. It has been seen quiescent, more or less central, and surrounded by a cell membrane, in Plates XI and XII (figs. 12, 13, 14). It has been observed in an excentric position and apparently devoid of a membrane (figs. 5, 6), it has also been noted more or less central and membraneless (fig. 2). These different positions and conditions are associated with various of the stages connected with the formation of the polar bodies. The nuclear contents have been seen to include chromatin granules, isolated, paired, massed in groups, or arranged in filamentous threads. In one instance the chromatin appears in the form of a definite, thick, rod-shaped chromosome (fig. 14).
10. The nucleolus. A clearly defined nucleolus, displaying evidence of internal structure, is exhibited in fig. 12, which represents the condition of the mature oocyte in which the nucleus displays"‘the characters associated with a resting stage. In fig. 5, representing the egg at a stage corresponding to that of the extrusion of the second polar body, there is in the nuclear mass a rounded, deeply stained body, which resembles in its mass and appearance a nucleolus; What the significance of this structure is, is doubtful. Within the nuclear mass of the same oocyte there are two or three deeply stained granules which from their size might be dignified by the name of nucleoli.
11. The karyoplasm (nuclear protoplasm) has retracted from the surrounding cytoplasm in figs. 1 and 5 and from the nuclear membrane in figs. 11 and 14, so as to leave a space. Whether this be due to an artefact or is a vital process is a moot point——if the latter, its contents are probably of the nature of karyolymph or nuclear juice.
In all instances, the karyoplasm is paler and more finely granular than the surrounding cytoplasm.
In fig. 15 the karyoplasm seems to be occupied by four little clearer vesicles, each with darker stained granules around its circumference.
The Polar Bodies
The first Polar Body
What is assumed to be the first polar body immediately after its expulsion into the perivitelline space, is shown in fig. 3. What, there is justifiable reason for believing, is the first polar body undergoing division (coincident with the extrusion of the second, polar body) is seen in figs. 8 and 9. In figs. 10 and 17 the first polar body, now divided, so that there are two, is shown in conjunction with the second polar body, now extruded into the perivitelline space, though not necessarily occupying the same position.
The Second Polar Body
The second polar body, in process of expulsion from the nuclear mass, is shown in fig. 7. Here what is assumed to be the daughter centrosphere connected with it, recognised as the dumb-bell shaped small clearer vesicular mass abutting against the inner wall of the egg envelope (see p. 187), is also evident. The second polar body has been identified, in what are assumed to be mature oocytes owing to the nuclei having resumed a more or less central position and each having associated with it a nuclear membrane, in figs. 11 and 18, in each of which oocytes the divided first polar bodies have each been located as seen in figs. 10 and 17.
If these observations be admitted, there is distinct evidence that in man the expulsion of both polar bodies takes place before the oocyte has left the Graafian follicle, in other words, that the maturation of the oocyte with its female pronucleus is complete before it has been subjected to the influence of the spermatozoon. This is contrary to the general rule as applicable to Vertebrates, according to which, as stated by Jenkinson, “It is a rule for the sperm to enter during the second maturation division of the ovum.”[10] But it may possibly be explained by the fact that in the human female congress with the male is not limited, as it is in most other mammals, to the period of “heat.” In the human female, ovulation appears to take place at any stage of the menstrual cycle, whereas in lower animals as a rule it occurs in association with the condition known as oestrus or “ heat.” We are, therefore, justified in assuming that in our own species, the greater licence conferred upon us as to sexual union, has led to a modification of the processes involved in the maturation of the egg. An observation worthy of note in this connection is that of Heape[11], who draws attention to the fact that “ in spite of the regular recurrence of the oestrus, monkeys have only a limited season during which conception is possible. . . .this is due to the fact that the ovary is not active all the year round,” which of course under the existing conditions of higher civilisation is not applicable to the human female.
A further point regarding the position of the polar bodies in relation to the axis of the egg is interesting. The evidence in the present series of observations seems to point to the fact that the point of extrusion of the first polar body through the cytoplasm need not necessarily bear any relation to the “polar” position which it subsequently assumes, but that that may be determined by the influence of gravity as it lies within the fluid occupying the perivitelline space (see p. 179). Nor is there any evidence to prove that the point of extrusion of the second polar body from the cytoplasm corresponds to that of the exit therefrom of the first polar body — the evidence, as has been pointed out (pp. 189, 203), enables us to decide that at the time of its extrusion the second polar body may occupy a position very different from that in which we see the first polar body, or its divisions, though it must be clearly borne in mind that this does not exclude the possibility of the two polar bodies having been extruded from the same site, since meanwhile, in the interval between the two extrusions, the first polar body may have altered its position by sinking to the most dependent part of the egg envelope within the perivitelline space.
The prophase condition in the formation of the first polar body is seen in figs. 1 and 2.
The anaphase condition of the formation of the first polar body is seen in fig. 3.
The metaphase condition of the formation of the second polar body is represented in Plate X and fig. 7.
Whilst the end of the telophase condition of the formation of the second - polar body is exemplified in Plates XI and XII and figs. 11 and 12, and 14 and 16.
References
- ↑ Vertebrate Embryology, Oxford, 1913, p. 72.
- ↑ The Cell in Development and Inheritance, New York, 1900, p. 156.
- ↑ 3.0 3.1 Sharpey W. Thomson A. and Schafer E.A. Quain's Elements of Anatomy. (1878) William Wood and Co., New York. Quain’s Anatomy, vol. I. “Embryology," T. H. Bryce, London.
- ↑ Cunningham, Textbook of Anatomy 1913.
- ↑ McMurrich The Development of the Human Body, 5th Edition, Philadelphia.
- ↑ Jenkinson Vertebrate Embryology, Oxford, 1913.
- ↑ Wilson The Cell in Development and Inheritance, New York, 1900.
- ↑ Paterson Manual of Embryology.
- ↑ Vertebrate Embryology, Oxford, 1913, p.40.
- ↑ Jenkinson Vertebrate Embryology, Oxford, 1913, p. 86.
- ↑ Heape “The ‘Sexual Season’ of Mammals.” Quarterly/Journal of Microscopical Science, New Series xLIv. 1900.1, p. 58.
Description of Plates
Plate X

Reproduction of complete series of sections of a human ovum in the discus proligerus (No. 453, Series A) exhibiting the extrusion of the second polar body, indicated by an arrow to the left side of the first section in the third row. The arrow in the fourth section of the first row points to the first polar body. x 150 (reduced from the photomicrographs taken with a magnification of 600 diameters).
Plate XI

Reproduction of a complete series of sections of a mature human ovum in the discus proligerus (No. 453, Series A). The arrows in the plate point to the polar bodies: the lower to the now divided first polar body, the upper to the second polar body.
Plate XII

Reproduction of a complete series of sections through a mature human ovum. The lower arrow points to the divided first polar body, the upper arrow indicates the position of the second polar body.
Magnification 150 diameters.
Figures
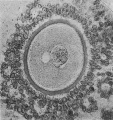
Fig. 1. Human ovum in discus proligerus in early part of prophase of first division
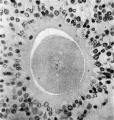
Fig. 2.
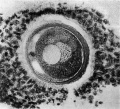
Fig. 3.
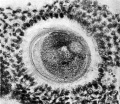
Fig. 4.
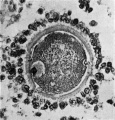
Fig. 5.
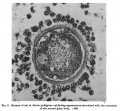
Fig. 6.
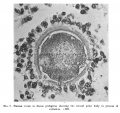
Fig. 7.
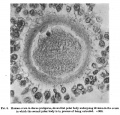
Fig. 8.
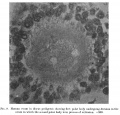
Fig. 9.
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Cite this page: Hill, M.A. (2026, April 18) Embryology Paper - The Maturation of the Human Ovum. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_The_Maturation_of_the_Human_Ovum
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G