Talk:Endocrine System Development
| About Discussion Pages |
|---|

Glossary Links
Cite this page: Hill, M.A. (2024, June 18) Embryology Endocrine System Development. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Talk:Endocrine_System_Development |
Human Endocrine - Embryonic
Based on data from O'Rahilly R. The timing and sequence of events in the development of the human endocrine system during the embryonic period proper. (1983) Anat. Embryol., 166: 439-451. PMID 6869855 [1]
By Stage
Pineal | Hypothalamus | Pituitary | Thyroid | Parathyroid | Thymus | Pancreas | Adrenal | Gonad |
Stage 13
- Hypophysis - basement membranes of the craniopharyngeal pouch and the brain are clearly in contact (O'Rahilly 1973).
- Thymus - Weller (1933) recognized already a thymic primordium "of considerable size" on the ventral part of the third pharyngeal pouch, whereas Norris (1938) considered this stage to be "preprimordial"
- Thyroid - median thyroid is now bilobed and is connected to the pharynx by a hollow pedicle (Weller 1933). The telopharyngeal body has been regarded as a "lateral thyroid component" by some workers (e.g. Weller 1933).
- Pancreas - ventral pancreas may perhaps be distinguishable (Politzer 1952).
Stage 14
- Hypophysis - craniopharyngeal pouch is prominent (Streeter 1945) and the notochord appears to be inserted into its dorsal wall. The craniopharyngeal pouch has become elongated and blood vessels are beginning to grow in between the basement membranes of the pouch and brain (O'Rahilly 1973a).
- Epiphysis - a slight irregularity in the surface outline of the intact head corresponds to the future pineal body (O'Rahilly et al. 1982). Thymus. Weller's (1933) "thymus" (the third pharyngeal pouch) becomes elongated.
- Parathyroids - "Parathyrogenic zones" (Politzer and Hann 1935) are recognizable (Streeter 1945). The parathyroid 4 primordium has been illustrated at this stage by Weller (1933, Fig. 16).
- Thyroid - thyroid pedicle shows further elongation but is still connected to the epithelium of the pharynx (Weller 1933). Right and left lobes and an isthmus may perhaps be presaged (ibid.).
- Adrenal Cortex - A change in the characteristics of the cells of the coelomic epithelium appears between the mesogastrium and the lateral end of the mesonephros.[2]
- Adrenal Medulla - paravertebral sympathetic ganglia increase in size as a result of cell division and the addition of nerve fibres from the rami communicantes. The ganglia contain three types of cells: MI, M2, and M3. The M3 cells are the" parasympathetic cells" of Zuckerkandl.[2]
- Pancreas - ventral pancreas (which may perhaps be distinguishable as early as stage 13) appears as an evagination from the bile duct at stages 14 (Blechschmidt 1973) and 15 (Streeter 1948). It is generally described as unpaired but, at least in some cases, may perhaps be bilobed[3] or even multiple (Delmas 1939).
Stage 15
- Epiphysis - pineal body is detectable in the roof of the diencephalon (Stadium I of Turkewitsch 1933) (O'Rahilly 1968).
- Thyroid - thyroid primordium may be detached from the pharyngeal epithelium in some instances. "At about the time" when the thyroglossal duct "becomes broken it loses its lumen" (Grosser 1912).
- Adrenal Cortex - primordium is first recognizable. A new type of cell (C1) from the coelomic epithelium is found in the subjacent mesenchyme. New cells (C2) appear in the medial wall of mesonephric glomeruli and begin to migrate into the suprarenal primordium.[2] Jirfisek (1980) denies a mesonephric contribution to the suprarenal.
- Adrenal Medulla - all types of cells (M1, M2, and M3) increase in number. From stage 15 to stage 18, the suprarenal primordium is cigar-shaped and extends from segment T6 to segment L1, lateral to the aorta and mesogastrium.[2]
Stage 16
- Hypophysis - slight indication of the infundibular recess may be seen in some embryos (O'Rahilly 1973 a).
- Epiphysis - cellular migration in an external direction occurs in the pineal body during stages 16 and 17 (Stadium 2 of Turkewitsch 1933) (O'Rahilly 1968).
- Thymus - according to Norris (1938), "not until the primordium of the parathyroid [3] has been outlined can the remaining portion of the third pouch be recognized, by exclusion, as the primordium of the endodermal thymus".
- Parathyroids - parathyrogenic zones are closely related to the third and fourth aortic arches at 9 mm (Politzer and Hann 1935, unstaged embryo). Parathyroid 3 is identifiable on the anterior wall of the third pharyngeal pouch (Weller 1933, Fig. 17) and "does not arise from a dorsal lobule" of the pouch (Norris 1937). The "sudden appearance of well-differentiated clear chief cells in the early primordia of the parathyroids" at 9 mm was emphasized by Norris (1937).
- Thyroid - has lost its continuity with the pharynx and it consists of two lobes, an isthmus, and a remnant of the pedicle (Weller 1933).
- Adrenal Cortex. Another type of cell (C3) arises from the coelomic epithe- lium. Both C1 and C3 cells enter the suprarenal primordium. An "enormous immigration" of C2 cells occurs.[2]
- Adrenal Medulla - cells of neural origin are migrating into the gland, separating the cortical cells into islands. Nerve fibres from the ganglia ac- company the M1 and M3 cells. The M2 cells remain in the ganglia and become sympathetic ganglion cells.[2]
- Pancreas - dorsal pancreas and the ventral pancreas are contiguous.[4]
Stage 17
- Hypophysis - juxtacerebral wall of the craniopharyngeal pouch is the thicker. The lateral lobes (future infundibular, or tuberal, part) and the anterior chamber (Vorraum) are clearly visible (O'Rahilly 1973 a). The infundibular recess displays a characteristically folded wall, namely the neurohypophysis (O'Rahilly 1973 a).
- Thymus - connection of the thymus with the pharynx has been severed (Weller 1933). The thymus is intimately approximated to the cervical duct (ibid.) According to Norris (1937), both third and fourth pouches make contact with the ectoderm, although only the third "receives an increment from the ectoderm".
- Parathyroids -parathyroid 4 is attached to the lateral surface of what Weller (1933) termed the "lateral thyroid component"
- Thyroid. The lobes of the thyroid curve around the carotid arteries and are connected by a delicate isthmus. Lacunae "should not be confused with lumina of follicles" (Weller 1933).
- Adrenal Cortex - dorsal part of the whole suprarenal primordium is disorganized by the invasion of sympathetic nerves and cells, while the band of C2 cells and the coelomic epithelium remain intact (Crowder 1957).
- Adrenal Medulla - first neural migration is at its height. Growth of the para-aortic complex is extensive. The plexiform complex is derived from paravertebral sympathetic ganglia T6-12 and usually L 1. Included in it are the primordia of the suprarenal medulla and of the celiac, superior mesenteric, and renal plexuses. Nerve fibres and "paraganglion" (M3) cells enter.
- Pancreas - ventral pancreas has now fused with dorsal (Streeter 1948). Perhaps the ventral and dorsal ducts have begun to blend (Russu and Vaida 1959).
Stage 18
- Epiphysis - cellular migration in the pineal body forms a distinct "anterior lobe" in which follicles appear (Stadium 3 of Turkewitsch 1933) (O'Rahilly 1973 a).
- Thymus- thymus makes contact with the thyroid gland and contains a series of canals internally (Weller 1933).
- Thyroid - median thyroid is in contact with "lateral thyroid compo- nents" (Weller 1933) but others have maintained that the telopharyngeal body should not be regarded as a thyroid component (Bejdl and Politzer 1953). The lobes of the thyroid are "composed of series of continuously communicating solid annectent bars" (Weller 1933). This is "the earliest stage of the definitive thyroid" (ibid.). First differentiation occurs in Weller's (1933) "lateral thyroid component," which is beginning to "blend into uniformly constituted thyroid tissue". Weller (1933) illustrated (Fig. 11) a thyroid gland that still showed continuity between its pedicle and the epithelium of the pharynx.
- Adrenal Cortex - gland becomes reorganized. The C1, 2, and 3 cells form cords as sinusoids develop. Cells divide at or near the surface, where new cells are added.[2]
Stage 19
- Hypophysis - the caudal part of the craniopharyngeal pouch is reduced to a closed epithelial stem (Andersen et al. 1971).
- Epiphysis - the "anterior lobe" of the pineal body shows a characteristic step and wedge appearance (Stadium 4 of Turkewitsch 1933) (O'Rahilly 1968).
- Parathyroids - 3 become detached from the pharyngeal endo- derm (Jirfisek 1980).
- Adrenal Cortex - C2 cells lie on the surface of the gland and form a "capsule".[2]
- Adrenal Medulla - Sympathicoblasts penetrate the cortex at stages 19 and 20, and form scattered islets of medullary tissue throughout the cortex (Jirfisek 1980).
Stage 20
- Hypophysis - the adenohypophysial epithelium adjacent to the neurohy- pophysis constitutes the beginning pars intermedia (O'Rahilly 1973 a).
The walls of the craniopharyngeal pouch bud into the mesenchyme (An- dersen et al. 1971 ; Jir/tsek 1980).
- Thymus - the right and left components are in contact with each other (Weller 1933)but are "never completely fused" (Norris 1938, Siegler 1969). Thymic cortex appears (in stages 20-22) as a result, according to Norris
(1938), of migration of and covering by "cells derived from the cervical sinus".
- Parathyroids - the parathyroid glands are attached to the lateral lobes of the thyroid (Weller 1933).
Weller (1933, Fig. 23) showed parathyroid 3 still rostral to parathyroid 4 at 23 mm, whereas (presumably due to variation in the "descent" of the thymus) Norris (1937, Fig. 4) showed parathyroid 3 rostral to, level with, and caudal to parathyroid 4 in embryos of 16-17 mm.
- Thyroid - the "annectent bars" of the thyroid are more compact then previously (Weller 1933). The thyroid now exhibits its definitive external form.
Stage 21
- Hypophysis - the pharyngeal stalk becomes fragmented (Jirfisek 1980).
- Adrenal Cortex - the cellular "capsule" becomes covered by a layer of fibrous tissue.[2]
Stage 22
- Parathyroids - Parathyroids 4 become detached from the pharyngeal endoderm (Jirfisek 1980).
- Adrenal Cortex - the C2 cells have changed and resemble fibrocytes.[2]
Stage 23
- Pituitary - adenohypophysis loss of the stalk and lobules of epithelium project into the mesodermal component of the gland, and oriented epithelial follicles are present (Streeter, 1951, plate 2). Abundant angioblasts and capillaries are found.
- Epiphysis - The pineal body has reached Stadium 5 of Turkewitsch (1933).[5]
- Thymus - The cortex is well-developed, "true lobulation" has begun with the appearance of" fine superficial scallops," lymphocytes are present sparsely in the subcortical zone, and vessels are found within the thymus (Norris 1938).
- Adrenal Cortex - It appears that C2 cells first enter the body of the gland at this stage. The pattern of the arterial supply is established. The cellular "capsule" is penetrated by arterial capillaries which join the sinusoids. Their points of entry give the surface of the gland an appearance of cobblestones. The zona glomerulosa is formed of CI and C3 cells. Cells from this zone and from the "capsule" migrate centrally into the cords.[2]
- Adrenal Medulla - Nerve fibres and neuroblasts are first seen in the body of the gland. The paragangtion (M3) cells are beginning to multiply rapidly and, from 30 mm (stage 23) until birth, some are differentiating into chromaffin cells.[2]
Table
| Week | Carnegie Stage | Pineal (Epiphysis) | Pituitary (Hypophysis) | Thyroid | Parathyroid | Thymus | Pancreas | Adrenal (Suprarenal) Cortex/Medulla | |
|---|---|---|---|---|---|---|---|---|---|
| Week 4 | Stage 13 | basement membranes of the craniopharyngeal pouch and the brain are clearly in contact (O'Rahilly 1973). | median thyroid is now bilobed and is connected to the pharynx by a hollow pedicle (Weller 1933). The telopharyngeal body has been regarded as a "lateral thyroid component" by some workers (e.g. Weller 1933). | Weller (1933) recognized already a thymic primordium "of considerable size" on the ventral part of the third pharyngeal pouch, whereas Norris (1938) considered this stage to be "preprimordial" | Pancreas - ventral pancreas may perhaps be distinguishable (Politzer 1952). | ||||
| Week 5 | Stage 14 | a slight irregularity in the surface outline of the intact head corresponds to the future pineal body (O'Rahilly et al. 1982). Thymus. Weller's (1933) "thymus" (the third pharyngeal pouch) becomes elongated. | craniopharyngeal pouch is prominent (Streeter 1945) and the notochord appears to be inserted into its dorsal wall. The craniopharyngeal pouch has become elongated and blood vessels are beginning to grow in between the basement membranes of the pouch and brain (O'Rahilly 1973a). | thyroid pedicle shows further elongation but is still connected to the epithelium of the pharynx (Weller 1933). Right and left lobes and an isthmus may perhaps be presaged (ibid.). | "Parathyrogenic zones" (Politzer and Hann 1935) are recognizable (Streeter 1945). The parathyroid 4 primordium has been illustrated at this stage by Weller (1933, Fig. 16). | ventral pancreas (which may perhaps be distinguishable as early as stage 13) appears as an evagination from the bile duct at stages 14 (Blechschmidt 1973) and 15 (Streeter 1948). It is generally described as unpaired but, at least in some cases, may perhaps be bilobed[3] or even multiple (Delmas 1939). | Cortex - A change in the characteristics of the cells of the coelomic epithelium appears between the mesogastrium and the lateral end of the mesonephros.[2]
Medulla - paravertebral sympathetic ganglia increase in size as a result of cell division and the addition of nerve fibres from the rami communicantes. The ganglia contain three types of cells: MI, M2, and M3. The M3 cells are the" parasympathetic cells" of Zuckerkandl.[2] | ||
| Week 5 | Stage 15 | pineal body is detectable in the roof of the diencephalon (Stadium I of Turkewitsch 1933) (O'Rahilly 1968). | thyroid primordium may be detached from the pharyngeal epithelium in some instances. "At about the time" when the thyroglossal duct "becomes broken it loses its lumen" (Grosser 1912). | Cortex - primordium is first recognizable. A new type of cell (C1) from the coelomic epithelium is found in the subjacent mesenchyme. New cells (C2) appear in the medial wall of mesonephric glomeruli and begin to migrate into the suprarenal primordium.[2] Jirfisek (1980) denies a mesonephric contribution to the suprarenal. Medulla - all types of cells (M1, M2, and M3) increase in number. From stage 15 to stage 18, the suprarenal primordium is cigar-shaped and extends from segment T6 to segment L1, lateral to the aorta and mesogastrium.[2] | |||||
| Week 6 | Stage 16 | cellular migration in an external direction occurs in the pineal body during stages 16 and 17 (Stadium 2 of Turkewitsch 1933) (O'Rahilly 1968). | slight indication of the infundibular recess may be seen in some embryos (O'Rahilly 1973 a). | has lost its continuity with the pharynx and it consists of two lobes, an isthmus, and a remnant of the pedicle (Weller 1933). | parathyrogenic zones are closely related to the third and fourth aortic arches at 9 mm (Politzer and Hann 1935, unstaged embryo). Parathyroid 3 is identifiable on the anterior wall of the third pharyngeal pouch (Weller 1933, Fig. 17) and "does not arise from a dorsal lobule" of the pouch (Norris 1937). The "sudden appearance of well-differentiated clear chief cells in the early primordia of the parathyroids" at 9 mm was emphasized by Norris (1937). | according to Norris (1938), "not until the primordium of the parathyroid [3] has been outlined can the remaining portion of the third pouch be recognized, by exclusion, as the primordium of the endodermal thymus". | dorsal pancreas and the ventral pancreas are contiguous.[4] | Cortex - Another type of cell (C3) arises from the coelomic epithe- lium. Both C1 and C3 cells enter the suprarenal primordium. An "enormous immigration" of C2 cells occurs.[2] Medulla - cells of neural origin are migrating into the gland, separating the cortical cells into islands. Nerve fibres from the ganglia ac- company the M1 and M3 cells. The M2 cells remain in the ganglia and become sympathetic ganglion cells.[2] | |
| Week 6 | Stage 17 | juxtacerebral wall of the craniopharyngeal pouch is the thicker. The lateral lobes (future infundibular, or tuberal, part) and the anterior chamber (Vorraum) are clearly visible (O'Rahilly 1973 a). The infundibular recess displays a characteristically folded wall, namely the neurohypophysis (O'Rahilly 1973 a). | lobes of the thyroid curve around the carotid arteries and are connected by a delicate isthmus. Lacunae "should not be confused with lumina of follicles" (Weller 1933). | parathyroid 4 is attached to the lateral surface of what Weller (1933) termed the "lateral thyroid component" | connection of the thymus with the pharynx has been severed (Weller 1933). The thymus is intimately approximated to the cervical duct (ibid.) According to Norris (1937), both third and fourth pouches make contact with the ectoderm, although only the third "receives an increment from the ectoderm". | ventral pancreas has now fused with dorsal (Streeter 1948). Perhaps the ventral and dorsal ducts have begun to blend (Russu and Vaida 1959). | Cortex - dorsal part of the whole suprarenal primordium is disorganized by the invasion of sympathetic nerves and cells, while the band of C2 cells and the coelomic epithelium remain intact (Crowder 1957). Medulla - first neural migration is at its height. Growth of the para-aortic complex is extensive. The plexiform complex is derived from paravertebral sympathetic ganglia T6-12 and usually L 1. Included in it are the primordia of the suprarenal medulla and of the celiac, superior mesenteric, and renal plexuses. Nerve fibres and "paraganglion" (M3) cells enter. | ||
| Week 7 | Stage 18 | cellular migration in the pineal body forms a distinct "anterior lobe" in which follicles appear (Stadium 3 of Turkewitsch 1933) (O'Rahilly 1973 a). | median thyroid is in contact with "lateral thyroid components" (Weller 1933) but others have maintained that the telopharyngeal body should not be regarded as a thyroid component (Bejdl and Politzer 1953). The lobes of the thyroid are "composed of series of continuously communicating solid annectent bars" (Weller 1933). This is "the earliest stage of the definitive thyroid" (ibid.). First differentiation occurs in Weller's (1933) "lateral thyroid component," which is beginning to "blend into uniformly constituted thyroid tissue". Weller (1933) illustrated (Fig. 11) a thyroid gland that still showed continuity between its pedicle and the epithelium of the pharynx. | thymus makes contact with the thyroid gland and contains a series of canals internally (Weller 1933). | Cortex - gland becomes reorganized. The C1, 2, and 3 cells form cords as sinusoids develop. Cells divide at or near the surface, where new cells are added.[2] | ||||
| Week 7 | Stage 19 | "anterior lobe" of the pineal body shows a characteristic step and wedge appearance (Stadium 4 of Turkewitsch 1933) (O'Rahilly 1968). | caudal part of the craniopharyngeal pouch is reduced to a closed epithelial stem (Andersen et al. 1971). | parathyroid 3 become detached from the pharyngeal endoderm (Jirfisek 1980). | Cortex - C2 cells lie on the surface of the gland and form a "capsule".[2] Medulla - Sympathicoblasts penetrate the cortex at stages 19 and 20, and form scattered islets of medullary tissue throughout the cortex (Jirfisek 1980). | ||||
| Week 8 | Stage 20 | adenohypophysial epithelium adjacent to the neurohy- pophysis constitutes the beginning pars intermedia (O'Rahilly 1973 a).
The walls of the craniopharyngeal pouch bud into the mesenchyme (An- dersen et al. 1971 ; Jir/tsek 1980). |
"annectent bars" of the thyroid are more compact then previously (Weller 1933). The thyroid now exhibits its definitive external form. | parathyroid glands are attached to the lateral lobes of the thyroid (Weller 1933). Weller (1933, Fig. 23) showed parathyroid 3 still rostral to parathyroid 4 at 23 mm, whereas (presumably due to variation in the "descent" of the thymus) Norris (1937, Fig. 4) showed parathyroid 3 rostral to, level with, and caudal to parathyroid 4 in embryos of 16-17 mm. | right and left components are in contact with each other (Weller 1933)but are "never completely fused" (Norris 1938, Siegler 1969). Thymic cortex appears (in stages 20-22) as a result, according to Norris
(1938), of migration of and covering by "cells derived from the cervical sinus". |
||||
| Week 8 | Stage 21 | pharyngeal stalk becomes fragmented (Jirfisek 1980). | Cortex - the cellular "capsule" becomes covered by a layer of fibrous tissue.[2] | ||||||
| Week 8 | Stage 22 | Parathyroids 4 become detached from the pharyngeal endoderm (Jirfisek 1980). | Cortex - the C2 cells have changed and resemble fibrocytes.[2] | ||||||
| Week 8 | Stage 23 | pineal body has reached Stadium 5 of Turkewitsch (1933).[5] | adenohypophysis loss of the stalk and lobules of epithelium project into the mesodermal component of the gland, and oriented epithelial follicles are present (Streeter, 1951, plate 2). Abundant angioblasts and capillaries are found. | cortex is well-developed, "true lobulation" has begun with the appearance of" fine superficial scallops," lymphocytes are present sparsely in the subcortical zone, and vessels are found within the thymus (Norris 1938). | Cortex - It appears that C2 cells first enter the body of the gland at this stage. The pattern of the arterial supply is established. The cellular "capsule" is penetrated by arterial capillaries which join the sinusoids. Their points of entry give the surface of the gland an appearance of cobblestones. The zona glomerulosa is formed of CI and C3 cells. Cells from this zone and from the "capsule" migrate centrally into the cords.[2] Medulla - Nerve fibres and neuroblasts are first seen in the body of the gland. The paragangtion (M3) cells are beginning to multiply rapidly and, from 30 mm (stage 23) until birth, some are differentiating into chromaffin cells.[2] |
Endocrinology - An Integrated Approach
| Thyroid | |
|---|---|

|
Endocrinology - An Integrated Approach Stephen Nussey and Saffron Whitehead
St. George's Hospital Medical School, London, UK Oxford: BIOS Scientific Publishers; 2001. ISBN-10: 1-85996-252-1 Copyright © 2001, BIOS Scientific Publishers Limited. Bookshelf |
Chapter 3. The thyroid gland
| |
|
|
Stephen Nussey and Saffron Whitehead.
St. George's Hospital Medical School, London, UK Oxford: BIOS Scientific Publishers; 2001. ISBN-10: 1-85996-252-1 Copyright © 2001, BIOS Scientific Publishers Limited. |
Chapter 1. Principles of endocrinology
Chapter 1. Principles of endocrinology
- Functions of hormones and their regulation
- Chemical signalling - endocrine, paracrine, autocrine and intracrine mechanisms
- Chemical classification of hormones and their synthesis
- Hormone synthesis
- Transport of hormones in the circulation and their half-lives
- Hormone receptors - cell surface
- Hormone receptors - intracellular
- Hormones and gene transcription
- Hormone receptor regulation
- Neuroendocrine interactions
- Hormones and the immune system
- Hormones, growth promotion and malignancy
- Genes, mutations and endocrine function
- Clinical evaluation of endocrine disorders
Chapter 2. The endocrine pancreas
Chapter 2. The endocrine pancreas
- Glucose turnover
- Anabolic and catabolic phases of glucose metabolism
- Actions of insulin and glucagon
- Lipid metabolism - insulinopenia and diabetic ketosis
- Protein metabolism and the anabolic actions of insulin
- Definition and diagnosis of diabetes mellitus
- Etiology of type 1 DM
- Prevention of type 1 DM
- Structure, synthesis and metabolism of insulin and glucagon
- Anatomical features of pancreatic islets in relation to hormone secretion and its control
- Control of insulin and glucagon secretion
- Type 2 DM
- Causes of DM
- Genetic disorders of β-cell function
- Counter-regulatory hormones and DM
- Complications of DM
- Macrovascular circulatory changes
- Microvascular changes - diabetic retinopathy, nephropathy and neuropathy
- Diabetes and the neuropathic foot
- Diabetes and insulin resistance of pregnancy
- Development of the pancreas: effects of DM on organogenesis
- Treatment of DM - rationale and practical considerations
- Hypoglycemia
- Physiological responses to hypoglycemia and its treatment
- Hypoglycemia and insulinoma
- Hypoglycemia in infancy
- Disorders of the α, γ and PP cells of the islets
- Clinical case questions
Chapter 3. The thyroid gland
- Iodine intake
- Anatomical features of the thyroid gland
- Iodine trapping and thyroid function
- Synthesis of thyroid hormones
- Actions of thyroid hormones
- Control of thyroid hormone synthesis and secretion
- Hyperthyroidism — Graves' disease
- Surgical anatomy and embryology of the thyroid gland
- Primary hypothyroidism — Hashimoto's disease and myxedema
- Secondary hypothyroidism
- Hypothyroidism in infancy and childhood
- Thyroid hormone resistance
- Non-thyroid illness (‘sick euthyroid’ syndrome)
- Transport and metabolism of thyroid hormones
- Biochemical measurements of thyroid hormone status
- Thyroid growth
- Nodular thyroid disease
- Thyroid cancer
- Clinical case questions
Chapter 4. The adrenal gland
- Specificity of the biological effects of adrenal steroid hormones
- Cholesterol and steroid synthesis in the adrenal cortex
- Anatomical and functional zonation in the adrenal cortex
- Glucocorticoid receptors
- Actions of glucocorticoids and clinical features of Cushing's syndrome
- Adrenal cortical androgens
- Hypothalamic control of adrenocortical steroid synthesis - CRH and vasopressin
- Pituitary control of adrenocortical steroids - ACTH
- Feedback control of glucocorticoids
- Excess glucocorticoids: biochemical investigation of Cushing's syndrome
- Measurements of cortisol in blood, urine and saliva
- Dynamic tests of endocrine function
- Imaging the adrenal gland
- Treatment of Cushing's syndrome
- Nelson's syndrome
- Excess adrenal androgens - congenital adrenal hyperplasia (CAH)
- Deficiency of adrenocortical secretions - Addison's disease
- Aldosterone and the control of salt and water balance
- Transport and metabolism of adrenocortical steroids
- Selective mineralocorticoid excess and deficiency
- The adrenal medulla and pheochromocytoma
- Catecholamine synthesis and secretion
- Diagnosis and treatment of pheochromocytomas
- Clinical case questions
Chapter 5. The parathyroid glands and vitamin D
Chapter 5. The parathyroid glands and vitamin D
- Calcium and phosphate in serum and its measurement
- Intracellular calcium concentration
- Calcium and phosphate balance
- Hormonal control of serum Ca2+ and Pi concentrations
- Sources, metabolism and transport of vitamin D
- Classical actions of vitamin D on intestine and bone
- Parathyroid glands and PTH synthesis
- Control of PTH secretion
- Actions of PTH
- Hypercalcemia and primary hyperparathyroidism
- Hyperparathyroidism and multiple endocrine neoplasia (MEN)
- Hypercalcemia and vitamin D excess
- Hypercalcemia and malignancy
- Parathyroid hormone-related peptide (PTHrp)
- Treatment of hypercalcemia
- Mutations of the Ca2+ or PTH receptors
- Hypocalcemia and its treatment
- Pseudohypoparathyroidism
- Vitamin D deficiency
- Non-classical actions of vitamin D
- Vitamin D resistance and rickets
- Hormones and the skeleton
- Structure, formation and function of bone
- Osteoporosis
- Paget's disease (osteitis deformans)
- Calcitonin and calcitonin gene-related peptide
- Clinical case questions
Chapter 6. The gonad
- Genetic determination of sexual differentiation
- Sexual differentiation of the gonads and internal reproductive tracts
- Sexual differentiation of the external genitalia
- Control of steroid production in the fetal gonads
- Puberty
- GnRH and the control of gonadotrophin synthesis and secretion
- The gonadotrophins - LH and FSH - and their actions
- Endocrine changes in puberty
- Precocious sexual development
- Delayed puberty
- Premature adrenarche
- Acne, hair growth and hirsutism
- The breast - premature development, hypoplasia and gynecomastia
- Testicular function
- Control of testicular function
- Transport, metabolism and actions of androgens
- Spermatogenesis
- Erection and ejaculation
- Ovarian control and the menstrual cycle
- Transport, metabolism and actions of ovarian steroids
- The ovary - folliculogenesis and oogenesis
- Non-steroidal factors in the control of the hypothalamic-pituitary-gonadal axis
- Ovulation, menstruation and its problems
- Polycystic ovary syndrome (PCOS)
- Contraception
- Infertility
- Ovulation induction and assisted conception
- Ovarian failure, the menopause and andropause
- Hormonal replacement therapy (HRT) and selective estrogen receptor modulators (SERMS)
- Clinical case questions
Chapter 7. The pituitary gland
Chapter 7. The pituitary gland
- Anatomical and functional connections of the hypothalamo-pituitary axis
- Embryology of the pituitary gland
- Craniopharyngioma
- Blood supply of the hypothalamo-pituitary axis
- Sheehan's syndrome
- Growth and somatotrophin deficiency
- Growth hormone - secretory patterns and control
- Actions of growth hormone and insulin-like growth factors
- GH replacement therapy
- GH excess - gigantism and acromegaly
- Pituitary adenomas - incidence and treatment
- Prolactinomas
- Prolactin and its control
- Circadian rhythms and the suprachiasmatic nucleus
- The pineal gland and melatonin
- Autonomic functions of the hypothalamus
- Obesity
- The neural lobe of the pituitary gland - AVP and oxytocin
- Clinical case questions
Chapter 8. Cardiovascular and renal endocrinology
Chapter 8. Cardiovascular and renal endocrinology
- Endocrinology of heart failure
- Paracrine and autocrine regulation of blood pressure: the endocrinology of sepsis
- Hormones and blood cell production - erythropoietin
- Carcinoid
- Clinical case questions
References
Effects of environmental endocrine disruptors on pubertal development
J Clin Res Pediatr Endocrinol. 2011 Mar;3(1):1-6. Epub 2011 Feb 23.
Ozen S, Darcan S.
Pediatric Endocrinology Unit, Mersin Children Hospital, Mersin, Turkey. Abstract The onset and course of puberty are under the control of the neuroendocrine system. Factors affecting the timing and regulation of the functions of this system may alter the onset and course of puberty. Several environmental endocrine disruptors (EDs) with significant influences on the normal course of puberty have been identified. Numerous animal and human studies concerning EDs have been conducted showing that these substances may extensively affect human health; nevertheless, there are still several issues that remain to be clarified. In this paper, the available evidence from animal and human studies on the effects of environmental EDs with the potential to cause precocious or delayed puberty was reviewed.Conflict of interest:None declared.
PMID 21448326
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3065309
2012
Neurobehavioral risk is associated with gestational exposure to stress hormones
Expert Rev Endocrinol Metab. 2012 Jul;7(4):445-459.
Sandman CA, Davis EP. Source Department of Psychiatry & Human Behavior, Women and Children's Health and Well-Being Project, University of California, Irvine, Orange, CA, USA.
Abstract
The developmental origins of disease or fetal programming model predict that early exposures to threat or adverse conditions have lifelong consequences that result in harmful outcomes for health. The maternal endocrine 'fight or flight' system is a source of programming information for the human fetus to detect threats and adjust their developmental trajectory for survival. Fetal exposures to intrauterine conditions including elevated stress hormones increase the risk for a spectrum of health outcomes depending on the timing of exposure, the timetable of organogenesis and the developmental milestones assessed. Recent prospective studies, reviewed here, have documented the neurodevelopmental consequences of fetal exposures to the trajectory of stress hormones over the course of gestation. These studies have shown that fetal exposures to biological markers of adversity have significant and largely negative consequences for fetal, infant and child emotional and cognitive regulation and reduced volume in specific brain structures.
PMID 23144647
External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
Histology
Adult
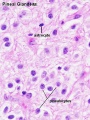
Pineal (high power)
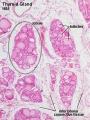
Thyroid (low power)
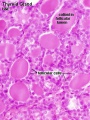
Thyroid (high power)
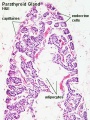
Parathyroid (low power)
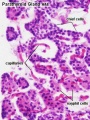
Parathyroid (high power)
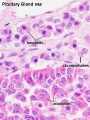
Pituitary - adenohypophysis
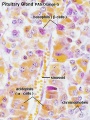
Pituitary - adenohypophysis
Pituitary - neurohypophysis
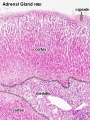
Adrenal - Cortex and Medulla
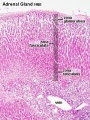
Adrenal - Cortical Zones
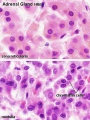
Adrenal - Zona Reticularis and Medulla
Pancreatic islet












Embryonic
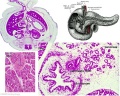
Stage 22 - Pancreatic duct
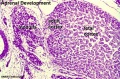
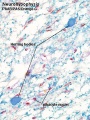
Stage 22 - Adrenal gland
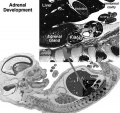
Week 10 - Adrenal gland



Terms
adrenocorticotropin - (ACTH or corticotropin) anterior pituitary, peptide hormone
antidiuretic hormone - (ADH) hypothalamus, peptide hormone
atrial natriuretic factor - (ANP) heart, , peptide hormone
calcitonin - (CT) C cells of thyroid, peptide hormone
follicle stimulating hormone - (FSH) pituitary, protein hormone
growth hormone - (GH) pituitary, peptide hormone
human chorionic gonadotropin - (hCG) pancreas glycoprotein hormone with 2 subunits (alpha and beta joined non covalently). Similar in structure to luteinizing hormone (LH), hCG exists in multiple hormonal and non-endocrine agents (regular hCG, hyperglycosylated hCG and the free beta-subunit of hyperglycosylated hCG). PMID: 19171054
lutenizing hormone - (LH) pituitary, protein hormone
melaocyte stimulating hormone - (MSH) pituitary, peptide hormone
prolactin - (PRL) pituitary, peptide hormone
parathyroid hormone - (PTH) parathyroid, peptide hormone
thyroid hormone - (TH) thyroid,amino acid derivative
thyroid stimulating hormone - (TSH) pituitary, protein hormone
- ↑ O'Rahilly R. The timing and sequence of events in the development of the human endocrine system during the embryonic period proper. (1983) Anat. Embryol., 166: 439-451. PMID 6869855
- ↑ 2.00 2.01 2.02 2.03 2.04 2.05 2.06 2.07 2.08 2.09 2.10 2.11 2.12 2.13 2.14 2.15 2.16 2.17 2.18 2.19 2.20 2.21 2.22 2.23 Crowder RE. The development of the adrenal gland in man, with special reference to origin and ultimate location of cell types and evidence in favor of the "cell migration" theory. (1957) Contrib. Embryol., Carnegie Inst. Wash. 36, 193-210.
- ↑ 3.0 3.1 Odgers PN. Some observations on the development of the ventral pancreas in man. (1930) J. Anat., 65(1): 1-7. PMID 17104298
- ↑ 4.0 4.1 Blechschmidt E. Die prdnatalen Organsysteme des Menschen. (1973) Hippokrates, Stuttgart.
- ↑ 5.0 5.1 O'Rahilly R. The development of the epiphysis cerebri and the subcommissural complex in staged human embryos. (1968) Anat. Rec., 160: 488-489.