|
|
| (23 intermediate revisions by the same user not shown) |
| Line 1: |
Line 1: |
| {{Header}} | | {{Header}} |
| {{Ref-Conklin1905}} | | {{Ref-Conklin1905}} |
| | {{Conklin1905 TOC}} |
| | {| class="wikitable mw-collapsible mw-collapsed" |
| | ! Online Editor |
| | |- |
| | | [[file:Mark_Hill.jpg|90px|left]] Ascidians ([[Sea Squirt Development|sea squirts]]) are the evolutionary link between invertebrates and vertebrates. Conklin's 1905 detailed developmental cell lineage study of the ascidian embryo, that inspired similar studies in other species. This also set the stage for identifying embryonic patterning. |
| | <br> |
| | Edwin Grant Conklin (1863 – 1952) was an American biologist and zoologist at the Department of Biology at Princeton University. He was Chair of Biology for twenty-five years and studied the embryonic development of marine animals. |
| | <br> |
| | <br> |
| | |
| | See also - {{Ref-Conklin1905b}} |
| | <br> |
| | <br> |
| | |
| | [[Sea Squirt Development]] | [[Media:Conklin EG-The Organization and Cell-Lineage of the Ascidian Egg-1905.pdf|Book PDF]] |
| | |
| | [[Historic Embryology Papers]] |
| | |} |
| {{Historic Disclaimer}} | | {{Historic Disclaimer}} |
|
| |
| =The Organization and Cell-Lineage of the Ascidian Egg= | | =The Organization and Cell-Lineage of the Ascidian Egg= |
| | [[File:Edwin Conklin.jpg|thumb|150px|alt=Edwin Conklin|Edwin Conklin (1863 – 1952)]] |
| | [[File:Conklin 1905 plate01.jpg|thumb|Living Egg of Oynthia partita - Maturation and Fertilization]] |
|
| |
|
| By Edwin G. Conklin. | | By Edwin G. Conklin. |
| | |
|
| |
|
| Professor of Zoology, University of Pennsylvania, | | Professor of Zoology, University of Pennsylvania, |
|
| |
|
| WITH PLATES I-XII.
| |
|
| |
|
| | With Plates I-XII. |
|
| |
|
| Philadelphia : 1905
| |
|
| |
|
| | Philadelphia - 1905 |
|
| |
|
| ==Contents== | | ==Contents== |
|
| |
|
| Introduction. | | [[#Introduction|Introduction]] |
| | | * A. Organization of the Egg |
| A. Organization of the Egg | | * B. Ascidian Embryology |
| | | * C. Methods and Material |
| B. Ascidian Embryology | |
| | |
| C. Methods and Material | |
| | |
| | |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 1|I. The Ovarian Egg]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 1|I. The Ovarian Egg]] |
| | | # Development of the Ovocyte |
| 1. Development of the Ovocyte
| | # Test Cells and Chorion . |
| | | # Structure of Fully Formed Ovocyte |
| 2. Test Cells and Chorion .
| |
| | |
| 3. Structure of Fullv Formed Ovocyte
| |
| | |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 2|II. Maturation and Fertilization]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 2|II. Maturation and Fertilization]] |
| | * A. Maturation |
| | # Disappearance of Nuclear Membrane |
| | # Chromosomes |
| | # Nucleolus |
| | # Spindle Formation |
| | # Movements of Spindle and Nuclear Plasm ; Formation of |
|
| |
|
| A. Maturation
| | * B. Fertilization |
| | | # Entrance of Spermatozoon |
| 1. Disappearance of Nuclear Membrane
| | # Movements of Ooplasm |
| | | ## Localization of Yellow Protoplasm |
| 2. Chromosomes
| | ## Localization of Clear Protoplasm and Yolk |
| | | # Development of Sperm Nucleus and Aster |
| 3. Nucleolus
| | # Path of Spermatozoon within the Egg . |
| | | # The Egg Nucleus and its Movements |
| 4. Spindle Formation
| | # Sperm Amphiaster and First Cleavage Spindle |
| | | # Dispermy |
| 5. Movements of Spindle and Nuclear Plasm ; Formation of
| | * Polar Bodies |
| | |
| B. Fertilization | |
| | |
| 1. Entrance of Spermatozoon
| |
| | |
| 2. Movements of Ooplasm
| |
| a. Localization of Yellow Protoplasm
| |
| b. Localization of Clear Protoplasm and Yolk
| |
| | |
| 3. Development of Sperm Nucleus and Aster
| |
| | |
| 4. Path of Spermatozoon within the Egg .
| |
| 5. The Egg Nucleus and its Movements
| |
| | |
| 6. Sperm Amphiaster and First Cleavage Spindle
| |
| | |
| 7. Dispermy
| |
| | |
| Polar Bodies | |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 3|III. Orientation of Egg and Embryo]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 3|III. Orientation of Egg and Embryo]] |
| | | # Van Beneden and Julin's System of Orientation |
| 1. Van Beneden and Julin's System of Orientation
| | # Seeliger's System |
| | | # Samassa's System |
| 2. Seeliger's System
| | # Castle's System |
| | | # Evidences in Favor of Van Beneden and Julin's System |
| 3. Samassa's System
| |
| | |
| 4. Castle's System
| |
| | |
| 5. Evidences in Favor of Van Beneden and Julin's System
| |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 4|IV. Cell-Lineage]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 4|IV. Cell-Lineage]] |
| Line 86: |
Line 79: |
|
| |
|
| B. Cleavage of the Egg ; First to Seventh Generation of Cells | | B. Cleavage of the Egg ; First to Seventh Generation of Cells |
| | | # First Cleavage ; 1-2 Cells |
| 1. First Cleavage ; 1-2 Cells
| | # Second Cleavage ; 2-4 Cells . |
| | | # Third Cleavage; 4-8 Cells . |
| 2. Second Cleavage ; 2-4 Cells .
| | # Fourth Cleavage; 8-16 Cells |
| | | # Fifth Cleavage ; 16-32 Cells |
| 3. Third Cleavage; 4-8 Cells .
| | # Sixth Cleavage ; 32-64 Cells |
| | |
| 4. Fourth Cleavage; 8-16 Cells
| |
| | |
| 5. Fifth Cleavage ; 16-32 Cells
| |
| | |
| 6. Sixth Cleavage ; 32-64 Cells
| |
|
| |
|
| C. Gastrulation ; Seventh to Ninth Generation of Cells | | C. Gastrulation ; Seventh to Ninth Generation of Cells |
| Line 104: |
Line 91: |
|
| |
|
| 8. Eighth Cleavage ; 112-132 Cells, 132-218 Cells | | 8. Eighth Cleavage ; 112-132 Cells, 132-218 Cells |
|
| |
|
| |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 5|V. Later Development]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 5|V. Later Development]] |
| | | # Closure of Blastopore |
| 1. Closure of Blastopore
| | # Development of Larva |
| | |
| 2. Development of Larva
| |
| | |
| | |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 6|VI. Comparisons with A.mphioxus and Amphibia]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 6|VI. Comparisons with A.mphioxus and Amphibia]] |
| | | # Axial Relations of Egg and Embryo |
| 1. Axial Relations of Egg and Embryo
| | # Entrance of Spermatozoon |
| | | # Cleavage |
| 2. Entrance of Spermatozoon
| | # Blastula and Gastrula |
| | | # Closure of Blastopore |
| 3. Cleavage
| | # Neural Plate |
| | | # Chorda |
| 4. Blastula and Gastrula
| | # Origin of Mesoderm |
| | |
| 5. Closure of Blastopore
| |
| | |
| 6. Neural Plate
| |
| | |
| 7. Chorda
| |
| | |
| 8. Origin of Mesoderm
| |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 7|VII. The Organization of the Egg]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 7|VII. The Organization of the Egg]] |
| | * A. Polarity |
| | * B. Symmetry |
| | * C. Cytoplasmic Localization |
| | # Localization in the Cleavage Stages Cytoplasmic Organization and the Nuclear |
| | # Localization before Cleavage |
| | * D. Genesis of the Organization of the Egg |
| | # Role of the Nucleus in Differentiation - Inheritance Theory |
| | # Factors of Localization |
| | ## Cytoplasmic Movements |
| | ## Cell Division as a Factor of Localization |
| | * Types of Germinal Localization ; Evolution of Types |
| | # Annelid-Mollusk Type |
| | # Ctenophore Type |
| | # Echinoderm Type |
| | # Ascidian Type |
|
| |
|
| A. Polarity
| | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 8|Summary]] |
| | # Ascidian Embryology |
| | # Cytological Results |
| | # Organization of the Egg |
|
| |
|
| B. Symmetry
| | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 9|Literature Cited]] |
|
| |
|
| C. Cytoplasmic Localization
| | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 10|Explanation of Figures]] |
|
| |
|
| 1. Localization in the Cleavage Stages
| | <gallery> |
| | File:Conklin 1905 plate01.jpg|Plate I |
| | File:Conklin 1905 plate02.jpg|Plate II |
| | File:Conklin 1905 plate03.jpg|Plate III |
| | File:Conklin 1905 plate04.jpg|Plate IV |
| | File:Conklin 1905 plate05.jpg|Plate V |
| | File:Conklin 1905 plate06.jpg|Plate VI |
| | File:Conklin 1905 plate07.jpg|Plate VII |
| | File:Conklin 1905 plate08.jpg|Plate VIII |
| | File:Conklin 1905 plate09.jpg|Plate IX |
| | File:Conklin 1905 plate10.jpg|Plate X |
| | File:Conklin 1905 plate11.jpg|Plate XI |
| | File:Conklin 1905 plate12.jpg|Plate XII |
| | </gallery> |
|
| |
|
| | <gallery> |
| | File:Conklin 1905 fig01-02.jpg|Fig 1-2 |
| | File:Conklin 1905 fig03-06.jpg|Fig 3-6 |
| | File:Conklin 1905 fig07-08.jpg|Fig 7-8 |
| | File:Conklin 1905 fig09-12.jpg|Fig 9-12 |
| | File:Conklin 1905 fig13-16.jpg|Fig 13-16 |
| | File:Conklin 1905 fig17-20.jpg|Fig 17-20 |
| | File:Conklin 1905 fig21-24.jpg|Fig 21-24 |
| | File:Conklin 1905 fig25-26.jpg|Fig 25-26 |
| | File:Conklin 1905 fig27-33.jpg|Fig 27-33 |
| | File:Conklin 1905 fig34-35.jpg|Fig 34-35 |
| | </gallery> |
| | ==Introduction== |
|
| |
|
| Cytoplasmic Organization and the Nuclear
| | ===A. Organization of the Egg=== |
| | | Recent years have witnessed a revival of the ancient controversy as to the nature and contents of the germ cells. On the one hand are those who with Weismann maintain that the egg must contain the elements or determinants of very many structures which will appear in the course of development; on the other hand are ranged the modern epigenesists who find in the egg cell only complex chemical substances which have the capacity under certain outer conditions of undergoing regular transformations into other substances which incidentally have peculiar forms, just as crystals have. |
| 2. Localization before Cleavage
| |
| | |
| D. Genesis of the Organization of the Egg
| |
| | |
| 1. Role of the Nucleus in Differentiation
| |
| | |
| Inheritance Theory
| |
| | |
| 2. Factors of Localization
| |
| | |
| a. Cytoplasmic Movements
| |
| | |
| b. Cell Division as a Factor of Localization
| |
| | |
| E. Types of Germinal Localization ; Evolution of Types
| |
| | |
| 1. Annelid-Mollusk Type
| |
| | |
| 2. Ctenophore Type
| |
| | |
| 3. Echinoderm Type
| |
|
| |
|
| 4. Ascidian Type
| |
|
| |
| Summary
| |
|
| |
| I. Ascidian Embryology
| |
|
| |
| II. Cytological Results
| |
|
| |
| III. Organization of the Egg
| |
|
| |
| Literature Cited
| |
|
| |
| Explanation of Figures
| |
|
| |
|
| |
|
| |
| ==Introduction==
| |
|
| |
| A. Organization of the Egg. Recent years have witnessed a revival of
| |
| the ancient controversy as to the nature and contents of the germ cells. On
| |
| the one hand are those who with Weismann maintain that the egg must contain
| |
| the elements or determinants of very many structures which will appear in the
| |
| course of development; on the other hand are ranged the modern epigenesists
| |
| who find in the egg cell only complex chemical substances which have the capacity
| |
| under certain outer conditions of undergoing regular transformations into other
| |
| substances which incidentally have peculiar forms, just as crystals have.
| |
|
| |
|
| But while this modern controversy recalls the ancient one between the | | But while this modern controversy recalls the ancient one between the |
| Line 204: |
Line 174: |
| somewhere between these two extremes ; the real problem is how much or how | | somewhere between these two extremes ; the real problem is how much or how |
| little of organization is present, and not whether the germ is organized at all. | | little of organization is present, and not whether the germ is organized at all. |
| | |
|
| |
|
| Though the controversy as to evolution and epigenesis has thus been narrowed within relatively small limits, and has therein- lost much of its startling and | | Though the controversy as to evolution and epigenesis has thus been narrowed within relatively small limits, and has therein- lost much of its startling and |
| Line 213: |
Line 184: |
| structures in earlier and earlier stages of development and so finally in the | | structures in earlier and earlier stages of development and so finally in the |
| unsegmented egg itself. | | unsegmented egg itself. |
| | |
|
| |
|
| It is not many years since all embryological studies were dominated by the | | It is not many years since all embryological studies were dominated by the |
| Line 226: |
Line 198: |
| the basis of any thorough study of development, inheritance and evolution. | | the basis of any thorough study of development, inheritance and evolution. |
|
| |
|
| B. Ascidian Embryology. Anyone who has observed the ascidian egg will | | ===B. Ascidian Embryology=== |
| understand why it has been such a favorite object of study. The cleavage of the | | Anyone who has observed the ascidian egg will understand why it has been such a favorite object of study. The cleavage of the |
| egg is so beautifully regular and can be observed so readily in life that it is not | | egg is so beautifully regular and can be observed so readily in life that it is not |
| surprising that ascidians were among the first animals to which the "cell-lineage" | | surprising that ascidians were among the first animals to which the "cell-lineage" |
| Line 265: |
Line 237: |
| me to say that I have spared no pains or labor to make them accurate. | | me to say that I have spared no pains or labor to make them accurate. |
|
| |
|
| ('. Material and Methods. Early in July. 1903, while working at the Marine
| | ===C. Material and Methods=== |
| Biological Laboratory, Woods Holl, Mass.. I began the study of the maturation | | |
| | Early in July. 1903, while working at the Marine Biological Laboratory, Woods Holl, Mass.. I began the study of the maturation |
| and fertilization of the egg of Ciona intestinalis (L.) Flemming, with the aim mentioned in the preceding paragraph. Only a small number of these animals was to | | and fertilization of the egg of Ciona intestinalis (L.) Flemming, with the aim mentioned in the preceding paragraph. Only a small number of these animals was to |
| be found at that time at Woods Holl. though they occurred more abundantly later | | be found at that time at Woods Holl. though they occurred more abundantly later |
| Line 283: |
Line 256: |
| I took up also the living eggs of Ciona and Molgula, and finally I fixed and prepared for microscopical examination, both as whole objects and as serial sections, | | I took up also the living eggs of Ciona and Molgula, and finally I fixed and prepared for microscopical examination, both as whole objects and as serial sections, |
| the eggs and embryos of all three of these genera. | | the eggs and embryos of all three of these genera. |
| | |
|
| |
|
| Castle (1896) has described in considerable detail the time and manner of egg | | Castle (1896) has described in considerable detail the time and manner of egg |
| Line 300: |
Line 274: |
| another individual, hut if fertilized with sperm from the same animal the eggs | | another individual, hut if fertilized with sperm from the same animal the eggs |
| rarely if ever develop, as Castle has shown. This is due to the fact that such | | rarely if ever develop, as Castle has shown. This is due to the fact that such |
| spermatozoa never enter the egg, though they may be quite active. Morgan (1904) | | spermatozoa never enter the egg, though they may be quite active. Morgan (1904) has recently discussed this interesting fact in a suggestive manner. |
| lias recently discussed this interesting fact in a suggestive manner.
| | |
|
| |
|
| The method which I employed in studying the living eggs of these ascidians | | The method which I employed in studying the living eggs of these ascidians |
| Line 317: |
Line 291: |
| of several eggs through consecutive portions of it, I chose the latter and easier | | of several eggs through consecutive portions of it, I chose the latter and easier |
| method. | | method. |
| | |
|
| |
|
| All my studies of the living eggs of these ascidians were made with a dry lens, | | All my studies of the living eggs of these ascidians were made with a dry lens, |
| Line 327: |
Line 302: |
| difficult" to explain. I can only account for it by supposing that he obtained the | | difficult" to explain. I can only account for it by supposing that he obtained the |
| eggs in the evening and studied them by yellow artificial light. | | eggs in the evening and studied them by yellow artificial light. |
| | |
|
| |
|
| Preserved material was fixed in various fluids, Perenyi's,. Kleinenberg's, PicroAcetic, Sublimate and Sublimate-Acetic. For the study of entire eggs and embryos | | Preserved material was fixed in various fluids, Perenyi's,. Kleinenberg's, PicroAcetic, Sublimate and Sublimate-Acetic. For the study of entire eggs and embryos |
| Line 338: |
Line 314: |
| satisfactory results. Such material was stained on the slide in Delafield's Hematoxylin and Eosin or in Iron Hematoxylin and Bordeau red. | | satisfactory results. Such material was stained on the slide in Delafield's Hematoxylin and Eosin or in Iron Hematoxylin and Bordeau red. |
|
| |
|
| Castle states that he found it necessary to remove the egg envelopes by | | |
| drawing the egg into a pipette through an opening so small that the egg alone could puss in, after the manner recommended l>\ Chabry. In most cases I have | | Castle states that he found it necessary to remove the egg envelopes by drawing the egg into a pipette through an opening so small that the egg alone could puss in, after the manner recommended by Chabry. In most cases I have |
| found that the presence of the egg envelope's (iocs not seriously interfere with clear | | found that the presence of the egg envelope's (iocs not seriously interfere with clear |
| seeing, possibly owing to the fact that in the study of preparations 1 have used an | | seeing, possibly owing to the fact that in the study of preparations I have used an immersion lens in which the depth of locus is relatively slight. In late stages, |
| | | however, the test cells are sometimes confusing, and in the case of Ciona I found |
| immersion lens in which the depth of locus is relatively slight. In late stages, | | that these, together with the other envelopes, could he removed by simply rolling the eggs under the cover glass. In Cynthia the envelopes may sometimes he removed in the same way, though not so easily as in Ciona. |
| however, the test cells are sometimes confusing, and in the case of Ciona 1 found | |
| that these, together with the other envelopes, could he removed by simply rolling | |
| the eggs under the cover glass. In Cynthia the envelopes may sometimes he | |
| removed in the same way, though not so easily as in Ciona. | |
| | |
| | |
| | |
| I. THE OVARIAN EGG.
| |
| | |
| Much has been written on the egg envelopes and ovarian eggs of ascidians
| |
| and I shall not here go over that ground in any detail. But in searching for the
| |
| earliest different iations of the egg substance it is necessary to go back to the
| |
| ovarian egg, and in so doing I have found some structures the real significance of
| |
| which has not hitherto been appreciated.
| |
| | |
| 1. Development of the Ovocyte.
| |
| | |
| In a young ovocyte the cytoplasm stains uniformly and there is no trace of
| |
| yolk or of test cells. Close around the nucleus is a granular mass which is deeply
| |
| colored by plasma stains, the yolk nucleus or "yolk matrix" of Crampton (1899).
| |
| As the egg grows, small spherules of yolk begin to appear in the vicinity of the
| |
| yolk matrix, and this yolk gradually fills the central portion of the egg surrounding
| |
| the nucleus, while the cytoplasm, which is free from yolk, occupies a peripheral
| |
| position. Some of the follicle cells which surround the egg at this stage then
| |
| invade the egg, thus forming the " test cells" which are located chiefly in the
| |
| peripheral layer of cytoplasm. My observations as to the origin of these " test
| |
| cells " agree with some of the most careful work, both ancient and modern, which
| |
| has been done on this subject (Kowalevsky, 18G6, 1871; Seeliger, 1882; Van
| |
| Beneden and Julin, 1886; Morgan, 1890; Floderus, 189G ; Bancroft, 1899).
| |
| | |
| The earliest appearance of polarity is found in the location of the yolk matrix
| |
| on one side of the nucleus and in a slight eccentricity of the latter. I consider it
| |
| very probable that the yolk matrix is derived from the attraction sphere of the last
| |
| ovogonic mitosis, and that the chief axis of the egg represents the cell axis which
| |
| passes through the centrosome and nucleus, and which, as I have previously shown
| |
| (Conklin, 1902), is preserved in every cell throughout the cleavage of the egg and
| |
| probably also in all later cell divisions. If this be true, the polarity of the egg is a
| |
| differentiation which is carried over from generation to generation, and as this chief
| |
| axis of the egg is identical with the gastrular axis, and bears a constant relationship
| |
| to the principal axes of the embryo and adult, it will be seen that at least one
| |
| important differentiation of an animal is predetermined (not predelineated) at all
| |
| stages. Although this chief axis of the egg is usually recognizable at all stages by a slight eccentricity of the nucleus, it is often difficult to observe it after the disappearance of the yolk matrix. No other axial differentiations of the egg are
| |
| recognizable until after the fertilization.
| |
| | |
| 2. Test Cells and Chorion.
| |
| | |
| In the fully formed ovarian eggs the test cells lie imbedded in a peripheral
| |
| layer of clear protoplasm ; this layer stains intensely with plasma stains, and in the
| |
| living eggs of Cynthia contains yellow pigment granules. In Cynthia the test cells
| |
| are distributed singly and pretty uniformly in this peripheral layer (fig. 61), and
| |
| the same is true of Ciona at an early stage in the formation of the ovocyte (figs. 168,
| |
| 169), but in the fully formed ovarian egg of Ciona the test cells are found in little
| |
| masses or "nests" of from three to six or eight cells each (fig. 170). A similar
| |
| grouping of the test cells has been described by Morgan (1890) in an unidentified
| |
| species of Clavellina. These cells are much smaller and more numerous than the
| |
| test cells of Cynthia, and are evidently formed by division of the original test cells.
| |
| | |
| The test cells of Cynthia become quite large and contain yolk spherules, though
| |
| they do not stain as densely as the yolk of the egg; in Ciona the test cells are very
| |
| much smaller and do not contain these spherules. About the time that the ovarian
| |
| eggs escape from the ovary the test cells are extruded from this peripheral layer of
| |
| protoplasm, and the outlines of the egg, which up to this time have been irregular.
| |
| become more nearly spherical. It is probable that the expulsion of the test cells
| |
| and the assumption of the regular spherical form by the egg have a common cause
| |
| in the increase of surface tension at this time.
| |
| | |
| At the time of the extrusion of the test cells I have observed in the ovarian
| |
| eggs of Ciona a faintly-staining, homogeneous layer which lies inside the outer
| |
| follicle cells and outside of the egg. The test cells lie on the inner border of this
| |
| homogeneous layer; from its general appearance it is highly probable that the substance of which it is composed is extruded from the egg along with the test cells.
| |
| This homogeneous material does not long persist as such but soon disappears and
| |
| probably goes to form the chorion. At this time the egg undergoes considerable
| |
| shrinkage in size, a distinct perivitelline space being formed, and the egg becoming
| |
| regularly spherical [cf. figs. 171 and 172). It is evident that this is clue to the
| |
| escape of fluid from the egg, probably the homogeneous substance described above.
| |
| | |
| In this connection a word or two as to the significance of the test cells may be
| |
| permissible. The fact that in Cynthia they contain yolk and grow to a considerable size, and that spermatozoa not infrequently enter them (figs. 80 and 85 sn.), may
| |
| be taken as evidence that these cells are rudimentan' eggs ; a view which is held by
| |
| Floderus (1896). Bancroft (1899) and others.
| |
| | |
| !. Structure of Fully Formed Ovocyte.
| |
| When first laid the living eggs of Cynthia are, exclusive of the egg envelopes,
| |
| about 150 /< in diameter; those of Ciona are about the same size, but in Molgula
| |
| they are much smaller, being about 100 /< in diameter. The very large germinal vesicle contains an abundant granular precipitate, an enormous nucleolus, and at
| |
| wide intervals within the vesicle, but chiefly near the unclear membrane, a few
| |
| deeply staining chromatic granules. These granules arc small at this time and it is
| |
| difficult to determine their exact shape, though many of them appear to be V- or
| |
| 5T-shaped; they are the bivalent chromosomes of the first maturation division.
| |
| Close around the germinal vesicle and extending out nearly to the periphery of the
| |
| egg is the yolk, which exists in the form of spherules, imbedded close together in
| |
| the granular cytoplasm. Finally there is the peripheral layer of deeply staining
| |
| protoplasm in which the test cells were formerly imbedded and which contains no
| |
| yolk, but numerous refractive spherules much smaller than those of the yolk.
| |
| | |
| In the living eggs of Cynthia this peripheral layer is clear and transparent and
| |
| contains uniformly but sparsely distributed yellow pigment, which seems to be associated with these small refractive spherules. This pigment is soluble in alcohol
| |
| and hence cannot be observed in fixed and prepared material ; on the other hand,
| |
| the alcohol in which large numbers of these eggs have been preserved, has the color
| |
| of a solution of potassium bichromate. The test cells of Cynthia also contain yellow pigment granules which are gathered close around the nuclei of these cells. It
| |
| is noticeable that most of the viscera of Cynthia contain this same yellow or orange
| |
| pigment, the ovaries being especially highly colored. This pigment is much denser
| |
| in some individuals than in others, and correspondingly one finds some ova in which
| |
| there is little or none of the pigment, while in others it is very abundant. In general the animals which have little of the pigment in their viscera are those which
| |
| produce eggs with little or no pigment, while those in which the viscera are deeply
| |
| pigmented produce well-pigmented eggs. The central yolk mass of the living egg
| |
| of Cynthia is of a slaty gray color, while the germinal vesicle is clear and transparent. Therefore, in the living egg of this species of ascidian, three areas can be
| |
| distinguished with great clearness before the maturation divisions begin, the
| |
| peripheral layer of protoplasm containing the yellow pigment, the central mass
| |
| of srrav volk and the clear germinal vesicle.
| |
| | |
| In Ciona and Molgula also these three areas are distinguishable in the living
| |
| egg before maturation, but not so clearly as in Cynthia. In Ciona the peripheral
| |
| layer is nearly transparent, the yolk is a brownish red, while the germinal vesicle
| |
| is also transparent. In Molgula both the peripheral layer and the germinal vesicle
| |
| are transparent, while the yolk is gray, with a faint lilac tinge. A brief inspection
| |
| of the eggs of Boltenia shows that in this genus the yolk is a bright red.
| |
| | |
| This peripheral layer of protoplasm, which is present in all the ascidian ova
| |
| which I have studied, is, both in living and in stained material, the most striking
| |
| feature of the egg before maturation and fertilization. It is surprising therefore
| |
| that in spite of this fact it has received so little attention from those who have
| |
| studied the ovarian history of the ascidian egg: in fact, with a single exception, I
| |
| cannot be sure that it has ever been mentioned by any previous writer on this subject. In his paper on the origin of the test cells, Morgan (1890) figures and describes this "peripheral /.one of protoplasm" in an unidentified species of Clavellina from Green Turtle Cay, Bahamas. What may perhaps be the same layer was described bj' Kowalevsky (1866) as a "gelatinous layer," in which the test cells are at
| |
| first imbedded ; later, about the time of the first cleavage, the test cells move to the
| |
| periphery of this layer. In these respects this gelatinous layer of Kowalevsky resembles the peripheral layer of protoplasm, but in other respects the differences
| |
| arc very great; for example, Kowalevsky says that this layer is formed by the
| |
| activity of the follicle cells, that it closely surrounds the blastomeres during cleavage, and that it becomes the gelatinous mantle of the adult, while the cells within
| |
| it (test cells) form the cells of the mantle. In none of these respects is this gelatinous layer like the peripheral layer of protoplasm described above, and I am in
| |
| doubt whether Kowalevsky actually saw this layer of protoplasm or whether he is
| |
| not describing the perivitelline space between the surface of the egg and the chorion.
| |
| It is interesting to note that Kowalevsky calls especial attention to the yellow color
| |
| of the test cells, though he nowhere indicates that he has seen any such yellow
| |
| pigment in the egg itself. Kupffer (1870), on the other hand, concluded that the
| |
| test cells were formed by free cell formation from the substance of the egg, because
| |
| " die Zellen gleich Anfangs genau die Farbe des Dotters haben' '; however, he
| |
| nowhere indicates that the peripheral layer of the egg differs in any way from the
| |
| remainder. In not one of the many later papers on the ascidian egg can I find anv
| |
| reference to this peripheral layer of protoplasm, except in that of Morgan already
| |
| referred to. In view of the ease with which it can be seen, both in living and in
| |
| stained material, and of the very important part which it takes in development,
| |
| this is most remarkable and inexplicable. If due attention had been given to this
| |
| feature of the ascidian egg, it is safe to say that some of the most conflicting accounts
| |
| of ascidian embryology would never have been written.
| |
| | |
| A peripheral layer of protoplasm, entirely similar in structure to that of the
| |
| ascidian egg, has been observed and described by Sobotta (1897) in the egg of Amphioxus. In this case Sobotta says that the peripheral layer gives rise in large part
| |
| to the inner egg membrane, which forms about the time of the maturation, but even
| |
| after the formation of this membrane a portion of this layer may remain at the
| |
| periphery of the egg. Sobotta speaks of the desirability of observing this layer in
| |
| the living egg, and from what I have seen in the ascidians I can but emphasize this
| |
| suggestion. In the ascidians this layer does not disappear with the formation of the
| |
| egg membrane, to which it contributes, but collects at the lower pole when the
| |
| egg is fertilized; from Sobotta's figures I judge that the same thing happens in
| |
| Amphioxus.
| |
| | |
| The colors of ascidian eggs deserve some notice at this place. In 1870
| |
| v. Kupffer observed in the living eggs of Ascidia canina {Ciona inlestinalis f) that
| |
| in the early stages of the ovarian egg the ooplasm is yellow ; later, as the egg ripens,
| |
| this color changes to a brownish red (Kupffer, 1870, p. 10). In the later stages of
| |
| development this red color is limited entirely to the walls of the alimentary tract.
| |
| In different animals and at different periods of the year Kupffer found that this
| |
| color varied from a bright red to an orange tone (p. 17).
| |
| | |
| | |
| | |
| Van Beneden and Julin (1884, pp. 4 and 5) call attention to the fact that Corella parallelogrannna produces two kinds of eggs, one yellow the other gray in
| |
| color. Both kinds of eggs develop normally and in the same manner and give rise
| |
| to larva', the endoderm cells of which are colored yellow or gray. Clavellina rissoana also produces two differently colored kinds of eggs, one pure rose the other
| |
| yellowish in tint. All eggs produced by the same individual have the same color.
| |
| | |
| In the species of Clavellina studied 1>\ Seeliger (1885) the color of the protoplasm surrounding the nuclei of the cleavage cells is yellow, as an inspection of his
| |
| figures of the living eggs shows (v. his plate I).
| |
| | |
| These cases, taken in conjunction with my observations on the eggs of Cynthia,
| |
| Ciona, Molgula and Bollema, show that the eggs of ascidians are frequently
| |
| colored; these colors are usually found in the yolk, and in the later development
| |
| pass into the endoderm cells. In Cynthia the peripheral layer is also colored, and
| |
| this fact leads me to hope that some other ascidian may be found in which still
| |
| other portions of the ooplasm may be differentially colored.
| |
| | |
| II. MATURATION AND FERTILIZATION.
| |
| | |
| These processes are so intimately associated in the ascidian egg that it is difficult and perhaps inadvisable to treat them entirely separately. As in so many
| |
| other eggs the entrance of the spermatozoon furnishes some stimulus to the egg
| |
| which leads to the completion of the maturation divisions. Without this stimulus
| |
| the egg may remain in the stage of the metaphase of the first polar spindle for
| |
| hours or even days.
| |
| | |
| A. Maturation.
| |
| | |
| 1. Disappearance of Nuclear Membrane.
| |
| | |
| The first steps in the formation of the polar spindle take .place before the
| |
| entrance of the spermatozoon. Almost as soon as the egg is laid, and sometimes
| |
| even before this, the wall of the germinal vesicle dissolves and the clear protoplasm
| |
| contained within the germinal vesicle moves up to the animal pole of the egg where
| |
| it may spread out into a cap or peripheral layer (Ciona), or may form merely a
| |
| somewhat flattened disk (Cynthia). As soon as the nuclear membrane has dissolved the chromosomes, nucleolus and a granular mass from which the spindle
| |
| fibres are formed gather together into the center of this area of nuclear protoplasm (figs. 62, 63, 77, 78) ; since the chromosomes lay at the periphery of the germinal vesicle before its membrane dissolved, this involves a considerable movement
| |
| on the part of these various constituents. No distinct linin network is visible
| |
| throughout the germinal vesicle, either before or after its membrane dissolves, and
| |
| the drawing together of these scattered elements into a central mass must be due
| |
| to something other than the contraction of the threads of such a network.
| |
| | |
| The chromosomes, when drawn together into a central mass, are connected by
| |
| a faintly staining, finely granular substance, which is much denser than the surrounding nuclear protoplasm. In the further development of the polar spindle this
| |
| mass gives rise to the spindle fibres, and from this fact, as well as from its staining
| |
| reactions, it may be identified with linin (figs. 62, 77).
| |
| | |
| | |
| The question as to the cause of the dissolution of the nuclear membrane is an
| |
| interesting one. In a recent work, R. Hertwig (1904) suggests that it is due to
| |
| the fact that the cytoplasm attacks the nucleus after the cell has ceased to grow.
| |
| From such observations as I have made 1 should be inclined to think that the
| |
| cause was a quite different one, viz., the continued growth of the nucleus at a
| |
| iin ire rapid rate than the cytoplasm. ' In most if not all cases the nuclear membrane dissolves only alter the nucleus has exceeded in volume a certain ratio to
| |
| the cell bodv. In the ascidian egg the germinal vesicle does not begin to dissolve
| |
| as soon as the egg ceases to grow ; on the other hand, therv is a considerable period
| |
| after the maximum size has been reached before the nuclear membrane disappears;
| |
| during this period the germinal vesicle continues to enlarge, the test cells are
| |
| extruded, the secretion which gives rise to the chorion is poured out, the entire
| |
| egg shrinks in volume, and finally the nuclear membrane grows very thin and disappears. This process is in no wise complicated by the presence of a centrosome,
| |
| since, according to my observations, no centrosomes are present at any stage of the
| |
| maturation divisions.
| |
| | |
| 2. Chromosomes.
| |
| Even before the wall of the germinal vesicle dissolves the chromosomes may
| |
| be distinguished as small deeply staining bodies, some of which at least are Vor Y-shaped (fig. 70). They are small and numerous, and I have not been
| |
| able to count them with any assurance of accuracy. After they have been drawn
| |
| together into the center of the nuclear area, as described above, they become a little
| |
| larger and are plainly V-shaped (tigs. 62, 63, 77). When the spindle fibres appear
| |
| they are at first widely scattered on or among these fibres (figs. 63, 79, SO), and
| |
| only in the metaphase do they become arranged in an equatorial plate (fig. 66).
| |
| In the splitting of the chromosomes the daughter halves first separate at the apex
| |
| of the V and remain longest connected together by the two limbs ; this double V,
| |
| with the apices pointing to the two poles of the spindle, is then stretched out
| |
| until the two limbs of each V come to lie near together, thus forming a double Y,
| |
| each with a long stem pointing to opposite poles; even the little space between the
| |
| limbs of the Y may disappear, thus forming cross-shaped chromosomes (fig. 66).
| |
| After the daughter chromosomes have separated they are plainly V-shaped (figs. 67,
| |
| 68) ; and this shape may also be seen in the first polar body and in the second
| |
| polar spindle (figs. 68, 69). In the second polar spindle each limb of the V is separated from the other, thus giving rise to rod-shaped chromosomes, which are found
| |
| in all the stages of the anaphase and in the second polar body (figs. 70-73).
| |
| Owing to the small size of the chromosomes it has not been possible to determine with certainty which of these maturation divisions is reducing and which
| |
| equational. If the two limbs of the V's in the first maturation represent two individual chromosomes united at one end, then the first maturation division is equational and the second reducing, for these limbs of the V's are not separated until
| |
| the second maturation ; if, on the other hand, the cleft in the original V's represents
| |
| the splitting of two original chromosomes placed side by side (a thing which seems likely, since such parallel chromosomes without any cleft arc found in early stages
| |
| ((-/. figs. iti. 62), then the first maturation division would separate whole chromosomes and hence be reducing, while the second would separate half chromosomes
| |
| and therefore be equational. Only a careful investigation of the manner of origin
| |
| of these \ -shaped chromosome- would finally solve this problem, and this material
| |
| is unfavorable for such work.
| |
| | |
| Nucleolus.
| |
| The nucleolus of the germinal vesicle is large and is frequently vacuolated;
| |
| | |
| it usually lies eccentrically in the germinal vesicle, though its position hears no
| |
| constant relation to the polarity of the egg. As is usually the ease, it begins
| |
| to dissolve at the same time that the nuclear membrane does, and it disappears, with great rapidity, so that no trace of it is left by the time that the first
| |
| maturation figure has reached the metaphase. In this respect it differs materially
| |
| from the nucleolus of many other eggs, where its solution is so slow that it may
| |
| not disappear until late in the first maturation division. In this case the solution
| |
| of the nucleolus is hastened by its breaking up into many small fragments (figs.
| |
| 62, 63, nl.).
| |
| | |
| 4. Spindle Formation.
| |
| My observations agree entirely with those of Boveri ( lS'.ldi. .1 ulin dS'JMi. Hill
| |
| (1895), Castle (1896) and Crampton 1 in showing that there is no trace of a centrosome at either pole of either of the maturation spindles at any time in their historv. These results are directly opposed to those of Golski (1809). who found
| |
| minute centrosomes at the poles of the maturation spindles of Ciona intestinalis. Not only are no centrosomes visible in my preparations at the poles of the
| |
| spindle, but all evidences of astral radiations are also absent. Under these circumstances the formation of the spindle is of unusual interest. The spindle fibres first
| |
| appear as lines of granules, which radiate in all directions from the finely granular
| |
| mass of linin substance which unites the chromosomes in the middle of the nuclear
| |
| area (fig. 62). These lines of granules are quickly transformed into fibres which
| |
| run through the linin substance ; these fibres are never parallel at their first appearance and frequently radiate in all directions, though they sometimes run in the
| |
| same general course (figs. 63, 64). As this mitotic figure with the surroundingnuclear plasm is moved nearer and nearer to the surface of the egg the fibres come
| |
| to be more nearly parallel, becoming paratangential with the egg surface (fig. 65).
| |
| In this rearrangement of the fibres they are at first farthesl apart at the ends, so
| |
| that the spindle has an hour-glass shape (fig. 65). Then certain of these fibres
| |
| unite at their ends into several groups or bundles, but the fibres which belong to
| |
| one group at one pole may be associated with different fibres at the opposite pole
| |
| (fig. CO). There is thus formed a kind of multipolar spindle, closely resembling
| |
| the mitotic figure present in many plants [cf. Osterhout, 1897: Mottier, 1897:
| |
| | |
| 1 I am indebted to Dr. Crampton for the privilege of seeing photographs of the beautiful pktes
| |
| of hi- completed but yet unpublished work on the maturation and fertilization of Molgula.
| |
| | |
| | |
| Nemec, 1899). Finally, in the metaphase all the spindle fibres are drawn together
| |
| at the poles; but even in this stage, though the spindle fibres lie close together,
| |
| they do not unite into a central body, and there are no astral rays (fig. 66). In
| |
| the anaphase a few rays may be seen running from the margin of the chromatic
| |
| plate toward the equator of the cell and lying on the periphery of the spindle (figs.
| |
| 67, 81, 82). These are the only fibres which are not continuous from pole to pole,
| |
| and are therefore the only ones which bear even a remote resemblance to astral rays ;
| |
| that they are not such, however, is shown by the fact that they radiate from no
| |
| center but lie only around the periphery of the spindle. In this respect they more
| |
| closely resemble peripheral spindle fibres than astral rays. Still, if one considers
| |
| that one of the characteristics of peripheral spindle fibres is that they are attached
| |
| to chromosomes, it will be seen that these fibres do not belong in that category. Of
| |
| course, since centrosomes are not present, there can be no central spindle. We have
| |
| in this case, therefore, a mitotic figure in which are neither central spindle, peripheral spindle nor astral rays in the strict significance of those terms. The spindle
| |
| which is present arises wholly from nuclear linin, and consists almost exclusively of
| |
| fibres which are continuous from pole to pole.
| |
| | |
| The small size of the maturation spindles of the ascidian egg is notable as contrasted with the great size of the germinal vesicle. Among many annelids and mollusks the first maturation spindle is at least as long as the diameter of the germinal
| |
| vesicle, whereas among the ascidians it is scarcely more than one-quarter as long.
| |
| However, in those animals in which the spindle is very long in the prophase or
| |
| metaphase it undergoes a great shortening in the anaphase, e. g., in Crepidula it
| |
| is not more than half as long in the anaphase as in the metaphase (Conklin, 1902).
| |
| This is probably true of all cases in which the maturation spindle is a large one ;
| |
| for, since division of the cell body regularly takes place through the equator of the
| |
| spindle, the spindle must be relatively short at the time of the division of the cell
| |
| body, or the polar body will be relatively large. In all those cases in which the
| |
| first polar spindle is a long one, centrosomes are present near the periphery of the
| |
| germinal vesicle before its membrane disappears and the loose linin network of the
| |
| nucleus is transformed into the spindle fibres, thus forming a large, loosely constructed spindle. Later, by contraction of these fibres, the spindle shrinks in all
| |
| dimensions. In the ascidians, on the other hand, no centrosomes are present and
| |
| the shrinkage of the linin takes place before the spindle is formed, so that from the
| |
| first it occupies but a small part of the volume of the germinal vesicle, and is no
| |
| larger at the beginning of mitosis than at its close.
| |
| | |
| The second maturation spindle arises in part at least from the remains of the
| |
| first, and is about half as large. Here also there is no trace of centrosomes or astral
| |
| radiations at any stage. The spindle is barrel-shaped, and a few peripheral fibres
| |
| are found around it in the anaphase (figs. 69-72) ; in all respects it closely resembles the first maturation spindle.
| |
| | |
| Such a case of mitosis as this, in which we have the formation of a spindle, the
| |
| separation of chromosomes and the division of the cell body entirely without the
| |
| presence of centrosomes, offers a valuable opportunity for the study of the mechanics of indirect cell division. Inasmuch as sonic of the cleavages also throw lighl on
| |
| this problem, its further consideration will be postponed to the section which deals
| |
| with the first clca\ age.
| |
| | |
| 5. Movements of Spindle and of Nuclear Plasm ; formation of Polar Bodies.
| |
| | |
| In Cynthia the tirst maturation spindle and the surrounding nuclear plasm remain indefinitely in the condition shown in figures 77 and 78 unless the egg be fertilized. In Ctona the stage at which the maturation processes come to rest is a little
| |
| more advanced than in Cynthia, as is shown in figure 17li: the peripheral layer of
| |
| protoplasm is here collected over the lower hemisphere of the egg, and the nuclear
| |
| plasm which has escaped from the germinal vesicle forms a layer over the entire
| |
| upper hemisphere. Unfertilized eggs may remain in this condition for at least
| |
| three or four hours and still he capable of fertilization and normal development ;
| |
| Imt if they remain unfertilized for ten or twelve hours the nuclear plasm spreads through the substance of the yolk in irregular masses {cf. text lig. I), and the eggs
| |
| thereafter are not capable of normal fertilization. The maturation and further
| |
| development of the egg are finally and forever halted in this early stage unless
| |
| the eo-g be fertilized. As soon as a spermatozoon enters the egg active movements of the protoplasm begin and a localization of different ooplasmic materials
| |
| occurs, which will be described later; at the same time the fust maturation spindle
| |
| moves to the animal pole and is turned from a paratangential to a nearly radial
| |
| position. The daughter chromosomes then separate and the first polar body is
| |
| extruded (figs. 66-68 and 79-82).
| |
| | |
| | |
| | |
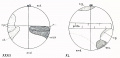
| Fig. I. Section of an egg of Cynthia partita which had lain twelve hours without being fertilized.
| |
| The first polar spindle (1 p. -. lies in tin- position in which it was first formed ; the peripheral layer
| |
| of yellow protoplasm ip. l.i remains uniformly distributed over the surface, hut the clear protoplasm
| |
| has spread throughout the yolk and broken it up into irregular masses (compare with figs. 77 and 78
| |
| showing unfertilized est:s in normal condition).
| |
| | |
| Fig. II. Stained preparation of an entire egg of Cynthia partita, showing small spindles at opposite poles (1. p. s.i. which are possibly two first maturation spindles, though more probably one of these
| |
| is a precociously developed sperm spindle.
| |
| | |
| | |
| | |
| The second maturation spindle is smaller than the first, as Castle has shown.
| |
| and. like the first, is paratangential in position in early stages and only later
| |
| becomes radial. The second polar bod) is extruded close to or immediately under the first (figs. 71-73). The two polar bodies are of approximately the same size,
| |
| and neither ever divides. They are at first composed of clear protoplasm in which
| |
| the chromosomes are free; later the chromatin is dissolved and diffused throughout
| |
| the cell body, so that they stain deeply and uniformly. They may at all times be
| |
| distinguished from the test cells by this staining reaction as well as by their being
| |
| closely attached to or imbedded in the egg. In Ciona they may further be distinguished from the test cells by the fact that they are larger than the latter. In
| |
| many eggs of Cynthia and in almost all of Ciona the polar bodies remain attached
| |
| to or imbedded in the egg at the point of their formation, and they thus constitute
| |
| a most important landmark.
| |
| | |
| B. Fertilization.
| |
| | |
| As has been said, the first maturation spindle remains in the metaphase until
| |
| the egg is fertilized. The egg remains capable of fertilization for three or four
| |
| hours at least after the first formation of this spindle. As Castle (1896) has
| |
| shown, self-fertilization rarely if ever occurs in Ciona, though artificial cross-fertilization is most easily accomplished. In Cynthia, on the other hand, artificial cross-fertilization is successful in only a small proportion of the eggs.
| |
| | |
| I have so far been unable to find any artificial means which will cause the
| |
| unfertilized eggs to develop beyond the metaphase of the first maturation division.
| |
| Violent shaking, various degrees of concentration or dilution of sea water, solutions
| |
| of sodium or magnesium chloride of varying strengths have all been without effect
| |
| in this regard. My experience in this matter is similar to that of Lyon (1903). who
| |
| reports that he was unable to cause parthenogenetic development among ascidians
| |
| at Naples by any artificial means.
| |
| | |
| 1. Entrance of Spermatozoon.
| |
| | |
| Of the multitudes of spermatozoa which may be seen burrowing between the
| |
| follicle cells outside of the chorion after spermatozoa have been mixed with the
| |
| ova, only a few ever pass through that membrane. I have never seen a spermatozoon in process of passing through the chorion and do not know how it is
| |
| accomplished. It is possible that there are one or more micropyles at the lower
| |
| pole, though I have never seen them. In whatever manner the spermatozoa pass
| |
| the chorion it is done very quickly and several frequently enter the perivitelline
| |
| space; dispermy or polyspermy, however, is very unusual. A spermatozoon enters
| |
| the egg in from two to five minutes after the spermatozoa are mixed with the ova,
| |
| and the presence of supernumerary spermatozoa in the perivitelline space is shown
| |
| by the fact that some of the test cells are occasionally fertilized (figs. SO, 85, sn).
| |
| | |
| The spermatozoon always enters the egg near the vegetal pole. I have not
| |
| found it possible to determine in living eggs whether the point of entrance lies
| |
| exactly at the vegetal pole or a little to one side of this. In stained preparations
| |
| of entire eggs, as well as in sections, the entering spermatozoon is usually seen to
| |
| lie eccentrically with reference to the vegetal pole (figs. 79, 173). In other cases, however, it lies almost exactly at that pole; in sections this appearance might
| |
| be due to obliquity of the plane of section to the egg axis, but in preparations of
| |
| entire eggs it can he seen that the spermatozoon does sometimes enter almost exactly at the vegetal pole. It is unquestionably true that the point of entrance is
| |
| usually eccentric as Castle affirms, hut the degree of eccentricity just as certainly
| |
| varies in different cases. It might he supposed that this eccentricity always lay in
| |
| a single dehnite meridian, were it not for the fact that in cases of dispermy and
| |
| polyspermy the various points of entrance lie in different meridians {cf. figs. 12, 94).
| |
| I conclude therefore that the spermatozoon may enter at any point on the vegetal
| |
| hemisphere within about 30 of the pole.
| |
| | |
| The fact that the spermatozoon always enters near the vegetal pole must be
| |
| due to some structural peculiarity ; the peripheral layer of protoplasm is a little
| |
| thicker at this pole than elsewhere at the time that the sperm enters, and this might
| |
| be held to be the cause of the sperm's entering at this pole, were it not for the fact
| |
| that the sperm enters at the vegetal pole in many other eggs, e.g. those of annelids
| |
| and mollusks, in which there is no periperal layer. It is probable that this very
| |
| general phenomenon is dependent upon some fundamental property, such as the
| |
| polarity of the egg or the direction of movement of the egg substance.
| |
| | |
| 2. Movements of Ooplasm.
| |
| | |
| With the entrance of the sperm the most astonishing series of changes takes
| |
| place in the egg. These changes are most striking in the living eggs of Cynthia,
| |
| where, owing to the yellow color of the peripheral protoplasm, the movements of
| |
| the egg substance can be directly observed ; but they may also be seen in the living
| |
| eggs of Ciona, and a detailed study of these changes may be made on fixed and
| |
| stained preparations. Almost immediately after the entrance of the spermatozoon the peripheral layer of protoplasm, which is nearly uniformly thick, and the
| |
| great area of nuclear plasm, in which the first maturation spindle lies (figs. 77, 78),
| |
| How around to the lower pole of the egg, leaving the first maturation spindle surrounded by only a small amount of protoplasm. Thus within some ten minutes
| |
| after the entrance of the sperm the protoplasmic pole of the egg is transformed into
| |
| the yolk pole and vice versa. Castle does not figure nor describe this flowing of the
| |
| protoplasm from the animal to the vegetal pole, and it is probably owing to the fact
| |
| that he had not observed the early stages in which this occurs that he describes the
| |
| polar bodies as being formed at the yolk pole of the egg and the spermatozoon as
| |
| entering at the protoplasmic pole. Although he says that the presence of a spermatozoon cannot be detected in the egg from which his figure 1 is drawn, I should suppose from the fact that the first polar body is being extruded that the sperm must
| |
| already have entered {cf. my fig. 173).
| |
| | |
| a. Localization of Yellow Protoplasm.
| |
| | |
| In Cynthia this downflow of protoplasm takes place so rapidly that it can
| |
| be seen in the living egg and with such force that the test cells, which lie between the surface of the egg and the chorion, are sometimes carried down with the streaming protoplasm to the lower pole of the egg, where they are crowded together and
| |
| heaped up in the perivitelline space (fig. 3, et seq.). While this flowing is most
| |
| active, streamers of yellow surface protoplasm may be seen radiating toward the
| |
| upper pole. The yellow protoplasm thus carried to the lower pole collects into a
| |
| deep orange-yellow spot which surrounds the sperm nucleus (figs. 4-6); it frequently
| |
| forms a prominence at the lower pole which recalls the polar lobe of the eggs of
| |
| annelids and mollusks. The clear nuclear protoplasm also flows to the lower pole,
| |
| where it lies beneath the yellow disk or spot and is visible around its periphery (figs.
| |
| 4-6). The yellow protoplasm then gradually spreads again until it covers most of
| |
| the lower hemisphere (figs. 6-10). Then the sperm nucleus moves to one side of this
| |
| yellow cap, and a large part of the yellow protoplasm is drawn over with it until it
| |
| forms a yellow band or crescent, in the middle of which the sperm nucleus lies.
| |
| This crescent lies just below the equator of the egg and its middle point marks
| |
| the posterior pole of the future embyro. while its two horns reach forward about
| |
| half-way around the egg to the middle of the right and left sides.
| |
| | |
| b. Localisation of Clear Protoplasm and Yolk.
| |
| | |
| At the same time that the yellow protoplasm is being formed into a crescent
| |
| and moved up toward the equator on the posterior side of the egg, the clear protoplasm which surrounds the sperm nucleus and aster is also drawn entirely away
| |
| from the lower pole to the posterior side of the egg and thence up to the equator
| |
| (figs. 82-92). Up to this time the sperm nucleus and the clear and yellow protoplasm have remained near to the egg surface ; finally, after the meeting of the germ
| |
| nuclei near the posterior pole of the egg, these nuclei and the clear protoplasm surrounding them move inward to the center of the egg, while the yellow protoplasm
| |
| is largely left at the surface.
| |
| | |
| When the clear and yellow protoplasm are withdrawn from the upper pole the
| |
| gray yolk is there exposed (figs. 4, 5, 11). After the protoplasm moves up to the
| |
| posterior pole the yolk is exposed over the entire egg, except for the area of the
| |
| yellow crescent and a narrow line of clear protoplasm, which comes to the surface
| |
| just above the crescent (figs. 13-18).
| |
| | |
| In sections, small spherules which probably represent the yellow granules
| |
| of the peripheral layer of protoplasm, may be seen heaped up around the entering
| |
| sperm (fig. 74), this aggregation corresponding to the yellow spot of the living egg
| |
| (fig. 6). This massing of the yellow spherules is most marked, while the sperm
| |
| head lies in the peripheral layer ; when it passes through this layer into the deeper
| |
| layer of clear protoplasm the yellow spherules again spread out into a flattened disk,
| |
| as shown in figures 75 and 80, which correspond to figures 7 and 8 of the livingegg (Plate I). Later, when the sperm nucleus moves to the posterior pole and the
| |
| yellow protoplasm is drawn over to that side to form the crescent, sections show
| |
| that this crescent does not lie entirely on the surface, but that it extends for some
| |
| distance inward toward the sperm nucleus (figs. 87, 90, 92).
| |
| | |
| | |
| In Ciona the same type of protoplasmic movement occurs as in t ynthia, 1 >ut
| |
| with certain minor differences. The peripheral layer is here decidedly thicker at
| |
| the lower pole than elsewhere, even before the fertilization of the egg; the nuclear
| |
| plasm or clear protoplasm is also at this stage distributed as a layer over the entire
| |
| upper hemisphere of the egg (fig. 172). After the entrance of the spermatozoon
| |
| the protoplasm of both these layers collects at the lower pole. The nuclear plasm
| |
| and peripheral protoplasm cannot easily be distinguished in living eggs of Ciona, but
| |
| in fixed and stained material the latter stains more deeply than the former (figs.
| |
| 172. 173). A crescent of peripheral protoplasm is formed here in the same way as
| |
| in Cynthia (figs. 175, 170). and it occupies the same relative position (figs. 179
| |
| 183). Though Castle did not observe the peripheral layer of protoplasm and its
| |
| movement to the lower hemisphere it is evident that he recognized at least a part
| |
| of the crescent. His figures 17 and 45-47 show the middle portion of the crescent in the 2-8 cell stages, and he describes this as an area of finely granular protoplasm, which is clear in the living egg, and out of which the small posterior
| |
| mesenchyme cells are formed. According to my observations these cells arise from
| |
| a small part only of the middle portion of this crescent, while the greater part of
| |
| the crescent gives rise to the muscle and mesenchyme cells of the tadpole. From
| |
| his figures, as well as his descriptions, it is evident that he recognized only a small
| |
| portion of the crescent, viz.. this median area of "clear protoplasm."
| |
| | |
| In many eggs of Ciona, if not in all, clear protoplasm, which is composed of
| |
| large alveoles, surrounds the entering spermatozoon (fig. 173). Later, when the
| |
| sperm nucleus moves to the posterior pole, this clear area moves with it. and in sections in the median plane (figs. 175, 170) forms a clear triangular area in the middle
| |
| of the deeply staining crescent. There is here shown a marked differentiation of the
| |
| substance of the crescent which continues to lie recognizable throughout most of the
| |
| cleavage. I have not observed this clear median portion of the crescent in the 4-cell
| |
| stage, but in the S-cell stage and thereafter it is plainly visible as a deeply staining
| |
| cap of protoplasm on each side of the mid-line. It corresponds in the main to the
| |
| 'clear protoplasm" described by Castle, which, as he discovered, marks the position of the sperm nucleus at the posterior pole and which ultimately gives rise to
| |
| the '-small posterior mesenchyme" cells (B 7 ") at the posterior pole of the gastrula.
| |
| This same clear protoplasm is present in the middle of the crescent in the Cynthia
| |
| egg, although it is here obscured by the surrounding yellow pigment; in the unsegmented e^/ it forms a layer of transparent protoplasm over the surface of the
| |
| crescent, and in the cleavage stages of prepared eggs it is visible as two deeply staining caps of protoplasm similar to those in the egg of Ciona ; it ultimately gives rise
| |
| to the small posterior mesenchyme cells which are formed from the middle of the
| |
| crescent and which are composed of clear protoplasm in which there is no yellow
| |
| pigment (fig. 48. inch.). The substance of the crescent is therefore plainly differentiated from the first into these two substances, clear and yellow protoplasm, which remain distinct throughout the entire development.
| |
| | |
| | |
| | |
| 3. Development of Sperm Nucleus and Aster.
| |
| Immediately after it has entered the egg the sperm head is rod-shaped and is
| |
| | |
| frequently coiled or twisted on itself (figs. 74, 79, 173). It decreases in length and
| |
| increases in width very quickly, and soon appears pear-shaped, the pointed end
| |
| being directed toward the sperm aster. At first densely staining throughout, it stains
| |
| less and less* densely as it swells in volume, until finally the chromatic and achromatic constituents are easily distinguishable (figs. 80-87). During this process there
| |
| are no evidences of chromosomal vesicles, the nucleus constituting a single vesicle.
| |
| In some cases there is a faint line between the head of the sperm and the egg
| |
| membrane which represents the middle piece and perhaps a portion of the tail (figs.
| |
| 74. 79). Very soon after the spermatozoon has entered the egg a small aster,
| |
| with central clear area and minute rays, appears in the position of the middle
| |
| piece, between the sperm head and egg membrane (figs. 80, 173). The sperm aster
| |
| then grows with great rapidity, the rays extend throughout the greater part of the
| |
| clear protoplasm and even into the yolk and a minute body, the centrosome, becomes
| |
| visible at the centre of the rays, while the whole aster stains moi'e deeply that the
| |
| surrounding protoplasm (figs. 81-87).
| |
| | |
| | |
| 4. Path of the Spermatozoon within the Egg.
| |
| | |
| | |
| The spermatozoon usually enters the egg in a radial direction and keeps right
| |
| on through the protoplasm at the lower pole until it reaches the deeper lying yolk
| |
| (figs. 74, 75, 80). This may be known as the penetration path (Roux). The sperm
| |
| nucleus and aster then rotate so that the aster is directed forward in all further
| |
| movements, as is true in so many other cases (figs. 80-83). The path described
| |
| after the rotation is the copulation path (Roux), and it always forms more or less
| |
| of an angle with the penetration path. While the penetration path may apparently
| |
| lie in any portion of the lower hemisphere within about 30 of the pole, the copulation path seems to be definitely determined by the structure of the egg. The sperm
| |
| nucleus and aster move in this path from the neighborhood of the lower pole up to
| |
| the equator of the egg on the posterior side, all the time keeping near to the surface
| |
| of the egg (figs. 81-87). But this path is not always the shortest path to the
| |
| equator; sometimes it is the longest, as in figures 81 and 85, in which the sperm
| |
| having entered to the left of the lower pole moves across to the right side in the
| |
| figure and then up to the equator. The point near the equator to which the sperm
| |
| nucleus moves invariably marks the posterior pole of the egg and of the future embryo, and the copulation path by which the sperm nucleus reaches this posterior pole
| |
| must lie along the posterior side of the egg ; but since the point of entrance of the
| |
| sperm and the penetration path may lie near to or far from the posterior side, it is
| |
| evident that they can have nothing to do in determining the position of the posterior
| |
| pole. And since the copulation path is not always the shortest path to the equator,
| |
| but may sometimes be the longest, it seems probable that the direction of the copulation path is not the cause but the result of the antero-posterior differentiation of
| |
| the egg. A further consideration of this subject will be found in the general part
| |
| of this paper.
| |
| | |
| | |
| 5. The Egg Nucleus and its Movements.
| |
| | |
| | |
| After the formation of the second polar bodv the chromosomes left in the egg
| |
| form chromosomal vesicles which then unite to form the egg nucleus (fig. 73). The
| |
| latter then moves away from the animal pole into the yolk, apparently in the direction of the axis of the second polar spindle (figs. 86, 87); it soon turns, however,
| |
| and moves toward the sperm nucleus and aster at the posterior pole. A't first a few
| |
| remnants of spindle fibres connect the egg nucleus with the animal pole (fig. 87),
| |
| but these are soon lost and thereafter this nucleus, without any surrounding area
| |
| of protoplasm or astral rays, is almost lost to view in the dense mass of yolk (fig.
| |
| 89). Finally the egg nucleus emerges from this yolk into the clear protoplasm
| |
| surrounding the sperm nucleus, and the two nuclei meet at the equator of the egg
| |
| about half way between the posterior pole and the center. The relative positions
| |
| of the two germ nuclei when they first meet is invariably the same ; the egg
| |
| nucleus always lying on the central (anterior) and animal pole (ventral) side of the
| |
| sperm nucleus (figs. 89-93).
| |
| | |
| 6. Sperm Amphiaster and first Cleavage Spindle.
| |
| | |
| | |
| About the time that the sperm nucleus has moved to the edge of the yellow cap
| |
| (fig. 8) and some time before the union of the two germ nuclei, the sperm centrosome divides as shown in figures 88, 89. 1 have not observed all the details
| |
| of this division, but it is evident that the centrosome here gives rise to a centrosomal spindle or netrum (Boveri L901 ). at the poles of which the daughter centrosomes lie. After the centrosome has divided, the sphere also divides (fig. 88), and
| |
| a well-marked central spindle is left connecting the two daughter centrosomes (fig.
| |
| 89). When these daughter centrosomes have moved to the poles of the sperm
| |
| nucleus the central spindle is curved around that nucleus, and finally its fibres
| |
| become indistinct (fig. 90) and then disappear altogether (fig. 91 ). The sperm aster,
| |
| at the time of its division, invariably lies on the central side of the sperm nucleus,
| |
| and the axis of the amphiaster thus formed is at right angles to the copulation path
| |
| and to the plane of the first cleavage (figs. 88-91). Up to the time when the
| |
| two germ nuclei meet, the sperm centrosomes lie at the poles of the sperm nucleus
| |
| (fig. 91), whereas no trace of centrosomes are ever found in connection with the egg
| |
| nucleus, and after the latter has moved away from the- animal pole, on its path to
| |
| the sperm nucleus, no trace of radiations, spindle fibres or even of surrounding
| |
| cytoplasm can he found near the egg nucleus. The cleavage centrosomes may be
| |
| traced without a break back to the sperm amphiaster, to the sperm aster and
| |
| finally to the middle piece of the spermatozoon. There could not possibly be a
| |
| clearer ease of the origin of the cleavage centrosomes from the middle piece of
| |
| the spermatozoon than is presented by the ascidian egg. This conclusion agrees
| |
| with the work of all who have ever studied the fertilization in these eggs (Boveri
| |
| 1890, Julin 1893, Hill 1895, Castle L896, Golski 1899, Crampton). In Ciona,
| |
| Castle observed an archoplasmic mass in connection with each of the germ nuclei,
| |
| though that found near the sperm nucleus was larger and more energetic than the one near the egg nucleus; the latter afterward completely degenerates, according
| |
| to Castle, and takes no part whatever in the formation of the lirst cleavage spindle.
| |
| I have been unable to find tins archoplasmic mass in connection with the ega;
| |
| nucleus unless the remnants of the second polar spindle (fig. 87) maybe interpreted
| |
| as such.
| |
| | |
| Any one \vli<> has studied the method of origin of the cleavage centrosomes in
| |
| the eggs of ascidians and of mollusks cannot fail to be impressed with the profound
| |
| differences between the two. In the one we have no centrosome or aster in connection with the egg nucleus at any stage, while the sperm centrosome and aster are
| |
| visible at all stages alter the entrance of the spermatozoon, and give rise directly
| |
| to the cleavage centrosomes; in the other, according to my observations, a centrosome and aster are found in connection with each of the germ nuclei, and coincidently with the union of these nuclei the asters or spheres also unite, while out of
| |
| this fused sphere material a single centrosome arises in connection with each genu
| |
| nucleus. It is recommended to those who maintain that in these details of fertilization all animals must conform to a single type that they study the fertilization of a gasteropod as compared with that of an ascidian.
| |
| | |
| | |
| 7. Dispermy.
| |
| | |
| | |
| Although it is a relatively rare thing for more than one spermatozoon to enter
| |
| an egg. still v^^ are occasionally found into which two spermatozoa have- penetrated. The entrance of more than two spermatozoa, if it occurs at all. must be a
| |
| very rare phenomenon. In stained preparations and in serial sections I have never
| |
| seen an undoubted case of it; unsegmented eggs are sometimes found in which
| |
| there are a number of nuclei, but in all cases it is possible that these may have
| |
| arisen from the division of two sperm nuclei. In living eggs I have sometimes
| |
| observed several yellow spots on the lower hemisphere. Such an egg is shown in
| |
| figure 12; there are here four yellow spots, each about equidistant from the vegetal
| |
| pole, and presumably there is a nucleus in each of these, though nuclei were actually
| |
| observed in only two of them. It is possible that these may have arisen by division from two original nuclei, and that this is therefore a case of dispermy and not
| |
| of polyspermy. It is an interesting fact that dispermic eggs never divide, though
| |
| the nuclei may do so repeatedly, and of course they never develop normally.
| |
| | |
| Dispermic eggs have been repeatedly observed both in living and in fixed
| |
| material, in entire preparations and in serial sections. Such eggs afford a valuable
| |
| means of testing the question as to whether the point of entrance of the sperm is
| |
| predetermined, and more important still, as to whether the posterior pole of the
| |
| egg and the plane of the first cleavage is pre-existent in the egg or is established
| |
| by the entering spermatozoon. So far as I have observed, (lie two sperm nuclei
| |
| always enter the egg near the vegetal pole, and at lirst they lie in a common protoplasmic field. As they move toward the equator, however, they frequently separate, and when they have reached the equator and have each given rise to a spindle they are often found on opposite sides of the egg with the surrounding protoplasmic fields quite separate. The two spindles are usually parallel to each other
| |
| and are always entirely independent, the pules never being united into a triaster
| |
| or tetraster.
| |
| | |
| Sections of two dispermic eggs are shown in figures 94 and 95; in the former
| |
| the sperm nuclei, which have not yel reached the equator of the egg, occupy symmetrical positions on each side of the mid-line, and the protoplasmic field in which the}
| |
| lie is located on that side of the egg which corresponds to the posterior pole of nor
| |
| | |
| | |
| | |
| | |
| | |
| | |
| FlGS. III-VI. Dispermic eggs of ('. partita; drawn from stained preparations of entire
| |
| eggs. Figs. III. and V are viewed from the vegetal pole, the polar bodies being seen
| |
| through the egg. Fig. IV is viewed from the animal pole and Fig. VI from the posterior
| |
| pole. The boundary between the protoplasm and yolk is indicated by a crenated line;
| |
| when seen through the egg this boundary is represented by a line of stipples.
| |
| | |
| mal eggs. The yellow protoplasm here forms a continuous crescent, and save for the
| |
| fact that the sperm nuclei do not lie at the middle of this crescent and that a small
| |
| tongue of yolk partly separates the two sperm asters, the egg is not unlike a normal
| |
| one. In figure 95 a later stage of a dispermic egg is shown, in which the sperm nuclei
| |
| have reached the equator and have moved in from the surface toward the center of
| |
| the egg, while one of these nuclei has united with the single egg nucleus. There
| |
| is here also a symmetrical arrangement of the sperm nuclei and of the (dear and yellow protoplasm on each side of the mid-line. The protoplasmic areas are here further separated than in the preceding figure, though they still lie nearer one pole of the
| |
| egg than the other. In this case also there can be little doubt that the more richly
| |
| protoplasmic pole corresponds to the posterior pole of normal eggs while the yolkladen pole corresponds to the anterior one.
| |
| | |
| Other dispermic eggs arc shown in text figures III-VI, and here also one hemisphere of the egg contains more protoplasm than the other, and may probably be identified with the posterior pole. In still other eggs, especially those in which the
| |
| cleavage spindles are fully formed, the spindle and protoplasmic fields may lie on
| |
| opposite sides of the egg [cf. text fig. V). In these cases neither pole can be certainly identified as anterior or posterior. In normal eggs the cleavage spindle
| |
| always stands at right angles to the chief axis of the egg and to the plane of the
| |
| first cleavage; in dispermic eggs the spindles are frequently not at right angles to
| |
| the egg axis and if. as 1 believe, the plane between the two protoplasmic areas represents the normally median plane, they are more frequently parallel with this
| |
| plane than perpendicular to it.
| |
| | |
| The phenomena of dispermy demonstrate that the point of entrance of the
| |
| spermatozoon is not predetermined but that spermatozoa may enter at different
| |
| points on the vegetal hemisphere; they also render probable the view that the
| |
| plane of bilateral symmetry is not first established by the accidental path of the
| |
| spermatozoon within the egg, but that this plane is structurally present before fertilization. This problem will be more fully discussed in the general part of this paper
| |
| (Chap. VII).
| |
| | |
| | |
| III. ORIENTATION OF EGG AND EMBRYO.
| |
| | |
| As a preparation to the study of the cell-lineage and later development of the
| |
| ascidian vixix it is necessary to consider at once the orientation of the egg and early
| |
| cleavage stages. This is the more necessary since the utmost possible diversity of
| |
| opinion has been expressed with regard to this matter.
| |
| | |
| 1. Van Beneden and Jul Ms System of Orientation.
| |
| | |
| | |
| Van Beneden and Julin (1884) were the first to undertake to relate the
| |
| early stages of development of the ascidian egg to the later stages. Their work
| |
| was in fact one of the earliest and most admirable contributions to the subject
| |
| of cell-lineage. They followed the cleavage, cell by cell, as far as the 44-cell stage
| |
| and pointed out what they supposed to be the relations of each of these cells to the
| |
| germ Layers. They determined the relations of the axes of the egg and early cleavage stages to those of the gastrula and larva and, for the first time in the history
| |
| of embryology, established the fact that the principal axes of the larva may be
| |
| identified in the nnseginented egg. The evidence's upon which they based their
| |
| conclusions as to the axial relations of egg and embryo and as to the fate of the
| |
| cleavage cells are not fully stated in their brief paper of only fifteen pages; but
| |
| their statements of fact are perfectly clear and explicit. In brief these are as
| |
| follows :
| |
| | |
| (1) The (list cleavage spindle is eccentric toward the posterior pule of the egg,
| |
| and the median plane of the future embryo is marked out by the bilateral symmetry
| |
| of the unsegmented egg (p. ti).
| |
| | |
| (2) The plane of the lirst cleavage coincides with the plane of bilateral symmetry, and therefore divides the egg into right and left halves (p. G).
| |
| | |
| (3) The second cleavage plane is transverse to the long axis of the embryo
| |
| and separates two large anterior cells from two small posterior ones (p. 7).
| |
| | |
| (4) The intersection of these two planes marks the vertical axis of the egg; one
| |
| end of this axis corresponds to the middle of the dorsal, the other to the middle of
| |
| the ventral face of the gastrula (p. 7).
| |
| | |
| (5) The third cleavage separates 4 larger dorsal cells from 4 smaller ventral ones (p. i); the latter are ectodermal, the former ''mixed."
| |
| | |
| (ti) At the fourth cleavage these 8 cells give rise to l(i; X ventral cells, all
| |
| ectodermal, and S dorsal cells, 6 of which are mixed, and 2, which are smaller than
| |
| any of the others and lie at the posterior pole, ectodermal (p. 8).
| |
| | |
| (7) By division these 1 li cells give rise to 32; l(i ventral cells, all ectodermal
| |
| and Id dorsal cells. 4 ectodermal derived from the 2 posterior ectoderm cells of the
| |
| previous stage. 6 ectodermal derived from the 6 mixed cells. 4 endodermal and 2
| |
| still mixed. With regard to the identification of the dorsal and ventral faces at
| |
| this stage they say: "'On bien les cellules ectodermique torment ensemble une
| |
| calotte appliquee par sa concavite" contra les globes endodermiques et mixtes (comme
| |
| dans fig. 10, c), ou bien e'est le contraire qui a lieu, les globes endodermiques et
| |
| mixtes s'etalent en surface de facon a constituer ensemble une calotte moulee sur
| |
| l'ectoderme (fig, '*. c)."
| |
| | |
| (8) At the next stage there are 44 cells; 32 ectodermal, easily recognized by
| |
| their transparency, and 12 other cells very much larger. The ectodermal cap is
| |
| notably extended and tends to envelope the endoderm.
| |
| | |
| From this stage onward there is no question as to the identification of the dorsal and ventral faces or the anterior or posterior ends. As will presently appear,
| |
| my work, like that of Chabry (1887), entirely confirms the orientation adopted by
| |
| Van Beneden and Julin, though I cannot agree with them as to the fate of certain
| |
| individual cells.
| |
| | |
| | |
| 2. Seeliger's System.
| |
| | |
| | |
| Seeliger's (1885) later work was much less detailed and satisfactory with
| |
| regard to the orientation of the early cleavage stages, as Castle has shown. His
| |
| principal conclusions as to orientation are :
| |
| | |
| (1) The first cleavage plane coincides with the median plane of the embryo,
| |
| hut neither anterior nor posterior, dorsal nor ventral can he recognized at this
| |
| stage (p. 48).
| |
| | |
| (2) The second cleavage divides the egg into two smaller anterior cells and
| |
| two larger posterior ones (p. 48).
| |
| | |
| (3) The third cleavage separates4 dorsal endodermal cells from 4 ventral ectodermal ones; the two posterior ventral cells are larger than any of the others.
| |
| Structurally all these cells are alike (p. 49).
| |
| | |
| (4) In the 16-cell stage the 8 dorsal endodermal cells are yellow and have
| |
| small nuclei ; the 8 ventral ectodermal ones are clear (p. 50).
| |
| | |
| In the identification of individual cells and their axial relations Seeliger was
| |
| much at fault. The small cells of the 4-cell and later stages are certainly not anterior in position but posterior, as has been shown by Van Beneden and Julin, Chahry. Samassa, and Castle; while the two larger cells of the 8-cell stage are not ventral but dorsal in position, not posterior hut anterior, as their relations to the two
| |
| small posterior cells show. Seeliger therefore mistook anterior for posterior, dorsal
| |
| tor ventral and consequently right for left; in short, he committed all the mistakes
| |
| possible in orientation.
| |
| | |
| | |
| 3. Samassd's System.
| |
| | |
| | |
| Ten years after the publication of Van Beneden and Julin' s work, Samassa
| |
| (1894) working on Ciona and Clavellina reached very different conclusions from
| |
| those set forth by the first named authors. With the first four conclusions of Van
| |
| Beneden and Julin mentioned above he agrees, save that in the unsegmented egg
| |
| he claims that only the median plane and the anterior and posterior, but not the
| |
| dorsal and ventral, poles can he recognized. With regard to the identification of
| |
| the dorsal and ventral sides he held that Van Beneden and Julin were completely
| |
| in error and that they had mistaken the dorsal for the ventral, the endodermal for
| |
| the ectodermal pole in all stages up to the 44-cell stage. As the most important
| |
| evidence of this false orientation Samassa cites Van Beneden and Julin' s figures '.' c
| |
| and 10 c, which represent optical sections in the sagittal plane of a 32-cell and a
| |
| 44-cell stage respectively. In the first of these the ectoderm cells are shown as
| |
| columnar, the endoderm cells as flattened; whereas in the second, figure 10c, the
| |
| ectoderm cells are flattened and the endoderm columnar. '"The figures of these
| |
| two authors." says Samassa, "are sufficient to show that figure 10c is properly and
| |
| figure c falsely oriented ; in both cases the cylindrical cells belong to the endoderm
| |
| and are dorsal in position." The words of Samassa directed against Van Beneden
| |
| and .Julin apply with equal or even greater force to himself : "Van Beneden and
| |
| Julin have not once sought," he says, "to bring forward one fact in support of this
| |
| remarkable transformation." With the exception of the worthless a priori argument that cells which have once been cylindrical must always remain so Samassa
| |
| has not produced a single argument or fact in favor of his contention.
| |
| | |
| | |
| 4. Castle's System.
| |
| | |
| | |
| In the same year Castle (1894), in a preliminary paper and again in his
| |
| final paper (1896) on the early embryology of Ciona intestinalis, reversed the orientation maintained by Van Beneden and Julin and held with Samassa that in all
| |
| stages preceding the 44-cell stage the Belgian investigators had mistaken dorsal
| |
| for ventral and vice versa. Furthermore, after having studied the formation of
| |
| the polar bodies, he was lead to the truly remarkable conclusion that these bodies
| |
| in ascidians are formed at the endodermal pole, whereas in all other animals, so far as known, they are formed at the ectodermal pole of the egg. His conclusions
| |
| were stated in the most positive manner and have been widely accepted, notwithstanding that such an orientation is absolutel} unique, and for this very reason should have been received with caution. Inasmuch as Castle's work is the most
| |
| thorough and extensive treatment of the early development of ascidians since the
| |
| appearance of Van Beneden and Julin's paper, and since his conclusions are diametrically opposed to my own. it seems desirable to give with some fulness his conclusions as to orientation as well as the evidences upon which these conclusions are
| |
| based. In speaking of Van Beneden anil Julin's work he says ( 1894, p. 200): . . .
| |
| 'It is my purpose to show that by yielding themselves to conjecture in so small a
| |
| matter as these three cell divisions, the eminent authors fell into an error which
| |
| invalidates the most important conclusions of their otherwise excellent work. For
| |
| in correlating the 44-cell stage with the 32-cell stage they have changed the orientation so that they have identified the dorsal side of one with the ventral side of
| |
| the other, the endodermal half of one with the ectodermal half of the other. Their
| |
| orientation of all the stages prior to the jj-ce/l stage is accordingly wrong. Their
| |
| terms ectodermal and endodermaL ventral and dorsal, as employed up to this stage,
| |
| must be interchanged." Again with regard to the point at which the polar bodies
| |
| form he says (1894, p. 211) : "1 have repeatedly seen the polar bodies and observed
| |
| continuously the cleavage stages following their formation. These observations lead
| |
| to the surprising but unavoidable conclusion that the point on the surface of the
| |
| egg at which the polar bodies form becomes later the center of the dorsal or endodermal half of the eps." Again in his later work (1896, p. 22b) he says with regard to this matter: . . '-The form changes accompanying maturation occur, in
| |
| Ciona at least, and presumably in ascidians in general, at the pole of the egg opposite to that at which they occur in Amphioxus, and, so far as known, in all other
| |
| animals producing eggs with polar differentiation; for the changes connected with maturation are uniformly reported to take place at the animal, i.e., at the more
| |
| richly protoplasmic pole, whereas in Ciona they take place at the vegetative pole.
| |
| . . . The statement made in the preceding paragraph presents a condition of affairs
| |
| so directly contrary to that found in other groups of animals, as well as to what has
| |
| been assumed by all previous writers to he the case in ascidians, that it requires the
| |
| presentation of unmistakable evidence in its support. Such evidence I have to offer,
| |
| both from the study of the living egg and from that of preparations."
| |
| | |
| | |
| | |
| | |
| | |
| Fig. VII. Four-cell stuni' of Ciona iniestinalis viewed from the animal pole ; the creuated line
| |
| represents the boundary between the protoplasm and yolk; the dotted line marks the anterior
| |
| limit of the crescent at the vegetal pole ; the four cells are approximately equal in size.
| |
| | |
| Fig. VIII. Four-cell stage of Cynthia partita seen from the animal pole ; the limits of protoplasm and crescent are represented as in the preceding figure; the two posterior cells are a little
| |
| smaller than the anterior ones.
| |
| | |
| | |
| | |
| What is this evidence? So far as it relates to the origin of the polar bodies at
| |
| the vegetal pole it is twofold; (a) the polar bodies are formed at the yolk-rich
| |
| pole, (b) this pole becomes the endodermal pole of the gastrula. As to the first of
| |
| these propositions I have already shown that the germinal vesicle fades and the first
| |
| maturation spindle appeal's at the protoplasmic pole (tigs. 77, 78, 172). Only later,
| |
| after the entrance of the spermatozoon, does the protoplasm now away from this pole.
| |
| leaving the maturation spindle closely surrounded by yolk; still later, during the
| |
| first cleavage, the protoplasm Hows back again to near the center of the egg and at
| |
| the (dose of this cleavage it moves still nearer to the pole at which the polar bodies
| |
| lie (figs. 100, 102. 10(1. 107. 178); thereafter this pole is always the more richly
| |
| protoplasmic. Therefore, except for a brief period after the fertilization and before
| |
| the first cleavage, when the protoplasm is temporarily withdrawn from the maturation pole through the influence of the spermatozoon, the maturation or animal pole
| |
| and the more richly protoplasmic pole are one and the same in ascidians as in other
| |
| animals.
| |
| | |
| As to the statement that the polar bodies are formed at a point which corresponds
| |
| to the center of the dorsal or endodermal pole of the gastrula it is evident that
| |
| unless the polar bodies have been actually followed through the development to a
| |
| stage when the ectodermal and endodermal poles are unmistakable, this statement
| |
| must rest upon indirect evidence furnished by a study of the cleavage stages. As
| |
| a matter of fact, Castle has not figured nor described the polar bodies in any egg
| |
| later than the 1G to 24-cell stage, whereas there is no trace of gastrulation in Ciona
| |
| before the 76-cell stage (fig. 200). Undoubtedly therefore Castle's evidence that the
| |
| polar bodies are formed at the endodermal pole must be indirect rather than direct,
| |
| and must he derived from the study and orientation of the cleavage stages. We
| |
| may therefore turn at once to the evidences which led him to reverse Van Beneden
| |
| and Julin's orientation of these stages. So far as 1 am able to discover there are, in
| |
| addition to several minor considerations which could at best be considered only as
| |
| confirmatory, two and only two general lines of evidence which he brings forward
| |
| in favor of his contention. They are the following:
| |
| | |
| (1) The hemisphere in which division is earliest as the egg passes from the L6cell stage to the 32-cell stage, and from the latter to the 46-cell stage becomes later
| |
| the ventral or ectodermal hemisphere of the embryo ( 1S94, p. 206 ; 1896, pp. 229 and
| |
| 235). The second paper refers to this proposition as having been demonstrated in
| |
| the first. What is this demonstration'.' So far as 1 can ascertain it consists merely
| |
| in the assumption that the cells, which in the lG-cell and 32-cell stages divide earlier than the others, must continue to divide more rapidly and thus give rise to the more
| |
| numerous ectoderm cells of the gastrular stage. So far from there being any demonstration of this proposition there is actually no evidence offered in support of it.
| |
| Furthermore, I can affirm from my own studies that it is not true. The cells which
| |
| lag behind in division up to the 64-cell stage, thereafter divide much more rapidly
| |
| than the others and give rise to the ectoderm of the gastrula [cf. figs. 130134 and
| |
| 196-204).
| |
| | |
| (2) Castle's second reason for rejecting the orientation of Van Beneden and Julin
| |
| is the same as Samassa's, viz., the peculiar shape of the cells at the two poles. In the
| |
| 32-cell stage and even earlier the cells at the maturation pole are long and columnar
| |
| while those at the opposite pole are thin and superficially lame. "They [the columnar cells] retain this columnar form up to and throughout gastrulation " (1896, p.
| |
| 237). They thus give rise directly to the columnar endoderm cells which are ultimately invaginated. On the other hand. Van Beneden and Julin maintained that the
| |
| flattened cells of the 32-cell stage became the columnar cells of the 1 44-cell stage and
| |
| that the columnar cells of the earlier stage became the flattened ones of the latter
| |
| stage. Castle says that their figures show at a glance the absurdity of such an
| |
| interpretation (1894, p. 208; 1896, p. 237). Since the whole orientation which he
| |
| adopts as opposed to that of Van Beneden and Julin rests upon the establishment
| |
| of this one point, it passes belief that hi', as well as Samassa, should not have taken
| |
| the most evident and direct step to prove it. Van Beneden and Julin figure optical
| |
| sections in the sagittal plane of an egg in the 32-cell stage showing the columnar
| |
| cells at the ventral pole, and of one in the 44-cell stage showing them at the dorsal
| |
| pole. Castle figures actual sections of a 32-cell stage and of a 76-cell stage, but
| |
| none between these two. A study of actual or of optical sections of eggs transitional between the 32-cell and the 76-cell stages would have shown conclusively
| |
| that the columnar cells of the former are graduall} transformed into the flattened
| |
| (ells of the hitter, and the flattened cells of the one into the columnar cells of the
| |
| other, and would thus have completely established Van Beneden and Julius
| |
| orientation. Such a series of optical sections of the Ciona egg, viewed from the left
| |
| side and also from the posterior pole, is shown in text figures IX to XVI, and the
| |
| various stages in this change of shape can there he clearly followed. A similar
| |
| series of actual sections of the egg of Cynthia is shown in text figures XVII to XXIV.
| |
| I do not find that this transformation is quite as rapid in Cynthia and Ciona as is
| |
| indicated by Van Beneden and Julin's figures 9 c and 10 c for Clavellina. At the
| |
| 44-cell stage the cells at both poles are columnar and of nearly equal height (text
| |
| figs. XIII, XIV), and not until the 64-cell or even the 76-cell stage is this transformation complete. It must not he supposed, however, that this change in shape
| |
| of the cells at the two poles is a continually progressive one. since all the cells hecome more superficial during division and more columnar during rest. Consequently
| |
| every cell changes shape more or less during each cycle of division; this is well
| |
| shown in figures XVIII and XX.
| |
| | |
| Other details which Castle regards as confirmatory of his view will lie taken up Inter, hut enough lias now been said, in my opinion, to show the untrust worthiness of his principal evidence against Van Beneden and Julin's system of orientation and in favor of his own.
| |
| | |
| | |
| | |
| 5. Evidences in favor of Van Beneden and Julias System.
| |
| While it is evident from these many and serious differences of opinion that it
| |
| is easy to make mistakes in the orientation of the ascidian egg, it is not true that
| |
| the egg is an unusually difficult one to orient. In fact there are few eggs, except
| |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| Figs. IX-X1I. Camera drawings of entire stained eggs of Ciona iiitesthuiUs viewed as transparent
| |
| objects. Figs. IX and XI are seen from the posterior pole; Figs. X and XII from the left side. Figs.
| |
| IX and X represent a 16-cell stage passing into a 24-cell stage ; Figs. XI and XII a 32 -cell passing
| |
| into a 44-cell stage. The head and tail of the arrow mark the position of the equator (third cleavage
| |
| plane) at the anterior and posterior poles. The cells of the crescent (mesoderm) are B 6 -3, B 6 -> and B 6 = ;
| |
| all the other cells of the lower hemisphere are yolk laden and the boundary between protoplasm and
| |
| yolk is indicated by a crenated line; the stippled areas adjoining the median plane in the cells Bs- 2 and
| |
| B 6 -3 represent caps of deeply staining protoplasm (clear in life). The segmentation cavity is shaded by
| |
| vertical lines and the cells bordering it are seen in median optical section ; the cells at the upper pole
| |
| are columnar, those at the lower pole flattened. The polar bodies, although shaded diagramatically, are
| |
| present exactly where they appear in the drawings.
| |
| | |
| those in which the cleavage is markedly unequal, in which this can be so easily
| |
| done. All the embryonic axes are (dearly distinguishable in the unsegmented egg,
| |
| and at every stage in development there are numerous landmarks by which the
| |
| different poles of the egg may be recognized. With the exception of Seeliger, all
| |
| students of the early development of ascidians have recognized that from the 16cell stage onward, the posterior pole is marked by two cells much smaller than
| |
| any others in the entire egg. The chief difficultv has been, as evidenced by the work of Seeliger, Samassa, and Castle, in distinguishing the dorsal and ventral faces
| |
| in the pregastrular stages. This is due to the fact that the cells at these two poles
| |
| are of somewhat similar shape, size and arrangement, as may he seen by referring
| |
| to figures 117. 118, 120, L24, 13(1 and 131 of this paper. However, the differences
| |
| | |
| between these poles are so marked that there never need he any confusion regarding them.
| |
| | |
| (1) The most striking difference between the two poles is found in the fact
| |
| that at all stages of the cleavage and gastrulation one pole is rich in protoplasm, the
| |
| other rich in yolk. This is particularly noticeable in Cynthia and Molgula, but is
| |
| also true of Ciona, though in this genus the differences between the two poles are not quite so marked as in the other genera named. This difference is so great that in
| |
| properly stained eggs one can always tell at a glance which is the yolk pole and
| |
| which the protoplasmic. In eggs stained in picro-haematoxylin the protoplasm
| |
| is red or light purple, the yolk yellow and the two poles are so unlike that there can be no excuse for mistaking them. In Cyntliia, indeed, these differences
| |
| can be easily recognized in the living egg, the yolk being slate-gray and the protoplasm colorless or yellow (figs. 28, 32, 37, 38). The yolk spherules are not scattered
| |
| through the cytoplasm, but the limits of the yolk and cytoplasm are sharp and distinct. In the cells at the yolk pole the cytoplasm is limited to a small area around
| |
| the nucleus; at the protoplasmic pole the cytoplasm occupies a large part of the
| |
| cell, the yolk being limited to the inner ends of the cells. This is seen especially well in actual sections taken in the vj:'s axis
| |
| XVII to XXIV, and it can there be seen that the cytoplasm is largely found in those
| |
| cells which lie on that side of the egg where the polar bodies are found, while the
| |
| cells at the opposite pole are almost entirely filled with yolk. These yolk-laden cells are ultimately tnvaginated and form endoderm (text figs. XXI to XXIV) and
| |
| arc therefore dorsal in position, while the protoplasmic cells at the opposite pole
| |
| form ectoderm and are ventral in position. The cells which form the posterior
| |
| boundary of the yolk-rich hemisphere contain the small spherules, already described,
| |
| which are characteristic of the yellow protoplasm of Cynthia. The distribution of
| |
| the yolk shows conclusively, therefore, that the cells of the animal or ventral hemisphere contain most of the clear protoplasm and give rise to the ectoderm, while
| |
| the cells of the vegetal or dorsal hemisphere contain most of the yolk and yellow
| |
| protoplasm and give rise to the endoderm and mesoderm.
| |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| Figs. XIII-XVI. Camera drawings of eutire eggs of Ciona intestinalu viewed as transparent objects ;
| |
| | |
| | |
| Figs. XIII and XV are seen from the posterior pole ; Figs. XIV and XVI from the left side. Figs. XIII
| |
| and XIV represent a 44-cell stage passing into a 62-cell stage; Figs. XV and XVI a 76-cell passing into a
| |
| 110-cell stage. The position of the equator, the boundary between protoplasm and yolk and the segmentation cavity are represented as in the preceding figures. In Fig. XIV the cells A7-4 and A? 8 are neural
| |
| plate cells, A?-3 and A" " chorda cells; in Fig. XVI the neural plate cells are A 8 ?, A 8 - 8 , A 8 ."5, A 8 -' 6 , while
| |
| the chorda cells are A 8 -s, A 8 - 6 , A 8 -'3, A 8 - 1 '!. The crescent cells (mesoderm) are B 6 -3, BM, B?-t, B?-3 (Figs.
| |
| XIII and XIVl and their dirivatives B7-5, &<>, B7 7, B?- 8 , B 8 -5, B 8 - 6 , BK B 8 - 8 (Fig. XV and XVI). The
| |
| other cells of the lower hemisphere are endodermal. In Figs. XIII and XIV the cells bordering the segmentation cavity (seen in optical section) are of about equal height at the two poles; in Figs. XV and
| |
| XVI the cells at the loVer pole are columnar, those at the upper pole flattened. The polar bodies are
| |
| present exactly where they appear in the drawings.
| |
| | |
| | |
| | |
| Figs. XVII-XX. Actual sections of eggs of Cynthia partita; Figs. XVII anil XIX in the median
| |
| plane, Figs. XVIII and XX in a transverse plane. Figs. XVII and XVIII represent a 20-24 cell stage;
| |
| Figs. XIX and XX a 32-44 cell stage. Unshaded portions of cells represent clear protoplasm ; closely
| |
| crowded spheres, the yolk ; minute spherules, the yellow protoplasm of the crescent. The clear protoplasm
| |
| is located chiefly in the cells of the animal half of the egg (ectoderm) ; it is also found in the crescent
| |
| cells (mesoderm) 1 15; =, B 6! , B 6 '3) and neural plate cells (A7-4, upper half of A 6 -=0 of the lower hemisphere.
| |
| The remaining cells of the lower hemisphere, (endoderm B5- 1 , B 6 - 1 , A 6,1 , A fc -3, and chorda, A<-3 and lower
| |
| half of A 6-2 ) are filled with yolk ; yolk is also found in the central ends of all the other cells. The yellow
| |
| protoplasm is limited to the crescent cells and to a single pair of cells of the upper hemisphere (bs-3). In
| |
| Fig. XVII the cells at the two poles are approximately equal in height ; in Fig. XVIII the cells at the
| |
| animal pole are flatter, probably owing to the fact that they are dividing ; in Figs. XIX and XX the cells
| |
| at the animal pole are columnar, those at the vegetal pole flattened. The polar bodies are actually present where they are represented.
| |
| | |
| | |
| | |
| such sections are shown in text figures
| |
| | |
| | |
| | |
| | |
| Figs. XX1-XXIV. Sections of eggs of Cynthia partita; Figs. XXI and XXIV in the median plane.
| |
| Fig. XXIII a little to one side of the median plane at the posterior end, Fig. XXII in a transverse plane.
| |
| Fig. XXI represents a 64-cell stage, Fig. XXII a 64-76 cell stage. Fig.XXIII a 76-110-cell stage, and Fig.
| |
| XXIV a 110-cell stage. The clear protoplasm, the yellow protoplasm and the yolk are represented as in
| |
| the preceding figures. The clear protoplasm is localized chiefly in the ectoderm and neural plate cells,
| |
| the yellow protoplasm in the crescent cells (mesoderm) and the yolk in the endoderm and chorda cells;
| |
| yolk is also present in the inner ends of the ectoderm and mesoderm cells. The polar hodies shown in
| |
| Fig. XXIII in dotted outline do not lie in the plaueof the section drawn, but in that of the next section of
| |
| ilie series. It is probable that the neural plate and chorda cells of this figure (k~-* and A>3) have already
| |
| divided in a transverse plane {v. fig. 131), and that these cells should therefore be labelled A 8 *? and \
| |
| a> in Fig. XXIV. In Fig. XXII the mesoderm cell B 6 - 2 (Fig. XVIII.i has divided into a mesenchyme
| |
| cell (B"-3) and a muscle cell IB'"*), the former containing little and the latter much of the yellow protoplasm.
| |
| | |
| (2) This orientation is further confirmed by a study of the yellow crescent of
| |
| the Cynthia egg and of the cells which develop from it. As has been shown, the
| |
| yelkvw protoplasm of this egg collects at the Lower (vegetal) pole and then moves up to a position just below the equator on the posterior side where it forms a yellow crescent. At the first cleavage this crescent is divided in the middle into right and
| |
| left halves: at the second cleavage it passes into the two posterior cells of the 4-cell
| |
| stage; at. the third cleavage it goes into the two posterior vegetal cells of the 8-cell
| |
| stage. In two subsequent divisions the yellow protoplasm is separated from the
| |
| yolk with which it is associated and thereafter forms a crescent of yellow cells which
| |
| surrounds the posterior side of the egg just below the equator (figs. 37, 39, 41, 42).
| |
| At all stages of development this crescent or. the cells which arise from it, lies in
| |
| the posterior half of the vegetal hemisphere, and the yellow cells are never separated from the mid-dorsal line by more than a single row of yolk cells (figs. 44-48).
| |
| On the other hand these yellow cells are separated from the mid-ventral line by an
| |
| ever increasing number of clear protoplasmic cells (figs. 43, 45, 122, 129, 137, et
| |
| seq.). The single row of yolk cells mentioned above as lying between the yellow
| |
| cells and the dorsal mid-line invaginates during: gastrulation and aives rise to the
| |
| ventral cord of endoderm in the tail of the larva, while the yellow cells, which are
| |
| also invaginated, trive rise to the mesoderm. A study of this yellow crescent and
| |
| of the cells which develop from it shows conclusively that it always lies on the posterior border of the yolk-rich or dorsal hemisphere, that at the 16-cell and 32-cell
| |
| stages, it is separated from cells which give rise to the ventral endoderm, and that
| |
| it is invaginated with the endoderm and forms the muscle cells and mesenchyme of
| |
| the tadpole.
| |
| | |
| (3) Wholly similar results as to the orientation of the egg and embryo follow
| |
| from a study of the lineage of all the other cells of the embryo. I believe that I
| |
| have seen every division of every cell up to the 21 8-cell stage, and in the critical
| |
| period between the 32-cell and 7(>-cell stages I have seen these divisions in hundreds
| |
| of cases. The evidence from this source as to the orientation cannot here be presented in detail but must be deferred to that portion of this paper which deals
| |
| particularly with the cell-lineage ; however, it can be said that in not a single instance have I found any evidence against the orientation according to Van Beneden
| |
| and Julin, while every observation which I have made on the cell-lineage speaks in
| |
| favor of that orientation.
| |
| | |
| (4) Finally a most direct and convincing evidence in favor of this system
| |
| of orientation is found in the position of the polar bodies throughout development. In preparations of the eggs of the three genera of ascidians which I have
| |
| studied, the polar bodies are easily distinguishable from the test cells by their
| |
| deeper stain ; in Ciona they are also larger than the test cells. In the last named
| |
| genus I have seen the polar bodies attached to the egg or imbedded in it at every
| |
| stage from the unsegmented egg to the gastrula (Plates XI and XII). In every
| |
| single instance they have been found at a point on the ectodermal (ventral) hemisphere which a study of the cell-lineage shows to correspond to the animal pole of
| |
| the unsegmented egg. I have not observed the polar bodies in every egg of Cynthia
| |
| which I have studied or drawn, possibly because they do not in this genus remain
| |
| attached to the egg so persistently as in Ciona. but wherever I have been able to identify them they have been found at the same polo of the egg as in Ciona (figs.
| |
| 87. 92, 96 3 102, 106, HIT. ins. 1 It). I ir,. I hi. L30, L33, L39, I 13). In stages later
| |
| | |
| than figures L39 and I !"> the protoplasm of the polar cells becomes vesicular and
| |
| stains so faintly that they can no longer be identified with certainty.
| |
| | |
| Castle maintained from indirect evidence, as 1 have already shown, that the
| |
| polar bodies of Ciona are formed at the middle of the endodermal or dorsal half of
| |
| the egg. I have never in a simile instance observed anything which might he mistaken for a polar body at this pole, whereas I have found the most positive and oft
| |
| repeated evidence that the polar bodies lie at the ectodermal or ventral pole from
| |
| the time of their formation to the gastrular stage. These ascidians therefore form
| |
| no exception to the genera] rule that the polar bodies are formed at the middle of
| |
| the ectodermal hemisphere of the egg.
| |
| | |
| It is not necessary in this place to point out in more detail than has been given
| |
| already the sources of error in the work of Seeliger and of Samassa, especially as their
| |
| work does not undertake to follow the exact cell-lineage of every cell up to the
| |
| gastrular stage or later. With Castle's work, however, the case is quite different,
| |
| for while the considerations already mentioned probably explain the sources of his
| |
| error of orientation, they do not explain the way in which he has incorporated this
| |
| error in the cell-lineage which he has followed to an advanced stage. In brief,
| |
| I find almost all of Castle's figures correctly drawn, and I can without difficulty correlate his drawings cell for cell with my own. The most important exceptions to this statement are found in his figures 53 and 54, hut even here the differences are not great. His gastrular stages are of course correctly oriented since the
| |
| dorsal and ventral faces of the embryo are unmistakably marked out as soon as the
| |
| invagination begins. All of his pregastrular stages, however, with the exception
| |
| of a 48-cell and a 64-cell stage, shown in his figures 57 to GO. are erroneously
| |
| oriented, dorsal being mistaken for ventral and ventral for dorsal. With the
| |
| lineage which he gives of every cell up to the 4G-cell stage (his fig. 56), I entirely
| |
| agree, but in passing to the 48-cell stage (his figs. 57 and 58) he inverts the egg
| |
| and shifts the equator one cell-row r nearer the vegetal pole than it should be,
| |
| consequently all of the lineage of the later stages is wrong. While therefore
| |
| the stages from 48 cells on are correctly orientated, the lineage of the individual
| |
| cells is incorrect; before the 48-cell stage the lineage is correct but the orientation
| |
| wrong. The evidence for this grows in part out of the general considerations
| |
| already mentioned, but it is also founded upon a detailed study of the cell-lineage, to which we now turn.
| |
| | |
| | |
| IV. CELL-LINEAGE.
| |
| | |
| | |
| A. Nomenclature. In order to facilitate reference to the work of others, it
| |
| is desirable that some good system of naming the individual cleavage cells be
| |
| adopted and thereafter adhered to even if it he not ideally perfect. The system
| |
| which has been employed with only slight modifications in all the recent cell-lineage work on annelids and mollusks is not well suited to the ascidian egg because in the latter there is no distinction of macromeres and micromeres, because there are
| |
| no "quartets" which arise from stem cells, because this system is not well adapted
| |
| to show the perfect bilateral symmetry of the egg and embryo, which is one of the
| |
| must characteristic features of ascidian development, and finally because of the great
| |
| number of figures which must be used as exponents in the representation of later
| |
| stages {eg. the letter designating each of the cleavage cells of the ninth generation,
| |
| figs. 140-143, wotdd need to be followed by no less than six exponents). Owing to
| |
| these reasons I early saw the difficulty of attempting to apply this system to the
| |
| cell-lineage of the ascidian egg. The most complete system of nomenclature which
| |
| has heretofore been used in the study of the cell-lineage of ascidians is that of
| |
| Castle, which is a modification of a system devised by Kofoid (1894). In this system, as is w^ell known, the four quadrants of the egg are designated by the letters
| |
| A, B, C, D ; after the third cleavage the cells nearer the vegetal pole are designated
| |
| by capitals, those near the animal pole by lower case letters. The first exponent following a letter indicates the generation to which a cell belongs, the second
| |
| exponent the position of the cell relative to the vegetal pole. With this system it
| |
| is always difficult to determine at a glance the lineage of any cell since "to ascertain the mother cell of any particular cell, its first exponent must be diminished by
| |
| one; and its second exponent, if an even number, must be divided by two, but if
| |
| an odd number it must be first increased by one and then divided by two. In
| |
| order to determine the daughter cell of a particular cell, simply reverse this process ;
| |
| that is, increase the first exponent by one and double the second exponent. To
| |
| determine the other daughter cell diminish this second exponent by one" (Castle.
| |
| L896, [>. 227). While it is thus difficult to determine at a glance the lineage
| |
| of any cell, the number of exponents required is relatively small, and this fact,
| |
| more than any other, has led me to adopt Castle's system, with the following modifications : The right and left halves of the embryo are designated by the same
| |
| letters, the names of cells on the right being underscored as compared with those
| |
| on the left. This method of designating the cells of the right and left sides is
| |
| essentially similar to that employed by Chabry (1887). In this way but two
| |
| letters are needed for the whole cell-lineage, one for the anterior and another for
| |
| the posterior quadrants. For these I desired to use the letters employed by
| |
| Van Beneden and Julin and by Chabry. viz. A and 1\ but owing to the difficulty of distinguishing between lower case and capital P. I finally chose the letter
| |
| B instead of P. The right anterior quadrant is A, the left A ; the right posterior quadrant is B, the left B. After the third cleavage all cells lying on the
| |
| polar body side of that cleavage plane are designated by lower case letters, while
| |
| those on the opposite side of that plane continue to be designated by capitals. This
| |
| modification not only emphasizes the bilateral character of the ascidian egg, but it
| |
| also simplifies the nomenclature. Furthermore, it facilitates reference to Castle's
| |
| work, for when in his orientation of the 48-eell stasre the egg is inverted as compared with earlier stages the right side is substituted for the left, and the letters A
| |
| and D, which in the earlier stages designate the actual left side, are used after the 48-cell stage to designate the actual right and vice versa. In my modification of his
| |
| system this substitution of one side for the other will be indicated only by the presence or absence of a line under the letter. In all stages later than the 48-cell stage
| |
| I continue to use lower case letters to designate cells of the animal or ectodermal
| |
| hemisphere, and capitals for those of the opposite hemisphere, whereas Castle
| |
| reverses this rule.
| |
| | |
| | |
| B. Cleavage of the Egg; First to Seventh Generation of Cells (1-G4 Cells).
| |
| | |
| | |
| Although the details of the early cleavages of the ascidian egg have been
| |
| treated at considerable length by previous writers on this subject, I have determined to present the subject here, cleavage by cleavage, both because my results
| |
| differ in many respects from the conclusions heretofore reached and because I wish
| |
| to call attention to certain features of these cleavages which have not as yet been'
| |
| noticed. I shall shorten the account wherever possible by references to previous
| |
| work.
| |
| | |
| As is well known, ascidian eggs develop with great rapidity; there are certainly few other eggs which develop so rapidly. Both Ciona and Cynthia reach
| |
| the fully formed tadpole stage in about twelve hours, while Molgula reaches this
| |
| stage in not more than eight hours after fertilization. Certainlv there are few things
| |
| more wonderful than the origin of a complex animal. of a chordate , from an egg
| |
| in the short space of from eight to twelve hours ! The portions of this brief period
| |
| devoted to the different stages of development are interesting and suggestive. In
| |
| Cynthia about 40 minutes elapse between the fertilization and the appearance of
| |
| the first cleavage , furrow ; about 140 minutes between the first cleavage and the
| |
| beginning of gastrulation ; and about 140 minutes between this stage and the young
| |
| tadpole stage shown in figure 103. The different generations of cleavage cells are
| |
| separated from one another by intervals about as follows : 1
| |
| | |
| Fertilization to first cleavage ( 1-2 Cells).
| |
| | |
| First to second cleavage
| |
| | |
| Second to third cleavage
| |
| | |
| Third to fourth cleavage
| |
| | |
| Fourth to fifth cleavage
| |
| | |
| Fifth to sixth cleavage
| |
| | |
| Sixth to seventh cleavage
| |
| | |
| Seventh to eighth cleavage (112-218
| |
| | |
| Eighth cleavage to young tadpole stage (fig. 163),
| |
| | |
| 1. First Cleavage; 1-2 cells. (Figs. 18-27, 96-100, 177. 178. 181.)
| |
| | |
| After the meeting of the germ nuclei, which occurs about midway between the
| |
| | |
| center of the ci'i and the posterior pole (figs. 91, 92, 93), the cleavage spindle
| |
| | |
| develops rapidly and moves inward from the posterior pole until it lies almost
| |
| | |
| 1 These figures are based upon the study of eggs laid and fertilized at 5 p.m. and put up at intervals thereafter "until 11 p.m. In each of these hits eggs were found in several different stages and the
| |
| results can be aecejitr.l u- only approximately correct.
| |
| | |
| | |
| | |
| ( 2-4 "
| |
| | |
| | |
| >.
| |
| | |
| | |
| I 4-8
| |
| | |
| | |
| ),
| |
| | |
| | |
| | 8-16 "
| |
| | |
| | |
| ),
| |
| | |
| | |
| t 16-32 "
| |
| | |
| | |
| ),
| |
| | |
| | |
| i .",2-64 "
| |
| | |
| | |
| ),
| |
| | |
| | |
| ( 64-112
| |
| | |
| | |
| )
| |
| | |
| | |
| (112-218
| |
| | |
| | |
| )
| |
| | |
| | |
| | |
| 40
| |
| | |
| | |
| min
| |
| | |
| | |
| 30
| |
| | |
| | |
| a
| |
| | |
| | |
| 30
| |
| | |
| | |
| u
| |
| | |
| | |
| 20
| |
| | |
| | |
| n
| |
| | |
| | |
| 20
| |
| | |
| | |
| a
| |
| | |
| | |
| 20
| |
| | |
| | |
| a
| |
| | |
| | |
| 20
| |
| | |
| | |
| u
| |
| | |
| | |
| 20
| |
| | |
| | |
| a
| |
| | |
| | |
| 2
| |
| | |
| | |
| hrs.
| |
| | |
| | |
| exactly in the center of the egg (fig. 96), always heing oriented so that it lies at
| |
| right angles to the plane of the first cleavage and with its equator in that plane.
| |
| At the same time the clear protoplasm and a small portion of the yellow protoplasm
| |
| move inward from the posterior pole toward the center of the egg (figs. 92, 96).
| |
| . The larger part of the yellow protoplasm remains at the surface in the form of a
| |
| crescent, but the clear protoplasm is entirely withdrawn from the surface except for
| |
| a narrow zone which lies just above (ventral to) the crescent on the posterior side
| |
| (figs. 13-18, 96). During the formation of the first cleavage furrow, even this
| |
| narrow zone of clear protoplasm is withdrawn from the surface to the center of the
| |
| egg, so that the yolk now covers the entire surface of the egg except for the area
| |
| of the crescent (tigs. 100-102 and 178-179). This condition is just the reverse of
| |
| that which prevailed at the beginning of development, when the yolk was central
| |
| in position and the protoplasm peripheral (figs. 76-79).
| |
| | |
| The centrosomes and asters are larger and more easily studied in Ciona than in
| |
| Cynthia. Proceeding from the periphery to the center, the following parts of the aster
| |
| may be recognized (figs. 177, 179) : (1) The deeply staining, peripheral layer of the
| |
| aster, (2) the clear inner layer of the aster traversed by radiating fibres, (3) a granular
| |
| central body upon which the astral fibres end. The latter is the centrosome, and
| |
| is plainly composed of two parts, (a) an outer granular zone and (b) a central clear
| |
| area from which the netrum arises. In Cynthia the outer and inner layers of the
| |
| aster are not distinguishable and the centrosome itself is not so large as in Ciona ;
| |
| the latter is, however, composed of the same parts, viz., a peripheral granular zone
| |
| and a central clear area which gives rise to the netrum (figs. 98, 99). In these
| |
| ascidians, just as in the gasteropods which I have studied (Conklin, 1902), the
| |
| centrosome undergoes a decided growth and metamorphosis during the cycle of
| |
| division ; in the early stages of the cycle it is a small, deeply staining body, in the
| |
| Later stages it becomes much larger and differentiates into the outer granular zone
| |
| and the central clear area (cf. figs. !)7, 98).
| |
| | |
| In Ciona both the first and second cleavage spindles are remarkable in that at
| |
| all stages of the division the nuclear part of the spindle can be clearly distinguished
| |
| from the polar or astral part (figs. 177, 17'.l). The portion of the spindle derived
| |
| from the linin of the germ nuclei is short, deeply staining and barrel-shaped, and in
| |
| all respects resembles a maturation spindle (compare figs. 177 and 179 with figs. 67.
| |
| 70, 71). Even in the possession of a few peripheral fibres which radiate from the
| |
| slightly rounded ends of the spindle toward the equator, this spindle resembles
| |
| those of the maturation divisions. These peripheral fibres are not in line with the
| |
| astral radiations, and hence are all the more striking. The astral rays which run
| |
| from the centrosomes to the ends of this nuclear spindle are small and faintly staining as contrasted with the heavy, deeply staining fibres of the nuclear spindle. Nowhere else, so far as I am aware, is this double character of the mitotic figure so
| |
| clearly shown as in these cleavages of Ciona. This is due to the small size of the
| |
| nuclear spindle and to the large size of the astral systems, so that the ends of the
| |
| nuclear spindle are not easily confused with the astral rays, and also to the great difference in the staining reactions of the two. Except for the presence of these astral systems, this cleavage spindle is almost exactly like a maturation spindle.
| |
| | |
| In Cynthia the distinction between these two parts of the cleavage spindle is
| |
| not so clear because here the nuclear portion is much Longer and reaches nearly to
| |
| the centrosomes (fig. 97); but even here the nuclear part can be distinguished from
| |
| the astral by the stronger character of its fibres. In this genus also the astral systems are not so Large as in Ciona and the individual fibres are stronger, so that the
| |
| contrast between the nuclear and the astral portions of the spindle are but faintly
| |
| marked.
| |
| | |
| These cleavage spindles of Ciona, like the maturation spindles already described,
| |
| are of especial interest for the study of the mechanics of mitosis. 1 have not attempted
| |
| to make a detailed study of this subject, but it is quite evident that the separating
| |
| chromosomes move only as far as the ends of the nuclear spindle fibres (fig. 99).
| |
| They are never drawn up into contact witli the centrosomes, but remain at the
| |
| border of the aster, where they are transformed into chromosomal vesicles. In the
| |
| maturation divisions there are no centrosomes or asters at all to complicate the
| |
| problem, and liere also the chromosomes move only to the ends of the nuclear spindle fibres. These fibres elongate somewhat in the later stages of mitosis (figs. 70,
| |
| 71, 72), thus separating more widely the daughter chromosomes.
| |
| | |
| The fact that in the maturation divisions the chromosomes separate without
| |
| the aid of centrosomes or asters may be taken as evidence that in the similar spindles of the first and second cleavages the centrosomes and asters, although present,
| |
| take no part in this work. Anything which will explain the movements of the
| |
| chromosomes in such spindles as those shown in figures 69 to 72 will also explain
| |
| their movements in such cleavage spindles as those shown in figures 177 and 179.
| |
| In the maturation mitoses there are neither centrosomes nor asters, and yet the
| |
| separation of the chromosomes occurs in the usual manner. The spindle fibres
| |
| apparently serve only as a guide for this movement, and must be considered the
| |
| result rather than the cause of stresses in the cell substance. This is shown by
| |
| the fact that when they first appear these spindle fibres are not parallel but run
| |
| in all directions (figs. 02, G4). Later, under the influence of stresses in the cytoplasm, they become parallel. Under these circumstances there is no reason to
| |
| believe that the movement of the chromosomes is caused by other factors than
| |
| those which bring about movements in the cell body.
| |
| | |
| Tile constriction of the cell body first occurs, as indicated by figure 20. at the
| |
| posterior pole. This is probably due, as Castle says, to the fact that at this stage
| |
| tliis pole is the more richly protoplasmic one. Very soon the constriction extends
| |
| all the way around the egg and, as the mitotic spindle lies in the middle, it follows
| |
| that the constriction must be about as deep on one side as on another (fig. 99).
| |
| This constriction divides the ooplasm with exact ecpiality, not only quantitatively
| |
| but qualitatively also. During and immediately following this division the yellowcrescent undergoes some very remarkable transformations. These changes are
| |
| shown in figures 21 to 26, which represent consecutive stages of the same egg drawn in intervals of about two or three minutes. The crescent is first constricted in the
| |
| middle (figs. 20 and 21) ; then the gray yolk penetrates into the lower part of each
| |
| half of the crescent and approaches near to the surface, being covered only by a
| |
| thin layer of the yellow protoplasm (fig. 22). Above and below this intrusion of
| |
| yolk the crescent remains deep yellow in color ; in the region of the intrusion the
| |
| color is gray with a superficial covering of yellow pigment granules. The lower
| |
| (dorsal) portion of the crescent thus cut off from the remainder is small as compared
| |
| with the upper part, and is median in position. Not more than two minutes afterward this lower part unites with the upper along the median line (fig. 23), thus
| |
| forming a deep yellow semicircle in each blastomere. The intrusion of yolk may
| |
| still be seen entering this semicircle through its open half, which is dorsal and lateral in position. Then these two semicircles come into contact with each other
| |
| along the first cleavage plane, the free ends enlarge into rounded knobs, and the
| |
| intrusion of yolk is less large (fig. 24). Finally, the intrusion becomes still smaller,
| |
| the open ends of each semicircle join, and the crescent is reestablished (figs. 25 and
| |
| 26). Observation of the living egg during this period of division gives the impression of remarkable cytokenetic activity in all the ooplasm ; not only does the crescent take pai't in this activity but the yolk and the clear protoplasm undergo
| |
| marked movements, in the course of which the clear protoplasm is divided into two
| |
| areas which are entirely separated from each other by a partition of yolk (figs. 2527). I have been unable to analyze all of these movements; one thing, however,
| |
| seems very probable, viz., that they are in the main of a vortical nature and that
| |
| they are comparable with the movements in the constriction of the cell body which
| |
| I have observed in gastropods (Conklin. 1902). I have not thoroughly studied
| |
| these movements by the aid of serial sections ; since they take place with such
| |
| rapidity, this would be possible only by sectioning and studying a very large number
| |
| of eggs during the period of the first cleavage. In figure 99, which is an equatorial
| |
| section of an egg of the stage shown in figure 22 or 23, the substance of the crescent
| |
| (Cr) can be recognized on the posterior side of the egg; it does not. however, show
| |
| any of the thickenings or thinnings indicated in the surface views mentioned.
| |
| Beneath the superficial layer of yellow protoplasm in this figure is an area of clear
| |
| alveolar protoplasm, while still deeper is the radiating protoplasm which constitutes
| |
| the astral systems.
| |
| | |
| In the telophase of the first cleavage the centrosomes, daughter nuclei, and the
| |
| surrounding areas of clear protoplasm rotate toward the animal pole (fig. 100) in a
| |
| manner similar to that which occurs in the blastomeres of gasteropods (Conklin, 1902).
| |
| Through the agency of these telokinetic movements of the first cleavage the nuclei,
| |
| centrosomes and clear protoplasm are carried above the equator of the egg toward
| |
| the animal pole. The pole at which the polar bodies lie thus becomes more richly
| |
| protoplasmic than the opposite pole and ever after continues to be so (fig. 102.^7 seq.).
| |
| Castle has observed this telokinetic movement in Ciona, and describes it in the following words (1896, p. 233): "The first cleavage spindle arises, as has been stated, not
| |
| far from the center of the egg. As its first cleavage is nearing completion, however, the attraction spheres and nuclei begin to move toward the dorsal surface of the
| |
| egg, me ay fro?) i its more richly protoplasmic animal pole, from which the plane of
| |
| separation cuts it more rapidly." I find, however, that in Ciona, as in Cynthia,
| |
| this telokinetic movement of the nuclei and spheres is not away from the more
| |
| richly protoplasmic pole, unless the substance of the crescent he considered as constituting the protoplasmic pole, hut that the clear protoplasm also moves with the
| |
| nuclei and spheres toward tin' animal (ventral) pole (fins. ITS. 182). Sobotta (1807)
| |
| has also observed a similar bending of the spindle axis and movement of the
| |
| daughter nuclei and centrosomes in the anaphase or telophase of the first cleavage
| |
| of the egg of Amphioxus. He describes it as an attempt on the part of the centrosomes and nuclei to regain the center of the blastomeres ; but it is probable that this
| |
| is only another case of the telokinetic rotation of the cell contents with the consequent establishment of a new cell axis [cf. Conklin, 1902).
| |
| | |
| 2. Second Cleavage; 2-4 cells. (Figs. 28-30, 101-105. 179, 182, 183).
| |
| | |
| During the anaphase of the first cleavage each centrosome becomes elongated
| |
| at right angles to the spindle axis and to the chief axis of the egg and gives rise to a
| |
| minute centrosomal spindle or netrum at each pole of the mitotic figure (figs. 98, 99).
| |
| These netra elongate in the antero-posterior axis until the daughter centrosomes
| |
| come to lie at the anterior and posterior poles of the nucleus of each blastomere ; the
| |
| nuclear membrane is then dissolved and the second cleavage spindles are formed (figs.
| |
| 101, 102, 177, 179). These spindles, like those of the first cleavage, are composed
| |
| of a nuclear and an astral portion, the two being distinguishable with especial ease
| |
| in Ciona (fig. 179). The spindles lie in an area of clear protoplasm above the equator of the egg, and in Cynthia are slightly eccentric toward the posterior pole (fig. 102).
| |
| The areas of clear protoplasm become elongated in the antero-posterior axis (figs. 28,
| |
| 101, 102, 179) ; they are surrounded on all sides by yolk, which forms a peripheral
| |
| laver over the whole surface of the egg, except in the region of the crescent. The
| |
| substance of the crescent is directly continuous with the clear protoplasm in the
| |
| region of the posterior pole of the second cleavage spindle (figs. 102, 17'.)). Although
| |
| the yolk surrounds the areas of clear protoplasm it is not uniformly thick on all
| |
| sides; on the upper or ventral side of these areas the layer of yolk is very thin, on
| |
| the lower or dorsal side and anterior to the crescent it is especially thick. This distribution of the yolk can be seen not only in sections, such as figures 102 and 17!), but
| |
| also in entire preparation and in living eggs. In the latter the light gray of the
| |
| upper hemisphere (figs. 28, 29 and 30), as contrasted with the dark gray of the lower
| |
| hemisphere, indicates that the layer of yolk which surrounds the areas of clear protoplasm is thin over the upper hemisphere and thick over the lower. The area at
| |
| the vegetal pole, where the layer of yolk is thickest, gives rise to the endoderm cells
| |
| which are always yolk laden : the upper hemisphere, where the yolk layer is thin.
| |
| gives rise to the ectoderm cells which contain a relatively small amount of yolk ;
| |
| the substance of the crescent gives rise to the muscle and mesenchyme cells, and in
| |
| Cynthia is always characterized by the presence of the yellow spherules. In later stages the yolk ceases to be peripheral in position and comes to lie in the central
| |
| portions of the cleavage cells (text figs. XVII XXIV) ; this change in position is
| |
| brought about by the flowing of the peripheral layer of yolk inward along all the
| |
| developing cleavage furrows (figs. 104-107, et seq.) until finally the yolk comes to
| |
| occupy a central position in all the blastomeres, while the clear protoplasm is
| |
| brought once more to the surface. These cytokinetic movements which accompany
| |
| cell division do not change the relative distribution of yolk and cytoplasm in the
| |
| different hemispheres and quadrants of the egg, but only its location in the individual cleavage cells.
| |
| | |
| In the second cleavage the constriction of the cell begins at the periphery or
| |
| free surface and proceeds inward through the cell body (figs. 104, 105). The
| |
| peripheral layer of yolk is thus carried inward along the cleavage furrow, as has
| |
| been said, and the middle of each spindle is bent in toward the center of the egg
| |
| (fig. 105). At the same time the yolk and yellow protoplasm continue to be carried in along the first cleavage furrow. The inflow of yolk along a developing
| |
| cleavage furrow leaves a protoplasmic connection between the two daughter cells,
| |
| into which the yolk does not penetrate for a considerable time ; this protoplasmic
| |
| connection is frequently of service in determining the lineage of cells since it always
| |
| connects daughter cells (figs. 104-107). Finally the inflow of yolk completely cuts
| |
| oft' this connection.
| |
| | |
| The four cells which are formed by the second cleavage are all approximately
| |
| of the same size in Ciona ; in Cynthia the two anterior cells are slightly larger than
| |
| the posterior ones, just as Van Beneden and Julin found to be the case in Clavellina (text figs. VII, VIII). But though the daughter cells are of nearly the same
| |
| size they are of very different quality. The posterior cells contain about the same
| |
| quantity of clear protoplasm as the anterior ones, but they contain little yolk and
| |
| practically all of the yellow crescent substance; the anterior cells on the other hand
| |
| contain a great deal of yolk, but practically none of the crescent substance. The
| |
| second cleavage is therefore differential in a very marked degree (r/". figs. 29 and 30).
| |
| | |
| | |
| 3. Third Cleavage; 4-8 cells. (Figs. 31-35, 106-100, 184).
| |
| | |
| In the anaphase of the second cleavage the centrosomes elongate in the vertical axis and the daughter centrosomes, moving to the upper and lower poles of the
| |
| nuclei, form the centrosomes of the third cleavage spindles. In an abnormal egg
| |
| shown in figure 103 this division of the centrosomes occurs in one of the blastomeres
| |
| in the prophase of the second cleavage and not in a vertical but in a horizontal direction. The position of these third cleavage spindles is peculiar and of great prospective
| |
| significance. They are slightly eccentric toward the animal pole, and accordingly
| |
| the four cells which are cut oft* at this pole are smaller than those at the vegetal
| |
| pole. When the egg is viewed from either the right or the left side the spindles in
| |
| the anterior and posterior quadrants seem to be parallel and are both slanted forward at the upper pole ; accordingly the four upper cells, when formed, lie slightly
| |
| anterior to the four lower ones (figs. 108, 184). When the egg is viewed from the anterior pole it is seen that the spindles in the anterior quadrants are not parallel.
| |
| but that they converge toward the animal pole. The reverse is the case if the egg is
| |
| viewed from the posterior pole, i.e., the spindles in the posterior quadrants diverge
| |
| toward the animal pole. Thus it comes about that the nuclei in the anterior-vegetal cells (A 41 ) are relatively far apart, those in the anterior-animal cells (a 4 ' 2 ) (dose
| |
| together (figs. 106 and 109); whereas the reverse is the case in the posterior cells,
| |
| i.e., the nuclei in the posterior-vegetal cells (B 41 ) are near together, those in the
| |
| posterior-animal cells (b 4-8 ) far apart (tig. 107). *
| |
| | |
| Every one of these matters is of prospective significance in the further development of the embryo; associated with the forward slant of the spindles toward
| |
| the animal pole is the tact that the cells of the animal hemisphere overhang those
| |
| of the vegetal hemisphere at the anterior pole; whereas the posterior cells of the
| |
| vegetal hemisphere are not completely covered by those of the animal hemisphere when the egg is viewed exactly from the animal pole (figs. 110. 112, 116).
| |
| Associated with the convergence of the spindles in the anterior quadrants toward
| |
| the animal pole and the convergence of the spindles of the posterior quadrants
| |
| toward the vegetal pole is the fact that in later stages the anterior half of the
| |
| vegetal hemisphere is broad from side to side, its posterior half narrow, while the
| |
| anterior half of the animal hemisphere is narrow from left to right, its posterior
| |
| half broad (figs. 109-118, et seq.). While the position of these spindles is therefore
| |
| indicative of important prospective characteristics of the embryo, it must not be
| |
| regarded as the initial cause of these characteristics. Indications of these features
| |
| may be seen in the distribution of the yolk and protoplasm at the four-cell stage,
| |
| and there can be no doubt that the position of the spindles is itself the result of
| |
| cytoplasmic localization.
| |
| | |
| One of the features of this stage to which Castle calls particular attention is the
| |
| presence of a " cross-furrow " on the right and left sides between the anterior dorsal
| |
| and the posterior ventral cells (A 4 - 1 and b 4 '", figs. SI, 32, 108, 184). I find, as did
| |
| Castle and Chabry, that this cross-furrow is constant in position and that it marks a
| |
| downward bend in the equator, which may be observed as late as the gastrula stage ;
| |
| in the region of this downward bend the ectoderm cells grow down over the cells of
| |
| the vegetal hemisphere in advance of the neighboring ectoderm cells (figs. 116-119,
| |
| 123-126, 128, 130, 134, et seq.). I observed the process of formation of this crossfurrow r in the living egg, and have represented this in figure 31. When the third
| |
| cleavage furrows first appear, they are all in nearly the same plane, the furrows
| |
| between the daughter cells of the posterior quadrants being nearly perjiendicular to
| |
| the egg axis, as indicated by the faint line between the cells B 41 and b 4 8 of figure 31,
| |
| which line represents the position of the furrow between those cells when it first
| |
| appears. A minute or two afterward this furrow is tilted downward at its anterior
| |
| end and upward at its posterior, as indicated by the heavy line between those two
| |
| cells ; in this way the cross-furrow arises on the right and left sides of the egg
| |
| hetween the anterior dorsal and the posterior ventral cells.
| |
| | |
| | |
| 1 Figs. 106 and 107 represent two sections of one and the same egg, in the 8-cell stage, the former through the nuclei of the anterior cells, the latter through the nuclei of the posterior ones.
| |
| | |
| | |
| During telokinesis the movements in the cell body are similar to those which
| |
| occur at the close of the second cleavage, i. e.. the middle of the spindle is carried
| |
| in toward the centre of the egg while the poles of the spindle move outward toward
| |
| the surface (figs. 106, 107). By this movement the spindle axis is much bent on
| |
| itself. I have not observed in these eggs any tendency for the sphere substance at
| |
| the poles of the spindles to be carried as near as possible to the animal pole, a
| |
| thing which is very apparent in gasteropod eggs.
| |
| | |
| I have already called attention to the fact that the four cells at the animal pole
| |
| are smaller than those at the vegetal pole; this disparity is most marked between
| |
| the upper and lower cells of the anterior quadrants (figs. 106, 107). The anterior
| |
| dorsal cells (A 41 ) are the largest in the egg at the eight-cell stage, the anterior
| |
| ventral cells (a 42 ) the smallest. The posterior dorsal cells (B 41 ) are but little, if any,
| |
| larger than the posterior ventral ones (h 42 ), and both are intermediate in size
| |
| between the upper and lower anterior cells.
| |
| | |
| The different cell substances are distributed to the eight cells as follows : The
| |
| clear protoplasm is found in all the cells, but is most abundant in the four ventral
| |
| cells and least abundant in the two posterior dorsal cells (B 41 ); yolk is found in
| |
| all of the cells, but is most abundant in the two anterior dorsal cells (A 41 ) and least
| |
| abundant in the four ventral cells ; the yellow protoplasm or crescent substance is
| |
| confined almost entirely to the two posterior dorsal cells (B 41 ), but a very small
| |
| amount of it is found around the nuclei of all the cells (figs. 106, 107, Cr. s.). My
| |
| attention was first drawn to the yellow protoplasm around the nuclei in my study
| |
| of the living eggs of Cynthia (figs. 32-40, et seq.)\ since then I have found, in
| |
| preserved material, a few of the spherules of this yellow protoplasm around the first
| |
| cleavage spindle and around the resting nuclei of the 2-cell, 4-cell and 8-cell stages
| |
| (figs. 96, 106, 107). In later stages of development it is found around the nuclei
| |
| of a few of the ectoderm cells even as late as the young tadpole stage (plates IV
| |
| and V). In spite of this perinuclear distribution of some of this crescent substance, it
| |
| is largely limited to the two posterior dorsal cells (B 41 ) of the 8-cell stage, where it
| |
| constitutes more than half of all the substance of those cells (figs. 31-35, 106, 107).
| |
| | |
| Van Beneden and Julin lirst observed that the four ventral cells of the
| |
| 8-cell stage are smaller than the lour dorsal ones; Seeliger, Samassa, and Castle
| |
| observed this same fact, though they incorrectly called these smaller cells dorsal in
| |
| position. Castle is quite right when he says (1896, p. 228) the " four cells which Lie
| |
| nearest the polar globules are smaller than those more remote," but I cannot understand how it was possible for him to reach the conclusion that- the smaller cells
| |
| ' are more abundantly supplied with yolk," while the larger cells are richer in protoplasm (pp. 234 and 235). According to my observations this is not true of Cz'ona,
| |
| Cynthia or Molgnla.
| |
| | |
| During the 8-cell stage two little bosses or caps of clear but deeply staining
| |
| protoplasm which will give rise at the 64-cell stage to the small posterior mesenchyme cells (B 7,; fig. 130 et seq.) are visible on the posterior surface of the cells B 41
| |
| and B 4 ' (fig. 184). These little caps lie in contact with each other on each side of the mid-line and right at the middle of the crescent, of which they form an
| |
| extremely small part. They arc formed by the aggregation at this point of a little
| |
| clear protoplasm which first appeared at the time of fertilization as a clear area
| |
| around the spermatozoon, and which afterwards lies at the middle of the crescent
| |
| (figs. 173, 17->. 17li). In Cynthia this area of clear protoplasm does not usually
| |
| take the form of the deeply staining hosses or caps before the 1 (i-cell stage (figs.
| |
| 113. 110), though these may sometimes appear, as they do in Ciona, at the 8-cell
| |
| stage. Although they arise from the surface of the crescent they contain no yellow
| |
| pigment, and in the living egg this small spot of protoplasm ami the cells to which
| |
| it gives rise are almost perfectly transparent and are therefore difficult to see. In
| |
| stained preparations they always stain deeply and thus form an excellent landmark (figs. 116-120, et seq).
| |
| | |
| Chabry and Castle have called particular attention to these prominences of
| |
| clear protoplasm which are found at the posterior pole of the egg, and Castle traces
| |
| them hack to the 2-cell stage, and ives good reason for believing that this area
| |
| of clear protoplasm marks the point of entrance of the spermatozoon, and was
| |
| caused by it. I entirely agree with Castle that this aggregation of clear protoplasm
| |
| is caused by the entering spermatozoon, since I have seen it surrounding the spermatozoon immediately after its entrance (fig. 173); but it can scarcely be said to
| |
| mark the point of entrance, as it does not remain stationary hut moves with the
| |
| crescent from a point near the vegetal pole to one near the equator on the posterior
| |
| side of the erg. So far as I am able to determine from a study of Castle's figures and description, the area of finely granular protoplasm, which he represents in his
| |
| figures 17, 45, 40 and 47, is the middle portion of the crescent. The large area of
| |
| clear protoplasm represented in each of these figures and marked x gives rise to
| |
| the middle cells of the crescent (C 5 - 2 , D 52 of his fig. 49), therefore the small posterior mesenchyme cells, C" r ' and D 75 of later stages, can represent but a small part
| |
| of the area marked x in the earlier stages. The earliest stage in which Castle
| |
| represents the substance of these future mesenchyme cells is at the posterior pole
| |
| of the cells C 63 and D 63 in his figure 51. 1 conclude, therefore, that he observed
| |
| the middle portion of the crescent (x of his figures) in the earliest stages of the
| |
| development, but that be did not recognize the substance of the small mesenchyme
| |
| cells as distinguished from the substance x (crescent substance) before the 24-cell
| |
| stage (his fig. 51).
| |
| | |
| All students of ascidian embryology agree that the first plane of cleavage is
| |
| median in position, the second transverse, and the third horizontal or coronal, but
| |
| beyond this there are few agreements among them, as has been pointed out. In
| |
| the matter of the relations of these cleavage planes to the germ layers there are as
| |
| many opinions as there are concerning the orientation of the v^<j:. Van Beneden
| |
| and Julin (1884) maintained that the four ventral cells of the 8-cell stage are
| |
| purely ectodermal, but that the four dorsal cells are still mixed." each of them
| |
| containing ectoderm and endoderm, while not until the 44-cell stage is the separation of ectoderm and endoderm in these dorsal cells completed. Seeliger ( 1 885) held that the third cleavage plane separates the ectoderm from the endoderm, the four
| |
| ventral cells being ectodermal, the four dorsal endodermal. Davidoff (1891 ) found
| |
| that in Distaplia the tour ventral cells are ectodermal, the four dorsal endodermal.
| |
| and a similar view is maintained by Samassa (1S'.>4). Castle (189G), on the other
| |
| hand, maintains that both the ventral and the dorsal cells of the 8-cell stage are
| |
| mixed, the ventral cells containing ectoderm and mesoderm, and the dorsal cells
| |
| endoderm and mesoderm, and not until the 48-eell stage are the substances of these
| |
| layers finally separated.
| |
| | |
| My work, like that of Castle, places hut little weight upon the idea of germ
| |
| layers, since it undertakes to trace specific organs to certain cleavage cells, and
| |
| even to certain regions of the unsegmented egg. Emphasis is therefore placed upon
| |
| organs and upon organ-forming cells and substances rather than upon the more
| |
| indefinite germ layers. However, I find that the tour ventral cells of the 8-cell
| |
| stage are purely ectodermal, while the four dorsal cells are endodermal and mesodermal, save for the fact that four neural plate cells (A 74 , A 78 , figs. 120, 121, 123)
| |
| will arise from the anterior portion of the dorsal hemisphere at the 44-cell stage.
| |
| The mesoderm and endoderm are first completely separated at the 22-cell stage
| |
| (figs. 117, 118). I find that only four ectodermal (neural plate) cells come from the
| |
| dorsal hemisphere, whereas Van Beneden and Julin hold that at a corresponding
| |
| stage (44-cells), sixteen ectodermal cells have been derived from the dorsal hemisphere. Of these sixteen cells four only are really ectodermal (the neural plate
| |
| cells), eight are mesodermal, and four are endodermal. Castle's conclusion that a
| |
| portion of the mesoderm is derived from the ventral cells is due to his erroneous
| |
| lineage of the cells after the 48-cell stage; all of the ventral cells are ectodermal,
| |
| and all of the mesoderm and endoderm are derived from the dorsal cells. With ,
| |
| the exception therefore of these four neural plate cells, which arise at the 44-cell
| |
| stage on the dorsal side of the third cleavage plane, all of the ectoderm lies on the
| |
| ventral side of that plane, and all of the endoderm and mesoderm on its dorsal side.
| |
| This conclusion, it will he observed, is very similar to that of Seeliger, Davidoff,'
| |
| ami Samassa.
| |
| | |
| 4. Fourth Cleavage; 8-i6 cells. (Figs. 36-38, 110-115, 186-188.)
| |
| The spindles for the fourth cleavage appear in all of the eight cells at about
| |
| the same time, though the dorsal cells sometimes divide slightly in advance of the
| |
| ventral ones. All the spindles are approximately horizontal in position, and all are
| |
| oblique to the median and transverse planes (first and second cleavage planes).
| |
| As a result of the fact, stated on page 45, that the dorsal hemisphere is broad in front
| |
| and narrow behind, while the ventral hemisphere is broad behind and narrow in
| |
| front, we find that the obliquity of the spindles of one hemisphere is reversed as
| |
| compared with that of the other. Thus the spindles in the anterior-dorsal cells
| |
| approach in direction a transverse plane, in the posterior-dorsal cells they approach
| |
| an antero-posterior plane ; whereas in the anterior-ventral cells they approach an
| |
| autero-posterior plane, while in the posterior-ventral cells they approach a transverse plane (figs. 110-113, 180, 187).
| |
| | |
| | |
| | |
| Corresponding with these positions of the spindles the subsequent divisions of
| |
| the cells are such as to lead to an inverse position of the cleavage cells in the
| |
| dorsal as compared with the ventral hemisphere. In the anterior-dorsal cells the
| |
| fourth cleavage furrows run from the anterior border of the cells to the transverse
| |
| (second cleavage) plane, and are approximately antero-posterior in direction: in
| |
| the posterior-dorsal cells these cleavage furrows run from the lateral borders of the
| |
| cells to the median (first cleavage) plane and are approximately transverse in direction. In the ventral hemisphere the reverse is the ease; in the anterior-ventral
| |
| cells the fourth cleavage furrows are approximately transverse in direction, in the
| |
| posterior-ventral cells approximately antero-posterior. Thus it comes about that
| |
| two of the anterior-dorsal cells do not reach the mid-line, while all of the posteriordorsal cells do; and that two of the posterior-ventral cells do not reach the midline, while all of the anterior-ventral ones do.
| |
| | |
| The fact that each hemisphere is thus the mirrored image of the other with
| |
| respect not only to the width of the anterior and posterior parts, but also as to
| |
| the direction of the fourth cleavage spindles and in the positions of the resulting
| |
| cleavage cells, this fact has contributed to the difficulties which most students of
| |
| ascidian embryology have experienced in distinguishing the dorsal and ventral
| |
| hemispheres, and has probably been responsible in some cases for the confusion of
| |
| those hemispheres. However, at this stage as at ever}- other, the two hemispheres
| |
| are easily distinguished by the relative amounts of yolk and protoplasm at the two
| |
| poles as well as by the position of the crescent and of the polar bodies.
| |
| | |
| All of the cell divisions of this cleavage are approximately equal, except that
| |
| of the posterior-dorsal cells, B 41 and B 41 . These cells divide very unequally, giving
| |
| rise to two small posterior cells, B 52 and B 52 , which are the smallest in the entire
| |
| egg (figs. Ill, 113, 180). Since the work of Van Beneden and Julin. these cells
| |
| have been observed by all who have studied the ascidian cleavage, and they have
| |
| served as the most important landmark in the orientation of the cleavage stages.
| |
| | |
| In this stage as in the preceding one. the yolk is most abundant in the cells
| |
| of the dorsal hemisphere; the protoplasm, in those of the ventral hemisphere; while
| |
| the yellow protoplasm is almost entirely confined to the posterior cells of the dorsal
| |
| hemisphere (figs. 36, 37, 38). In stained preparations the limits of the yolk and
| |
| protoplasm are sharp and distinct, and are represented in the drawings by a crenated line (figs. 108, et seq.). The relative amounts of yolk and protoplasm at the
| |
| two poles can he readily seen by comparing figure 110 with 111, and 112 with 113.
| |
| The yolk and protoplasm of the four ventral cells are about equally distributed to
| |
| their eight daughter cells; the same is true of the two anterior-dorsal cells, which
| |
| divide so that each of their four daughter cells contains about the same proportion
| |
| of yolk and protoplasm (A 51 , A 5 - 2 , Figs. 36, 37, 113). However, in the division of
| |
| the posterior-dorsal cells, the daughter cells are qualitatively very disimilar ; the
| |
| small posterior cells (B 5 - 2 ) consist almost entirely of yellow protoplasm, while their
| |
| larger sister cells (B 5 - 1 ) are about half and half, yellow protoplasm and yolk. The
| |
| outlines of the yellow protoplasm or crescent are perfectly distinct as shown in figure 37, and the formation of the .small posterior cells shows most beautifully that
| |
| the cleavage planes do not necessarily follow the lines of demarcation between the
| |
| yellow protoplasm and the yolk ; for in this case they cut across those lines so that
| |
| the small posterior cells contain a wedge of yolk in addition to the yellow protoplasm (figs. 37, 113, 117). This yolk is later obscured by being covered by the
| |
| yellow protoplasm (fig. 39, et seq.), but when the posterior cells are first formed it
| |
| is quite distinct. These small posterior cells contain not only yellow protoplasm
| |
| and yolk, but also those caps of clear superficial protoplasm which later go into the
| |
| small posterior mesenchyme cells. These cannot be seen in the living egg, but are
| |
| very evident in stained preparations (tigs. 113, 115, ISO, 187).
| |
| | |
| The localization of yolk and protoplasm at the vegetal pole is now practically
| |
| the same as at the beginning of gastrulation, and it is clearly indicative of the location of definitive organs. The relative positions of the yolk and yellow protoplasm
| |
| are the same in the 16-cell stage shown in figure 37, as in the early gastrula stage
| |
| shown in figure 46. The area of yolk, free from protoplasm, which surrounds the
| |
| vegetal pole (figs. 37. 111. 113), gives rise to the endoderm of the gastrula, the
| |
| tongue of yolk which runs back between the arms of the crescent (figs. 37. 113)
| |
| gives rise to the caudal endoderm cord of the larva, while the greater breadth of
| |
| the yolk in front of the second cleavage plane (fig. 37, 113) is indicative of the
| |
| great transverse extent of the endoderm of this region in later stages (fig. 4(i, ct
| |
| seq.). The protoplasm of the anterior-dorsal cells is located at the anterior borders
| |
| of those cells (figs. 37. 113). and in this region the notochord and neural plate cells
| |
| later arise. In all these respects the localization of these substances is of direct
| |
| prospective significance : in fact one may go further and say that all the regions of
| |
| the gastrula and certain important organs of the later larval stage are here
| |
| actually marked out on the egg at the i6-cell stage. This is no ideal mapping out
| |
| of the egg into organ forming germ regions, but an actual localisation of strikingly
| |
| different substances which need only to be followed through the development to
| |
| prove that they give rise to definite organs which occupy the same relative positions
| |
| in the larva, and are composed of the same peculiar substances, as in the early
| |
| cleavage stages or even in the unsegmented egg.
| |
| | |
| 5. Fifth Cleavage; i6- 3 2 cells. (Figs. 3'.>-42, 116-11'.). 1S'.)-195).
| |
| | |
| The fifth cleavage does not occur simultaneously in all the cells of the fifth
| |
| generation, but divisions appear in the cells of the vegetal or dorsal hemisphere
| |
| before they do in those of the animal or ventral hemisphere digs, llli-118. lS'.i.
| |
| I'M)). In Cynthia the anterior-dorsal cells divide a little earlier than the posteriordorsal ones (fig. 117). and the anterior-ventral cells a little in advance of the posterior-ventral ones (lii:. 118). In Cioua. also, the cells of the dorsal hemisphere
| |
| divide before those of the vental, but there is practically no difference in the time
| |
| of division of the anterior and posterior cells of this hemisphere. Neither at this
| |
| stage nor at any preceding or succeeding one are the cleavages more rapid or the
| |
| cells more numerous at the posterior than at the anterior pole, as claimed by Van Beneden and Juliti for Clavellina (1884, p. 13). Although this cleavage may be subdivided into 20-cell, 22-cell and 24-cell stages, the duration of each of those
| |
| stages is very brief, and the fifth cleavage is completed in all the cells before the
| |
| | |
| sixth appeal's.
| |
| | |
| Castle ( IS'.IC. p. 229), in particular, has described the differences in the time of
| |
| division of the cells of the dorsal and ventral hemispheres, and has made it a principal evidence in favor of his scheme of orientation. The fact that at this and at
| |
| the succeeding cleavage the cells of one hemisphere divide earlier than those of the
| |
| other has been accepted by him as proof that the earlier dividing hemisphere is
| |
| ventral and ectodermal, while the more slowly dividing one is dorsal and endodermal, since, at the time of gastrulation, the number of cells of the ectodermal
| |
| hemisphere is greater than that of the endodermal. But neither the fifth nor
| |
| the sixth cleavage results in the formation of more cells in one hemisphere than
| |
| in the other, since all the cells of both hemispheres divide before the next cleavage
| |
| begins; at the close of the fifth cleavage there are sixteen cells in each hemisphere,
| |
| and at the close of the sixth cleavage thirty-two cells in each hemisphere. In the
| |
| seventh cleavage, as we shall see. the hemisphere in which divisions were slower at
| |
| the two preceding cleavages becomes the more rapidly dividing one, and thereafter
| |
| the number of cells is more numerous in this hemisphere than in the opposite one.
| |
| | |
| In the anterior-dorsal cells the fifth cleavage spindles are parallel with the
| |
| median plane and are obliquely posterior-dorsal and anterior-ventral in direction
| |
| (fig. 117); four of the resulting daughter cells (A 6,2 , A 6 - 4 ) lie around the anterior
| |
| border of the egg just below the equator, while the other four (A 111 , A 6 - 3 ) form a rowacross the dorsal surface of the egg just in front of the second cleavage plane (fig.
| |
| 117). The former are composed of yolk and protoplasm in about equal parts, and
| |
| give rise to chorda and neural plate cells; the latter are rich in yolk, but have little
| |
| protoplasm and give rise to endoderm.
| |
| | |
| The four posterior-dorsal cells divide a little later than the anterior ones, and the
| |
| spindles lie approximately in a transverse direction (figs. 117, 189). The protoplasm
| |
| of these cells is chiefly crescent substance ; the small posterior cells (B 52 ) are almost
| |
| entirely composed of this substance, while the larger cells (B 3-1 ) are composed of this
| |
| substance and yolk in about equal proportions, the former occupying the outer half
| |
| of the cell and the latter the median half. These larger cells divide equally so as to
| |
| cut off all of this crescent substance and a small amount of yolk in the lateral
| |
| daughter cells and to leave hut little protoplasm and much yolk in the median ones
| |
| (figs. 37. 39). This division occurs at the 20-cell stage, and when it is completed all
| |
| of the mesodermal or crescent substance is finally and completely separated from
| |
| the endoderm, and, except for a small amount of yellow protoplasm which lies close
| |
| around the nuclei of many of the blastomeres, all the crescent substance is contained
| |
| in the four cells which form the posterior border of the dorsal hemisphere (figs. 39,
| |
| 40). The small posterior cells divide a little later than these larger ones and
| |
| unequally, the median daughter cells being smaller than the lateral ones (figs. 41,
| |
| \'i. 119). Thus there come to he six mesodermal cells, three on each side of the
| |
| mid-line, during this cleavage.
| |
| | |
| | |
| Divisions begin in the ventral hemisphere before they are finished in the dorsal (figs. 118, 19] ). Tn the most anterior and posterior pairs of cells of this hemisphere (a 5-3 , b 5-4 ) the spindles are nearly parallel with tlie median plane; in the two
| |
| remaining pairs of cells (a 5 - 4 and b 5-3 ) the spindles are oblique from posterior-ventral
| |
| to anterior-dorsal (figs. 118, 191). The division of the anterior pair of cells (a 53 ,
| |
| tins. 1 IS, 119) gives rise to a couple of cells (a 6-5 , a'"') which lie just above the equator and in contact with the chorda-neural-plate cells of the dorsal hemisphere.
| |
| Later development shows that these cells form part of the anterior portion of the
| |
| neural plate; the only other cells of the ventral hemisphere which enter into the
| |
| formation of this plate are portions of the cells a 6r , which lie on the lateral borders
| |
| of the cells a 6,5 . All of the cells of the ventral hemisphere are of practically the
| |
| same size and constitution ; each consists of a superficial layer of protoplasm, in
| |
| which the nucleus lies, and a deeper layer of yolk, the cells of this pole being
| |
| decidedly protoplasmic as compared with those of the opposite pole.
| |
| | |
| The result of this cleavage is the formation of sixteen cells in each hemisphere
| |
| which may be tabulated as follows :
| |
| Ventral hemisphere
| |
| | |
| 14 ectoderm cells, protoplasmic.
| |
| 2 neural plate cells, protoplasmic.
| |
| Dorsal hemisphere
| |
| | |
| 6 endoderm cells, yolk laden.
| |
| | |
| 4 chorda-nerve 1 cells, yolk and protoplasm.
| |
| | |
| 6 mesoderm cells, yellow protoplasm or crescent substance.
| |
| | |
| 32 cells.
| |
| | |
| At the (dose of this cleavage the cells of the ventral hemisphere are smaller in
| |
| superficial area than those of the dorsal hemisphere; when viewed from the ventral
| |
| pole the dorsal cells are seen around the entire periphery of the egg, except at a
| |
| point on the right and left sides where a single ventral cell (b liD , b 6-B ) occupies the
| |
| periphery; this is the only cell of the ventral hemisphere which can lie seen from
| |
| the dorsal pole (figs. 11!), 192, 193). A similar condition prevailed at the close of
| |
| the preceding cleaving (figs. 116, 117, 190), the only cells of the ventral hemisphere which could be seen from the dorsal pole being b 5:i and b 5 - 3 . This condition
| |
| may be traced still farther back to the 8-cell stage (figs. 108, 1 L0, L84) where the
| |
| ventral cells are smaller than the dorsal ones and where the only portion of the
| |
| ventral hemisphere which lies below the general plane of the equator is that part
| |
| of each of the posterior-ventral cells (b 4,2 , b 4 - 2 ) which meets the anterior-dorsal cell
| |
| in the cross furrow (figs. 108, 184).
| |
| | |
| At the close of the fifth cleavage the superficial area of the ventral cells is
| |
| smaller than at any preceding stage and that of the dorsal cells is greater; this is
| |
| due to a change in t lie shape of the cells, the ventral cells becoming long and col
| |
| 1 Throughout this paper the cells which are to give rise to chorda, nerve, muscle and mesenchyme are, for the sake of brevity, frequently referred to as if they had already given rise to these
| |
| structures.
| |
| | |
| | |
| | |
| umnar, while the dorsal cells become relatively broad and lint (text figs. XI. XII,
| |
| XIX. XX). This change of shape has been observed and commented upon bj
| |
| Van Beneden amilulin. Samassa, and Castle, and both of the latter authors attribute the columnar form of the ventral cells to the pressure exerted upon them by
| |
| the overgrowth of the cells of the dorsal hemisphere; both regard this overgrowth
| |
| as the beginning of gastrulation (epibole). Whatever may he the cause of the
| |
| shapes of the cells at the two poles, whether purely mechanical or not. it is certain
| |
| that this is not the beginning of gastrulation, since, as I will show later, the
| |
| columnar cells of this stage become the flattened ectoderm cells of Later stages,
| |
| while the flattened cells of this stage become the columnar endoderm cells of the
| |
| gastrula.
| |
| | |
| 6. Sixth Cleavage; 32-64 <*lk. (Figs. 4:5-45, 120-130, 194-197).
| |
| | |
| In this cleavage the divisions are not synchronous, the cells of the dorsal hemisphere dividing before those of the ventral as in the preceding cleavage, and some
| |
| of the cells in the posterior half dividing later than those in the anterior one.
| |
| Accordingly it would be possible to sub-divide the period between the 32-cell and
| |
| (14 -cell stages into a 44-cell, a 46-celI and a 48-cell stage, as Castle does. These
| |
| stages, however, are of brief duration and all the cells of the sixth generation
| |
| divide before any of the seventh do; therefore, the sixth cleavage is distinct from
| |
| preceding and succeeding ones.
| |
| | |
| The spindles appear in the four chorda-neural-plate cells at the anterior border
| |
| of the dorsal hemisphere in a nearly dorso-ventral direction. The four ventral produets of this division (A' 4 , A 78 ) form a band of small cells around the anterior border of the egg just dorsal to the equator; these cells ultimately give rise to the
| |
| posterior part of the neural plate; the dorsal products (A 7-3 , A 7 - 7 ) give rise to the
| |
| chorda (figs. 119-123). The neural plate cells are small and contain little or no
| |
| yolk, whereas the chorda cells are larger and are yolk-laden (text figs. XIX, XXI.
| |
| XXIII); this cleavage of these cells is therefore markedly differential.
| |
| | |
| While these cells are dividing, all of the endoderm cells divide; these are the
| |
| four median cells which meet at the vegetal pole (A" 1 . A" 1 . IV' 1 . ]\ M ). and a single
| |
| pair of cells which lie lateral to these and in front of the transverse (second cleavage) plane (A 6-3 , A 6 - 3 ). The spindles in the median cells are antero-posterior in
| |
| direction, while those in the lateral cells are nearly transverse (figs. 120, 121, 123).
| |
| These divisions are equal and non-differential in the median cells; in the lateral
| |
| cells the division is differential, the inner product (A 75 ) being rich in yolk, the
| |
| outer (A 7 - 6 ) containing more protoplasm : the former is an endoderm cell, the latter,
| |
| according to Castle, mesenchyme. By these divisions ten endoderm cells are produced, live on each side of the mid-line, and two mesenchyme cells (figs. 44-46).
| |
| | |
| While the divisions of the endoderm and chorda-neural-plate cells are occurring, the most anterior mesoderm cell (B 02 , B 1 '-). forming the point of the crescent
| |
| on each side, divides, the spindle lying in a nearly dorso-ventral direction (figs. 43,
| |
| 44, 120-129, 193, 194). This division, in fact, sometimes slightly precedes that of the anterior cells. The products of this division are nearly equal in size hut are
| |
| qualitatively dissimilar, the dorsal one (B 7,3 ) containing less of the yellow protoplasm and more yolk than the ventral one (B 74 ). This difference between these
| |
| daughter cells is plainly visible in the living condition, the dorsal cell being a
| |
| fainter yellow than the ventral one (figs. 43-48, et seg.); in preparations the dorsal
| |
| cells always stain more deeply than the ventral ones, owing to the greater quantity
| |
| of clear protoplasm which they contain (text fig. XXII). This difference in the
| |
| constitution of these cells corresponds to a difference in their fate; the dorsal cells
| |
| give rise to mesenchyme, while the ventral ones produce some of the muscle cells of
| |
| the tail of the tadpole.
| |
| | |
| The division of these twelve cells of the dorsal hemisphere are practically synchronous, and they advance the egg from the 32-cell to the 44-cell stage. A little
| |
| later the second cell of the crescent on each side of the mid-line (B 6 - 4 ) divides, its
| |
| spindle standing in a nearly dorso-ventral direction (figs. 45, 127-42'.>). The dorsal
| |
| daughter cell (B 77 ) in this case also contains less yellow protoplasm and more yolk
| |
| than the ventral one (B 78 ), and like the cell which immediately adjoins it anteriorly
| |
| (B 73 ) gives rise to mesenchyme, while the ventral moiety becomes a muscle cell.
| |
| By this division the mesenchyme and muscle substance of the crescent are finally
| |
| and completely segregated into separate cells, and the number of cells in the crescent is advanced to ten, and in the entire egg to forty-six. This division of the cell
| |
| B 64 is sometimes delayed until the cells of the ventral hemisphere are dividing
| |
| (figs. 127-129), and a 46-cell stage is therefore not always present. The division
| |
| of the last remaining cells of the dorsal hemisphere, the middle cells of the crescent
| |
| (B 63 B 63 ), is delayed until divisions are well advanced in the ventral hemisphere,
| |
| and it may occur even after the ventral cells have divided (fig. 47). I do not find,
| |
| therefore, that there is commonly a 48-cell stage such as Castle describes.
| |
| | |
| The divisions of the cells of the ventral hemisphere are all synchronous, as
| |
| figures 124 to 121) show. The direction of the spindles in the different cells is so
| |
| different that it is difficult to give an exact description of them. In the four median
| |
| cells which surround the animal pole (a 6-8 , b' is and their fellows of the right side)
| |
| the spindles are transverse; the spindles are also nearly transverse in the most
| |
| anterior and most posterior pairs of cells (a 65 . b a7 ); in the only other pair of cells
| |
| which meet along the mid-line, the second pair in front of the animal pole (a 65 ), the
| |
| spindles are nearly antero-posterior. In the other three pairs of cells of this hemisphere (a 6-7 , b 65 , b 66 ) the spindles are oblique in position, and their directions can he
| |
| best appreciated by consulting the figures {v. figs. 124, L96). The most anterior
| |
| pair of cells (a 6,5 ) are neural plate cells; these cells divide transversely (figs. 124126), forming a transverse band of four cells just above the equator; on each side
| |
| of these a single cell (a 7 - 13 , fig. L30) is added at the close of this cleavage which
| |
| completes the number of neural plate cells that are derived from the ventral
| |
| hemisphere. In figure 130 the hand of six cell (a 7 ' 9 , a 710 , a 713 and their fellows of
| |
| the right side) which lie around the anterior border of the ventral hemisphere are
| |
| these neural plate cells.
| |
| | |
| All the divisions of the cells of the ventral hemisphere are equal, and all the daughter cells are similar in appearance. By this cleavage the cells of the
| |
| ventral hemisphere are increased to thirty-two, and when the small posterior cells
| |
| (B 6,8 , B 6-8 ) of the dorsal hemisphere have divided there are thirty-two cells in this
| |
| hemisphere also, or sixty-four in the entire embryo, all in the seventh generation
| |
| (figs. 130, 131). Tabulating these facts we find that there are at the close of the
| |
| sixth cleavage the following cells :
| |
| Ventral hemisphere
| |
| | |
| 26 ectoderm cells, protoplasmic.
| |
| | |
| 6 neural plate cells, protoplasmic.
| |
| Dorsal hemisphere
| |
| | |
| Hi endoderm cells, yolk laden.
| |
| | |
| 4 chorda cells, yolk laden.
| |
| | |
| 4 neural plate cells, protoplasmic.
| |
| | |
| 4 mesenchyme cells, light yellow protoplasm.
| |
| | |
| 2 anterior mesenchyme cells, clear protoplasm.
| |
| | |
| 2 posterior mesenchyme cells, clear protoplasm.
| |
| | |
| 6 muscle cells, deep yellow protoplasm.
| |
| ti4 cells.
| |
| At the beginning of the sixth cleavage the cells of the ventral hemisphere are
| |
| narrow and columnar, while those of the dorsal hemisphere are broad and flat (text
| |
| figs. XI. XII, XIX, XX). This condition prevails up to the 44-cell stage when
| |
| the cells of the ventral hemisphere begin to divide. During their division the
| |
| ventral cells become shorter and broader, and at the same time the dorsal cells,
| |
| which have passed into a resting stage, grow more columnar and much smaller in
| |
| surface area, and before the close of this cleavage the cells at both poles are columnar and of about the same height (text figs. XIII. XIV, XXI). This change
| |
| in the shape of the cells of the two hemispheres, which begins during the sixth
| |
| cleavage, is not completed until the seventh cleavage of the ventral cells (figs. 133.
| |
| 134. 198-204). During this change of shape there is no difficulty in distinguishing the two hemispheres, for the endoderm cells are filled with yolk ami the mesoderm cells with yellow protoplasm or crescent substance, whereas the cells of the
| |
| ventral hemisphere are largely protoplasmic (text figs. XVII-XXIV). Moreover,
| |
| the polar bodies are often attached to the egg at its animal pole throughout the
| |
| whole of this period (cf. figs. 1U0-204).
| |
| | |
| I have already discussed the views of Van Beneden and Julin, of Samassa and
| |
| of Castle relative to the shape of the cells of the two hemispheres. Although Van
| |
| Beneden and Julin showed by their figures that the dorsal cells of the 32-cell stage
| |
| of Clavellina are fiat and the ventral ones columnar, whereas the dorsal cells of the
| |
| 44-cell stage are columnar and the ventral ones fiat, they did not observe nor
| |
| attempt to explain this change of shape. On the other hand, as we have seen,
| |
| Samassa and Castle denied that such a change of shape took place, and they therefore reversed Van Beneden and Julin's orientation of all stages before the 44-cell
| |
| stage. I have already given what seems to me satisfactory and sufficient evidence
| |
| in favor of the orientation of Van Beneden and Julin, and against that of Samassa
| |
| and Castle, and I need not repeat that evidence here.
| |
| | |
| | |
| | |
| Turning now to a detailed study of the observations of Samassa and Castle
| |
| during this critical sixth cleavage, we find that Samassa did not attempt to follow
| |
| the cell-lineage further than the 48-cell stage (his tig. 9), but jumped at once from
| |
| this stage to one with at least 76 cells (his fig. 10). His orientation of all stages up
| |
| to and including the 48-cell stage (his fig. 9) is the reverse of that of Van Beneden
| |
| and Julin, and is wrong; his orientation of the gastrula, shown in Lis figure 10, is
| |
| right. Therefore, in the interval between his figures 9 and 10 he has inverted the
| |
| egg so that the dorsal face of his figure 10 corresponds to the so-called ventral face
| |
| of all preceding figures. 1
| |
| | |
| Castle, on the other hand, has traced the cell-lineage much further than the
| |
| 48-cell stage, and it is therefore possible to follow in detail the manner in which lie
| |
| passes from the erroneous orientation of earlier stages to the correct orientation of
| |
| later ones. He has given correctly the lineage of every cell up to and including
| |
| the 46-cell stage (his figs. 55 and 5(i), as I have convinced myself by comparing his
| |
| figures, cell tor cell, with my own. but his orientation of these stages should be
| |
| reversed. On the other hand his orientation of all stages later than the 46-cell
| |
| stage is correct, but the cell lineage of these stages is wrong. This is due to the
| |
| fact that between the 46-cell and the 48-cell stages (his figs. 56 and 57) he has
| |
| inverted the egg so that the dorsal surface of all stages later than the 46-cell stage
| |
| corresponds with the so-called ventral surface of all earlier stages. 2 This inversion
| |
| of the egg introduces many profound errors in the cell-lineage after the 46-cell stage.
| |
| | |
| Considering in detail Castle's account of this sixth cleavage we find that be
| |
| has correctly represented the divisions of the cells of the real dorsal hemisphere
| |
| which bring these cells up to the seventh generation and the entire egg up to the
| |
| 46-cell stage (his fig. 55). At this stage the cells of the ventral hemisphere are
| |
| still in the sixth generation (:-. his fig. 56). and this stage is almost exactly comparable with my figures 1 1 9 to 1 23. Immediately after this, in the 4S-cell stage (his figs.
| |
| | |
| 57 and 58), Castle supposes that the cells of the real dorsal hemisphere, which are
| |
| now in the seventh generation, divide again, thus passing into the eighth generation,
| |
| while the sixth generation cells at the opposite pole are supposed by him to remain
| |
| undivided. It is absolutely essential to his scheme of orientation that the cells of
| |
| one hemisphere should remain in the sixth generation, while those of the other hemisphere are advancing to the seventh and eighth generation. If it could be shown
| |
| that all the cells of both hemispheres divide during this sixth cleavage it would completely break down Castle's orientation of the earlier stages and his cell-lineage of the
| |
| later ones. In all of his figures of this cleavage (figs. 55, 56. 57, 58) Castle represents
| |
| the cells at one pole in process of division while those at the other pole are in the
| |
| resting condition. However, in two of my figures of this cleavage in Ciona (figs.
| |
| | |
| 1 In his explanation of figures he says that figure '> is viewed from the cephalic pole; this is, .if
| |
| course, a verbal error, since his lettering of the cells shows plainly that the egg is viewed from the
| |
| caudal pole.
| |
| | |
| 2 Unfortunately Castle gives no dorsal, ventral nor lateral views of this critical 48-cell stage at
| |
| which the inversion occurs, but only an anterior and a posterior view (his tigs. 57 and 58, reproduced in
| |
| text tigs. XXV and XX VI (if this paper).
| |
| | |
| | |
| L 96 and 197), which represent ventral and dorsal views of one and the same egg,
| |
| the cells at both poles are seen to be in process of division, and the only cells in
| |
| the entire embryo which are not dividing are the small posterior cells (B 6-3 ). The
| |
| cells of the dorsal hemisphere are in the late anaphase or telophase, and their nuclei
| |
| are still small and densely chromatic; the cells of the ventral hemisphere are all in
| |
| the equatorial plate stage. These figures show most conclusively that all the cells of
| |
| the embryo divide during this sixth cleavage and are advanced from the sixth to
| |
| the seventh generation, and they therefore make impossible Castle's view that the
| |
| eel Is of the dorsal hemisphere remain undivided, while those of the ventral hemisphere divide twice. Another evidence that the cells which are shown dividing in
| |
| his figures 57 and 58 are not the same ones which have just divided in his figure 55
| |
| may he found in the fact that but sixteen of these cells are shown dividing in the
| |
| former figures, whereas the other sixteen cells, which, according to Castle, belong
| |
| to the ventral hemisphere, are in the resting stage, exactly as are the sixteen cells
| |
| which immediately surround the dorsal pole; at the two previous cleavages, and as
| |
| I have found also, at the two subsequent ones, all the cells of the ventral hemisphere
| |
| divide simultaneously, and this fact speaks against Castle's view that at the 48-cell
| |
| stage one-half of these cells divides ami the other half does not.
| |
| | |
| Since the dorsal hemisphere, shown in his figure 55, contains twenty-eight cells
| |
| of the seventh generation and two of the sixth, while the ventral hemisphere shown
| |
| in figure 56 contains only sixteen cells of the sixth generation, it is evident that if
| |
| the egg is inverted in its orientation at this stage the equator must be shifted nearer
| |
| to the dorsal hemisphere so as to reduce the number of dorsal cells to sixteen and
| |
| to increase the number of ventral cells to thirty, or, after the division of the two
| |
| small posterior cells, to thirty-two. This is just what Castle has done; in his
| |
| description of the 48-cell stage (pp. 238, 239) he says that at this stage the embryo
| |
| is composed of three zones of sixteen cells each, as follows :
| |
| Ventral hemisphere
| |
| | |
| 16 cells of the seventh generation, ectodermal group.
| |
| | |
| 16 cells of the seventh generation, equatorial hand.
| |
| Dorsal hemisphere
| |
| | |
| 16 cells of the sixth generation, endoderm, chorda and mesoderm.
| |
| | |
| 1 8 cells.
| |
| Immediately after this stage the 64-cell stage is reached by the division of the
| |
| sixteen cells of the ectodermal group. Castle has tabulated the cells of this stage
| |
| as follows :
| |
| | |
| Ventral hemisphere
| |
| | |
| 32 cells in the eighth generation, ectodermal group.
| |
| 16 cells in the seventh generation, the equatorial band.
| |
| | |
| 48
| |
| Dorsal hemisphere
| |
| | |
| 16 cells in the sixth generation.
| |
| lil cells.
| |
| | |
| As it can be proved that no cells of this stage remain in the sixth generation,
| |
| but all have passed into the seventh, it is certain that the equator both here and in
| |
| the 48-cell stage is in the wrong place, that it really lies between his equatorial
| |
| hand and the ectodermal group, and that there are therefore thirty-two cells in each
| |
| hemisphere in the 64-cell stage.
| |
| | |
| Wholly apart, therefore, from the perfectly conclusive evidence as to the orientation of the egg and embryo which may be drawn from the histological character
| |
| of the cells at the two poles, as well as from the location of the polar bodies, it can
| |
| lie shown b^' a detailed stud \ of the cell-lineage that Castle has inverted the egg at
| |
| the 48-cell stage, transferred sixteen cells from the dorsal to the ventral hemisphere
| |
| and consequently shifted the equator of the embryo at least one cell row nearer the
| |
| dorsal pole than it should be. Of course the lineage of every cell is thereby profoundly changed ; the oi
| |
| | |
| | |
| Figs. XXV and XXVI. Surface views of eggs of Cionn intesHnalis; copied from Castle's figures 5T and
| |
| 58 (1896). Fig. XXV represents an anterior view ; Fig. XXVI a posterior one of the same egg. Tlie orientation and cell-lineage, according to Castle, are indicated by the designations of the cells in that half of the egg
| |
| under his name; the designations of the corresponding cells in the other half of each figure shows the system
| |
| of orientation and cell-lineage adopted in this paper. Owing merely to differences in nomenclature the cells
| |
| in the right half of Fig. XXVI are designated by the letter D, those on the left by the letter B. Everywhere lower-ease letters designate cells of the animal (maturation) hemisphere; capitals, cells of the hemisphere
| |
| opposite the maturation pole. The equator lies between the cells designated by lower-case and capital letters.
| |
| | |
| | |
| | |
| | |
| | |
| | |
| lis which retain a semblance of their former names
| |
| | |
| | |
| | |
| throughout this revolution arc the small posterior cells (C 75 . D 75 of Castle's system).
| |
| and their sisters (C 7,li . D"' 1 ), the most anterior cells of the crescent of each side (C 74 ,
| |
| D 7 - 4 ), and the most anterior pair of cells of the dorsal hemisphere (A 7-4 , B 74 ). Even
| |
| in the case of these four pairs of cells the right and left cells of each pair are interchanged, so that everywhere A should replace B, and C, D.
| |
| | |
| In subsequent stages Castle does not always preserve the same designations for
| |
| given cells. For example, the cell which in his figure 58 is labelled A 7-8 becomes
| |
| a 6,7 in fig. 00 ; a 6-7 of figure 58 becomes d 6 - 5 of figure GO, while the one labelled d' ! - 5
| |
| in the former figure becomes A 7-6 in the latter. Strangely enough this last cell
| |
| which had been variously located in the dorsal and ventral hemispheres, and in the anterior and posterior quadrants is finally brought back to its right position and
| |
| given its true designation. In all stages later than his figure 60 the designation
| |
| A" stands for the same cell in Castle's figures and in my own.
| |
| | |
| The changes in the designations of the cells which arc brought about by this
| |
| inversion of the orientation at the 48-cell stage may be most easily seen and appreciated by a reference to the accompanying text figures XXV and XXVI, where the
| |
| designations of the cells, according to my interpretation, are given on the righl side
| |
| of figure XXV and on the left side of figure XXVI, while Castle's designations of the
| |
| corresponding cells are given on the Left side of figure XXV and on the right side
| |
| of figure XXVI. Barring the exceptions mentioned in the preceding paragraph.
| |
| Castle has followed with substantial accuracy the subsequent lineage of the dorsal
| |
| hemisphere up to a stage of about one hundred and twelve cells, though always
| |
| upon the basis of his erroneous lineage of the 48-cell stage. With the exception of
| |
| a single pair of cells, I need not further explain my departure from Castle's nomenclature of the later stages. This exception is the pair of small posterior mesenchyme cells which Castle designates C 7,5 , D 75 ; inasmuch as I find that they lie
| |
| ventral to their sister cells. I shall designate them B' 6 , B 76 , and their more dorsally
| |
| placed sister cells B 75 , J3 r ' 5 .
| |
| | |
| With the completion of the sixth cleavage we reach a period when the gastrulation is ready to begin. Already preparations for the gastrulation are apparent in
| |
| the changing shapes of the cells of the dorsal and ventral hemispheres, in the relative positions of the cells and in the directions of their divisions. Even the peculiar type of the chordate gastrula, with its overgrowing anterior lip and its nearly
| |
| stationary posterior one, is foreshadowed at a very early stage in the eccentric position of the animal and vegetal poles in the two hemispheres of the egg.
| |
| | |
| In the 32-cell and 64-cell stages it is apparent that the animal and vegetal poles
| |
| do not mark the middle of the ventral and dorsal faces of the embryo. This was
| |
| first noticeable in the 4-cell stage of Cynthia where the two posterior cells are
| |
| smaller than the anterior ones. In the 8-cell stage the anterior-ventral cells are
| |
| elongated antero-posteriorly. while the posterior-ventral ones are elongated transversely: this brings the animal pole still farther back of the middle of the ventral
| |
| face. In the 16-cell stage there are two pairs of cells adjoining the mid-line in
| |
| front of the animal pole and but one pair behind it ; in the dorsal hemisphere there
| |
| is one pair of such cells in front of the vegetal pole and two behind, but the most
| |
| posterior pair is smaller than the others, so that the vegetal pole lies near the middle of the dorsal face. In the 32-cell stage three pairs of cells adjoining the midline are in front of the animal pole, two behind it; while in the dorsal hemisphere
| |
| there are two pairs of such cells in front of the vegetal pole and two behind it.
| |
| though here again the most posterior cells are very small ones. In the 64-cell
| |
| stage (digs. 130, 131) there are four pairs of cells adjoining the mid-line in front of
| |
| the animal pole and only two pairs behind it; while in the dorsal hemisphere there
| |
| an- four pairs of such cells, both in front of and behind the vegetal pole, but the
| |
| most posterior pair are the tin\ mesenchyme cells (B 7 - 6 , B 7 - 6 ) which are partly covered by their sister cells (B 75 , B 75 ). Thus the vegetal pole is slightly posterior to
| |
| the middle of the dorsal lace and the animal pole is decidedly posterior to the middle of the ventral face in all of the stages mentioned, and this condition becomes
| |
| even more pronounced in later stages; thus in the 124-cell stage (fig. 139) there are,
| |
| ventral to the equator, six pairs of cells adjoining the mid-line in front of the animal
| |
| pole and four behind it. while in the 184-cell stage (fig. 143) there are eight pairs
| |
| of such cells in front of the animal pole and five behind it; in the 2lN-cell stage (fig.
| |
| 144-147) there are ten such pairs of cells in front of the animal pole and only five
| |
| behind it. All of the cells of the ventral hemisphere are of approximately the same
| |
| size, so that in these later stages it is evident that the animal pole lies far back of the
| |
| middle of the ventral hemisphere. This location of the animal pole posterior to the
| |
| middle of the ventral hemisphere is due in the first instance to the smaller size of
| |
| the posterior cells in the 4-cell stage and then to the fact that the prevailing position
| |
| of the spindles in the anterior cells of tins hemisphere is parallel with the median
| |
| plane, while in the posterior cells it is transverse. It is not due. as might at first
| |
| thought seem to be the case, to the more rapid growth and division of the anterior
| |
| cells of the ventral hemisphere since all of these cells divide at nearly the same
| |
| time and are of approximately the same size. The irrespective significance of this
| |
| eccentric location of the animal pole maybe found in the greater length of the anterior lip of the blastopore, as compared with the posterior lip.
| |
| | |
| | |
| C. Gastrulation ; Seventh to Ninth Generation of Cells, 64-218 Cells.
| |
| | |
| | |
| In both Ciona and Cynthia the gastrulation actually begins during the seventh
| |
| cleavage and it is far advanced by the close of the eighth, though the closure of the
| |
| blastopore and the completion of the gastrulation does not occur until about the end
| |
| of the tenth cleavage. 1 have followed the lineage of every cell through the seventh
| |
| cleavage and of almost all the cells through the eighth, and have therefore been able
| |
| to determine the part which each cell takes in the formation of the gastrula. At
| |
| no time after the 64-cell stage are all the cells of the embryo in the same generation.
| |
| From this time toward the endoderm cells lag behind the ectoderm and mesoderm
| |
| cells in division; the eighth cleavage occurs in the ectoderm and mesoderm before
| |
| the seventh is finished in the endoderm. Therefore the periods of the seventh and
| |
| eighth cleavages cannot be sharply separated, but for the sake of convenience we
| |
| shall consider these two cleavages as if they were distinct.
| |
| | |
| 7. Seventh Cleavage ; 64-76, 76-112 cells. (Figs. 46-51, 130-139. 198-204.)
| |
| The seventh cleavage begins in the anterior quadrants of the dorsal hemisphere
| |
| in the two pairs of chorda cells (A 7:i , A 77 ) and in the two pairs of neural plate cells
| |
| (A 74 , A 78 ); in the posterior quadrants it begins in the two most anterior cells of the
| |
| m\scent on each side, the pair of muscle cells, B 74 . and the pair of mesenchyme
| |
| cells, B 73 (figs. 130-132). With the exception of the two mesenchyme cells the
| |
| spindles in all of these cases are parallel with the plane of the equator and with the
| |
| surface of the egg; in the mesenchyme cells the spindles, when seen from the dorsal side, are directed obliquely forward, outward and ventralward (figs. 131, 132).
| |
| These divisions, when completed, give rise in the anterior quadrants to four chorda
| |
| cells (A 8 - 5 , A 8 - 6 , A 818 , A. & "), and to four neural plate cells (A 87 , A 8 - 8 , A. 815 , A 818 ) on
| |
| each side of the mid-line, which are arranged in two rows of eight cells each
| |
| running around the anterior border of the dorsal hemisphere (figs. 134, 200). All
| |
| these divisions are equal and non-differential.
| |
| | |
| In the posterior quadrants the division of the anterior muscle cell on each side
| |
| gives rise to two daughter cells (IT'. I'> ss ). one in front of the other, which are alike
| |
| in size and quality. The mesenchyme cell (B 7r ) divides unequally and differentially
| |
| giving rise to a small yolk-laden cell (B 88 ), lying anterior, lateral and ventral to its
| |
| large sister cell (B 8 '). which is more protoplasmic (figs. 134. 200). The former is,
| |
| according to Castle, the "posterior chorda fundament" while the latter is mesenchyme. I have been unable to find sufficient evidence that this small cell is later
| |
| incorporated in the chorda, but on the other hand do not wish to deny that this is
| |
| the case. Since it is derived from the mesenchyme I prefer to class it with the
| |
| mesenchyme cells until its fate is more certainly known. These divisions advance
| |
| the embryo to the 7G-cell stage and the distribution and generations of the cells
| |
| may be summarized as follows :
| |
| Ventral hemisphere
| |
| | |
| Ectoderm 7th generation 26 cells.
| |
| | |
| Neural plate 7th " G "
| |
| | |
| Dorsal hemisphere
| |
| | |
| Endoderm 7th " 10 "
| |
| | |
| Chorda . . . . 8th generation 8 cells.
| |
| | |
| Neural plate 8th " 8 "
| |
| | |
| . Muscle . . 8th " 4 " 7th " 2
| |
| | |
| Mesenchyme 8th " 4 " 7th " 8
| |
| | |
| | |
| | |
| a
| |
| | |
| | |
| | |
| sth generation 24 cells. 7th generation 52 cells.
| |
| | |
| | |
| | |
| | |
| | |
| | |
| o
| |
| | |
| | |
| | |
| 76 cells.
| |
| The classification of the cells of this stage into chorda, neural plate, muscle
| |
| and mesenchyme. 1 must of course be based upon the later history of these cells, but
| |
| even at this early stage important differences may be recognized in the histological
| |
| characters of the cells named. In the living eggs of Cynthia the endoderm cells
| |
| are slate-gray in color and are filled with yolk; the chorda cells are lighter
| |
| gray and contain less yolk than the endoderm cells; the ectoderm and neural
| |
| plate cells are clear and protoplasmic ; while the muscle and mesenchyme cells
| |
| are yellow, the former being more deeply colored than the latter. In the main
| |
| my classification of the cells of this stage agrees with that of Castle; the most
| |
| important difference concerns the muscle cells which he classes as " neuro-muscular
| |
| cells." Owing to the striking color of these cells in Cynthia their later history can
| |
| be followed with relative ease; they ultimately give rise to the three rows of muscle cells in the tail of the tadpole (figs. 58, 59), and so far as 1 am able to determine do
| |
| not contribute anything to the nervous system. In general the mesenchyme cells
| |
| may also be traced by their faint yellow color until they give rise to the mesenchyme of the tadpole, and, with the exception of the small cell which Castle calls
| |
| the posterior chorda fundament," I agree with him as to the fate of these cells.
| |
| Castle figures and describes the cell, A 70 , as the most anterior of the mesenchyme
| |
| cells. 1 do not find that it contains yellow protoplasm in Cynthia, but its histological structure is different from that of the other endoderm cells; I shall therefore
| |
| follow Castle in classing it as a mesenchyme cell. The median cells of the crescent
| |
| (B 7S ) resemble in their deep yellow color, the muscle cells rather more than the
| |
| mesenchyme, and Castle reckons these cells with the " neuro-muscular ring." but
| |
| the later history of these cells shows that they lie just beneath the notochord and
| |
| at the hinder end of the ventral endodermal cord in the tail of the tadpole (figs.
| |
| 161-165) ; therefore, they do no give rise to the lateral muscles, and they are probably to be counted as mesenchyme cells.
| |
| | |
| | |
| See note p. 52.
| |
| | |
| | |
| | |
| | |
| At this stage, therefore, the endoderm consists of four pairs of cells meeting
| |
| along the mid-line (figs. 134, 200), and of one pair of laterally placed cells which lie
| |
| just in front of the second cleavage plane ; the chorda consists of an arc of eight cells
| |
| bounding the endoderm in front; in front of the chorda cells and below the equator
| |
| is an arc of eight neural plate cells. Posteriorly the endoderm is bounded by an
| |
| arc of twelve mesenchyme cells, while just outside these is an arc of six muscle
| |
| cells (eight, in lig. 134). The neural arc in front is separated from the muscle arc
| |
| behind by the most dorsally situated of all the ectoderm cells (b 817 ). But for this
| |
| lateral interruption it would be possible to speak of a " neuro-muscular ring" as
| |
| Castle does. The chorda and mesenchyme arcs form a continuous chorda-mesenchynie ring, as Castle has shown.
| |
| | |
| Castle asserts (1896, p. 246) that the mesenchyme "is made up of cells
| |
| derived from both hemispheres and all four quadrants," and again that two cells
| |
| " vis. d' '- and c 712 [my B 85 ] are the sole contribution of the dorsal hemisphere to
| |
| the mesoderm of the larva" (p. 242). This is certainly not the case; the mesenchyme and muscle cells are derived entirely and exclusively from the dorsal hemisphere and largely from the posterior quadrants. The most posterior cells of the
| |
| crescent on each side (B S1,; and its mate) are counted b\ Caslle as part of the ectoderm; their histological structure, color in the living egg of Cynthia and later
| |
| history show that they are the most posterior of the muscle cells. Of two other
| |
| cells of tbis stage, Castle says (p. 242). " it is noticeable that d 65 and its mate c 65
| |
| have been shoved forward out of their own quadrants to a position beside the endoderm cells derived from the anterior quadrants." These cells are really A 75 and
| |
| its mate, as is shown by their origin and later history (figs. 120, et set/.), and do not
| |
| belong to the posterior but to the anterior quadrants.
| |
| | |
| The 76-cell stage is of very short duration, for immediately after those divisions in the dorsal hemisphere which advance the embryo from the 64-cell to the
| |
| 76-cell stage, all the cells of the ventral hemisphere divide simultaneously. The direction of the spindles in these cells is indicated by the equatorial plates represented in figures L98 to 203. In the posterior quadrants all the spindles are
| |
| nearly antero-posterior in direction, except in those three ventral cells on each side
| |
| (b 7,9 , b rj ". W "i which lie nearest to the dorsal surface and between the muscle cells
| |
| behind and the neural plate cells in front (figs. 201, 203); in these cells the spindles are nearly vertical. In the anterior quadrants the spindles are antero-posterior in a transverse row of four cells which lies just in front of the animal pole
| |
| (a 7-16 , a'''i: in the row of four cell just in front of this the spindles are transverse
| |
| (a ;i \ a' "l: they are also transverse in a single pair of cells which meet at the midline just in front of the last mentioned row (a'"); in the most anterior row of the
| |
| ventral hemisphere, consisting of six neural plate cells (a 7 - 10 , a 7,9 , a 7-13 ), the spindles
| |
| are dorso-ventral in position, therefore, at the close of this division there are
| |
| twelve neural plate cells in the ventral hemisphere, arranged in two rows of
| |
| six cells each.
| |
| | |
| All of these divisions of the cells of the ventral hemisphere are synchronous,
| |
| equal and non-differential, and they increase the number of cells in the ventral
| |
| hemisphere to sixty-four and bring the whole number of cells in the embryo up to
| |
| one hundred and eight.
| |
| | |
| Very soon after these divisions in the ventral hemisphere the posterior muscle
| |
| cell (B 7-8 ) and one of the mesenchyme cells (B 7,7 ) of each side divide; in the former
| |
| the spindles converge posteriorly and ventrally toward the median plane, in the
| |
| latter posteriorly and dorsally. By this division two mesenchyme and two muscle
| |
| cells are added to the total in the embryo which at this stage consists of one bundred and twelve cells (fig. 133. 134 1 ) which may be tabulated as follows :
| |
| Ventral hemisphere
| |
| | |
| Ectoderm . .8th generation, 52 cells.
| |
| | |
| Neural plate 8th generation, 12 cells.
| |
| Dorsal hemisphere '
| |
| | |
| Endoderm 7th gen., 10 cells.
| |
| | |
| Chorda .... 8th generation, 8 cells.
| |
| | |
| Neural plate . 8th " 8
| |
| | |
| Muscle .... 8th 8 "
| |
| | |
| Mesenchyme 8th " 8 " 7th " 6 "
| |
| | |
| 8th generation, 96 cells. 7th gen., lb cells.
| |
| | |
| 112 cells.
| |
| The only cells in the entire embryo which have not passed into the eighth
| |
| generation at this stage are the ten endoderm cells, and six mesenchyme cells, two
| |
| of which are anterior, and four median and posterior: all of these cells except two,
| |
| the small posterior mesenchyme cells ( l? 7 !. divide soon after this stage and thus
| |
| pass into the eighth generation, but not until after other cell- have passed into
| |
| the ninth. This stage, therefore, max be taken as representing, as nearly as may
| |
| be found, the close of the seventh cleavage ami the transition to the eighth.
| |
| | |
| 'The division in a pair of mesenchyme cells B 7-7 ) of this stage i> not completed; therefore, in
| |
| the explanation of figures, tin- is called a LlO-cell stage.
| |
| | |
| | |
| | |
| i,l ORGANIZATION AND CELL-LINEAGE OF ASCIDIAN EGG.
| |
| | |
| This stage is important as marking the beginnings of gastrulation for which
| |
| preparations were made in preceding stages. The endoderm cells and the four
| |
| posterior mesenchyme cells which remain in the seventh generation now lie at a
| |
| considerably lower level than the surrounding cells. This, although usually spoken
| |
| of as an invagination, can scarcely with right be called such, for as sections show
| |
| there is neither at this stage, nor at any preceding one, any considerable blastocoel ;
| |
| since there is no cavity into which the cells can push it is scarcely permissible to
| |
| speak of their invaginating. In reality the gastrulation is due to two factors,
| |
| neither of which is invagination. The first and most important is the change of
| |
| shape of the cells, which has been described in part already; the second is the
| |
| overgrowth of the cells him:' around the endodermal area.
| |
| | |
| As to the first we have already seen that in the 16-cell stage the cells at both
| |
| poles are of about equal height ; during the fifth cleavage the cells at the animal
| |
| pole become long and columnar, while those at the vegetal pole are broad and flat;
| |
| during the sixth cleavage the cells at the two poles change shape so that at the
| |
| close of this cleavage (64-eell stage) the cells of both poles are of nearly equal
| |
| height, those at the vegetal pole being perhaps slightly longer than those at the
| |
| animal pole. At this stage the cells of the dorsal (vegetal) hemisphere still have a
| |
| larger surface area than those of the ventral (animal) hemisphere, so that when
| |
| viewed from the vegetal pole only cells of the dorsal hemisphere^ can be seen, but
| |
| when viewed from the opposite pole a peripheral row of dorsal cells can be seen
| |
| around those of the ventral hemisphere (figs. 130, 131). During the seventh
| |
| cleavage this change of shape progresses rapidly so that at the 76-cell stage the
| |
| surface area of the dorsal cells is less than that of the ventral ones ; the endoderm cells in particular grow long and narrow, whereas the ectoderm cells
| |
| become broad and fiat (figs. 198-204). After the seventh cleavage of the ectoderm and mesoderm cells (112-cell stage) a row of ventral hemisphere cells is
| |
| visible all around the periphery of the dorsal hemisphere when the embryo is
| |
| viewed from the dorsal pole (figs, loo, 134). The remarkable reduction in the
| |
| surface area of the endoderm cells, which occurs without any division in these cells,
| |
| and wholly by their change of shape will be best appreciated by comparing figures
| |
| I'll with 134, and figures 197 with 200; in all of these figures the endoderm cells
| |
| are in the seventh generation, but the superficial area of these cells in the two older
| |
| stages is not more than half as great as in the two earlier ones. In proportion as
| |
| these cells decrease in surface area they increase in depth, their inner ends become
| |
| enlarged, and at the same time their nuclei are withdrawn from the surface to a
| |
| deeper level in the cells (text figs. XXI-XXIV). The flattening of the ectoderm
| |
| cells leads to their covering a larger and larger surface area until they finalty overgrow the marginal cells of the dorsal hemisphere. Samassa undertook to explain the
| |
| columnar form of the cells at the animal pole in the 32-cell stage by the pressure
| |
| exerted on them by the overgrowing cells of the opposite pole, but it is obvious
| |
| that neither at this early stage, nor during the later one just described, can the
| |
| cause of this change of shape be located in the cells of one hemisphere rather than
| |
| in those of the other.
| |
| | |
| | |
| The second factor of the gastrulation, viz., the overgrowth of the cells surrounding the endodermal area has been well described by Castle. It is the result
| |
| in part of the first factor and also of the more rapid division of the cells of the ectoderm and the corresponding retardation of division in the endodermal cells. This
| |
| overgrowth occurs in the anterior quadrants from in front, the chorda cells overgrowing the endoderni and the neural plate cells the chorda : in the posterior quadrants it occurs from the sides, the muscle cells overgrowing the mesenchyme, and
| |
| the ectoderm the muscle cells. At three points this overgrowth is long delayed, at
| |
| the posterior pole where there is a dee]) notch In the blastoporic rim which persists
| |
| until the blastopore has nearly closed, and at the light and left sides of the blastopore where the overgrowth is slow. This leads to the peculiar form of blastopore,
| |
| wide in front and narrow behind, which is found among ascidians.
| |
| | |
| | |
| 8. Eighth Cleavage; 1 12-132 cells, 132-218 cells. (Figs. 135-147, 205).
| |
| | |
| The eighth cleavage first appears in the two anterior muscle cells of each side
| |
| ( B 88 , B 87 ), the spindles being nearly transverse to the antero-posterior axis of the
| |
| embryo (figs. 135, 136). This division is equal and non-differential, and there result
| |
| four muscle cells on each side, an anterior pair (B 915 , B 916 ) and a posterior pair (B 913 ,
| |
| B 914 ). When first formed the median cells of each of these pairs lie at a higher
| |
| level than the lateral ones (fig. 135) ; soon afterward the lateral and median cells
| |
| are at the same level (fig. 136); still later the lateral ones lie at a higher level
| |
| than the median ones (fig. 140). This is, of course, a result of the overgrowth,
| |
| whereby the cells which were lateral in the blastopore lip come to overly those
| |
| which were median in position.
| |
| | |
| | |
| At the same time that these muscle cells are dividing the pair of large mesenchyme cells, B 85 , divides, the spindles being obliquely antero-posterior and dorsoventral in direction (figs. 135, 136). This division is approximately equal and
| |
| non-differential, and gives rise to the mesenchyme cells B", .B 910 , which lie on each
| |
| side of the caudal endoderm cells.
| |
| | |
| | |
| While these divisions are proceeding in the mesoderm, and thereby advancing
| |
| these cells to the ninth generation, the delayed seventh cleavage appears in the
| |
| mesenchyme and endoderm cells. The first of these cells to divide is the most
| |
| anterior mesenchyme cell (A 7 - 6 ) ; the- spindles are here nearly dorso-ventral in
| |
| direction, and the resulting daughter cells (A su , A 8 - 12 ) are of about the same size,
| |
| though the dorsal cell contains more protoplasm than the ventral one, as Castle has
| |
| shown.
| |
| | |
| Coincidently with these divisions two pairs of endoderm cells (A 72 and B 72 )
| |
| divide, the spindles being approximately transverse in the anterior pair and anteroposterior in the'posterior one (fig. 135).
| |
| | |
| A little later the four endoderm cells which meet at the vegetal pole (A 71 , B 71 , fig.
| |
| 1 36) divide, the spindles being antero-posterior in direction. The last remaining pair
| |
| of endoderm cells of the seventh generation to divide is the lateral one (A 75 , figs. 135,
| |
| 136) ; I have not seen this cell in division, but it is probable from a study of later stages that the spindle lies in it also in an anteroposterior direction. All of these
| |
| divisions of the endoderm are equal and non-differential-.
| |
| | |
| Finally the median mesenchyme cells, B 75 . divide, the spindles being anteroposterior (fig. 136), and the resulting daughter cells alike in size and quality.
| |
| | |
| With this division all the cells of the embryo have passed into the eighth
| |
| generation, except the small posterior mesenchyme cells (B 7 - 6 ) which never again
| |
| divide, so far as I have observed ; and excepting eight muscle cells and four mesenchyme cells which have passed into the ninth generation. The following tabular
| |
| statement summarizes the character and location of the cells at the close of this stage:
| |
| Ventral hemisphere
| |
| | |
| Ectoderm 8th gen., 52 cells.
| |
| | |
| Neural plate 8th " 12 "
| |
| | |
| Dorsal hemisphere
| |
| | |
| Endoderm 8th " 20 "
| |
| | |
| Chorda 8th " 8 "
| |
| | |
| Neural plate 8th " 8 "
| |
| | |
| Muscle .... 9th gen., 8 cells. . . 8th " 4 "
| |
| | |
| Mesenchyme. 9th " 4 " ...8th " 14 " 7th gen., 2 cells.
| |
| | |
| 9th gen., 12 cells. 8th gen., 118 cells. 7th gen., 2 cells.
| |
| | |
| 132 cells.
| |
| | |
| The 132-cell stage is not a sharply defined one. for before all the divisions
| |
| which have been described above have been finished, other divisions are begun
| |
| which lead to the 184-cell stage (figs. 136-143). The cells which divide first in
| |
| this period are the four median neural plate cells A 87 , A 88 (fig. 136) ; shortly afterward the four lateral ones A 815 , A 816 (fig. 140), also divide. The spindles in all
| |
| these cells lie in a radiating position around the blastopore, and as a result of this
| |
| division there are produced in the dorsal hemisphere two rows of neural plate cells,
| |
| eight cells in a row, situated at the anterior border of the blastopore and dorsal to
| |
| the chorda cells.
| |
| | |
| About the same time forty-four of the fifty-two ectoderm cells divide ; the
| |
| spindles are approximately transverse in all these cells, except in the most posterior
| |
| row of the ventral hemisphere, where they are dorso-ventral, and in two transverse
| |
| rows of four cells each, which are the third and fourth rows in front of the animal
| |
| pole (figs. 139, 143), where the spindles are antero-posterior in direction.
| |
| | |
| By these divisions the dorsal neural plate cells are increased to sixteen, and
| |
| the ectoderm to one hundred and eight cells, so that at this stage the entire embryo
| |
| contains one hundred and eighty-four cells. Twenty cells, forming two rows often
| |
| each around the anterior border of the embryo just ventral to the equator, remain
| |
| undivided for some time and are conspicuous for the large size of their resting
| |
| nuclei and their more deeply staining cytoplasm (figs. 138, 142). The four hindmost of these cells on each side belong to the posterior quadrants (b 8-17 , b 818 , b 819 ',
| |
| b 8?0 ) ; the other six pairs (a 8-25 , a 826 , a 8 - 17 , a 818 , a 8-19 , a s ' 20 ) which form the median anterior portions of these rows, are derived from the anterior quadrants, and they
| |
| form the anterior portion of the neural plate.
| |
| | |
| Finally in the stage shown in figures 144 to 147. these twenty cells divide, all
| |
| the spindles being approximately dorso-ventral in direction. About the same time
| |
| the eight chorda cells divide, the spindles standing in a dorso-ventral direction (fig.
| |
| 1 Km. and a little later the six most anterior endoderm cells (A s -, A 83 , A 8,4 ) divide,
| |
| the spindles being nearly transverse (fig. 147). These thirty -four divisions advance
| |
| the embryo from 1S4 to 218 cells.
| |
| | |
| Beyond this point I have not attempted to follow the lineage of each and
| |
| every cell; this could be done successfully only by a most exacting study of serial
| |
| sections in connection with whole preparations. With sufficient labor and material
| |
| I believe that the lineage of every cell could be traced through to the tadpole stage,
| |
| but I have lacked both the time and the material for such a study. Tabulating
| |
| the cells of this stage we find that there are :
| |
| Ventral hemisphere
| |
| | |
| Ectoderm . . . 9th gen., 104 cells.
| |
| | |
| Neural plate . 9th " 24 "
| |
| Dorsal hemisphere
| |
| | |
| Endoderm. . . 9th " 12 " . . . . 8th gen., 14 cells.
| |
| | |
| Chorda 9th " 16 "
| |
| | |
| Neural plate . 9th " 16 "
| |
| | |
| Muscle 9th " 8 " .... 8th " 4 "
| |
| | |
| Mesenchyme . 9th " 4 " . . . . 8th " 14 " 7th gen., 2 cells.
| |
| | |
| 9th gen., 184 cells. 8th gen., 32 cells. 7th gen., 2 cells.
| |
| | |
| 218 cells.
| |
| At this stage the gastrula is nearly circular in outline when viewed from the
| |
| dorsal or ventral pole ; the gastrocoel is a cavity deep and wide in front and narrow behind, where it opens to the exterior through a deep groove between the
| |
| mesenchyme and muscle cells of each side. The endoderm lies at a deep level, in
| |
| contact with the ectoderm of the ventral side; in the anterior quadrants it consists
| |
| of nine pairs of cells which become the gastric endoderm of the larva, in the posterior quadrants are four pairs of endoderm cells which meet along the median
| |
| plane; the two posterior pairs become the caudal endoderm of the tadpole, the two
| |
| anterior pairs are added to the gastric endoderm. Three pairs of cells at the hinder
| |
| end of this cord of caudal endoderm are mesenchyme, while on each side of these
| |
| and running forward lateral to the caudal endoderm are seven pairs of large mesenchyme cells (in fig. 147 only five pairs are visible, since two lie at a deeper level,
| |
| one beneath the cell B 814 and another beneath A 8 - 12 ). The muscle rudiment consists
| |
| of six pairs of large cells, dorsal to the mesenchyme, on each side of the blastopore
| |
| en >ove and still uncovered by ectoderm. The chorda consists of sixteen cells arranged
| |
| in two rows of eight each, one ventral to the other, and covered superficially by
| |
| cells of the neural plate. Sixteen cells of the neural plate which cover the chorda cells .and form the anterior border of the blastopore belong to the dorsal hemisphere ; the rest of this plate is composed of cells of the ventral hemisphere,
| |
| arranged in four rows of six each, and lying just in front of the chorda region ;
| |
| therefore in the 218-cell stage the nerve plate consists of forty cells arranged in six
| |
| transverse rows, each containing six cells except, the two most posterior rows which
| |
| contain eight each. The animal pole, with the polar bodies still attached, is still
| |
| situated back of the middle of the ventral face, six cell rows posterior to the
| |
| anterior edge of the neutral plate ; there are but five rows of cells of the ventral
| |
| hemisphere posterior to the animal pole, while there are ten such rows in front
| |
| of it.
| |
| | |
| | |
| V. LATER DEVELOPMENT.
| |
| | |
| My observations on the later stages of development agree in the main with
| |
| those of previous workers and particularly with those of Castle, who made a thorough
| |
| study of these stages by means of serial sections. It is true of these stages, however, as it is of the cleavage stages, that many topographical relations can be made
| |
| out more satisfactorily by a study of entire preparations. I have therefore
| |
| devoted especial attention to such preparations, and my observation, both on living
| |
| and on stained material, ai*e embodied in plates V and X.
| |
| | |
| 1. Closure of Blastopore.
| |
| | |
| During the closure of the blastopore the embryo changes shape and at the
| |
| same time the egg axis is shifted. This stage is therefore an important one in
| |
| the orientation of the later stages. The gastrula is at first disk-shaped (fig. 134), it
| |
| then becomes saucer-shaped (fig. 136) and then cup-shaped (figs. 144, 145). During
| |
| this change as the embryo increases in depth it decreases in its other dimensions so that it becomes more nearly spherical (fig. 145). The closure of the
| |
| blastopore takes place more rapidly from the anterior than from the posterior
| |
| side ; in fact after the general drawing together of the margins of the saucershaped gastrula the posterior lip remains nearly stationary until the last stage in
| |
| the closure of the blastopore.
| |
| | |
| Soon after the 218-cell stage the gastrula becomes elongated and egg-shaped,
| |
| the posterior end being somewhat narrower than the anterior. The anterior lip of
| |
| the blastopore continues to grow 'posteriorly while the lateral lips draw nearer
| |
| together; thus the blastopore becomes T-shaped (fig. 148), and finally, by the
| |
| further growth of the anterior lip, the anterior part of the blastopore, represented
| |
| by the bar of the T, is covered and the blastopore is reduced to a longitudinal
| |
| groove between the lateral lips (figs. 152, 153). In the growth of the lateral lips
| |
| they come to lie at a higher level than the anterior lip. and consequently as the
| |
| latter continues to. grow posteriorly, the former are tilted up at their anterior
| |
| ends until they become vertical in position. These lateral lips are at first formed
| |
| only of the muscle cells, but later the ectoderm cells completely overgrow them.
| |
| | |
| | |
| In this process the ectoderm does not, for some time, close up the notch at the posterior end of the blastopore (fig. 52); this is one of the last steps in the process
| |
| of closure.
| |
| | |
| The overgrowth of the anterior lip continues until it has covered about threequarters of the dorsal face; meanwhile the animal pole is shifted nearer to the
| |
| point of greatest curvature at the anterior end. and the blastopore is transported
| |
| from the dorsal side toward the posterior end. In this process the rows of muscle
| |
| cells which at an earlier stage stretched from the posterior pole to the second
| |
| cleavage plane, and were antero-posterior in direction, are tilted up at their anterior
| |
| ends and pushed backwards until they lie at the hinder end of the embryo and run
| |
| in a dorso-ventral direction (figs. 52, 53, 50, 157). This complete change in the
| |
| direction of the rows of muscle cells I found most perplexing and difficult to understand. In early stages the crescent, and the mesoderm cells which form from it. lies
| |
| just below the equator of the egg, and in the anteroposterior plane; in these later
| |
| stages the muscle cells are transverse to the anterior-posterior axis. A detailed
| |
| study of intermediate stages shows how this change is brought about. After the
| |
| closure of the anterior part of the blastopore, corresponding to the bar of the T.
| |
| the anterior lip does not overgrow the blastopore groove (stem of the T) and its
| |
| lateral walls, which are composed of the muscle cells; on the other hand, these
| |
| lateral walls lie at a higher level than the anterior lip, and the continued growth
| |
| of this lip pushes the muscle cells and the groove before it. As the posterior lip
| |
| remains stationary during this process it happens that the entire dorsal portion of
| |
| the posterior quadrants is tilted up in front and pushed backward until it forms the
| |
| posterior end of the embryo, the posterior lip becoming vental and the anterior lip
| |
| dorsal in position. Thus the blastopore groove, which lay on the dorsal side posterior to the middle, comes to lie at the* posterior end of the embryo and the walls
| |
| of the groove, containing the muscle cells, come to be terminal in position and vertical in direction (figs. 50-53). The mesenchyme cells, as well as the caudal
| |
| endoderm. lie at so low a level that they are not disturbed by the overgrowth of the
| |
| anterior lip, consequently the rows of these cells still preserve an antero-posterior
| |
| direction (fig. 157). Thus the mesenchyme and muscle cells, which in earlier
| |
| stages lay side by side, come to be separated anteriorly, and only remain in contact with one another at the hinder end of the strand of caudal endoderm cells;
| |
| the mesenchyme cells in this region are derived from the median part of the crescent and they ultimately become separated from the remaining portion of the
| |
| mesenchyme which comes to lie in the trunk (figs. 161-167).
| |
| | |
| These general changes in the shape of the embryo at this stage are accompanied by divisions of many of the cells, some of which we may now consider. In
| |
| all the ectoderm cells the ninth cleavage is nearly synchronous ; in the posterior
| |
| quadrants the spindles are approximately antero-posterior in direction, and the
| |
| same is true for the two hindmost rows of the anterior quadrants, but in most
| |
| of the other cells of the anterior quadrants the spindles are transverse, thus it
| |
| happens that the animal pole is shifted forward (fig. 149). As compared with the eighth cleavage there is therefore a regular alternation in the direction of
| |
| division in most of the cells. 1 have not observed the tenth cleavage of these
| |
| cells, but it seems probable that the direction of division is, in many of the cells,
| |
| the same as at the ninth cleavage, if one may judge by the longitudinal rows
| |
| of cells as well as by the number of rows which are present in the posterior
| |
| quadrants (figs. 155, 160). The animal pole is, therefore, shifted still further
| |
| forward during this cleavage. The posterolateral ectoderm cells slowly overgrow
| |
| the muscle cells, but for a long time they do not overgrow the median posterior
| |
| mesenchyme cells, and there is therefore left the deep notch in the blastopore at
| |
| the hinder end of the embryo which has already been described.
| |
| | |
| The neural plate in the stage shown in figure 148 consists of six transverse
| |
| rows of cells, only four of which show in the figure. The two posterior rows are
| |
| derived from the dorsal hemisphere and consists of eight cells each; the four anterior rows consist of six cells each, anil are derived from the ventral hemisphere.
| |
| In subsequent divisions of the posterior rows of this plate the spindles are anteroposterior in direction, thus adding to the number of rows but not to the number of
| |
| cells in each row; for example, in figure 152 there are eight rows of cells, but
| |
| apparently only six cells in each row. 1
| |
| | |
| The endoderm cells of the anterior quadrants divide chiefly in a transverse
| |
| plane; those of the posterior quadrants in an antero-posterior plane (figs, 150,
| |
| 151, 156). This fact, taken in conjunction with the direction of division in the
| |
| ectoderm cells, contributes to the lengthening of the posterior part of the embryo
| |
| and to the widening of the anterior part, and consequently to the shifting of the
| |
| animal pole further toward the anterior end.
| |
| | |
| Of the two rows of chorda cells established at the eighth cleavage one has
| |
| come to lie posterior to the other, and both bend so as to become horse-shoe-shaped
| |
| (fig. 153). Later these cells divide again (fig 157) and, pushing backward with the
| |
| anterior lip. carry the muscle cells before them, as already described.
| |
| | |
| In the 218-cell stage there were twenty mesenchyme cells: in the next stage
| |
| shown (fig. 150) these are increased by one or two divisions so that there are twentytwo or twenty-four cells. As in the preceding stage, they still lie on the ventral
| |
| side next to the ectoderm and along the posterior border of the gastral endoderm.
| |
| In figure 150 only one row of mesenchyme cells is found lateral to the caudal endoderm ; in figures 154 and 15G there are two such rows. In all these figures there
| |
| are three pairs of mesenchyme cells at the hinder end of the caudal endoderm ; the
| |
| most posterior of these is the small posterior mesenchyme cells (B 76 ), the others are
| |
| B 89 and B 810 . All of these cells are protoplasmic, stain deeply and art' strikingly
| |
| different in appearance from the endoderm cells.
| |
| | |
| The muscle cells, which in the 218-cell stage consisted of six pairs of cells, are
| |
| shown in figures 51 and 153 increased to eight pairs which are arranged in two
| |
| rows on each side of the blastopore groove. By the continued growth of the anterior lip these rows are tilted up into a dorso-ventral direction. An optical section
| |
| of the caudal region at this static shows four muscle cells on each side, one above
| |
| another (fig. 158) ; a lateral view shows three rows of muscle cells with four or five
| |
| cells in each row (figs. 56, 157). I have not observed the exact manner in which
| |
| this change from two rows to three takes place, but it is evident that it must be
| |
| associated with the division of the cells of the original two rows. In figure 51
| |
| there are eight muscle cells on each side; in figure 55. thirteen ; in figure 56,
| |
| fourteen; in figure 157. fifteen ; in figure 165, eighteen; and in figure 59, twenty;
| |
| therefore each of the cells shown in figure 51 must have divided once and some
| |
| of them twice during the period represented by these figures.
| |
| | |
| | |
| 1 In this figure it is possible that a single row of cells on each side of the stippled area should be
| |
| reckoned as part of the neural plate.
| |
| | |
| | |
| | |
| | |
| | |
| 2. Development of Larva (Figs. 57-60, 160-167).
| |
| | |
| After the growth of the anterior lip has carried the notochord to a position approximately corresponding to that of the blastopore groove in figure 153
| |
| and has shifted the rows of muscle cells into a nearly vertical direction (figs.
| |
| 157. 158), these rows of muscle cells again come to be antero-posterior in direction (figs. 58, 59, 161-165). This change takes place rather suddenly and I have
| |
| not observed all the steps in the process. It seems probable, however, that
| |
| it is due to two factors; (a) the depression of the dorsal ends of the muscle
| |
| rows to a position alongside of the notochord, and (b) the outgrowth of the
| |
| tail of the larva from the region of the ventral ends of the muscle rows. This
| |
| outgrowth, which is associated with the lengthening of the ventral side of the
| |
| embryo, carries the caudal mesenchyme cells and the ventral ends of the muscle
| |
| rows backward into the tail and thus the rows of muscle cells again assume an
| |
| anteroposterior direction (figs. 58, 59, 161-167). Usually six cells are seen in
| |
| each row and in addition there are two or more cells at the hinder ends of
| |
| these rows which do not fall specifically into any one of them. In livingembryos the muscle and mesenchyme cells retain their yellow color and the
| |
| individual muscle cells may be plainly seen ; all the figures shown in plate V
| |
| represent camera drawings of living embryos and in all of them the yellow cells
| |
| were distinctly visible as drawn. 1 When seen from the caudal pole (fig. 60), the
| |
| three rows of muscle cells are seen to be only one-layered and the cells of one side
| |
| are connected with those of the other by a group of small yellow cells (the caudal
| |
| mesenchyme), "which lie ventral to the notochord at its hinder end.
| |
| | |
| In stained preparations of young tadpoles these caudal mesenchyme cells can
| |
| be seen to consist of two or three pairs of cells at the posterior end of the caudal
| |
| endoderm and ventral to the notochord (figs. 101-165). The other mesenchyme
| |
| cells, which in a former stage (figs. 156, 157) were continuous with this caudal
| |
| group, are now separated from it by the whole length of the muscle rows. These
| |
| mesenchyme cells at the anterior ends of the muscle rows are found in later stages cells ; in figures 161-165 they. consist of eight or ten cells on each side. Whether
| |
| there may be a few scattered mesenchyme cells between the caudal and the trunk
| |
| groups and ventral to the muscle rows must still be left an open question, but there
| |
| can be no doubt that most of the mesenchyme cells are located in these two groups.
| |
| The separation of the caudal from the trunk mesenchyme must have been accomplished in part by the same means which brought the muscle rows from a vertical to
| |
| a horizontal position, viz., by the outgrowth of the tail. In addition there seems to
| |
| have been an actual forward movement of the trunk mesenchjme, as is indicated
| |
| by a comparison of such figures as 156 and 161. This is probably part of the general
| |
| forward shifting of the animal pole. In later stages when the tail is bent toward
| |
| the ventral side, the trunk mesenchyme is found ventral to the anterior ends of the
| |
| muscle rows (figs. 59, 166, 167). In these later stages the mesenchyme cells are
| |
| frequently found dividing; they are smaller and more numerous than the muscle
| |
| cells and are more than one cell-layer thick.
| |
| | |
| | |
| | |
| * Since this paper was sent to press Misses I'd. it and Strobell have prepared for me a series of
| |
| more than thirty photomicrographs of the living eggs and embryos of Cynthia. These photographs
| |
| show in the most striking manner the yellow protoplasm and the cells which arise from it ; even in the
| |
| tadpole stage these individual cells are plainly recognizable in the photographs.
| |
| | |
| | |
| in the trunk of the larva, and they may therefore be known as trunk mesenchyme
| |
| | |
| | |
| In the formation of the larva the ventral cord of endoderm increases greatly in
| |
| length, being composed in very young tadpoles (fig. 161) of six or seven pairs of cells.
| |
| These cells form a double row between the muscle cells of each side and ventral to
| |
| the notochord. In front of the caudal endoderm and notochord lies the gastral
| |
| endoderm consisting of yolk cells which form a single but rather irregular layer
| |
| around a small central cavity, the enteron (figs. 161, 162, 164-166).
| |
| | |
| In young larva 1 the chorda cells are wedge-shaped and form two or more rows
| |
| of cells which interdigitate, as has been described by previous writers. In the
| |
| latest stage which I have studied (fig. 167) these cells interdigitate to such an extent
| |
| that the}- form a single row of disk-shaped or slightly wedge-shaped cells. I have
| |
| not followed in detail the method by which the two arcs of chorda cells shown in
| |
| figure 153 are transformed into the double row shown in figure 162, but I see no
| |
| reason to question the account given of this by Van Beneden and Julin and also
| |
| by Castle.
| |
| | |
| The neural plate grows backward with the notochord nearly to the posterior
| |
| end of the embryo. I can find no evidence in favor of the view that any portion
| |
| of the nervous system is derived from cells which bound the blastopore groove posterior to the neural plate (figs. 152, 153), nor is there any evidence for the existence
| |
| of a nerve ring surrounding the blastopore. Since the neural plate, six or eight
| |
| cells wide at its hinder end, is carried back with the chorda nearly to the hinder
| |
| end of the embryo where the last trace of the blastopore is found (fig. 53), and since
| |
| no portion of the nerve cord is found posterior to the blastopore and notochord
| |
| (figs. 163, et seq.), it seems most probable that the hinder portion of the nerve
| |
| cord, as well as all the rest of it, is derived from the neural plate and not from
| |
| the lateral lips of the blastopore groove. That the muscle cells do not give
| |
| rise to the posterior part of the nerve cord, as claimed by Castle, is made probable
| |
| by the fact that this portion of the nerve cord is not yellow, as are the muscle
| |
| cells: I cannot therefore accept without further evidence Castle's statement that the posterior portion of the nerve cord is formed from the muscle cells (his "neuromuscular" cells). Furthermore, I am unable to find satisfactory evidence that
| |
| the ectoderm which covers the muscle cells and closes the blastopore notch behind
| |
| contributes to the formation of the nerve cord. Therefore, it is probable that
| |
| the entire central nervous system conies from the neural plate, which is a portion
| |
| of the anterior lip of the blastopore.
| |
| | |
| After having overgrown the muscle cells and closed up the posterior notch
| |
| of the blastopore the ectoderm forms a pair of Y-shaped folds (figs. 52-54), the
| |
| apex of the V lying just behind the blastopore and the limbs diverging anteriorly
| |
| and laterally. By the forward extension of these folds the neural plate is rolled
| |
| up into a tube which is covered with a layer of ectoderm, in the manner characteristic of vertebrates. These folds are at first V-shaped, but after they have extended
| |
| around the anterior end of the nerve plate the}' inclose an oval area which is pointed
| |
| behind (fig. 55). The folds close from behind forward and ultimately convert the
| |
| entire neural plate into a tube, which retains a lumen in its anterior portion (the
| |
| sense vesicle) and an opening to the exterior (the neuropore). but which contains no
| |
| lumen back of the anterior end of the notochord (figs. 16G, 167). That portion of
| |
| the nervous system dorsal to the notochord and which contains no lumen is derived
| |
| from those neural plate cells which belong to the dorsal hemisphere and which
| |
| in origin ivere intimately associated with the chorda cells ; the anterior half
| |
| of the enlarged portion of the tierve tube lying in front of the notochord (sense
| |
| vesicle) is derived from those cells of the neural plate which belong to the ventral
| |
| hemisphere. As tiearly as I can determine the anterior end of the neural plate lies
| |
| about jo above the original equator oj the egg and 6o below the anitnal pole.
| |
| The cephalic pole of the larva lies ventral to the anterior end of the neural plate
| |
| but dorsal to the animal pole ; therefore, the antero-posterior axis coincides neither
| |
| with the egg axis nor with the equatorial plane but lies mid-way between the two.
| |
| The egg axis is therefore not dorso-ventral in the larva but is, strictly speaking,
| |
| postero-dorsal and antero-ventral in direction. Inasmuch as the forward shifting
| |
| of the animal pole by which this position of the axes is brought about occurs at a
| |
| late period in the development, and also for the sake of simplicity of expression I
| |
| have, in accord with all my predecessors, described the egg axis as dorso-ventral in
| |
| direction in all the early stages.
| |
| | |
| | |
| | |
| VI. COMPARISONS WITH AMPHIOXUS AND AMPHIBIA.
| |
| | |
| The remarkable differentiations apparent in the egg and early cleavage stages
| |
| of ascidians. the relatively small number of cells present during gastrulation and
| |
| organogeny, and the comparative ease and certainty with which the axial relations
| |
| of the egg and embryo can he determined at all stages. these conditions render
| |
| the ascidian egg the most favorable in the whole phylum of the chordata for an
| |
| exact study of the early development. In no other chordate has the cell-lineage
| |
| been followed in detail up to the formation of definitive organ cases, and no where in the phylum has it been possible to determine with the same degree of certainty as here the relations of the axes of the egg to those of the gastrula and larva.
| |
| | |
| | |
| It is therefore worth while to compare the early development of ascidians
| |
| with that of other primitive chordates in order to see what light may thereby he
| |
| thrown on certain disputed problems. It must, of course, be understood from the
| |
| beginning that such comparison can have the weight only of suggestion ; the problems winch have been raised in the study of any group can be solved only by the
| |
| further study of that group, but comparisons with other forms may be of great
| |
| service. If evolution be true, if ascidians are genetically related to other chordates.
| |
| then it must be true that their modes of development are related. Whether the
| |
| mode of development of ascidians as compared with Amphioxus and amphibians is
| |
| palingenetic or coenogenetic is largely a matter of opinion, and need not concern us
| |
| here so only it be granted that there is a relationship between these classes in the
| |
| matter of their development as well as in their later structure.
| |
| | |
| Klaatsch (1896) has attempted to elucidate certain disputed points in the
| |
| develojnnent of Amphioxus by a comparison with the ascidians, proceeding upon
| |
| the principle that it is well to reason from the relatively known to the relatively
| |
| unknown, from conclusions in which all agree to questions upon which there is
| |
| diversity of opinion. Samassa (1898), on the other hand, holds that the ascidian
| |
| ontogeny has been so greatly shortened and modified as compared with that of
| |
| Amphioxus that it would be much better to explain the former by the latter than
| |
| the reverse. All this might be true without destroying the value of comparison,
| |
| but when Samassa further proceeds, as he does in the following sentence, to deny
| |
| that there is any relationship between the two forms except in a single stage, he
| |
| takes away all basis of comparison except for that single stage. He says, p. 20,
| |
| " Nun ahnelt aber die Ascidienentwicklung der des Amphioxus nur in dem einen
| |
| Stadium, wenn der Urmund geschlossen ist, der Chorda nach hinten auswachst und
| |
| die Organe der Larva die fiir Wirbelthiere characteristische gegenseitige Lagerung
| |
| ziegen . . . Bis zu diesem Stadium ist aber die Entwicklung des Amphioxus und
| |
| der Ascidien so verschiedenen wie mdglich." We have here, if I understand
| |
| Samassa correctly, homologies which are found only in a single stage of the ontogeny, which have had no beginnings in homologous parts or processes, have neither
| |
| homological antecedents nor consequents and have therefore arisen de novo. This.
| |
| it seems to me. is the logical conclusion to be drawn from Samassa' s statement, and
| |
| it is one as indefensible on zoological as on philosophical grounds. There are many
| |
| points of resemblance in the early development of Amphioxus and ascidians. as is
| |
| well known, and such differences as exist are explicable on the general principle of
| |
| evolution through divergent modification.
| |
| | |
| The study of the cell-lineage and early development of a large number of
| |
| annelids and mollusks has shown that in such general matters as the relations of
| |
| the axes of the egg to those of the gastrula and larva, and the origin of the germ
| |
| Layers and of specific organs from certain blastomere or regions of the egg, there is
| |
| a high degree of uniformity among members of the same phylum and even among
| |
| related phyla, ft would certainly be surprising if the development of Amphioxus
| |
| and the ascidians should he found to be more dissimilar than that of annelids
| |
| and gasteropods.
| |
| | |
| | |
| | |
| 1 . Axial Relations of Egg and Embryo.
| |
| | |
| In considering the axial relations of egg and embryo one is confronted at once
| |
| with the difficulty of determining what is meant by the anterior pole, unless it be
| |
| defined in terms of structure rather than function. The animal pole is a structurally definite point, hut the anterior end of the embryo, who can say what it is?
| |
| In the early development of Amphioxus and ascidians the point which at one time
| |
| is most anterior does not continue such for any considerable period, and it is practically impossible to determine the exact point of this rounded anterior portion of
| |
| the embryo which will become the most anterior part of the body. Not only is the
| |
| animal pole a structurally definite point but the anterior limit of the neural plate
| |
| is also, and the relative positions of these two can be determined with considerable
| |
| accuracy. The result of such a determination shows that there is great similarity
| |
| among the lower Chordata in that the anterior limit of the neural plate is always
| |
| some distance retnovedfrom the animal pole. In ascidians the chief axis of the egg is
| |
| plainly marked out not only by the well differentiated cleavage cells but also by the
| |
| polar bodies which in some cases remain attached to the egg at the point of their
| |
| formation until the blastopore has closed. In the early gastrula the animal pole is
| |
| slightly posterior to the middle of the ventral hemisphere, the vegetal pole marks
| |
| the middle of the dorsal hemisphere, and the deepest point of the gastrocoel (text
| |
| fig. XXVIII). In the closure of the blastopore the anterior lip overgrows the
| |
| archenteron. and the blastopore, being closed from in front backwards, is finally
| |
| limited to a longitudinal groove in the posterior half of the dorsal face of the
| |
| gastrula.
| |
| | |
| The relation of the egg axis to the embryonic axis is not a simple one, i. e.,
| |
| they neither coincide nor is one at right angles to the other. During the overgrowth of the anterior lip the animal pole is shifted nearer to the anterior end of
| |
| the gastrula. This may be, and probably is in part, due to a shifting of the point
| |
| of greatest curvature at the anterior end to a point nearer the animal pole. The
| |
| anterior edge of the neural plate never reaches farther forward than about onethird of the way from the equator to the animal pole, and consequently the animal
| |
| pole lies on the ventral side of the larva but near the head end. Correspondingly
| |
| the opposite pole of the extended egg axis lies near the posterior end of the dorsal
| |
| side and consequently not far from the place where the last trace of the blastopore
| |
| can be seen.
| |
| | |
| Previous students of ascidian embryology, and particularly Castle and Samassa,
| |
| have considered that the egg axis was dorso-ventral and hence perpendicular to the
| |
| embryonic axis. I at first held the same opinion, but observations on the change
| |
| of shape of the gastrula and particularly upon the anterior limit of the neural
| |
| plate during the closure of the blastopore have convinced me of the truth of
| |
| the position here taken (cf text figs. XX VII-XXIX).
| |
| | |
| The axial relations are not so evident in Amphioxus and amphibians, since
| |
| the animal and vegetal poles are not so clearly marked as in the ascidians. Hatschek (lSSl ) supposed that the animal pole of the egg in Amphioxus was ventral to the cephalic pole of the embryo; and this view has been supported by
| |
| Garbowski (1898); on the other hand. Kowalevsky (1867) and many recent
| |
| writers on the development of Amphioxus (Lwoff 1894, Klaatsch 1896, Samassa
| |
| 1898, Morgan and Hazen 1900). have considered that the most highly arched
| |
| portion of the late gastrula represents the animal pole. Since this point is said
| |
| to become the anterior end of the embryo it is evident that according to this
| |
| view the chief axis of the egg coincides with the chief axis of the embryo and
| |
| is antero-posterior in direction, whereas in ascidians it has heretofore been claimed
| |
| that the egg axis is dorso-ventral in direction and hence perpendicular to the
| |
| same axis in Amphioxus.
| |
| | |
| Sueh diversity in this most fundamental of all axial relations seems very
| |
| improbable considering the many points of resemblance between these groups,
| |
| and at least sueh conflicting results should be supported by the best of evidence
| |
| before being given general credence.
| |
| | |
| Korschelt and Heider in their excellent text book attempt to harmonize these
| |
| differences in axial relations between Amphioxus and the ascidians by regarding
| |
| the anterior pole of the ascidian gastrula as the animal pole, but I agree with
| |
| Samassa (1894). and Castle (1896), that the animal pole never comes to lie at the
| |
| anterior end of the embryo, though unlike them I hold that it does move in that
| |
| direction.
| |
| | |
| In Amphioxus as in the ascidians the anterior limit of the neural plate is
| |
| situated some distance behind the most highly arched portion of the gastrula,
| |
| and even if the latter be regarded as the animal pole it would still be true of
| |
| Amphioxus as of the ascidians that the neural plate does not reach as far forward
| |
| as the animal pole. But there are reasons for thinking that the animal pole lies
| |
| ventral to the most highly arched portion of the Amphioxus gastrula. Many investigators agree that the animal pole lies opposite the blastopore ; Samassa has observed
| |
| in a small percentage of eggs that the polar body is still attached to the embryo at
| |
| a time when the blastopore is growing smaller, and in all sueh cases he found it at
| |
| t be pole opposite the blastopore (although, as he maintains, at the anterior end of
| |
| the embryo). But the point opposite the blastopore lies ventral to the most highly
| |
| arched portion of the embryo. Even if it should be assumed that both ventral and
| |
| dorsal lips grow equally, the animal pole would still be located on the ventral side
| |
| of the most highly arched portion, owing to the peculiar shape of the embryo; if
| |
| the dorsal lip grows more rapidly than the ventral, which in the light of what
| |
| takes place in ascidians and amphibians seems probable, the animal pole must lie
| |
| still farther toward the ventral side. In any event a considerable space must intervene between the anterior limit of the neural plate and the animal pole.
| |
| | |
| The work of Garbowski (1898), shows that the longitudinal axis of the larva
| |
| of Amphioxus forms an angle of about 70 with the gastrular axis. a result which,
| |
| like that of Eatschek and Sobotta, agrees very closely with my observations on
| |
| ascidians. and which practically removes the supposed discrepencies in axial relations between these two classes.
| |
| | |
| | |
| | |
| On the whole it seems to me that then' is every reason for believing that the
| |
| relations of the egg axis to the embryonic axis are essentially the same in Amphioxus and ascidians, that in both the egg axis is postero-dorsal and antero-ventral
| |
| in direction and that in neither does the neural plate extend more than one-third
| |
| of the way from the equator to the animal pole (cf. text figs. XXVII-XXXII).
| |
| | |
| It' the same axial relations exist in amphibians as in ascidians, the middle
| |
| of the pigmented hemisphere of the frog's egg does not correspond to the cephalic
| |
| pole of the embryo but lies ventral to this pole, while the white hemisphere corresponds in the main to the dorsal side. This is approximately the orientation which
| |
| has been maintained by Pfiuger, Roux. Morgan, Kopsch and H. V. Wilson. Kopsch
| |
| | 1 '.'00), in particular, has shown that the anterior margin of the neural plate lies
| |
| some distance below the animal pole, and judging from his figures the axial relations
| |
| in the embryo of the frog must be almost identically like those in the ascidian {cf
| |
| text tigs. XXXlII-XXXYh
| |
| | |
| | |
| 2. Entrance of Spermatozoon.
| |
| | |
| Among ascidians the sperm enters the egg near the vegetal pole ; it then moves
| |
| to the posterior pole where it meets the egg nucleus, and the sperm amphiaster is
| |
| formed at right angles to the copulation path. The outer pigmented layer of protoplasm collects around the sperm nucleus and moves with it to the posterior pole
| |
| where the mesodermal crescent is formed.
| |
| | |
| In Amphioxus the sperm also enters near the vegetal pole according to Sobotta
| |
| (1897), hut whether it then moves to the posterior pole and whether there is a
| |
| collection of superficial protoplasm around the sperm nucleus is unknown.
| |
| | |
| In the frog the sperm enters on the posterior side just below the equator and.
| |
| according to Roux. the point of entrance determines the posterior pole of the embryo.
| |
| Schultze, on the other hand, thought that the point of entrance lay at the anterior
| |
| pole, hut since he also with Roux holds that the entrance occurs at the pole opposite that at which gastrulation begins, it is evident that this difference with regard
| |
| to the pole of entrance is only part of the larger difference between these authors as
| |
| t<> the general orientation of the embryo. The conditions which are found in the
| |
| ascidian ess closely auree with the orientation of Roux as against that of Schultze.
| |
| | |
| In another important respect Roux's observations find a parallel in the ascidian
| |
| __ : he observed that after fertilization the pigment cap of the fro.i;'- egg shifts so
| |
| that its margin lies below the equator on the side of the egg where the sperm
| |
| enters while at the opposite pole it comes to lie above the equator. I believe that
| |
| this movement of the pigment is comparable to the movements of the layer of
| |
| yellow protoplasm in the egg of Cynthia.
| |
| | |
| ?> . Cleavage.
| |
| | |
| There are many differences in the cleavage of the egg in these three classes
| |
| of chordates, hut some fundamental characteristics are essentially similar in all of
| |
| them. The most important of these is that the cleavage is usually bilaterally symmetrical. The first cleavage always coincides with the median plane among
| |
| ascidians, and every subsequent cleavage is perfectly bilateral, one-half of the egg
| |
| being the mirrored image of the other. In the frog's egg the first cleavage usually
| |
| lies in the plane of symmetry 1 , and although the subsequent cleavages grow more
| |
| and more irregular, bilaterality is sometimes strongly expressed even in the later
| |
| stages [cf. M. Schultze, 1863; Rauber. 1882).
| |
| | |
| In Amphioxus, if I correctly understand Wilson (1893, p. 600), the first cleavage
| |
| coincides with the median plane. In the subsequent cleavages, both Wilson and
| |
| Samassa (1898) have been unable to find the remarkably regular alternation of
| |
| meridional and latitudinal cleavages described by Hatschek. These cleavages are
| |
| extremely variable in form; among them Wilson recognizes three principal types,
| |
| one radial and two bilateral. After the 16-cell stage, however, almost all the eggs
| |
| become bilateral, whereas in the 8-cell stage three-fourths of them are radial.
| |
| Wilson suggests that variations from the bilateral type may occur among ascidians,
| |
| but I agree with Castle and Samassa that under normal conditions this is never the
| |
| case. In the 8-cell, 16-cell and 32-cell stages of the bilateral types there are many
| |
| striking resemblances to corresponding stages of the ascidian ; this applies particularly to Wilson's bilateral type II (cf. his figs. 13-18, 33. 34, 36, 37-30, 41-43, and
| |
| Samassa' s figs. 2, 6, 7, 9). In these figures the form of the cleavage is so similar
| |
| and the position of the cells and even the direction of the spindles within the
| |
| cells so remarkably like what is found in the ascidians that the individual cleavage
| |
| cells can be correlated in these two animal classes.
| |
| | |
| Too little is known of the cell-origin of the germ layers in At>iphio.\us to
| |
| determine accuratelv how close is the likeness to ascidians in this regard.
| |
| Wilson holds that the eight animal cells of the 16-cell stage are purely ectodermal
| |
| and that the "secondary macromeres " (A 2 , B 2 , C 2 , D 2 , ) which surround, and were
| |
| derived from, the four basal cells at the vegetal pole are of mixed character,
| |
| giving rise to both endoderm and ectoderm, and perhaps also mesoderm. He does
| |
| not give the evidence upon which this conclusion rests, but its similarity to the conditions which exist in the ascidians should not be overlooked. Here also the
| |
| eight animal cells are purely ectodermal, while the " secondary macromeres." and
| |
| in fact, all the cells of the vegetal hemisphere in the 16-cell stage are of mixed
| |
| character, the four anterior ones containing endoderm and ectoderm (neural plate
| |
| substance), and the four posterior ones, endoderm and mesoderm. Wilson expressly
| |
| states that he uses the terms macromere and micromere "solely for the sake of convenience," and he concludes that the cleavage is very unlike that of annelids ;
| |
| Samassa also emphasizes this same conclusion.
| |
| | |
| We may conclude, then, that there are certain fundamental resemblances between Amphioxus and ascidians in the matter of cleavage and that the most notable
| |
| differences between them are found in the number of cells and the degree of their
| |
| differentiation at any given embryogenic stage ; in ascidians this number is relatively small ami the degree of differentiation high as compared with Amphioxus ;
| |
| e.g.. at the stage when invagination begins in Amphioxus there arc according to
| |
| Wilson about 512 cells, at a corresponding stage in Ciona there are 76 cells. It
| |
| may be presumed that the relative constancy or variability of cleavage in these
| |
| two classes depends upon the two features just contrasted, viz.. the number of the
| |
| cleavage cells and the deirree of their differentiation.
| |
| | |
| | |
| | |
| 1 In the newt, Diemyctylus, Jordan (1893) found that the first cleavage is perpendicular to the
| |
| median plane.
| |
| | |
| | |
| | |
| | |
| | |
| In a general way the same kinds of likenesses ami differences exist between
| |
| ascidians and amphibians in the matter of cleavage as between the former and . Xmphioxus. Among amphibians, however, these differences are further increased
| |
| by the presence of a relatively large quantity of yolk. Whether the ectoderm
| |
| comes entirely from the four upper cells of the 8-cell stage in these animals cannot
| |
| be affirmed, but it is evidently derived in chief part from these cells.
| |
| | |
| | |
| 4. Blastula and Gastrula.
| |
| | |
| The firm of the blastula and gastrula is much influenced by the relative
| |
| amount of yolk in different cases. A large coeloblastula, such as is present in
| |
| Amphioxus, does not occur in the ascidians or amphibians. In the ascidians this
| |
| is due not merely, nor largely, to the amount of the yolk but rather to the shape of
| |
| the cells which are always elongated either at one pole or the other so as to nearly
| |
| fill the blastocoel ; the latter is small at all stages and the embryo and larva very
| |
| compact. In the amphibians the relatively small size of the blastocoel is due
| |
| not only to the quantity of yolk, but also to the many-layered character of the
| |
| blastula wall.
| |
| | |
| In all three classes the ectoderm arises from the upper hemisphere of the blastula, the endoderm and mesoderm from the lower hemisphere, but the precise relation of these germ layers to the third cleavage plane is not known in the cases of
| |
| Amphioxus and the Amphibia.
| |
| | |
| Most investigators affirm that the gastrula invagination in Amphioxus is at
| |
| first radially symmetrical, and only in the later stages does it become unsymmetrical. Samassa (1898), on the contrary, finds that the gastrula is bilateral from the
| |
| beginning and concludes that this bilaterality is the direct outcome of the bilaterality of the cleavage stages. In both ascidians and amphibians it is bilateral from
| |
| the first, the invagination appearing near the anterior border of the dorsal face and
| |
| then extending so as to include most of the dorsal area.
| |
| | |
| 5. Closure of Blastopore.
| |
| | |
| In ascidians the closure of the blastopore results largely from the progressive
| |
| posterior growth of the anterior (dorsal) lip, while the posterior (ventral) lip
| |
| remains relatively fixed in position. Owing to the peculiar differentiation of the
| |
| cells of the blastopore lip they can be individually followed through a large part of
| |
| this process; the number of cell rows between the posterior lip and the animal
| |
| pole and between the anterior lip and that pole can also be determined with accuracy during the earlier stages of the closure: from both of these facts it is certain that the posterior lip takes only a small part in the closure of the blastopore, except
| |
| in the final stages of that process. The posterior border of the blastopore is formed
| |
| of mesodermal cells derived from the crescent ; these cells are larger and more
| |
| rounded than the cells of the anterior border and are easily distinguishable by
| |
| their color and texture. In the closure of the blastopore they are rolled in at
| |
| the lateral margins but not at the hinder end. and owing to the large size of
| |
| these " myoblasts " the posterior portion of the blastopore is reduced to a longitudinal groove. Finally, this groove is closed by growth from all sides, the posterior lip growing more rapidly than the anterior one in the final stages of the
| |
| process.
| |
| | |
| | |
| | |
| Figs. XXVII XXXVIII. Schematic representations of three stages in the gastrulatiou of an ascidian, of Amphioxits
| |
| and of an amphibian to show their supposed resemblances in (1) Axial Relations, (2) Closure of Blastopore, (3l Origin of
| |
| Neural Plate, (4) of Chorda and (5) of Mesoderm. The position assigned to the polar bodies in Amphioxus and the amphibian is to a certain extent hypothetical. The head of the arrow marks approximately the anterior limit of the neural
| |
| plate; the tail of the arrow, the median mesenchyme cells in the posterior (ventral) lip of the blastopore. The mesodermal cells or areas are shaded by entire or broken lines.
| |
| | |
| FlQS, XXVII XXIX. Right halves of bisected gastrulae of Cynthia ; the chorda cells are shaded by coarse stipples,
| |
| the neural plate cells in. p.) by line stipples. The muscle cells (m.s.) lie lateral to the mesenchyme cells (m'cfa l and by the
| |
| overgrowth of the anterior (dorsal) lip of the blastopore are separated from the mesenchyme cells anteriorly (XXIX),
| |
| | |
| FlGS. XXX XXXII, Right halves of bisected gastrula- of Amphioxus (mainly after Hatschek I. In the two earlier
| |
| stages the existence and position of the mesoderm is hypothetical, being based upon the conditions found in ascidians.
| |
| | |
| Figs, XXX I II -XXX V. Right halves of bisected gastruke of the frog 'mainly after Kopsch I. The areas shaded by
| |
| stippled lines represent the supposed position of the mesoderm, here covered hv endoderm and yolk.
| |
| | |
| Figs. XXXVI XXXVIII. Dorsal views of late gastrnte of ascidian (XXXVI), Amphioxus (XXXVII), and frog
| |
| | |
| (XXXVIII), The circles marked 1, 2, 3. 4 indicate successive stages in the closure of the blastopore. The actual position
| |
| | |
| of the mesoderm in the ascidian and its supposed position in Amphioxus and the frog is shown by the radiating lines
| |
| | |
| around the blastopore. The unshaded area (Ch.) anterior to the blastopore and between the halves of the mesoderm rep
| |
| nts the plate of chorda cells.
| |
| | |
| | |
| | |
| | |
| | |
| The earliest trace of the anterior lip appears just posterior to the chorda
| |
| cells, the endodermal cells here becoming depressed (fig. 134); at this stage the
| |
| chorda cells and the neural plate cells which lie just anterior to them are at the
| |
| same level, but in the posterior growth of this lip the chorda cells are rolled in so
| |
| that they form the inner, as the neural plate cells form the outer, layer of the anterior (dorsal) lip. None of the neural plate cells and none of the ectodermal cells
| |
| are ever inrolled, the only cells which suffer this fate being the chorda cells and
| |
| the muscle cells (myoblasts).
| |
| | |
| There has been much controversy as to the part played by the anterior and
| |
| the posterior lips in the closure of the blastopore in Amphioxus and the amphibians. Kowalevsky supposed that the closure in Amphioxus occurred in a radially
| |
| symmetrical manner, the entire bolder of the blastopore growing equally ; Hatschek
| |
| thought that the growth of the anterior (dorsal) lip was the chief factor in the
| |
| closure; Lwoff, Klaatsch, Samassa, Morgan and Hazen agree in the main with
| |
| Kowalevsky. MacBride (1898) finds that in the final stages of closure the ventral
| |
| lip grows more rapidly than the dorsal.
| |
| | |
| Among the amphibians, observation and experiment show that the overgrowth of the dorsal lip is greater than that of the ventral, but the relative
| |
| amount of growth of each lip is not certain. In early stages of closure the dorsal
| |
| lip is alone concerned, as is also the ease with ascidians ; in later stages growth
| |
| takes place from all sides. According to Pfliiger the dorsal lip sometimes moves
| |
| through an are of 180 in the case of the frog, according to Roux 170; Morgan
| |
| estimates this movement at 120, Kopsch at 75. and H. V. Wilson at 722. In
| |
| the ascidians there is no doubt whatever that the closure is due chiefly to the
| |
| growth of the dorsal lip, though owing to the changing shape of the embryo it
| |
| is difficult to estimate the angular amount of that growth.
| |
| | |
| In Amphioxus and the ascidians the growth of the dorsal lip occurs as rapidly
| |
| in the middle as at the sides and there is therefore no indication of concrescence of lateral lips. At no stage during the closure of the blastopore in these
| |
| animals is there any indication whatever of such concrescence, either in the
| |
| form of a notch at the edge of the dorsal lip or of a seam along the middle of
| |
| the neural plate. In Cynthia and Ciona I have seen every division of the cells
| |
| of this lip up to an advanced stage and these divisions take place as rapidly
| |
| and uniformly along the mid-line as at the sides. Practically all investigators,
| |
| who have studied the embryology of Amphioxus or the aseidians are in agreement
| |
| upon this point, and if concrescence occurs among the amphibians, as is claimed
| |
| by some investigators, though denied by others, it can only be said that in this
| |
| respect the amphibians are very different from these other classes. The evidence
| |
| that the amphibians do form such an exception is by no means conclusive, as Ziegler
| |
| (1902) points out,
| |
| | |
| The question whether and to what extent there is an actual inrolling of cells
| |
| from the outer to the inner layer in the closure of the blastopore is one which has
| |
| been much discussed. In all three of these chordate classes an inrolling of cells at
| |
| the margin of the blastopore has been repeatedly observed, but the relative number,
| |
| the origin and the character of such cells are matters of dispute. Lwoff (1894)
| |
| maintains that the entire dorsal lip of Ampktoxus, inner as well as outer layer, is
| |
| formed from ectoderm cells which are inrolled. All of these inrolled cells he counts
| |
| as ectoderm and consequently concludes that the chorda and mesoderm are of ectodermal origin. The invagination of the endoderm is, in his opinion, the real gastrulation. whereas the turning in of the ectodermal cells is a coenogenetic process
| |
| which has nothing to do with the formation of the enteron but is concerned only
| |
| with the formation of chorda and mesoderm. This conclusion has been criticised
| |
| by Samassa (1898). Klaatsch (1896), Morgan and Hazen (1900). et al., on the
| |
| ground that there is no sufficient evidence that the inrolled cells are ectodermal.
| |
| With this conclusion, when extended to the aseidians. I heartily agree. Here
| |
| the cells which are inrolled at the anterior border of the blastopore are chorda
| |
| cells which are yolk laden and resemble endoderm and not ectoderm. The cells
| |
| which are inrolled at the posterior lateral borders are mesenchyme and muscle cells
| |
| and in histological structure are very unlike the ectoderm. While therefore agreeing with Lwoff that the chorda and mesoderm cells are inrolled (though from
| |
| opposite portions of the blastopore lip in aseidians) I agree with his critics that
| |
| these cells, judged by their lineage and histological character, are certainly not
| |
| ectodermal.
| |
| | |
| | |
| G. Neural Plate.
| |
| | |
| | |
| In aseidians the neural plate material becomes segregated into six cells at the
| |
| 44-cell stage ; four of these cells lie in a transverse row at the anterior border of
| |
| the dorsal hemisphere, just below the third cleavage plane and two of them lie
| |
| just above this plane and therefore in the ventral hemisphere. The four dorsal
| |
| cells lie just anterior to the four chorda cells from which they were separated at the
| |
| sixth cleavage. Both- the dorsal and ventral cells divide transversely, the former
| |
| sjivimz; rise to an arc of eight cells the latter to one of four cells, and to these a
| |
| single additional cell is added on each side making an arc of six neural plate cells
| |
| in the ventral hemisphere. In subsequent divisions the neural plate increases much
| |
| in length and its anterior portion also increases in breadth, but it is never more than
| |
| eight cells wide in its posterior part, Soon after gastrulation begins the neural
| |
| cells overgrow the chorda cells and thereafter cover the dorsal lip to its posterior margin. During all this time the anterior margin of the plate reaches only about
| |
| one-third of the way from the equator to the animal pole. The posterior margin of
| |
| the plate reaches nearly to the hinder cud of the embryo, and when the blastopore
| |
| doses a pair of V-shaped folds runs forward from the region of the blastopore
| |
| inclosing the neural plate between them. These neural folds then fuse from behind
| |
| forwards thus converting the plate into a tube. Dorsal to the notochord the neural
| |
| tube becomes solid ; in the region in front of the notochord it retains its lumen.
| |
| There is no nerve ring around the blastopore and probably none of the ectoderm cells around the posterior margin of the blastopore are added to the neural
| |
| plate.
| |
| | |
| In Amphioxus and amphibians the neural plate is first recognizable about the
| |
| the time of the closure of the blastopore. As in ascidians it arises in the outer layer
| |
| of the dorsal lip and extends back as far as the blastopore, but whether its cells
| |
| arise in close connection with the chorda and from both dorsal and ventral hemispheres as in the ascidians is unknown; furthermore, the distance of the anterior
| |
| edge of the plate from the animal pole is unknown. The work of Kopsch (1900)
| |
| indicates that in the frog the anterior margin of the plate is situated less than
| |
| half the distance from the equator to the animal pole, and H. V. Wilson (1000) in
| |
| particular has shown that the anterior part of the neural plate is formed from the
| |
| black hemisphere, the posterior part from the white hemisphere. a result which
| |
| agrees precisely with my observations on ascidians. As is well known the method
| |
| of closure of the neural tube in Amphioxus is peculiar, while the solid character
| |
| of the hinder part of the tube is peculiar to the ascidians, but with these exceptions
| |
| the later history of the neural plate and tube is essentially similar in all three
| |
| classes.
| |
| | |
| | |
| 7. Chorda.
| |
| | |
| In ascidians the substance of the chorda is segregated into a single transverse row of cells just posterior to the neural cells at the 44-cell stage, before
| |
| there is a trace of gastrulation. These chorda cells are generally clearer and
| |
| contain rather less yolk than the endoderm cells which lie immediately posterior to
| |
| them. These four chorda cells divide transversely forming an arc of eight cells and
| |
| soon thereafter a depression of the endoderm occurs posterior to this arc. which is
| |
| the beginning of the gastrulation. These chorda cells are flanked on each side by
| |
| the most anterior cells of the mesenchyme arc. the two arcs together forming the
| |
| chorda-mesenchyme ring of Castle. The eight chorda cells then divide anteroposteriorly forming two rows of eight cells each. This plate of cells by shoving,
| |
| by interdigitation and perhaps to a limited extent by folding, decreases in width and
| |
| increases in length, the cells finally, in a late larval stage, becoming arranged in a
| |
| single linear series. When they first arise the chorda cells are superficial in position, but in the overgrowth of the dorsal lip they are inrolled so as to lie in the
| |
| roof of the gastrocoel. The posterior growth of the dorsal lip carries the entire
| |
| chorda into the hinder half of the embryo, and it afterwards extends to the tip of
| |
| the developing tail.
| |
| | |
| | |
| In Amphioxus the earliest stage at which the chorda has been positively identified is one when the blastopore is small and the embryo elongated. According to
| |
| Hatschek it consists at this stage of a plate, about six cells wide, in the roof of the
| |
| archenteron and extending along the mid-line of the dorsal lip throughout its entire
| |
| length. This plate is narrower and longer than it is in the ascidians. but is otherwise much the same in appearance. The later history of the chorda is essentially
| |
| the same in both forms. With regard to the origin of the chorda cells in
| |
| Amphioxus, Morgan and Hazen (1900) have shown that the cells which are inrolled
| |
| in the formation of the dorsal lip and some of which must take part in the formaation of the chorda, are clear and contain less yolk than the endoderm cells.
| |
| Whether these cells form at this stage a plate which is wider from side to side
| |
| than it is long, as is true of the ascidian, is not known. Lwoff (1804) has also
| |
| recognized the fact that the chorda cells are rolled in at the margin of the dorsal lip,
| |
| and for that reason he regards them as of ectodermal origin.
| |
| | |
| In Amphioxus and in some amphibians the definitive roof of the enteron arises
| |
| from cells which lie along each side of the chorda plate, and which finally grow
| |
| under that structure and thus separate it from the gastric cavity ; in the ascidians the
| |
| chorda lies ultimately in the posterior part of the body where the gastric cavity is
| |
| almost entirely lacking and there is no growth of endoderm cells under it to form the
| |
| roof of the enteron. In most amphibians the chorda does not form a broad plate of
| |
| cells, but is a narrow rod closely united ventrally with the endoderm. which forms
| |
| the roof of the enteron, and connected laterally with the mesoderm. In these
| |
| three groups of chordates the chorda plate is widest in ascidians and narrowest
| |
| in amphibians. In all three it lies in the dorsal lip and is connected laterally
| |
| with mesoderm (text figs. XXXVI XXXVIII). The later history of the chorda
| |
| is essentially the same in all three classes.
| |
| | |
| The question whether the chorda is of endodermal or of mesodermal origin is.
| |
| as has been frequently said, one of definition of terms. Castle concludes that it is
| |
| mesodermal because in Amphioxus and lower vertebrates it " is derived from a
| |
| common fundament with what is universally regarded as mesoderm" and also
| |
| because it "comes to occupy a position between the inner and outer layers of the
| |
| embryo." On the other hand, the histological structure of the chorda cells in
| |
| Cynthia and Ciona is much more like endoderm than mesoderm, and they are
| |
| unquestionably derived from cells of the gastric endoderm at the 32-cell stage (fig.
| |
| 117, 103). I believe that special importance should attach to the structure of the
| |
| cells which form the chorda, and if this be accepted as a guide the chorda, at least
| |
| among ascidians. should be regarded as endodermal.
| |
| | |
| | |
| 8. Origin of Mesoderm.
| |
| | |
| The exact place and manner of origin of the mesoderm of ascidians can be recognized with the greatest certainty in the gastrula, cleavage stages and even in the unsegmented egg. The crescent, from which most if not all of the mesoderm arises, lies just below the equator of the unsegmented egg, and on the posterior side, its arms extending forward to the second cleavage plane. It occupies this
| |
| position throughout the whole of the cleavage, its substance becoming localized in
| |
| a number of large rounded cells. In the gastrulation these cells are inrolled along
| |
| the lateral-posterior borders of the blastopore, thus reducing the posterior portion
| |
| of the blastopore to a groove and rendering the whole blastopore pear-shaped. No
| |
| such appearance is found in Amphioxus or amphibians where the blastopore retains
| |
| its circular form until a late stage; this may be interpreted as due to the fact that
| |
| in these animals the mesoderm is not so largely developed at an early stage, but
| |
| it furnishes no satisfactory reason for supposing that the mesoderm is not formed
| |
| in corresponding positions in all three classes. We know that the neural plate and
| |
| the notochord come from similar regions in all three, and it is most unlikely that
| |
| the mesoderm arises froin wholly different regions.
| |
| | |
| Hatsehek's account of the origin of the mesoderm of Ampliioxus shows some
| |
| important resemblances to what occurs among ascidians. He found that running
| |
| back on each side from the first appearing primitive segments was a mesodermal
| |
| fold which led to a pair of pole cells in the ventral (posterior) lip of the blastopore.
| |
| All recent investigators nave denied the existence of these pole cells, and there
| |
| can be little doubt that Hatschek was mistaken with regard to them. Even in the
| |
| ascidian there are, strictly speaking, no pole cells in this region, nor anywhere
| |
| else in the embryo. The cells which in the ascidian occupy the position assigned
| |
| by Hatschek to the pole cells are the posterior mesenchyme cells. These cells form
| |
| the middle of the crescent, and from them a band of mesoderm cells runs forward
| |
| on each side, but these bands were not formed by the teloblastic growth of the posterior cells; on the contrary, their substance was localized in the crescent before
| |
| cleavage began. However the non-existence of the pole cells of Ampliioxus does
| |
| not destroy belief in Hatsehek's account of the mesodermal folds which run backward from the primitive segments to the blastopore. Several investigators have
| |
| recognized such folds or bands, and their existence can scarcely be doubted. These
| |
| bands have been seen only in older gastrulse, and they here occupy a position
| |
| which corresponds very closely with the mesenchyme bands in the ascidian gastrula. The separation of the muscle band from the mesenchyme band in the older
| |
| gastrulse of the ascidian [v. p. 69) is evidently a coenogenetic phenomenon, since
| |
| nothing of this sort is known to occur elsewhere. If the mesodermal bands of
| |
| Ampliioxus are present in earlier stages than those in which they have been represented by Hatschek, and if they occupy the same relative position as in the ascidian they would surround the posterior border of the blastopore, and only by overgrowth of the dorsal lip and the narrowing of the whole blastopore would they
| |
| come to lie alongside of the notochord. That mesodermal cells are present in the
| |
| posterior lip of the gastrula of Amphioxus at an early stage is made probable by
| |
| the observations of Lwoff, Klaatsch, Morgan and Hazen. Lwoff has found that
| |
| the longitudinal musculature of Amphioxus arises along the hinder lateral parts of
| |
| the blastopore, where it comes from ectodermal cells, as he thinks, which are inrolled. Klaatsch agrees with this and compares the "pole cell bands" of ascidians with these mesodermal folds of Amphioxus. He has observed that in both ascidians and Amphioxus these cells are more rounded than other cells of the gastrula.
| |
| Like Klaatsch, Morgan and Hazen find that around and within the ventral lip of
| |
| the blastopore, during the early gastrula stages, there are frequently found small
| |
| rounded cells which contain little yolk. They affirm that the form of these cells
| |
| is not the result of cell division, as Samassa had assumed, but that they preserve
| |
| their rounded form even in the resting stage.
| |
| | |
| Samassa ( 1S98), however, says that in Amphioxus the origin of the mesoderm
| |
| has no relation to the blastopore. The fact that the mesoderm has it growth
| |
| zone at the caudal end of the embryo, in the vicinity of the blastopore, is, he says,
| |
| a condition which it shares in common with all other organs of the embryo. In
| |
| the face of the positive evidence adduced by Lwoff, Klaatsch, Morgan and Hazen
| |
| this negative conclusion of Samassa's seem to me to lose much of its weight.
| |
| | |
| It seems probable from these accounts that mesoderm cells are present in
| |
| the ventral lip of the early gastrula of Amphioxus just as in the ascidians, and
| |
| that they give rise to the longitudinal mesodermal folds of later stages ; it remains
| |
| to be seen whether these mesoderm cells may not be traced back to a still earlier
| |
| stage, comparable with the crescent in the ascidian egg (cf. text (ins. XXVIIXXXII).
| |
| | |
| The origin of the mesoderm in amphibians is a much more difficult and complicated question and one into which I cannot enter full}' here. It is generally
| |
| believed, however, that in the frog's egg the cells which are to form the mesoderm
| |
| are present when the dorsal lip first appears, and even prior to that time. They
| |
| are the deeper layer of cells of the blastoporic ring and, therefore, surround the
| |
| egg below the equator. Whether at their first appearance they surround the entire
| |
| blastopore is not plain, but in later stages this is said to be the case. According to
| |
| this view the notochord is a mesodermal structm*e differentiated out of the continuous ring of mesoderm surrounding the blastopore. There is here resemblance
| |
| to the chorda-mesenchyme ring which is present in the ascidians and probably also
| |
| in Amphioxus, but in the amphibians this ring appears to give rise at once to a
| |
| sheet of mesoderm and not to mesodemal bands such as are found in Amphioxus
| |
| and ascidians (text figs. XXXIII-XXXVI1 1 ).
| |
| | |
| On the whole it is probable that there is fundamental agreement between
| |
| Amphioxus and ascidians in the place and manner of mesoderm formation, and
| |
| though the amphibians differ in some important respects from the other two classes
| |
| it is possible to interpret their method of mesoderm formation in the same general
| |
| terms.
| |
| | |
| Referring to Raid's (1892) " Theorie des Mesoderms," Samassa (1898), and
| |
| Garbowski (1898) maintain that there is no peristomal" mesoderm in Amphioxus,
| |
| but that all the mesoderm is "gastral." If the view here taken is correct, all the
| |
| mesoderm of this animal is at first peristomal while the gastral mesoderm is later
| |
| derived from this. This is exactly the conclusion which has been readied by Davidoff
| |
| (1891), and Castle (1896), with regard to the ascidian, a conclusion which I can fully confirm from my own work. Furthermore, it is not improbable that the same
| |
| thing is true of the amphibians. This is in confirmation of Rabl's view that the
| |
| peristomal mesoderm is palingenetic, the gastral coenogenetic, and the suggestion
| |
| is raised that in all these rases the gastral mesoderm is derived from the peristomal
| |
| through the manner of overgrowth of the dorsal lip of the blastopore (text figs.
| |
| XXXY1 XXXVIII).
| |
| | |
| Although 1 have made no special study of the subject, and cannot therefore
| |
| speak with assurance. I have seen no evidence in favor of Van Beneden and Julin's
| |
| view thai enterocoels are present in ascidians as in Amphioxus\ in this respect 1
| |
| am in accord with the more recent students of ascidian development (Davidoff,
| |
| Castle).
| |
| | |
| From these comparisons 1 think it may be safely concluded that there are
| |
| many fundamental resemblances in the early development of Tunicata, Amphioxus
| |
| and Amphibia, and that in consequence of the early differentiation of the ascidian
| |
| egg and embryo and because of the known cell-lineage of some of its important
| |
| organs the development of these animals throws light upon the embryology of other
| |
| chord ate classes.
| |
| | |
| | |
| VII. THE ORGANIZATION OF THE EGG.
| |
| | |
| It is interesting to observe how recent studies of development have led to the
| |
| recognition of morphogenetic differentiations at earlier and earlier stages in the
| |
| ontogeny; a dozen years ago the germ layers were the earliest differentiations of
| |
| this sort which were generally recognized. It was in the attempt to determine the
| |
| cellular origin of the germ havers that it became evident that the cleavage cells
| |
| themselves were of morphogenetic value. Some of the differentiations of the
| |
| cleavage cells could be traced back to the ver} r first cleavage or even to the
| |
| unsegmented egg ; thus the study of cell-lineage led logically and unavoidably to
| |
| the conclusion that the cleavage cells and even the unsegmented egg must be organized with reference to the parts and axes of the future animal.
| |
| | |
| For our present purposes the organization of the germ cells lias reference only to
| |
| such differentiations as are of direct value in the building of the embryo, in other
| |
| words, such as are morphogenetic, and it may be held to include phenomena of
| |
| polarity, symmetry and localization ; it obviously includes other things also, such as
| |
| regeneration and regulation, which are not. however, objects of investigation in this
| |
| work.
| |
| | |
| | |
| A. Polarity.
| |
| | |
| Fifty years ago Remak showed that the pigmented hemisphere of the frog's
| |
| egg gave rise to the cells of Von Baer's animal germ-layer," while the white
| |
| hemisphere gave rise to the 'vegetative germ-layer." The middle of the ectodermal hemisphere hasever since been known as the animal pole, the middle of the endodermal hemisphere as the vegetative (vegetal) pole. It is a remarkable fact that with a few possible exceptions, which arc by no means well established, the
| |
| polar bodies arc formed at the animal pole of the egg in all cases. This is a fact of
| |
| the most general occurrence and of the highest significance : it indicates that before
| |
| or during the maturation of the egg there occurs a polar differentiation or localization of the egg substance of such a kind that in all cases the future ectoderm is
| |
| formed at the maturation pole and the endoderm at the opposite pole.
| |
| | |
| | |
| | |
| 1 A more complete discussion of this suhjert, especially that portion of it which relates to experimental work, is reserved for a subsequent paper, only such matters being treated hen- as are the
| |
| outgrowth of the observations recorded in the preceding pages.
| |
| | |
| | |
| | |
|
| |
| | |
| The apparent exceptions to this rule are few in number and may be examined
| |
| in some detail ; they are limited to the eggs of certain insects, Petromyzon,
| |
| copepods, Ascaris, echinoderms and aseidians. The only reason for supposing, as
| |
| Korschelt and Heider (1903, pp. 545, 546) do, that the polar bodies are not formed
| |
| at the animal pole in insects and in Petromyzon is that they here lie to one side of
| |
| the pointed end of the egg; there is no proof that they do not lie at the middle
| |
| of the ectodermal area. Hacker (1899) says that in the larger species of Cyclops
| |
| "neither the place of formation of the polar bodies, the place of entrance of the
| |
| sperm nor the position of the first cleavage spindle are preformed in the egg, but
| |
| are secondarily determined by the position of the egg in the egg sack" (pp.
| |
| 193, 194). However, this egg is one which is not easy to orient, and it has by no
| |
| means been proven that the polar bodies do not form in this case at the middle of
| |
| the ectodermal area. Even if the justice of all of Hacker's statements bo admitted
| |
| it has not been shown that the cleavage spindle may not rotate so as to cause the
| |
| first and second cleavage furrows to pass through the maturation pole, as is usually
| |
| the case. Such a rotation of the first cleavage spindle takes place in nematodes,
| |
| and a somewhat similar rotation of the entire egg, after the formation of the first
| |
| cleavage spindle, has been described by Bigelow (1902) in the case of Lepas, where
| |
| it had previously been held that the first cleavage was equatorial. Hacker's observations do not show that the chief axis of the egg is not predetermined, and they
| |
| certainly do not prove that the maturation pole and the ectodermal pole do not
| |
| coincide.
| |
| | |
| In Ascaris megalocephala, Boveri (1887) observed that the second polar body
| |
| is usually formed at some distance from the first "whether through wandering in
| |
| the protoplasm or through a turning of the entire egg I could not determine" (p.
| |
| 32). His figures (1888, pi. IV) show that the first cleavage furrow frequently
| |
| passes through the point of attachment of the second polar body. The study of
| |
| the cell lineage of Ascaris has shown that most of the ectoderm is segregated in
| |
| one of the first two cleavage cells (the "primary ectoderm cell" of Zur Strassen,
| |
| 1896). This would seem to indicate that in this animal the polar bodies-do not lie
| |
| at tin' middle of the ectodermal pole; however the relations of the maturation
| |
| pole to the ectodermal pole and to the first cleavage arc not clear in this case, and
| |
| it may not be impossible that Ascaris may yet be found to conform to the general
| |
| | |
| rule.
| |
| | |
| As for the echinoderms. Wilson (1895) supposed from indirect evidence that
| |
| the maturation pole and the future animal pole did not usually coincide in
| |
| Toropneustes, and further that the chief axis of the egg was established only after fertilization. However, the evidence in favor of this is not conclusive as Wilson
| |
| admits. On the other hand. Boveri i L901 > has shown in the most convincing manner that in Strongylocentrotus the polarity of the egg may be traced hack to the
| |
| ovocyte, and that this polarity determines the gastrular axis. It is. therefore,
| |
| possible that in all echinoderms the polaritj of the egg is predetermined in the
| |
| ovary, and not after the maturation and fertilization, and that in all cases the
| |
| maturation and ectodermal poles coincide. 1
| |
| | |
| The most remarkable ami apparently well established of these exceptions to
| |
| the rule that the polar bodies are formed at the animal pole is that of the ascidians
| |
| studied by Castle (1894, 1896), where the polar bodies were said to he formed at
| |
| the vegetal or endodermal pole of the egg. However, this conclusion rests upon
| |
| erroneous orientation, as I have shown in the preceding pages ; in ascidians as in
| |
| other animals the polar bodies are formed at the ectodermal pole. There are, therefore, no well established exceptions to this general law. 2
| |
| | |
| In many cases it is known that the polar differentiation of the egg may he
| |
| recognized while the egg is still in the ovary. Reference has just been made to
| |
| the condition in Strongylocentrotus in which the pole of attachment to the ovarian
| |
| wall becomes the maturation pole of the egg and the ectodermal pole of the larva.
| |
| Boveri says that in all known cases the pole toward which the germinal vesicle is
| |
| eccentric becomes the animal pole. In Unto, Lillie (1900) has demonstrated that it is
| |
| the free pole of the egg which becomes the maturation and ectodermal pole, while the
| |
| pole of attachment becomes the vegetal pole. In a number of gasteropoda (Lzmn&a,
| |
| Succinea. Polygyra, Limax, Phvsa. Planorbis. Ancylus) I have found that there
| |
| is a marked polar differentiation of the vfi'j: in the ovary, the germinal vesicle being
| |
| eccentric toward the tree pole of the ovocyte. 1 have elsewhere (1903) shown
| |
| reason for believing that in dextral snails the polar bodies are formed at the free
| |
| pole and in sinistral snails at the attached pole of the ovocyte. In his work on
| |
| Cerebratiihis, Wilson ( 1 9 >> > , found that the polar bodies were formed at the live
| |
| pole of the ovocyte, and again in his recent paper on Dentalimn (1904), he finds
| |
| the side of attachment in the ovary represents the lower or vegetal hemisphere.
| |
| We find then that the chief axis of the egg is very generally present in the ovocyte,
| |
| and that the free side usually uives rise to the maturation and ectodermal pole,
| |
| while the attached side becomes the vegetal pole; but in echinoderms and probably
| |
| also in sinistral gasteropods these conditions are reversed, the side of attachment
| |
| becoming the ectodermal pole.
| |
| | |
| In the gasteropods named above, 1 found it possible to recognize this polarity
| |
| of the ovocyte at a very early stage ; in general it coincides with the "organic
| |
| axis" (Van Beneden), or the ''cell axis" (Heidenhain) i.e.. the axis passing through
| |
| the centrosome or sphere, and the center of the nucleus. This cell axis is a general characteristic of many, if not of all cells, and as it is present in all the cells of
| |
| the cleaving egg, where it is preserved from one cell generation to another (v.
| |
| Conklin, 1902), it may be considered to be a differentiation which is continuous
| |
| from generation to generation. But while the cell axis determines the egg axis
| |
| and this the gastrular axis, it is not necessary to suppose that in the early
| |
| ovarian history of the egg one pole is composed of ectodermal substance and the
| |
| other of endodermal. On the contrary, this is probably not the case. My observations on the living eggs of ascidians and snails leads to the view that it is not
| |
| the extrusion of the polar bodies at one pole which causes that pole to become
| |
| the ectodermal one, but rather that it is the movement of the germinal vesicle with
| |
| its contained clear protoplasm to o?ie pole, and the spreading of this pro/op/asm
| |
| at this pole, which is the determining factor. In short it is the localization of
| |
| ectodermal substance at the maturation pole which causes that pole to give rise
| |
| to ectoderm. I shall return to this subject in the section on localization.
| |
| | |
| | |
| | |
| 1 However, Garbowski (1904) affirms that in Asterias (jlaeialis the polarity of the egg is net
| |
| determined even in the 8-eelI and 16-cell stages, and thai the blastomeres are equipotential up v> the
| |
| 500-cell stag '
| |
| | |
| Wheeler 1897, p. 4 1 4t> has discussed in an admirable manner the apparent exceptions to
| |
| this law ni' polar differentiation and concludes that these exceptions are by no means well established.
| |
| | |
| | |
| | |
| Whether other axes of the egg are predetermined before cleavage is in most
| |
| instances unknown. In a lew cases all the axes of the future animal are marked
| |
| out before fertilization ; for example, among insects and cephalopods, as is well
| |
| known, it is possible to identify anterior and posterior, right and left, dorsal and
| |
| ventral axes of the egg while it is yet in the ovary. In most cases, however, the
| |
| only axis which is recognizable before fertilization is the chief axis of the egg.
| |
| This is true of the ascidians. but here there are certain evidences, which will be
| |
| presented in the next section, that the other axes are already established, though
| |
| not directly recognizable until after fertilization.
| |
| | |
| | |
| B. Symmetry.
| |
| | |
| Van Beneden and Neyt (1887) suggested that bilateral symmetry may be
| |
| characteristic of all cells of bilateral animals, and Lillie (1901) has expressed a
| |
| similar view regarding the eggs of such animals. This hypothesis, if true, would
| |
| materially simplify the problem of the earliest differentiations and localizations of
| |
| the egg, but it is supported by little direct evidence ; in fact, it is surprising that in
| |
| most bilateral animals bilaterality appears so late in development, In most annelids
| |
| and mollusks the egg and early cleavage stages are to all appearances radially symmetrical, and in many cases bilateral symmetry first appears with the formation of
| |
| the mesentoblast cell, 4d. In echinoderms bilaterality is said to appear first in the
| |
| gastrula stage ; in Amphioxus during cleavage ; in ascidians it appears immediately
| |
| after fertilization and before the first cleavage ; while in cephalopod and insect
| |
| eggs it appears during the growth of the ovocyte in the ovary. Wilson has
| |
| repeatedly expressed the view that characteristic differentiations, such as bilateral symmetry, arise at different periods of development in different cases, and it
| |
| cannot be denied that the ocular evidence is in favor of this view. On the other
| |
| hand, there are certain considerations which lead to the conclusion that bilateral
| |
| organization may be present in the developing egg or embryo long before it is directly
| |
| visible. For example, in Neritina there are two groups of granules in the protoplasm of the unsegmented egg, one on each side of the polar bodies. Blochmann
| |
| (1882) observed that these granules were ultimately localized during cleavage
| |
| in the right and left " Urvelarzellen." They therefore mark out a bilateral organization of the unsegmented egg, although the cleavage up to the time of the formation of the Urvelarzellen" is typically spiral and radially symmetrical. In other
| |
| gasteropod eggs, where these granules are lacking, not a trace of bilateral organization is visible before the formation of the mesentoblast cell ; yet it can scarcely be
| |
| supposed that the eggs of these gasteropods are so unlike those of Neritina as to be
| |
| actually radially symmetrical as they appear to be. Rather it seems probable that
| |
| the bilateral organization which appears in this one respect in the Neritina egg is
| |
| characteristic of other gasteropod eggs also, though it does not usually become
| |
| apparent until a later stage.
| |
| | |
| Crampton (1894) discovered that the cleavage of the egg in sinistral snails is
| |
| reversed as compared with that of dextral forms. I have shown elsewhere (1903)
| |
| that the inverse symmetry of sinistral snails is traceable to the inverse organization
| |
| of the unsegmented 'egg. Of this fact there can be no doubt, though it is not yet
| |
| certain how this inverse organization may have been produced. But an inverse
| |
| organization of the egg, such as would produce inverse symmetry of the embryo
| |
| and adult, implies of necessity a bilateral organization to begin with ; it must be,
| |
| therefore, that the eggs of these gasteropods are bilateral, though this fact is not
| |
| directly evident.
| |
| | |
| In the aseidian egg the first appearance of bilaterality which I have been able
| |
| to detect occurs soon after fertilization when the sperm nucleus moves toward one
| |
| side of the egg which later becomes the posterior pole. One might, therefore, lie
| |
| inclined to consider that in this case the egg before fertilization was radially symmetrical, and that the chance movement of the sperm into one meridian determined
| |
| the median plane of the embryo, were it not for the fact that all the movements of
| |
| the sperm within the egg seem to be directed by the organization of the cytoplasm.
| |
| The sperm always enters the egg near the vegetal pole, but the fact that the point
| |
| of entrance is nearer that pole in some instances than in others shows that that
| |
| point is not a fixed and constant one. After the sperm has penetrated the peripheral layer of protoplasm, and has turned so that its centrosome is directed forward in its movement through the egg it moves up to the equator of the egg in a
| |
| path nearly parallel with the surface. Arrived at the equator, the upward movement ceases and the sperm nucleus and centrosome. after meeting the egg nucleus,
| |
| turn in toward the center of the egg. These movements are of such a constant character that they cannot be the result of chance; they must be directed
| |
| and probably by the cytoplasm of the egg. Furthermore, it seems probable from the
| |
| evidence of such cases as figures 81 and 85 that the sperm nucleus does not
| |
| always take the shortest path to the equator as it should do if the egg were
| |
| radially symmetrical and the median plane were really determined by the path
| |
| <>f the spermatozoon . On the other hand, it sometimes appare)itly takes the longest
| |
| path as if it must needs move in a certain meridian. This seems to indicate that the median plane of tlie embryo is not determined by tlie chance path of the
| |
| spermatozoon within the egg, but rather that both the median plane and the
| |
| path of the spermatozoon are determined by the structure of the cytoplasm.
| |
| | |
| Finally, in cases of normal or artificial parthenogenesis the median plane
| |
| cannot be determined by the path of the spermatozoon. In eggs of this kind
| |
| the establishment of bilateral symmetry must be held to be due to the structure
| |
| of the egg itself or to environment, and whichever of these views may be accepted
| |
| it follows that the path of the spermatozoon cannot be regarded as a general factor
| |
| in determining the median plane of the embryo.
| |
| | |
| These and other similar considerations lead to the view that bilateral organization is frequently present in the egg before it becomes visibly manifest, and
| |
| they lend support to the hypothesis of Driesch (189G) that the eggs of all bilateral
| |
| animals are bilaterally organized, there being a "polar bilateral direction of particles" in the "intimate structure of the egg." // this be true, the eggs, the cleavage stages and the blastulce of annelids and mollusks, of echinoderms and Amphioxus are as truly bilateral as they are in the ascidians, thohgh this bilaterality
| |
| may be masked by a radial form of cleavage and by an apparently radial organization of the egg.
| |
| | |
| I cannot pass over this subject without referring to the extensive work of
| |
| Roux (1883, 1885, 1887, 1902, 1903) on the determination of the median plane in
| |
| the frog's egg. This work is too widely known to require more than passing
| |
| notice. By means of " localized fertilization," i. e., the application of spermatozoa
| |
| to any meridian of the egg, Roux has determined that the first cleavage plane
| |
| passes through the entrance point of the spermatozoon and that the median plane
| |
| of the embryo usually coincides with the first cleavage plane. He therefore considers that the median plane is in typical conditions, determined by the path of the
| |
| spermatozoon. Moskowski (1902), on the other hand, holds that the first cleavage
| |
| plane and the median plane of the embryo are determined by definite movements
| |
| of the egg substance and not by the path of the spermatozoon. Castle (1896)
| |
| believed that the plane of the first cleavage and the median plane of the embryo
| |
| were determined, in the ascidians studied by him, by the place of entrance of the
| |
| spermatozoon, the point of entrance marking the posterior pole ; I mt since the point
| |
| of entrance is near the vegetal pole, while the posterior pole lies near the equator,
| |
| it is evident that the point of entrance cannot mark that pole. It is true that the
| |
| protoplasm which gathers around the head of the sperm as soon as it enters the egg
| |
| moves with the sperm to the posterior pole and there remains permanently, but the
| |
| location of this protoplasm at this pole is evidently due to something other than the
| |
| point of entrance of the spermatozoon. There is no question whatever that, in the
| |
| ascidians, the path of the sperm within tlie egg coincides with the plane of the
| |
| first cleavage and with the median plane of the embryo, but there is evidence, as
| |
| I have shown, that this path is itself determined by the structure of the e^'i.
| |
| | |
| | |
| | |
| C. Cytoplasmic Localization.
| |
| | |
| | |
| 1 . Localization in Cleavage Stages.
| |
| | |
| That there is a specification and localization of those portions of the protoplasm of the egg which are destined in development to give rise to definite organs
| |
| has been repeatedly affirmed and denied since His first propounded the doctrine of
| |
| "organ forming germ regions" in 1 874. At first this doctrine .took the form of a
| |
| mental projection of the early embryonic organs back upon the unsegmented egg.
| |
| Later the study of cell-lineage showed that definite organs of the larva or adult
| |
| ai'ose from definite blastomeres, which in turn came from definite portions of the
| |
| unsegmented egg. But although it was thus possible to map out the cleavage cells
| |
| ami the unsegmented egg into regions corresponding to certain organs of the embryo.
| |
| it was not usually possible to show that these regions were visibly different from one
| |
| .another. Nevertheless the fact that certain blastomeres constantly gave rise to
| |
| certain parts, and that other blastomeres developed very differently and gave riseto other parts, led students of cell-lineage generally to the view T that there must be
| |
| some protoplasmic difference between such blastomeres. though it might not be
| |
| directly visible.
| |
| | |
| On the other hand were those who maintained that the protoplasm of the early
| |
| cleavage stages was undifferentiated and that specifications which determined the
| |
| fate of these cells arose only at a later period and under the influence of environmental or extrinsic conditions, such as mutual interaction between the cells, position
| |
| in the developing embryo, etc. Such views were maintained on the ground of
| |
| experimental work, especially that of Driesch, Hertwig, Morgan. Wilson and others,
| |
| but it should not be forgotten that the experimental work of Roux furnished
| |
| important evidence in favor of the independent differentiation, " Selbstdifferenzirni/o-" . of different blastomeres.
| |
| | |
| Thus while the study of cell-lineaue showed conclusively that certain cells
| |
| were destined in the course of normal development to give rise to certain organs
| |
| and that the individual blastomeres were more or less differentiated from one another,
| |
| the results of experimental work showed that in many animals individual cleavage
| |
| cells were capable of giving rise to an entire embryo, and it was. therefore, affirmed
| |
| by some investigators that these cells could not lie differentiated for any particular
| |
| end. Inasmuch as these facts of cell-lineage and of experimental embryology were
| |
| well established, it was only possible to harmonize these discordant results by some
| |
| form of interpretation. This was undertaken from two different standpoints: (1)
| |
| It was affirmed that the early cleavage cells were not really differentiated lor any
| |
| specific end and that each might develop into any part of the embryo; if in any
| |
| ea-e certain parts or organs came from certain blastomeres it was due merely to the
| |
| '*' continuity of development" (Hertwig. ().. 1.892).
| |
| | |
| (2) On the other hand, it was suggested that these discordant results as to
| |
| the differentiation of tin/ early cleavage cells might be explained by the fact that
| |
| the eggs of different animals might differ in the time at which differentiations
| |
| arise. In the eggs of echinoderms. Amphioxits.. fishes and frogs, which had been chiefly employed in experimental work, the cleavage was not known to be constant
| |
| and differential in character; whereas in all forms the cell-lineage of which was
| |
| known, the cleavage was both constant and differential. I therefore suggested
| |
| i ! "- '. ' 7 1 that for the present it would be advisable to recognize two types of cleavage,
| |
| a determinate type in which the blastomeres are differentiated from one another
| |
| and are constant in their manner of origin and development, and an indeterminate
| |
| type.in which such differentiation and constancy are not known to occur. At the
| |
| same time I was careful to state that this indeterminateness might be only apparent
| |
| and not real, and "that the denial of a definite prospective value to each blastomere might rest upon the curious basis that no one had followed a single blastomere through the development" (1897, p. 191). In favor of such a distinction was
| |
| the experimental work which had been done on the eggs of ctenophores and gasteropods ; the cleavage in these animals is known to be determinate, and it was found
| |
| that from a part of an egg only a part of an embryo would develop. In all cases
| |
| constant and differential features appear sooner or later in the course of development, but if in some cases they appear late in the cleavage while in others they
| |
| appear early this would explain the fact that in some species a whole embryo may
| |
| he produced from one of the first two or first four blastomeres, whereas in other
| |
| cases only a partial embryo results. Wilson in particular has defended the view
| |
| that specifications arise at different times in different eggs, and that these differences in the time of specification may explain the different potencies of blastomeres
| |
| or portions of the egg.
| |
| | |
| While it is entirely possible that differentiations may appear in some cases
| |
| earlier than in others, experiments on the development of parts of eggs are no
| |
| satisfactory test of the presence or absence of such differentiations as the eggs of
| |
| echinoderms and ascidians well show. The echinoderms were supposed to present one of the best examples of an indeterminate form of cleavage; fragments of
| |
| the egg or isolated blastomeres here give rise to entire embryos, and it was concluded that differentiations must appear in these eggs relatively late in development.
| |
| But Boveri (1901) has shown that in Strongyloceiitrotus, and presumably in other
| |
| echinoderms also, 1 a remarkable stratification of the egg. corresponding to the primary organs of the larva, appears at the time of the maturation of the egg. These
| |
| observations have taught us more with regard to the actual differentiations of this
| |
| egg. as contrasted with the potencies of its parts, than all the experiments which
| |
| have ever been made. Again, the ascidian egg has one of the most determinate
| |
| and morphogenetic forms of cleavage known and the differentiations of the various
| |
| parts of the unsegmented egg are very great, and yet the experiments of Driesch
| |
| i L895, 1903) and Crampton (1897) have shown that entire embryos may be produced from isolated blastomeres of this egg ; such experiments apparently demonstrate the totipotence of the first four blastomeres of the ascidian egg, 2 but all the
| |
| | |
| See foot-note, p. 89.
| |
| | |
| ! Since this paper was written I have carefully studied the potency of individual blastomeres of
| |
| the ascidian egg by the experimental method. My result, which will be published elsewhere, show
| |
| that nothing resembling a normal embryo or larva is ever produced from any fragment of an egg which experiments in the world could not have shown as satisfactorily as direct observation
| |
| has done the remarkable cytoplasmic differentiations and localizations of this egg.
| |
| | |
| | |
| It seems, therefore, that this apparent conflict between the results of observation and of experiment on the early development of the egg, between the prospective tendency and the prospective potency of its various parts, can be harmonized
| |
| neither by the claim that differentiations do not exist in the early stages of development nor by the assumption that differentiations appear earlier in some eases
| |
| than in others.
| |
| | |
| (3) It seems rather that the true explanation of this discrepancy is the one
| |
| originally suggested by Roux (1892, 1895), viz., that there is a difference in the
| |
| regenerative or regulative capacity of different ova and that in the experimental
| |
| studies referred to we are dealing with indirect development or regeneration, as contrasted with direct or normal development . Just as some adult forms show little
| |
| capacity tor regeneration or regulation while others of equally complex differentiation show this power in a high degree, so it seems that the capacity for regulation
| |
| shown by eggs is more or less independent of the degree of their differentiation.
| |
| To all appearances the ascidian egg is more highly differentiated than those of mollusks or ctenophores, and yet the former has a much higher regulative capacity
| |
| than the latter. If this view of the relative independence of differentiation and
| |
| regulation be correct the conflict between the results of cell-lineage and of experimental e/ubryology disappears, for the prospective tendency or the actual differentiation of a blastomere and its prospective potency deal with two distinct things.
| |
| | |
| | |
| 2. Localisation before Cleavage.
| |
| | |
| The phenomena of germinal localization have heretofore been studied for the
| |
| most part during the cleavage and subsequent periods of development ; only within
| |
| the last few years has this study been extended to the egg before cleavage. Nevertheless the brilliant researches of Driesch, Lillie, Boveri, Fischel, Wilson and Carazzi
| |
| in this field have already yielded most important results, and are full of promise for
| |
| future work. In some cases this localization of different kinds of protoplasm or of
| |
| organ-forming substances has been directly observed, in other cases it has been
| |
| inferred from the results of experiment, but in many instances both observation and
| |
| experiment lead to the conclusion that the morphogenetic processes begin before
| |
| cleavage. The work of Lillie on Unio (1901) and Chestopterus (1902), and especially
| |
| experiments of Fischel (1897, L898, 1903) on the ctenophore egg, and of Wilson
| |
| (1903), and Yatsu (1904) on the nemertine egg have shown that definite regions
| |
| of the unsegmented egg give rise to definite organs or regions of the embryo.
| |
| | |
| Apart from the early separation of protoplasm and }*olk which occurs in many
| |
| yolk-laden eggs, localization of visibly different kinds of protoplasm in the unsegmented egg has been observed in relatively few cases.
| |
| Among the earliest observations of this sort are those of Robin (1875) and Whitman (1878) on the eggs of
| |
| leeches. Here peculiar aggregations of protoplasm occur at the two poles of the egg,
| |
| after maturation and fertilization, which have been called " polar rings." Vejdovsky
| |
| (1888) discovered that these polar rings arise in Rhynchelmis from a peripheral
| |
| layer of brown protoplasm, which has a great affinity for stains. The substance of
| |
| this layer collects at the two poles of the egg after maturation and fertilization and
| |
| thus constitutes the polar rings. During the cleavage most of this substance is segregated into the large posterior macromere of the 4-cell stage, and it ultimately
| |
| passes into the mesomeres (probably the first and second somatoblasts of Wilson).
| |
| Nevertheless other portions of this protoplasm go into the micromeres; in fact it
| |
| forms " the general material for the building of the body, with the exception of the
| |
| intestinal epithelium" (Vejdovsky. 1888, p. 123). Polar rings have also been
| |
| observed by Foot (1S04. 1896) in AUolobophora, and their method of formation in
| |
| this form has been determined in a most careful and satisfactory manner ; this work
| |
| will be discussed more fully in the next section on the genesis of egg organization.
| |
| | |
| | |
| | |
| does not include the whole of the right or lilt half. Individual blastomeres produce rounded masses cf
| |
| cells but have <> power '" give rise to muscle, chorda, neural plate or senst organs, if they do nut contain
| |
| thost portions of the egg which normally givt rise to these parts.
| |
| | |
| | |
| | |
| | |
| One of the most remarkable cases on record of the localization of visibly different kinds of ooplasm is found in Myzostoma glabrum in which Driesch (1S!J0). and
| |
| more recently Carazzi (1004), observed two conspicuous zones of protoplasm in the
| |
| egg before maturation, an upper one which is of a redish tint and a lower one which
| |
| is green. During the maturation of the egg the upper zone differentiates into
| |
| two. an upper red zone and an equatorial colorless one. According to Driesch
| |
| (1896. p. 120) the red zone gives rise principally to the substance of the micromeres (ectoderm), the clear zone to endoderm. and the green one to the substance
| |
| of the somatoblasts (ectoderm and mesoderm).
| |
| | |
| Another case of visible localization of the substances of the unsegmented egg
| |
| was observed by Boveri ( L901) in the ovocyte, egg and larva of Strongylocentroius
| |
| lividus ; here before maturation and fertilization the surface of the egg is covered
| |
| by a uniformly distributed red pigment; after maturation this gathers into an equatorial zone leaving an area of clear protoplasm at the upper pole and another at
| |
| the lower one. Later development shows that the upper clear cap gives rise to the
| |
| ectoderm, the red zone to endoderm and the lower cap to mesenchyme.
| |
| | |
| A visible localization of differently colored substances in the unsegmented eggalso occurs in fresh water snails belonging to the genera Physa, Planorbis and
| |
| Limncza. In these animals I have found (Conklin. 1903) that a clear cap of protoplasm appears at the upper pole during maturation and then gradually spreads,
| |
| over the upper hemisphere of the egg; the upper hemisphere thus becomes milkywhite in the living; egg, while the lower half remains yellow. 1 have followed these
| |
| | |
| O 00- 7 J
| |
| | |
| white and yellow substances through the development and find that the white substance gives rise to the ectoderm, the yellow to the mesoderm and endoderm.
| |
| | |
| Quite recently Wilson ( L904) has observed in Dentalium a localization of unlike
| |
| substances in the unsegmented egg and by a series of experiments he has shown
| |
| the part which some of these substances take in the formation of certain organs of
| |
| the larva. As in the case of Strongylocentroius there is here an accumulation of clear protoplasm at each pole of the egg with a broad pigment band around the
| |
| equatorial region. The clear polar areas, the lower of which forms a prominent
| |
| lobe, Wilson regards as comparable with the "polar rings" of leeches and oligochaetes. 1 In the course of development the upper white area is allotted to the three
| |
| quartets of ectomeres; the middle pigmented zone is mainly allotted to the four
| |
| basal entomeres, while the lower zone passes mainly into the first somatoblast (2d),
| |
| and possibly also into the second somatoblast (4d) and the left posterior micromere
| |
| (3d). This work is the most complete and important which has yet been done on
| |
| the subject of cytoplasmic localization and it firmly establishes the fact that different substances and areas of the unsegmented egg are causally related to different
| |
| organs and parts of the larva.
| |
| | |
| It is doubtful whether any other case of cytoplasmic localization hitherto
| |
| reported is more remarkable than that which has been described in the preceding
| |
| pages for the ascidian egg. The most striking features of this localization are the
| |
| great differences in the substances localized, the manner in which this localization
| |
| is accomplished and its bilateral character.
| |
| | |
| (1) The first of these features is the result of the different pigments which
| |
| are assoeiated with the different kinds of protoplasm, and which mark out as on a
| |
| map the various germinal areas of the egg. In Cynthia the pigment in the peripheral layer of protoplasm is yellow, the yolk is a blueish gray, while the protoplasm
| |
| which escapes from the germinal vesicle is colorless. Not the pigment but the protoplasm with which it is associated is of differential value, for the pigment may
| |
| differ most remarkably in different genera of ascidians. but the organs which arise
| |
| from similar areas are in all eases similar. What has been said of the pigment may
| |
| also be said of the yolk; this inert substance is not in itself of differential value,
| |
| but it lies in a definite region of the egg and probably in a particular kind of protoplasm, which it marks out as the yellow pigment does the peripheral layer.
| |
| | |
| Of these three kinds of protoplasm the yellow (mesoplasm) goes almost entirely
| |
| into the muscle and mesenchyme cells, though a small portion of it may be found
| |
| around the nuclei of other cells, the clear protoplasm (ectoplasm) is chiefly distributed to the ectoderm and the gray yolk-laden protoplasm (endoplasm) 2 to the endoderm, though here also some of these substances are distributed to all the cells. It
| |
| is not to be supposed that these three kinds of protoplasm are the only ones present
| |
| in the Q^iX. rather it is probable that others are present which are not visibly distinguishable. In fact, soon after the cleavage begins, it is noticeable that the protoplasm in the dorsal part of the crescent is a fainter yellow than that in the ventral
| |
| part, while from the time of the fertilization onward the middle of the crescent is
| |
| marked by a small area of clear protoplasm {v. p. 21) ; the deeply pigmented portion of the crescent gives rise to the muscle cells, the lighter or clearer portions
| |
| to mesenchyme. Inasmuch as the protoplasm which enters into the muscle cells
| |
| and mesenchyme is localized with such definiteness in the unsegmented egg it can
| |
| scarcely be supposed that the substances which are to give rise to the neural plate
| |
| and notochord are not also definitely localized though they may not be directly visible. 1
| |
| If this presumption is correct the visibly different organ-forming substances are
| |
| by no means the only ones present.
| |
| | |
| | |
| | |
| Several years ago I suggested (Conklin 1897, p. 39) that the yolk lobe (" polar lobe," Wilson
| |
| was comparable to the polar rings of leeches.
| |
| | |
| - It should lie observed that these names are given with reference to the part which these
| |
| different portions of the ooplasm play in the development of the animal; the peripheral layer of the
| |
| ovocyte, which would be called ectophtzm if the ovocyte alone were under consideration, is mesoplasm
| |
| when regarded from the standpoint of its fate in development.
| |
| | |
| | |
| | |
| | |
| (2) The striking effect of this cytoplasmic differentiation is heightened by the
| |
| manner in which localization takes place. The downrush of the peripheral la}'er
| |
| of yellow protoplasm to meet the entering sperm, the subsequent movement of this
| |
| protoplasm together with the sperm nucleus to the posterior pole and the formation
| |
| there of the crescent, the migration of the clear protoplasm to the lower pole, thence
| |
| to the posterior pole and then to the center of the egg. these phenomena are so
| |
| evident and they occur so rapidly that they strike the observer with amazement.
| |
| | |
| (3) Finally the bilateral character of this localization is most notable. In all
| |
| other recorded cases of cytoplasmic localization the various substances become
| |
| arranged in zones around the chief axis of the egg and the symmetry is apparently
| |
| radial; here the early stages of localization are also of this sort, and the gray upper
| |
| pole, the clear middle zone and the yellow lower pole of the Cynthia egg immediately after fertilization are not unlike the localizations in the eggs of Myzosloma or
| |
| Strongylocent rotus, but in the ascidian this apparent radial symmetry gives place
| |
| almost immediately to a marked bilateral symmetry which is brought about by the
| |
| movement of the protoplasm from the lower hemisphere to the posterior pole and
| |
| the formation there of the crescent.
| |
| | |
| Certain fundamental resemblances which run through all these cases of cytoplasmic localization are so striking that the}- scarcely need any emphasis hei'e. The
| |
| existence in the unsegmented egg of a peripheral layer of protoplasm which is
| |
| clearly distinguishable from the remainder of the egg is a phenomenon of very wide
| |
| occurrence. In most of the cases just named this peripheral layer aggregates at
| |
| one or both poles of the egg after fertilization, and in animals belonging to phyla
| |
| as far apart as annelids, echinoderms. mollusks and chorda tes the substances at the
| |
| upper pole give rise to ectoderm, those at the lower pole to mesoderm, while the
| |
| endoderm arises frotn the region intermediate between these two. Although many
| |
| differences appear in the later development of these animals they do not detract
| |
| from the value of these fundamental resemblances which apparently afford a sound
| |
| basis for a comparative morphology of ova.
| |
| | |
| 1 Since this was written I have heen able to distinguish the chora neural-plate substance as early
| |
| as the 2-cell stage ; it is the light gray protoplasm at the anterior border of the dorsal hemisphere
| |
| (figs. 28, 32 et seq.) Photomicrographs of living egg of this stage will be published soon in which
| |
| this substance is clearly shown.
| |
| | |
| | |
| | |
| D. Genesis of the Ohganization of the Egg.
| |
| | |
| It is probable that the differentiations of egg cells, of blastomeres, and possibly
| |
| t)f all types of cells, are reducible to two fundamental processes: (1) the genesis of
| |
| unlike substances, and (2) the localization of these substances in definite parts.
| |
| Few observations or experiments have been made on the former of these processes
| |
| and probably no other problem of development would better repay a thorough
| |
| investigation; the localization problem has been approached from many sides and
| |
| has yielded results of great interest and importance.
| |
| | |
| It is a significant fact that localization in the unsegmented egg takes place in
| |
| so many cases at the time of maturation and fertilization. This is the case in certain ascidians, fresh-water snails, nemerteans and echinoderms ; in Myzostoma and
| |
| Dentalium the two poles of the egg are dissimilar while the egg is still in the ovary,
| |
| but here also active localization goes on during maturation. In ascidians and
| |
| fresh-water snails it is not possible to determine whether the movements which
| |
| lead to localization are dependent upon the maturation or upon the fertilization of
| |
| the egg. since as yet it has not been possible to separate experimentally these
| |
| processes; they certainly seem to be associated with the entrance of the spermatozoon, but since the maturation does not here occur until after the fertilization,
| |
| it is not possible to determine with certainty the relative importance of these two
| |
| processes in causing localization. In Strongylocentrotus the movements which
| |
| lead to the formation of the red pigment zone occur after the extrusion of both
| |
| polar bodies and before fertilization ; in this case therefore the localization is
| |
| associated with the maturation.
| |
| | |
| 1 . Role of the Nucleus in Differentiation ; Cytoplasmic Organisation and the Nuclear Inheritance Theory.
| |
| | |
| The localization which is effected in the ascidian egg upon the entrance of the
| |
| spermatozoon is by no means the initial localization in this egg. In the ovocyte
| |
| 1) 'tore maturation and fertilization the mesoplasm, which later give rise to the mesoderm, exists as a peripheral layer of protoplasm, the ectoplasm, which in later
| |
| stages is chiefly distributed to the ectoderm, is in large part contained within the
| |
| germinal vesicle, while the yolk-laden portion of the egg, the endoplasm, which
| |
| later passes largely into the endoderm, is nearly central in position (figs. 61, 76).
| |
| At an earlier stage neither the peripheral layer nor the yolk are recognizable as
| |
| such ; the cell body is composed of granular deeply-staining protoplasm, and around
| |
| the nucleus is a distinct granular mass, the "yolk matrix" of Crampton (1899).
| |
| In the very young ovocyte this granular mass is situated chiefly on one side of the
| |
| nucleus, and frequently contains at its center a large granule, surrounded by a clear
| |
| aria, which I take to be the centrosome ; the granular mass surrounding this is
| |
| accordingly sphere material or archoplasm.
| |
| | |
| In the growth of the ovocyte the sphere material enlarges and spreads around
| |
| the nucleus, forming the yolk nucleus or matrix; it then begins to disintegrate into granules or larger masses. 1 as described by Crampton, which wander out into the
| |
| cell body. Crampton has observed that these granules give rise to the yolk spherules which first appear in the protoplasmic ground substance around the nucleus,
| |
| leaving the peripheral layer of the egg free from yolk.
| |
| | |
| I am of the opinion that the peripheral layer also contains portions of the
| |
| archoplasm or sphere material; the staining reactions of this layer are like those
| |
| of the archoplasm; in the disintegration of the sphere flocculcnt masses of archoplasm pass into this layer; finally, comparison with other forms favors this view.
| |
| The careful observations of Foot ( 1 S < > ) on the yolk nucleus and polar rings
| |
| of Allohbophora show that in this animal the polar rings may he traced back
| |
| step by step to a substance in the vicinity of the nucleus of the very young ovocyte, which Foot identifies with archoplasm. In the later stages this substance
| |
| becomes distributed throughout the cell and forms a more or less irregular peripheral layer; finally the substance of this layer aggregates at the two poles of the
| |
| egg to form the polar rings, as previously described.
| |
| | |
| Among gasteropods the sphere material is largely of nuclear origin, containing
| |
| nuclear sap and dissolved oxj'chromatin, which have escaped from the nucleus
| |
| during the period of mitosis (Conklin, 1902) ; if the same be true of the ascidians
| |
| both the peripheral layer of protoplasm (mesoplasm) and the yolk (endoplasm)
| |
| contain elements which were ultimately derived from the nucleus at the last ovogonic division.
| |
| | |
| The clear protoplasm (ectoplasm) which is apparent in the egg after maturation, and which, in the course of development, passes mainly into the ectoderm is
| |
| largely contained within the nucleus of the ovocyte. In the first maturation division an extremely large quantity of nuclear sap, containing an unusual amount of
| |
| dissolved oxychromatin, escapes into the cell body where it can be recognized as an
| |
| area of clear protoplasm. This clear protoplasm can be followed through a large
| |
| part of the development, both in ascidians and in gasteropods. In the latter particularly this clear nuclear plasm is plainly visible in the living egg. It forms a
| |
| fusiform or columnar area around the first maturation spindle, and after the formation
| |
| of the polar bodies it flattens out at the surface of the egg, forming first a cone,
| |
| then a lenticular mass, and finally a cap of clear protoplasm. This cap extends
| |
| down over the egg to a region a little below the equator, and finally during cleavage
| |
| it is largely localized in the three quartets of ectomeres.
| |
| | |
| In the ascidians the later history of this nuclear plasm is not so easily followed
| |
| as in the gasteropods, owing to the presence of a peripheral layer of mesoplasm.
| |
| and to the fact that its movements here are more extensive and complicated.
| |
| In Cynthia it flows to the lower pole along with the yellow mesoplasm. then it
| |
| moves with the sperm nucleus to the posterior side of the egg and finally to its
| |
| center. Here it surrounds the cleavage spindle, and at the close of the fust
| |
| cleavage moves toward the animal pole so that the larger part of it comes to
| |
| lie in the upper hemisphere. In subsequent divisions it surrounds all the nuclei
| |
| though the most of it goes into the ectodermal cells as in the case of gasteropods.
| |
| | |
| | |
| * These fragments of the yolk nucleus are larger and more easily seen in Molgula than in either or Cynthia.
| |
| | |
| | |
| | |
| This truly remarkable condition in which considerable portions of the cytoplasm are traceable to the nucleus is of the utmost theoretical importance. From
| |
| all sides the evidence has been accumulating that the chromosomes are the seat
| |
| of the inheritance material, until now this theory practically amounts to a demonstration. On the other hand, all students of the early history of the egg have
| |
| observed that the earliest visible differentiations occur in the cytoplasm, and that the
| |
| position, size and quality of the cleavage cells and of various organ liases are controlled by the cytoplasm. However, in the escape of large quantities of nuclear
| |
| material into the cell body and the /or/nation there of specific protoplasmic substances we have a possible mechanism for the nuclear control of the cytoplasm, and
| |
| when, as in the case of the ascidians and fresh water gastcropods, these substances
| |
| are definitely localized in the egg, and ca?i be traced throughout the development
| |
| until they cuter into the formation of particular portions of the embryo, a specific
| |
| mechanism for the nuclear control of development is at hand, and the manner of
| |
| harmonizing the pacts of cytoplasmic organization with the nuclear inheritance
| |
| theory is clearly indicated.
| |
| | |
| Of course substances which enter the nucleus and contribute to its growth
| |
| must reach it through the cytoplasm, but this does not signify that the same substances are given hack to the cytoplasm as are taken up from it ; on the contrary
| |
| we know that some of the substances which escape from the nucleus (e. g., oxychromatin) are not identical with those which enter it. Considering the necessity
| |
| of the nucleus in assimilation and regeneration, it seems most likely that differentiations of the cytoplasm proceed in the first instance from the nucleus; and, indeed,
| |
| in the case of the egg cell, some of the important cytoplasmic substances can
| |
| be actually seen to come from the nucleus. This does not indicate that these substances exist from the beginning in the nucleus; on the contrary there is direct and
| |
| visible evidence that they arise epigenetically. Such epigenesis, however, does not
| |
| signify lack of primary organization; on the other hand all the evidence favors the
| |
| view that back of the organization of the cytoplasm is the organization of the
| |
| chromosomes, which is definite, determinate and primary.
| |
| | |
| What has been said with regard to the genesis of the different substances of
| |
| the cytoplasm applies in the main to their localization. It is evident that this
| |
| localization is progressive, and that it arises epigenetically. But though we may
| |
| push back this localization to earlier and earlier stages and to simpler and simpler
| |
| forms we cannot entirely do away with it, even though it may be traced to polarity
| |
| and chemotropism. Some basis of localization must be present in the earliest
| |
| stages of the oogenesis, but this may possibly be little more than is found in the
| |
| body cells in general. It does not seem improbable that the differentiations and
| |
| localizations of the ovocyte and of the tissue cells are comparable in their manner
| |
| of origin. The most remarkable difference between the two is that the tissue
| |
| cells having reached the limit of their differentiation are incapable of further
| |
| development whereas the egg cell having reached the limit of its differentiation in
| |
| the ovary may. under the conditions of a free cell, begin another scries of differentiations which lead to the production of an organism.
| |
| | |
| | |
| | |
| 2. Factors of Localization, a. Cytoplasmic Movements.
| |
| | |
| | |
| Undoubtedly the most important of all the localizing factors so far recognized
| |
| are cytoplasmic movements. Such movements have been observed in unsegmented
| |
| eggs as well as in the cleavage cells, and they are generally associated with localization of unlike substances and frequently with cell division. The importance of
| |
| such movements in the differentiations of the egg I first recognized in Crepidula
| |
| (1899), where the movements of the cytoplasm during cleavage are very extensive. In the ascidian egg, on the other hand, these movements are most pronounced
| |
| in the period between the fertilization and the close of the first cleavage. In both
| |
| the ascidian and gasteropod these movements are definitely directed and bringabout a constant and typical form of localization of the materials of the egg.
| |
| | |
| The fact that these movements are definitely directed shows that they are dependent upon a constant organization of the cell ; their immediate cause is unknown.
| |
| So far as I have observed, these movements always begin soon after the disappearance of the nuclear membrane and the consequent escape of nuclear material into the
| |
| cell body. In the case of the gasteropods, I have suggested (1902) that one of the
| |
| characteristic movements of the telophase of division is due to the affinity of the
| |
| sphere material for oxygen. After the formation of this sphere material, during
| |
| each cell division, it moves to the surface of the cell and as nearly as possible to the
| |
| animal pole. If, however, the eggs be placed in water, from which the oxygen has
| |
| been removed by boiling, this movement to the surface does not take place. In the
| |
| ascidian the entrance of the spermatozoon seems to be the inciting cause of the
| |
| movement. The peripheral protoplasm (mesoplasm) rushes down to the point of
| |
| entrance and masses around the spermatozoon ; then when the latter moves toward
| |
| the posterior pole this protoplasm goes with it and is thus gathered into the crescent; finally, when the sperm nucleus moves in toward the centre of the egg the
| |
| larger part of this protoplasm remains at the surface, while a small portion of it is
| |
| drawn in with the sperm toward the center of the egg. In these movements, as well
| |
| as in the subsequent ones during cleavage, the mesoplasm remains near the surface of
| |
| the cell and in this respect resembles the sphere substance of the gasteropod egg.
| |
| The flowing of the protoplasm to meet the entering spermatozoon is a phenomenon of rather general occurrence. In most cases this leads only to the formation
| |
| of a small protoplasmic field around the sperm and sometimes to the formation of
| |
| an entrance cone ; in the ascidian practically all the protoplasm of the egg takes
| |
| part in this movement leaving the maturation spindles with only a trace of protoplasm around them. This withdrawal of the protoplasm from the animalpole
| |
| may be associated with the fact that there are no centrosome or asters in the
| |
| maturation spindles, whereas there is a large centrosome and aster in connection
| |
| with the sperm nucleus. Certainly the clear protoplasm is usually found in the
| |
| region of the asters. What the exact nature of this attraction between the protoplasm and the spermatozoon is, is not known, but the important point here is that
| |
| the cause of the remarkable movements of the protoplasm which follow the fertilization of the ascidian egg is not unique, and that the whole movement is peculiar
| |
| only because of its extent and the definite manner in which it is directed.
| |
| | |
| The movements which take place during cleavage are in part merely the
| |
| general movements which accompany cell division and in part they are of a localizing character. In the former class are the vortical movements which probably
| |
| cause the separtion of the chromosomes and the division of the cell body (Conklin,
| |
| 1902); in the latter are such movements as that which occurs at the close of the
| |
| first cleavage by which the clear protoplasm is carried from a central position into
| |
| the upper hemisphere of the egg. After the cleavage has begun the localizations
| |
| due to movement are strictly limited to the individual cells, no movements of a
| |
| localizing character occurring through cell w r alls.
| |
| | |
| b. Cell Division as a Factor of Localization.
| |
| | |
| This brings us to the much discussed question of the role of cell division in
| |
| development, and more particularly of the influence of cell division on phenomena of
| |
| localization. There can he no doubt that in many eggs the localization which
| |
| begins before cleavage continues during that process.
| |
| | |
| To a certain extent cleavage may he regarded as a localizing factor, but its
| |
| importance in this respect is certainly far less than that of the active movements
| |
| just described. Inasmuch as localizations may take place in the absence of cleavage or before it begins, and since many cleavages are non-differential it is evident
| |
| that there is no close nor necessary connection between the two. Furthermore
| |
| the cleavage planes do not always coincide with the lines of localization ; this is
| |
| shown especially well in the ascidian. where the localization in the unsegmented
| |
| egg is particularly distinct. Thus the cleavage planes do not follow closely the
| |
| boundaries of the crescent ; the first and second cleavage planes are placed symmetrically with reference to the crescent, but they do not coincide with any of its
| |
| boundary lines. The third cleavage plane lies above the upper border of the
| |
| crescent when first formed; later the crescent extends up to the equatorial plane
| |
| so that the cleavage plane and the upper boundary of the crescent coincide (fig. 31,
| |
| 32). The fourth cleavage cuts off the median posterior crescent cells from the
| |
| lateral ones, but leaves an area of yolk in both of these cells (fig. 37). In the
| |
| median posterior cells this is a small wedge-shaped mass of yolk which is later
| |
| covered and obscured by the yellow crescent substance (fig. 39). The neural plate
| |
| arises on the anterior side of the egg from cells which lie both above and below the
| |
| equator, or plane of the third cleavage ; these neural plate cells are rich in protoplasm, and correspondingly the area from which they arise is richly protoplasmic.
| |
| The third cleavage cuts right through this protoplasmic area leaving a portion of
| |
| it above and a part below the equator. In the 8-cell and 16-cell stages the anterior
| |
| dorsal cells contain both neural-plate and chorda substance; the portion of each of
| |
| these cells turned toward the equator is protoplasmic, that turned toward the vegetal
| |
| pole yolk-laden (A 63 , AH figs. XVII. XIX. 110, 117). At the next cleavage
| |
| these two portions are separated, the upper protoplasmic part becoming the neural plate cells. A 74 and A rs , while the lower yolk-laden part becomes the chorda cells,
| |
| A 73 and A 77 . The chorda-neural substances arc thus contained in the same cells
| |
| until the sixth cleavage, though their substances are distinct at a much earlier
| |
| period. Still other instances might be cited to show that the planes of localization and the planes of cleavage do not always coincide. This is in part due to the
| |
| fact that the boundaries of the different kinds of germinal material, e.g.. the yellow
| |
| protoplasm of the Cynthia egg, are not as sharp as are the boundaries of the cells,
| |
| and consequently the cleavage furrows cannot precisely separate different kinds of
| |
| germinal material. Nevertheless the cleavage planes are, under normal conditions,
| |
| constant in position and character and bear a constant relation to the planes of differentiation. But that this relationship is not a casual one is further indicated by
| |
| experimental studies on cleavage in which the position of the cleavage furrows may
| |
| be altered without altering the localization of germinal materials or the typical form
| |
| of development. Therefore the factors which determine localization and those
| |
| which determine the form of cleavage are more or less independent.
| |
| | |
| All of these tacts speak unmistakably for the view that localization is more
| |
| fundamental than cleavage as Whitman (1893) has so ably maintained, and that
| |
| such correspondence as may exist between the two is of secondary origin and of
| |
| minor importance. Nevertheless the extreme constancy of cleavage forms shows
| |
| that we have here a phenomenon, which if of secondary importance to germinal
| |
| localization, is still of real significance. I have shown that in Crepidula the
| |
| cleavage is a localizing factor, though secondary in importance to protoplasmic
| |
| movement, and it seems probable that Wilson (1003) is right when he argues that
| |
| the relative isolation produced by cleavage gives opportunity for the increase of
| |
| any initial differences which may exist in the cells at the time of their formation.
| |
| | |
| Finally it must be concluded as a result of both observation and experiment
| |
| that the type of cleavage is less constant and less fundamental than the type of
| |
| localization, but that cleavage may itself be a factor in the progressive specification
| |
| of cells {cf. Wilson, Lillie, Conklin, et al.).
| |
| | |
| E. Types of Germinal Localization ; Evolution of Types.
| |
| | |
| The wonderful resemblances in the germinal localization of annelids and
| |
| mollusks, as shown especially in the cleavage, have been repeatedly commented
| |
| upon. Furthermore this localization is for shadowed in the egg before cleavage
| |
| begins, and this suggests the inquiry as to whether the resemblances between types
| |
| of localization grow closer as one approaches the ovocyte, and whether the manner as well as the results of localization are comparable in the different types. At
| |
| present our knowledge of the localization in these earliest stages of development
| |
| is very incomplete, and a comparison can be drawn only between annelids, mollusks, ctenophores, echinoderms, ascidians and possibly nemerteans and nematodes.
| |
| | |
| In most of these phyla a peripheral layer of protoplasm is present before
| |
| maturation, which after maturation and fertilization collects at one or both poles
| |
| of the egg; also with the possible exception of the ctenophores and nematodes, there is remarkable uniformity in the localization of the substances of the germinal
| |
| layers in all of these groups, the ectodermal substances being located in the upper
| |
| hemisphere, and the endodermal and mesodermal in the lower hemisphere of the
| |
| | |
| egg. But in the localization of important organ bases there are many differences
| |
| between these phyla.
| |
| | |
| | |
| | |
| 1. Annelid-Mollusk Type.
| |
| | |
| The pattern of localization in annelids and mollusks is very similar during the
| |
| cleavage stages and, so far as can be judged from present knowledge, it is much the same
| |
| in the unsegmented eggs of these two phyla. The fact that the ectoderm, mesoderm
| |
| and endoderm come from cells which are identical in origin, position and number;
| |
| that the umbrella, prototroch, cerebral ganglion, sub-oesophageal ganglion, mesodermal bands, blastopore, stomodaium and intestine come from corresponding region of
| |
| the egg in the two groups, these facts speak strongly in favor of the regional homologies of the eggs of these phyla, whatever may be thought of their cell homologies
| |
| (Conklin, 1897; Child, 1900). But regional homologies as well as cell homologies
| |
| must be based upon similarities of germinal localization, and we would, therefore,
| |
| be justified in concluding that the types of localization were similar in the unsegmented eggs of annelids and mollusks even in the absence of any direct knowledge
| |
| upon that subject. But the experiments of Crampton (1896) on Illyanassa and
| |
| of Wilson (1904) on Dentalium as well as the observations of Lillie (1899, 1901)
| |
| on [/mo, and my own observations on localization in the eggs of Crepidula, Pliysa,
| |
| Planorbis and Limncva furnish considerable information as to the time, the manner
| |
| and the nature of localization in the molluscan egg during and before cleavage, while
| |
| numerous works on the cell-lineage of the annelids as well as the observations of
| |
| Wheeler (1897), Driesch (1890) and Carazzi (1904) on the unsegmented egg of
| |
| Myzosioma show that the nature of localization is here very similar to that found
| |
| in the mollusks.
| |
| | |
| In all of these cases the only formative substances which are directly recognizable before cleavage are those of the future germ layers. In the main the ectodermal substances are located in the upper hemisphere and the endodermal in the
| |
| lower, though Wilson (1904) has found that the apical organ does not form in the
| |
| larva of Dentalium when the polar lobe at the vegetal pole is removed. The mesodermal substances are also located in the lower hemisphere, and since the primary
| |
| mesoderm cell (4d) always come from the left posterior macromere of the 4-cell
| |
| stage and from the posterior blastomere of the 2-cell stage it may be inferred that
| |
| immediately before cleavage it lies posterior to the vegetal pole; whether it may be
| |
| located exactly at the vegetal pole in still earlier stages and then later shift to the
| |
| posterior side, as in ascidians, cannot be determined at present.
| |
| | |
| When a polar lobe is present the mesodermal substance is probably located in
| |
| it. Crampton (1896) found in Illyanassa that the mesoblast cell (4d) did not form
| |
| when the lobe had been removed; Wilson (1904) holds that in Dentalium the substance of the lobe is allotted to both the first and second somatoblasts'(2d and 4d),
| |
| and that its size is proportional to the size of those cells and of the parts to which they give rise. He calls particular attention to the fundamental resemblances
| |
| between the eggs of Dcntalium and of Mysostoma'va the matter of the polar
| |
| lobe and the " pillar of protoplasm." Furthermore this lobe is comparable to
| |
| the polar rings of leeches and oligochastes. Such a lobe, although present in some
| |
| annelids and mollusks, is not present in all of them, and this would at first thought
| |
| seem to mark some important difference in localization. But the presence or
| |
| absence of such a lobe probably indicates no fundamental dissimilarity in the localization, but rather variations in the surface tension and fluidity of different eggs.
| |
| Although there are many interesting differences between various annelids and
| |
| mollusks in the size of the polar lobe, of the blastomeres and of larval organs, these
| |
| differences mark variations in the proportions of parts rather than in the type of
| |
| localization. In all known cases among annelids and mollusks corresponding organs
| |
| arise from corresponding regions of the egg.
| |
| | |
| It may be concluded also from the work of Wilson (1903) and Yatsu (1904) on
| |
| Cerebratulns that the character of the localization in the nemertine egg is essentially
| |
| like that of the annelid and mollusk, though many of the details of localization
| |
| are less accurately known in this case than in the others named.
| |
| | |
| | |
| 2. Ctowphore Type.
| |
| | |
| If Fischel (1903) is right regarding the localization which he ascribes to
| |
| the unsegmented ctenophore egg there is one fundamental difference between the
| |
| ctenophore and other animals whose types of localization are known. On the
| |
| authority of Metschnikoff he derives the mesoderm (somewhat doubtfully it must
| |
| be said) from the micromeres at the upper pole of the egg, and consequently in his
| |
| fig. 21 (p. 708) he localizes the mesodermal material at the upper pole of the unsegmented egg. A zone beloAv this, reaching to the equator or a little lower, represents the ectodermal substance, and in it is located the material for the ciliated
| |
| plates. At the lower pole and in the central part of the egg is the material substratum of the endoderm. In all other well established cases the ectodermal substances lie near the animal pole, while the mesodermal and endodermal substances
| |
| lie near the vegetal pole. Inasmuch as an apical sense organ is formed at the
| |
| animal pole in ctenophores in much the same way as in annelids, mollusks and
| |
| nemerteans, it is difficult to resist the conclusion that the localization of the ectodermal, mesodermal and endodermal substances in the ctenophore egg will ultimately be found to be similar to that which prevails in other types.
| |
| | |
| 3. Echitiodcrm Type.
| |
| | |
| The form of localization in the eohinoderm egg, as shown by Boveri's
| |
| (1901) work on Strongylocentrotits, is in many respects similar to the annelidmollusk-nemertean type. In this case, however, the mesoplasm is located at the
| |
| lower pole of the egg and is sorrounded hy an equatorial zone of endoplasm, whereas
| |
| in annelids and mollusks, after the first two cleavages, the endoplasm lies at the
| |
| lower pole, and the mesoplasm on the posterior side of this pole, and in one only of the first four blastomeres. Whether the mesoplasm lies on one .side of the vegetal
| |
| pole in the unsegmented egg of annelids and mollusks cannot be affirmed on direct
| |
| evidence, but it seems not unlikely that this is the case. If this be true there is
| |
| here a difference between echinoderms and annelids or mollusks, in the form of
| |
| localization, though it is by no means impossible to derive one type from the other.
| |
| | |
| | |
| | |
| 4. Ascidian Type.
| |
| | |
| Finally, the type of localization in the ascidian egg differs in many respects
| |
| from that of the other phyla mentioned, though showing certain general resemblances to all of them and particularly to the annelid-mollusk type. Castle has
| |
| called attention to the fact that there are no important resemblances between
| |
| ascidians and annelids in their cell-lineage, and with this opinion I entirely agree.
| |
| Nevertheless, in the localization of ectoplasm, mesoplasm and endoplasm in the
| |
| unsegmented egg there are many similarities between these phyla, hut in the position of specific organ bases the differences are quite notable.
| |
| | |
| Among ascidians the ectoplasm which escapes from the germinal vesicle at the
| |
| animal pole does not remain there, as in the fresh-water snails, but flows rapidly
| |
| to the lower pole, then to the posterior side of the egg, then into the center and finally
| |
| into the upper hemisphere of the egg; in other phyla the ectoplasm becomes directly localized at the upper pole, here only indirectly. The mesoplasm is first segregated
| |
| at the lower pole in a manner which recalls the egg of Strongylocentrotus, and then
| |
| finally becomes localized on the posterior side, a result which somewhat resembles
| |
| the condition in annelids and mollusks ; in the ascidians the cells of the mesodermal crescent lie in the posterior lip of the blastopore, in annelids and mollusks the
| |
| teloblasts and mesodermal bands lie in a similar position in the early gastrula
| |
| stages, but owing to the closure of the blastopore from behind forward they are
| |
| ultimately removed some distance from the blastopore lip. The mesoderm and
| |
| mesodermal organs may therefore be said to arise from corresponding regions of the
| |
| egg in these two groups of animals (text figs. XXXIX, XL). The endoplasm also
| |
| is localized in corresponding regions of the egg in these phyla.
| |
| | |
| When, however, we come to compare the positions in the eggs of these phyla
| |
| of important organ bases the differences are very marked. For example, in
| |
| annelids and mollusks the apical plate and cerebral ganglion are formed near the
| |
| animal pole, the sub-oesophageal ganglia from the ventral plate, which is derived
| |
| from the cell 2d, lying not far below the equator on the posterior-dorsal side and
| |
| just above the mesodermal teloblasts (text fig. XL) ; subsequently in the concresence
| |
| of the posterior lip of the blastopore, the bases of the sub-oesophageal ganglia are
| |
| carried to the ventral side. The nervous system of annelids and mollusks thus has
| |
| a double origin, one portion arising from the region of the animal pole, the other
| |
| from the posterior pole, and these two portions subsequently become connected
| |
| together by commissures which surround the oesophagus. Jn the ascidian the
| |
| entire central nervous system is formed as a continuous plate which lies along the
| |
| anterior side of the egg, stretching from a point about 60 from the animal pole and from the equator to a point a little below the equator (text fig. XXXIX); no
| |
| portion of the nervous system comes from the region of the animal pole and none
| |
| from the posterior pole. Furthermore, the mouth, which is here a new formation
| |
| and has nothing to do with the blastopore, does not open through the nerve plate
| |
| but lies between the anterior end of the neural plate and the animal pole.
| |
| | |
| We have here differences of a fundamental order, even in the earliest stages of
| |
| development, between the vertebrate, or rather chordate, and the invertebrate; the
| |
| early development throws no light upon the way in which the one may have been derived from the other. It is of course possible to conceive of a condition in which the
| |
| nervous system surrounded the entire blastopore as a ring, which in the case of the
| |
| annelids underwent concrescence from behind forward, thus forming the ventral
| |
| plate and ganglia, but which in the chordates underwent concrescence from in front backwards. But however probable such a theory may be it finds little support in
| |
| the early development of ascidians. It is true that a nerve ring has been described
| |
| as surrounding the blastopore in ascidians, but I have not been able to find evidence
| |
| of its existence. Furthermore, there is no evidence in the development of ascidians
| |
| that there is any concrescence of the anterior lip of the blastopore ; on the contrary
| |
| the anterior lip grows backward over the archenteron as rapidly in the mid-line as
| |
| at the sides, a view in which practically all writers on ascidian embryology agree.
| |
| Finally, the lack of an apical plate and cerebral ganglion at the animal pole in the
| |
| ascidian constitutes a notable difference from the condition found in most invertebrates. In his great work on Sa/pa, Brooks (1893) has shown in masterly fashion
| |
| | |
| | |
| | |
| | |
| | |
| Figs. XXXIX, XL. Diagramatic representations of the types of germinal localization in ascidians
| |
| and annelids. Mesodermal substance is shaded by lines, neural substance by fine stipples, and chorda
| |
| material by coarse stipples. Fig. XXXIX, the ascidian type; egg viewed from right side. The mesoplasm, composed of mesenchyme (m'ch) and muscle substance (msl, is represented in its final position,
| |
| which it assumes before the first cleavage. The neural plate (n. p.) and chorda (eh.) substances are not
| |
| distinguishable in the unsegmented egg. but are here shown in the positions in which they appear at the
| |
| 2-cell stage; the chorda and mesenchyme substances should be shown as meeting on the side of the egg,
| |
| thus forming a chorda-mesenchyme ring around the endoderm. The ectoplasm (ect.) and endoplasm
| |
| (end.) are localized, as here represented, at the close of the first cleavage. Fig. XL, the annelid type;
| |
| egg viewed from left side. The substances of the first and second somatoblasts (the former stippled along
| |
| one border, the latter shaded by lines) are shown in the positions in which these cells are ultimately
| |
| formed ; in the unsegmented egg the lobe which contains the substances of these cells lies nearer the
| |
| lower pole. The substances of the cerebral ganglion (c. g.), ventral ganglia (v. g.) and prototroch (proto.)
| |
| are not distinguishable in the unsegmented egg, but are shown in the regions to which they may be
| |
| traced by means of the cell lineage.
| |
| | |
| | |
| | |
| | |
| the weakness dl' ! he annelidan hypothesis of the origin of chordates and has adduced
| |
| much evidence in favor of the view that the great metazoic stems run back to
| |
| simple and minute pelagic ancestors whose common meeting placemust lie be found
| |
| | |
| in still more recent times. The earliest dift'erentations of the egg seem to me to
| |
| favor this view.
| |
| | |
| In conclusion, then, it seems necessary to recognize several types of cytoplasmic localization. Between annelids and mollusks the similarities of localization
| |
| extend to the bases of numerous parts and organs, thus confirming the view of
| |
| the phylogenetic relationship of these two phyla based upon the resemblances in their
| |
| cleavage stages and larvae. Between the annelid-mollusk type of localization and
| |
| tin' types found in the other phyla enumerated there are general agreements in the
| |
| localization of the materials of the germinal layers, but few, if any, resemblances
| |
| which extend to the bases of particular organs. The annelids do not approach
| |
| the chordates nor the echinoderms in the earliest stages of localization any more
| |
| closely than in their cleavage stages or later development. In all respects in which
| |
| localizations differ in the eggs of these animals they resemble the later differences
| |
| in their embryos. In short, there is no convergence toward a co?n)non type of localization as one goes back to earlier and earlier stages in the ontogeny.
| |
| | |
| Important results flow from this conclusion, for the doctrine that " Ontogeny
| |
| is a short recapitulation of Phylogeny" assumes that there is such convergence
| |
| toward a common type of structure in the early stages of development. If there be
| |
| no such convergence the causes of the resemblances which exist between certain
| |
| eggs, cleavage stages, embryos, larvas and adults must be sought in some other
| |
| direction. Students of the cell-lineage of annelids and mollusks have maintained
| |
| that homologies of cleavage must be due to similarities in the protoplasmic structure of the cleavage cells. The same must also be said of the organization of the
| |
| egg before cleavage begins. Similarities in the material substance of the egg and
| |
| in the form of its localization must lie at the bottom of all later appearing similarities. And this fact, upon which all students of cell lineage have insisted,
| |
| furnishes a possible explanation, as Morgan (1903) has recently pointed out, of
| |
| the resemblances between the embryos of related forms.
| |
| | |
| Speculations as to the origin and evolution of tyj3es of germinal organization are
| |
| likely to be more interesting than valuable in the present state of our knowledge.
| |
| Wilson (1892) first suggested that the localization of the materials of embryonic
| |
| parts or organs in certain cleavage cells was an illustration of the principle of "precocious segregation" first propounded by Lankester and afterward elaborated by
| |
| Hyatt, in its application to palaeontology, under the title of ''the law of acceleration." Lillie (1895) maintained that "it is parallel precocious segregation which
| |
| conditions cell homologies." and he further showed (1899) that the size and rate of
| |
| division of individual cells in every case possesses pi'ospeetive significance; in
| |
| short, that the cleavage forms are beautifully adapted to produce a given type of
| |
| adult structure. Recently Wilson (1903, 1904) has expressed the view that the
| |
| earliest differentiations and localizations of the egg, even before cleavage begins, are
| |
| examples of this same principle of "precocious segregation."
| |
| | |
| | |
| Although this principle is carefully stated so as not to directly affirm that the
| |
| organization of the egg is the result of the organization of the adult, or that the
| |
| adaptations of the early development have arisen secondarily after the adult structure was estafTHshed, these ideas are nevertheless plainly implied. The early
| |
| appearance of differentiations is usually explained as a " throwing hack of adult
| |
| characters upon the egg." The whole life cycle is viewed from the standpoint of
| |
| the adult ; the embryo and germ exist for the purpose of producing a certain end ;
| |
| the adult is primary, the germ secondary. But do not all such ideas put the cart
| |
| before the horse ? What is the evidence that any inherited modification of an adult
| |
| structure can arise without an antecedent modification of the germ ? We know that
| |
| the adult is moulded upon the egg, that specific modifications of the germ do, in
| |
| some cases, produce specific modifications of the adult, but the converse proposition
| |
| is certainly not established. "Precocious segregation" represents the backward
| |
| rather than the forward look; it is a teleological rather than a causal explanation.
| |
| | |
| As there can be no transmission of heritable qualities from one generation to
| |
| another except through the germ cells, so there can be no evolution of adult forms
| |
| except through the evolution of the germ cells. Any inherited modification of a
| |
| species implies some modification of the germ cells of the species. Even " acceleration" or " precocity" must be due to a modification of the germ in its earliest
| |
| stages, a modification of some unknown sort which hastens differentiation.
| |
| | |
| It cannot be maintained that all those animals in which differentiations and
| |
| localizations are present in the unsegmented egg are, for that reason, debarred
| |
| from any further evolution, but if this be not true then it must follow that the
| |
| type of egg organization must undergo modifications during the course of evolution,
| |
| and granted this we have no need of the principle of " parallel precocious segregation " for explaining any of the homologies of the earl}' development. If the
| |
| resemblances between annelids and mollusks are not due primarily to the similarities in the adults or larva? or cleavage stages, but to phylogenetic similarities in the
| |
| organization of the unsegmented egg, we have in this initial resemblance a sufficient
| |
| explanation of all later resemblances, whereas if we reverse this procedure and
| |
| hold that the similarities of the adults or larva? are the causes of the likenesses in
| |
| the earlier stages we must of necessity resort to some such teleological principle
| |
| as "precocious segregation" for an explanation.
| |
| | |
| In view of the fact that there are such definite types of differentiation and
| |
| localization in the eggs of many animals and that the causes which lead to the
| |
| evolution of animals must operate through modifications of this organization, the
| |
| character and manner of such modification become problems of the first importance.
| |
| If the nuclear inheritance theory is true, such modifications must in the first instance affect the chromosomes ; but how and in what respect is wholly unknown. In
| |
| the case of the cytoplasm it is evident that such modifications may concern the character, or quality, of the differentiations and the place and manner of their localization. Modification of any of these might be expected to produce modifications
| |
| in the resulting animal.
| |
| | |
| | |
| Relatively slight modifications of this localization, however produced, may lead
| |
| to profound modifications of the resulting embryo and adult. I have elsewhere
| |
| (1903) shown reason for believing that the cause of inverse symmetry is to be
| |
| found in the inverse organization of the egg, and that this inverse organization may
| |
| pjssibly be produced by the maturation of the egg at opposite poles in dextral and
| |
| sinistral forms. This ease shows that one of the most remarkable forms of mutation with which we are acquainted may be the result of modifications in the localization of germinal substances in the unsegmented egg.
| |
| | |
| One of the great difficulties in explaining the origin, on evolutionary principles, of different phyla has been the dissimilar locations of corresponding organs or
| |
| parts. These difficulties are well illustrated by the theories which attempt to
| |
| derive the vertebrates from annelids, or from any other invertebrate type. Without
| |
| assuming to defend any of these theories it may here be pointed out that if evolution takes place through modifications of germinal organization, it is no more
| |
| difficult to explain the different location of parts than their different qualities.
| |
| Changes in the relative positions of parts, which would be impossible in the adult,
| |
| may be readily accomplished in the unsegmented egg, as is shown by cases of inverse
| |
| symmetry. The question is here raised, whether some similar sudden alteration of
| |
| germinal organization may not lie at the basis of the origin of new types.
| |
| | |
| | |
| | |
| SUMMARY.
| |
| | |
| | |
| I. Ascidian Embryology.
| |
| | |
| 1. The orientation of the ascidian egg and embryo adopted by Van Beneden
| |
| and Julin is correct, that of Seeliger, Samassa and Castle is wrong (pp. 20-37).
| |
| | |
| 2. The cell-lineage given by Castle is correct for the early stages; from the
| |
| 48-cell stage on it is wrong (pp. 5 65 U ) .
| |
| | |
| 3. The egg axis corresponds very nearly with the gastrular axis; during the
| |
| closure of the blastopore this axis is shifted so that it is no longer dorso-ventral as
| |
| in the early stages, but is antero-ventral and postero-dorsal in direction in the larva
| |
| (pp. 73, 75-77).
| |
| | |
| 4. The relation of the germinal layers to the cleavage planes is very different
| |
| from the account given by Van Beneden and Julin and by Castle, and is more
| |
| nearly in accord with that of Seeliger, Davidoff and Samassa. All cells above the
| |
| equator (3d cleavage plane) are ectodermal and neural plate cells; all below are
| |
| endodermal, mesodermal and neural plate (pp. 47-48).
| |
| | |
| ''5. The factors of gastrulation are (a) change of shape of the cells of the
| |
| animal and vegetal hemispheres, (b) overgrowth of the marginal cells (pp. 64-65).
| |
| Peculiarities of the gastrula are foreshadowed in the egg at a very early stage
| |
| (pp. 45. 50. 59).
| |
| | |
| / 6. The muscle and mesenchyme cells arise from a common base, the mesodermal crescent, which surrounds the posterior side of the egg just below the
| |
| equator (pp. 19-21); ultimately these cells surround the posterior margin of the blastopore (pp. 51-55); the most laterally situated of these cells become the large
| |
| muscle cells of the tail of the tadpole, the portion of the crescent lying nearest the
| |
| dorsal mid-line becomes mesenchyme (pp. 61-07). In the overgrowth of the dorsal
| |
| lip of the blastopore the muscle cells become separated from the mesenchyme (pp.
| |
| 69, 84-87).
| |
| | |
| 7. The chorda arises at the anterior border of the blastopore from yolk-laden
| |
| cells which resemble endoderm (pp. 53, 61, 62, 70-72, 83, 84).
| |
| | |
| 8. The neural plate arises on the anterior side of the egg from cells of both
| |
| hemispheres ; it extends from the margin of the anterior lip of the blastopore to a
| |
| point about one-third of the way from the equator to the animal pole (pp. 52-54,
| |
| 61-63, 66-68, 70, 72, 73. 82, 83).
| |
| | |
| 9. The nervous and muscular systems do not arise from a common base as
| |
| claimed by Castle; there is no nerve ring around the blastopore (pp. 61, 72, 73).
| |
| | |
| ' 10. A comparison of the early development of ascidians with that of Amphioxus and amphibians shows that there is fundamental agreement among them in
| |
| axial relations of egg and larva, in bilaterality of cleavage, in the method of closure
| |
| of the blastopore and probably in the origin and position in the embryo of the
| |
| neural plate, the chorda and the mesoderm (pp. 73-87).
| |
| | |
| II. Cytologic al Results.
| |
| | |
| 11. The maturation spindles are peculiar; they have no centrosomes nor
| |
| asters ; they are formed entirely within the nuclear area from nuclear linin and
| |
| chromosomes ; their fibres at first radiate in all directions, and finally they form a
| |
| barrel-shaped spindle. The chromosomes separate without any possible influence
| |
| from centrosomes or traction on the part of spindle fibres (pp. 15, 16).
| |
| | |
| 12. In the first and second cleavages a small nuclear spindle like those present during maturation, lies between the two large asters, and in Ciojia it is quite
| |
| distinct, from them. The separation of the daughter chromosomes takes place here
| |
| as in the maturation divisions (pp. 40, 41).
| |
| | |
| 13. The spermatozoon enters near the lower pole and rotates after entering
| |
| the egg so that its centrosome is directed forward ; the centrosome is derived from
| |
| the middle piece of the spermatozoon and can be followed without interruption until
| |
| it divides, at right angles to the copulation path, and gives rise to the sperm amphiaster and finally to the cleavage centrosomes (pp. 22-24). A netrum is formed in
| |
| the division of all centrosomes (p. 40).
| |
| | |
| III. Organization of the Egg.
| |
| | |
| 14. In the ovocyte of Cynthia parti/a there is a peripheral layer of yellow
| |
| protoplasm, a central mass of gray yolk, and a large clear germinal vesicle, which is
| |
| eccentric toward the animal pole. These same parts are jiresent in the eggs of
| |
| other ascidians, but are differently colored (pp. 11, 12).
| |
| | |
| 15. When the wall of the germinal vesicle dissolves at the beginning of the
| |
| maturation divisions a large amount of clear protoplasm, containing dissolved oxyebromatin, is liberated into the cell body. This clear protoplasm is eccentric
| |
| toward the animal pole and is distinct from the yolk and peripheral layer (pp.
| |
| 13, 17).
| |
| | |
| lii. Immediately after the entrance of the spermatozoon the yellow and clear
| |
| protoplasm How rapidly to the lower pole, where the yellow protoplasm collects
| |
| around the point of entrance; the clear protoplasm lies at a deeper level. The
| |
| yellow protoplasm then spreads out until it covers the surface of the lower hemisphere. This flowing of protoplasm to the point of entrance of the sperm is comparable with what takes place in many animals, though here much more extensive
| |
| and rapid than elsewhere (pp. 1921, 77).
| |
| | |
| 17. The withdrawal of protoplasm from the upper pole leaves the maturation
| |
| spindles closely surrounded by yolk. The polar bodies are thus formed at the
| |
| middle of a yolk-rich hemisphere, which is, however, the animal pole and not
| |
| the vegetal pole as was claimed by Castle (pp. 19-21, 29, 30, 36, 37, 87-90).
| |
| | |
| 18. The sperm nucleus moves from the point of entrance toward the equator
| |
| in a path which is apparently predetermined. Tins path lies in the plane of the
| |
| first cleavage and the point, just below the equator, at which the sperm nucleus
| |
| stops in its upward movement, becomes the posterior pole of the embryo. The
| |
| median plane and the posterior pole are probably not determined by the path of the
| |
| spermatozoon, but by the structure of the egg. All the axes of the future animal
| |
| are now clearly established, antero-posterior, right-left, dorso-ventral (pp. 22, 26,
| |
| 90-93).
| |
| | |
| 19. As the sperm nucleus moves to the posterior pole the clear and the yellow
| |
| protoplasm move with it ; the latter collects into a yellow crescent with its middle
| |
| at the posterior pole and its horns extending about halfway around the egg just
| |
| below the equator. This position it retains throughout the whole development,
| |
| giving rise to the muscle and mesenchyme cells mentioned in 6 (pp. 19-21, 97, 98).
| |
| | |
| 20. After the sperm and egg nuclei have met at the posterior pole they move
| |
| in toward the center of the egg and the clear protoplasm goes with them ; the outplace where the latter remains in contact with the surface is along the upper border
| |
| of the crescent. At the close of the first cleavage the nuclei and clear protoplasm
| |
| move into the upper hemisphere, and thereafter, throughout development, this
| |
| hemisphere contains most of the clear protoplasm and gives rise to the ectoderm
| |
| (pp. 20, 21, 42, 102).
| |
| | |
| 21. The yolk which before maturation was central in position is shifted
| |
| toward the animal pole when the protoplasm flows down to meet the spermatozoon ;
| |
| when the sperm nucleus and surrounding protoplasm move to the posterior pole
| |
| the yolk is moved down around the anterior side of the egg to the lower pole, and
| |
| when the clear protoplasm moves into the upper hemisphere the yolk is largely
| |
| collected in the lower hemisphere. This yolk rich area gives rise to the endoderm
| |
| (pp. 20', 33-35, 12, 102).
| |
| | |
| 22. At the close of the first cleavage the principal germ regions of the embryo
| |
| are visible in their definite positions and proportions, viz. : the muscle-mesenchyme crescent and the ectodermal and endodermal areas. The chorda and neural plate
| |
| areas are also visibly different from surrounding areas at this stage (pp. 42, 50, 95,
| |
| 97, 98, 108).
| |
| | |
| 23. In many eases the cleavage planes do not follow the lines of differentiation but cut across them. Although cleavage is, under normal conditions, constant
| |
| in form, it is less constant and fundamental than the type of localization, and the
| |
| two are relatively independent (pp. 103, 104).
| |
| | |
| 24. The chief factor of localization is protoplasmic flowing; cell division is a
| |
| factor of subordinate value (pp. 102-104).
| |
| | |
| 25. Experiments which demonstrate the totipotence of blastomeres or regions
| |
| of the egg prove nothing with regard to the presence or absence of differentiation
| |
| in those parts. Some eggs with a high degreee of differentiation have at the same
| |
| time great capacity for regulation, e.g., those of ascidians ; 1 others with no greater
| |
| differentiation have little regulative capacity, e. g., ctenophores and mollusks.
| |
| Therefore the potency of any part of an egg or embryo is no satisfactory measure of
| |
| the degree of its differentiation (pp. 93-95).
| |
| | |
| 26. The organization of the ovocyte is not the initial organization. The yellow protoplasm (mesoplasm) of the Cynthia egg is probably derived, at least in part,
| |
| from sphere material (archoplasm) which arose from the nucleus at the last ovogonic division. / The yolk (endoplasm) is formed by the activity of the "yolk
| |
| matrix" (Crampton) which also is probably sphere material. The clear protoplasm
| |
| (ectoplasm) is derived from the germinal vesicle at the first maturation division. Thus
| |
| many important regions of the egg come, at least in part, from the nucleus, and a
| |
| method is therein suggested .of harmonizing the facts of cytoplasmic localization
| |
| with the nuclear inheritance theory (pp. 99-101)./
| |
| | |
| 27. There are several distinct types of germinal localization. /The annelidmollusk type does not approach that of chordates or echinoderms in the earliest
| |
| stages of localization more closely than in the cleavage or gastrular stages. There is
| |
| no convergence toward a common type in the earliest stages (p. 104-109).
| |
| | |
| 28. Embryonic repetitions (recapitulations), as well as many other homologies,
| |
| probably result from similarities of egg organization common to each type (p. 109).
| |
| | |
| 29. "Precocious segregation" is not a satisfactory explanation of the origin
| |
| of germinal organization (pp. 109, 110).
| |
| | |
| / 30. The evolution of animals must be accompanied by an evolution of the
| |
| type of germinal organization ; modifications of this organization are probably the
| |
| immediate causes of evolution. Transformations which would be impossible in
| |
| adults are readily brought about by modifications in the organization of the egg
| |
| {e.g., inverse symmetry). Perhaps profound mutations or even the origin of distinct types may be so explained (pp. 110, 111).
| |
| 1 See foot-note p. 95.
| |
| | |
| | |
| | |
| ==Literature Cited==
| |
| | |
| | |
| BANCROFT, F. \V. (1899).
| |
| | |
| Ovogenesis in Distaplia occidentalis, Ritter. Bull. Mus. Camp. Zoo/, xxxv.
| |
| BENEDEX, E. van kt CH. JULIN, (1884).
| |
| | |
| La segmentation chez les ascidens dans ses rapportes avec l'organization de la larve. Arch, de Biol., v.
| |
| BENEDEN, E. van et CH. JULIN, (1886).
| |
| | |
| Recherches snr la Morphologie des Tuniciers. Arch, de Biol., vi.
| |
| BENEDEN, E. van et A. NEYT, ( L887).
| |
| | |
| Nouvelles recherches sur la fecondation et la division mitosique chez 1'Ascaride Megalocephale. Bull.
| |
| /'Acad. Row Belgique, 3eser., XI V.
| |
| BIGELOVV, M. A (1902 .
| |
| | |
| The Early Development of Lepas. A study of Cell-Lineage and Germ Layers. Bull. Mus. Comp. Zoo/., xl.
| |
| BLOCHMANN, FR. (1882). ,
| |
| | |
| Beitriige zur Kenntniss der Gasteropoden. Zeit. wiss. Zoo/., xxxviii.
| |
| BOYERI, TH. (18S7, 1SSS, 1890).
| |
| | |
| Zellen Studien, I, II, III. Jena.
| |
| BOVERI, TH. (1901).
| |
| | |
| Ueber die Polaritat des Seeigeleies. Verh. Phys. Med. Ges. Wurzburg, xxxiv.
| |
| BOVERI. TH. (1901).
| |
| | |
| Die Polaritat von Ovocyte, Ei und Larve des Strongylocentrotus lividus. Zoo/. Jahrb., .xiv.
| |
| BROOKS, W. K. (1S93).
| |
| | |
| The Genus Salpa. Baltimore.
| |
| CARAZZI, D. (1904).
| |
| | |
| Ricerche embriologiche e citologiche sull 'uovo di Myzostoma glabrum Leuckart. Monitoro. Zool.
| |
| Italiano, xv.
| |
| CASTLE, W. E. (1894).
| |
| | |
| On the Cell-Lineage of the Ascidian Egg. A preliminary notice. Proc. Amer. Acad. Arts and Sci., xxx.
| |
| CASTLE, W. E. (1896).
| |
| | |
| The Early Embryology of Ciona intestinalis, Flemming (L. ). Bull. Mus. Comp. Zoo/., xxvii.
| |
| CHABRY, L, (1887
| |
| | |
| Contribution a l'embryologie normal et teratologique des Ascidies simples. Jour. Ana/, et Physiol, xxiii.
| |
| CHILD, C. M. (1900).
| |
| | |
| The Early Development of Arenicola and Sternaspis. Arch. Entw. Meek., ix.
| |
| C( INKLIN, E. G. (1897)
| |
| | |
| The Embryology of Crepidula. Jour. Morph., xiii.
| |
| CONKLIN, E. G. (1899).
| |
| | |
| Protoplasmic Movement as a Factor of Differentiation. Woods Ho// Bio/. Lec/ures, 1S98.
| |
| CONKLIN, E. G. (1902).
| |
| | |
| Karyokinesis and Cytokinesis in the Maturation, Fertilization and Cleavage of Crepidula and other
| |
| Gasteropoda. Jour. Acad. Nat. Sci. Philadelphia, xii.
| |
| CONKLIN, E. G. (1903).
| |
| | |
| The Cause of Inverse Symmetry. Anal. Anz., xxiii.
| |
| CONKLIN, E. G. (1903).
| |
| | |
| The Earliest Differentiations of the Egg. Abstract of paper before Amer. Philos. Soc, Science, xvii.
| |
| CRAMPTON, H. E. (1894).
| |
| | |
| Reversal of Cleavage in a Sinistral Gasteropod. Ann. New York Acad. Sci., viii.
| |
| CRAMPTON, H. E. (1896).
| |
| | |
| Experimental Studies on Gasteropod Development. Arch. Entiv. Mech., iii.
| |
| CRAMPTON, H. E. (1S97).
| |
| | |
| The Ascidian Half- Embryo. Ann. Nezv York Acad. Sci., x.
| |
| CRAMPTON, H. E. (1899).
| |
| | |
| Studies upon the Early History of the Ascidian Egg. Jour. Morph., xv. Supplement.
| |
| DAYIDOFF, M.v. (1889-1891).
| |
| | |
| Untersuchungen zur Entwicklungsgeschichte der Distaplia magnalarva, Delia Yalle. Mitth. Zool.
| |
| Stat. Neapel, ix.
| |
| | |
| DRIESCH, H. (1895).
| |
| | |
| Von der Entwicklung einzelner Ascidienblastomeren. Arch. Entw. Mech., i.
| |
| DRIESCH, H. (1896).
| |
| | |
| Betrachtungen iiber die Organization des Eies und ihre Genese. Arch. Eniw. Mech., iv.
| |
| DRIESCH, H. (1903).
| |
| | |
| Ueber Aenderung der Regulationsfahigkeiten im Verlauf der Entwicklung bei Ascidien. Arch. Entw.
| |
| Mech., xvii.
| |
| FISCHEL, A. (1897, 1898).
| |
| | |
| Experimentelle Untersuchungen am Ctenophorenei. Arch. Entw. Mech., vi, vii.
| |
| FISCHEL, A. (1903).
| |
| | |
| Entwicklung und Organ-Differenzirung. Arch. Entw. Mech., xv.
| |
| FLODERUS, M. (1896).
| |
| | |
| Ueber die Bildung der Follikelhiillen bei den Ascidien. Zeit. wiss. Zool., lxi.
| |
| GARBOWSKI, T. (1S98).
| |
| | |
| Amphioxus als Grundlage der Mesodermtheorie. Anat. Anz., xiv.
| |
| GOLSKI, St. (1899)'.
| |
| | |
| La Maturation et la fecondation de 1'oeuf de Ciona intestinalis F. Bull. Internat. Acad. Sci. Cracovie.
| |
| HACKER, V. (1899).
| |
| | |
| Praxis und Theorie der Zellen und Befruchtungslehre. Jena.
| |
| HATSCHEK, B. (1881).
| |
| | |
| Studien iiber die Entwicklung der Amphioxus. Arb. Zool. /fist, ll'ien, iv.
| |
| HERTWIG, O. (1892).
| |
| | |
| Urmund und Spina Bifida. Arch. mik. Anat., xxxix.
| |
| HERTWIG, R. (1904).
| |
| | |
| Ueber physiologiscbe Degeneration bei Actinosphaerium Eichorni. Fcstschr. zum siebzigsten Geburtstage v. E. Haeckel.
| |
| HILL, M. D. (1895).
| |
| | |
| Notes on the Fecundation of the Egg of Sphaerechinus granulans and on the Maturation and Fertilization of the Egg of Phallusia mammalata. Quart. Journ. Mic. Sci., xxxviii.
| |
| JORDAN, E. O. (1893).
| |
| | |
| The Habits and Development of the Newt. Jour. Morph., viii.
| |
| JULIN, CH. (1893).
| |
| | |
| Ovog^nfese, spermatogn<;se et fecondation chez Styelopsis. Bull. Sci. France et Belg., xxv.
| |
| KLAATSCH, H. (1896).
| |
| | |
| Bemerkungen iiber die Gastrula des Amphioxus. Morph. Ja/ub., xxv.
| |
| KOFOID, C. A. (1S94).
| |
| | |
| On Some Laws of Cleavage in Limax. Proc. Amer. Acad. Arts and Sci., xxix.
| |
| KOPSCH, Fr. (1900).
| |
| | |
| Ueber das Verhaltniss der embryonalen Achsen zu den drei ersten Furchungsebenen beim Frosch.
| |
| Internat. Mona/schr. f. Anat. u. Physiol, xvii.
| |
| KORSCHELT, u. HEIDER (1903).
| |
| | |
| Lehrbuch der vergleich. Entwicklungsgeschichte der wirbellosen Thiere. Allgemeiner Theil, 2te Lieferung. Jena.
| |
| KOVVALEVSKY, A. (1866)
| |
| Entwicklungsgeschichte der einfachen Ascidien. Mem. V Acad. St. Petersbourg, Ser. 7, x.
| |
| KOVVALEVSKY^ A. (1867).
| |
| | |
| Entwicklungsgeschichte der Amphioxus lanceolatus. Idem, xi.
| |
| KOWALEVSKY, A. (1871).
| |
| | |
| Weitere Studien iiber die Entwicklung der einfachen Ascidien. Arch. mik. Anat, vii.
| |
| KUPFFER, C. v. (1870).
| |
| | |
| Die Stammverwandschaft zwischen Ascidien und VYirbelthieren. Arch. mik. Aval., vi.
| |
| LILLIE, F. R. (1895).
| |
| | |
| The Embryology of the Unionidte. Jour. Morph., x.
| |
| LILLIE, F. R. (1899).
| |
| | |
| Adaptations in Cleavage. Woods Holt Biol. Led., 1898.
| |
| LILLIE, F. R. (1901).
| |
| | |
| The Organization of the Egg of Unio, etc. Jour. Morph., xvii.
| |
| LILLIE, F. R. (1902).
| |
| | |
| Differentiation without Cleavage in the Egg of the Annelid Chaetopterus pergamentaceus. Arch.
| |
| Entw. Mech., xiv.
| |
| | |
| | |
| LWOFF, B. (1894).
| |
| | |
| Die Bildung der primaren Keimblatter und die Entstehung des Chorda und des Mesoderms bei den
| |
| W'irbelthieren. Bull. Soc. Imp. Nat. d. Moscou, N. S., viii.
| |
| LYON, E. P. (1903).
| |
| | |
| Experiments on Artificial Parthenogenesis. Amer. Jour. Physiol., ix.
| |
| MAC BRIDE, E. \V. (1898).
| |
| | |
| The Early Development of Amphioxus. Quart. Jour. Mic. Sei., xl.
| |
| MORGAN, T. H. (1S90).
| |
| | |
| The Origin of the Test Cells of Ascidians. Journ. Morph., iv.
| |
| MORGAN, T. H. (1901).
| |
| | |
| Regeneration. New York.
| |
| MORGAN, T. H. (1903).
| |
| | |
| Evolution and Adaptation. New York.
| |
| MORGAN, T. H. (1904).
| |
| | |
| Self Fertilization Induced by Artificial Means. Jour. Exp. Zool., i.
| |
| MORGAN and HAZEN (1900).
| |
| | |
| The Gastrulation of Amphioxus. Jour. Morph., xvi.
| |
| MOSKOWSKI, M. (1902).
| |
| | |
| Ueber den Einfluss der Schwerkraft auf die Entstehung und Erhaltung der bilateralen Symmetric des
| |
| Froscheies. Arch. mik. Anat. Ix.
| |
| MOTTIER, D. M. (1897).
| |
| | |
| Beitriige zur Kenntniss der Kerntheilung in den Pollenmutterzellen. Jahrb. wiss. Bol., xxx.
| |
| NEMEC, B. (.1S99).
| |
| | |
| Ueber die karyokinetische Kerntheilung in den Wiirzelspitzen von Allium. Idem, xxxiii.
| |
| OSTERHOUT. \V. J. V. (1897).
| |
| | |
| Ueber Entstehung der karyokinetischen Spindel bei Equisetum. Idem, xxx.
| |
| PFLUGER, E. (18S3).
| |
| | |
| Ueber den Einfluss der Schwerkraft auf die Theilung der Zellen. Pfluger's Archiv., xxxi, xxxii.
| |
| RABL, C.'(1892, 1S96).
| |
| | |
| Theorie des Mesoderms. Morph. Jahrb., xv, xix, xxiv.
| |
| ROUX, W. (1883).
| |
| | |
| Ueber die Zeit der Bestimmung der Hauptrichtungen des Froschembryo. Gesammelte Werke, ii.
| |
| ROUX, W. (1885).
| |
| | |
| Ueber die Bestimmung der Hauptrichtungen des Froschembryo im Ei und iiber die erste Theilung des
| |
| Froscheies, Idem.
| |
| ROUX, W. (1887).
| |
| | |
| Die Bestimmung der Medianebene des Froschembryo d.urch die Copulationsrichtung des Eikerns und
| |
| des Spermakerns. Idem.
| |
| ROUX, \Y. (1S92).
| |
| | |
| Ueber das Entwicklungsmechanische Vermogen jeder der beiden ersten Furchungszellen des Eies.
| |
| Verh. d. Anal. Ges. zu Wien, 1892.
| |
| ROUX, W. (1895).
| |
| | |
| Einleitung. Arch. Entw. Mech., i.
| |
| ROUX, W. (1903).
| |
| | |
| Ueber die Ursachen der Bestimmung der Hauptrichtungen des Embryo im Froschei. Anat. Am., xxiii.
| |
| SAMASSA, P. (1894).
| |
| | |
| Zur Kenntniss der Furchung bei den Ascidien. Arch. mik. Anat. xliv.
| |
| SAMASSA, P. (1898).
| |
| | |
| Studien iiber den Einfluss des Dotters auf die Gastrulation und die Bildung der primaren Keimblatter
| |
| der W'irbelthiere. IY Amphioxus. Arch. Entw. Mech., vii.
| |
| SEELIGER, O. (1882
| |
| | |
| Zur Entuicklungsgeschichte der Ascidien. Sitzb. Akad. Wiss. Mien., Ixxxv.
| |
| SEELIGER, O. (1885
| |
| | |
| Die Entwicklungsgeschichte der Socialen Ascidien. Jena. Zeitschr., xviii.
| |
| SOBOTTA, J. (1S97).
| |
| | |
| Die Reifung und Befruchtung des Eies von Amphioxus. Arch. mik. Ana/., 1.
| |
| SOBOTTA. J. (1897 .
| |
| | |
| Beobachtungen iiber den Gastrulationsvorgang beim Amphioxus. Verh. Phys. Med. Ges. II Urzburg, xxxi.
| |
| VEJDOVSKY, Fr. (1888-1892).
| |
| | |
| Entwicklungsgeschichtliche Untersuchungen. Pi
| |
| | |
| | |
| WHEELER, W. M. (1897).
| |
| | |
| The Maturation, Fecundation and Early Cleavage in Myzostoma. Arch, dc Biol., xv.
| |
| WHITMAN, C. O. (1893).
| |
| | |
| The Inadequacy of the Cell Theory of Development. Woods Holl Biol. Lectures, 1893.
| |
| WILSON, E. B. (1892).
| |
| | |
| The Cell-Lineage of Nereis. Jour. Morph., vi.
| |
| WILSON, E. B. (1893).
| |
| | |
| Amphioxus and the Mosaic Theory of Development. Jour. Morph., viii.
| |
| WILSON, E. B. (1903).
| |
| | |
| Experiments on Cleavage and Localization in the Nemertine Egg. Arch. Entw. Meek., xvi.
| |
| WILSON, E. B. (1904).
| |
| | |
| Experimental Studies on Germinal Localization. Jour. Exp. Zoo/., i.
| |
| WILSON and MATHEWS (1895).
| |
| | |
| Maturation, Fertilzation and Polarity in the Echinoderm Egg. Jour. Morph., x.
| |
| WILSON, H. Y. (1900).
| |
| | |
| Formation of the Blastopore in the Frog's Egg. Anat. A?i~., xviii.
| |
| YATSU, N. (1904).
| |
| | |
| Experiments on the Development of Egg Fragments in Cerebratulus. Biol. Bull., vi.
| |
| ZIEGLER, H. E. (1902).
| |
| | |
| Lehrbuch der Vergleichenden Entwicklung^geschichte der niedereu Wirbelthiere. Jena.
| |
| ZUR STRASSEN, O. (1896).
| |
| | |
| Embryonalentwicklung der Ascaris megalocephala. Arch. Entw. Mcch , iii
| |
| | |
| | |
| | |
| ==Explanation Of Figures==
| |
| | |
| All figures represented in the plates were drawn with the aid of the camera lucida. Plates I-V
| |
| represent the living eggs of Oynthia (Styela) partita, and the colors used are in no sense diagrammatic
| |
| but are as nearly as possible those which appear in life; plate VI-X show fixed and stained' eggs of the
| |
| same species; plates XI anil XII, lixed and stained eggs of Citmn hitextintdk. The figures of the
| |
| living eggs of Cynthia i plates I-V) were drawn at the stage level under Zeiss Apochromatic Obj. 4
| |
| mm., Oc. 4. and are therefore magnified 250 diameters. With the exception of figs. 46-49 all the
| |
| details shown in these drawings were observed in living eggs without reference to fixed and stained
| |
| material; in the figures specified certain cell outlines were added to the drawings of the living eggs
| |
| after a study of prepared material. The figures of plates VI-XII, with the exception of figs. 61-75,
| |
| were drawn at the stage level under Zeiss Apochromatic Obj. 3 mm., Oc. 4; in the process of reproduction they have been reduced about one-fifth so that as they now appear they represent a magnification of about 266 diameters. Figs. 61-75, plate VI, were drawn under Zeiss Apochromatic Obj. 1.5
| |
| nun., Oc. 4, and therefore represent a magnification, after reduction, of about 535 diameters. The
| |
| chorion and test cells, though present in most of the eggs drawn, have been omitted from all the
| |
| figures, except those of plates I, VI and VII; the chorion, without the test cells, is shown in plates
| |
| II-V.
| |
| | |
| | |
| | |
| A.
| |
| a.p.bp,
| |
| ch.cn.c. p.Cr.
| |
| Cr. s,
| |
| | |
| D.
| |
| end.
| |
| f. c
| |
| g. v.kp.
| |
| mb.
| |
| m'ch.
| |
| ms.
| |
| n.
| |
| n. p.
| |
| | |
| | |
| Reference
| |
| | |
| -Anterior.
| |
| | |
| -Animal pole.
| |
| | |
| -Blastopore.
| |
| | |
| -Chorda
| |
| | |
| -Chorion.
| |
| | |
| -Clear protoplasm.
| |
| | |
| -Crescent of mesodermal substance (yellow
| |
| | |
| in Cynthia).
| |
| -Substance of crescent.
| |
| -Dorsal.
| |
| -Endoderm.
| |
| | |
| -Follicle cells (outside of chorion).
| |
| -Germinal vesicle.
| |
| -Karyoplasm (achromatic substance from
| |
| | |
| nucleus).
| |
| -Thick membrane which becomes chorion.
| |
| -Mesenchyme.
| |
| -Muscle cells.
| |
| -Nucleus.
| |
| -Neural plate.
| |
| | |
| | |
| | |
| Letters.
| |
| | |
| nl. Nucleolus.
| |
| n. t. Neural tube.
| |
| | |
| P. Posterior,
| |
| p. b. Polar body.
| |
| | |
| 1 p. b. First bolar body.
| |
| | |
| 2 p. b. Second polar body.
| |
| | |
| p. 1. Peripheral layer of protoplasm.
| |
| | |
| 1 P. S. First polar spindle.
| |
| | |
| 2 P. S. Second polar spindle.
| |
| | |
| Sn . Sperm atozoon .
| |
| t. c Test cells.
| |
| V. Ventral,
| |
| v. end. Ventral (caudal) endoderm.
| |
| v. p. Vegetal pole. y. h. Yellow hemisphere of egg.
| |
| | |
| ' yk. Yolk.
| |
| | |
| z Zwischenkorper (mid-body).
| |
| | |
| n. Sperm nucleus.
| |
| | |
| | |
| | |
| Nomenclature of Cells.
| |
| | |
| A Anterior half of egg; B Posterior half ; AB Right half; AB Left half ; A and A Right
| |
| and left anterior quadrants; B and B Right and left posterior quadrants. From the 8-cell stage
| |
| onward the cells of the eudodermal hemisphere are designated by capital letters, those of the ectodermal hemisphere by lower case. The first exponent indicates the cell generation, counting the unsegmented egg as the first ; the second exponent the position of the cell relative to the animal and vegetal
| |
| poles, cells which lie nearest the vegetal pole having the lowest exponent, those farthest away (nearest
| |
| the animal pole) having the highest. See p. 37.
| |
| | |
| | |
| | |
| Plate I.
| |
| Figures of the Living Egg* of Oynthia partita; Maturation and Fertilization.
| |
| | |
| Fig. 1. Unfertilized egg before the fading of the germinal vehicle, showing central mass of gray yolk,
| |
| peripheral layer of yellow protoplasm, test cells and chorion.
| |
| | |
| Fig. 2. .Similar egg after the disappearance of the nuclear membrane, showing the spreading of the
| |
| clear protoplasm of the germinal vesicle at the animal pole.
| |
| | |
| Fig. 3. Another egg about five minutes after fertilization, showing the streaming of the peripheral
| |
| protoplasm to the lower pole where the spermatozoon enters, thus exposing the gray yolk
| |
| of the upper hemisphere; the test cells are also carried by this streaming to the lower
| |
| hemisphere.
| |
| | |
| Figs. 4 and 5. Other eggs -bowing successive stages in the collection of the yellow and clear protoplasm at the vegetal pole; clear protoplasm lies beneath and extends a short distance
| |
| beyond the edge of the yellow cap.
| |
| | |
| Figs. 6-10. Successive stages of the same egg drawn at intervals of about five minutes; viewed from
| |
| the vegetal pole. In fig. 6 the area of yellow protoplasm is smallest, and the sperm
| |
| nucleus is a small clear area near its center. Figs. 7.-10 show stages in the spreading of
| |
| this yellow protoplasm until it covers nearly the whole of the lower hemisphere; at the
| |
| same time the sperm nucleus and aster move toward one side of the yellow cap and the
| |
| yellow protoplasm begins to collect into a crescent at this side.
| |
| | |
| Fig. 11. Side view of an egg of about the same stage as fig. 10, showing the eccentric position of the
| |
| sperm nucleus and a small area of clear protoplasm at the upper pole where the polar
| |
| bodies are being formed.
| |
| | |
| Fit;-. 12. Polyspermic (?) egg, viewed from the vegetal pole, showing four collections of yellow protoplasm around as many sperm (?) nuclei (see p. 24).
| |
| | |
| | |
| | |
| | |
| PLATE 1.
| |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| Plate II.
| |
| | |
| Living Eggs of Cynthia partita ; First Cleavage.
| |
| | |
| Figs. 13 and 14. Side views of egg, showing the formation of the crescent (cr.) from the yellow hemisphere ; in all the figures the animal pole is above, the vegetal pole below. Above the
| |
| yellow crescent is an area of clear protoplasm (c. p.).
| |
| | |
| Fig. 15. Similar stage of another egg showing the aggregation of test cells over the crescent and the
| |
| protrusion of the chorion at this place, an unusual phenomenon.
| |
| | |
| Fig. 16. Egg showing crescent, viewed nearly from the vegetal pule
| |
| | |
| Fig. 17. Another egg showing crescent and clear protoplasm, viewed from posterior pole; the animal
| |
| pole is above, the vegetal below.
| |
| | |
| Figs. 18, 19, 20. Successive stages of the same egg drawn at intervals of about two minutes, showing
| |
| the approaching division of the egg.
| |
| | |
| Figs. 21, 22, 23, 24. Succesive stages in the first cleavage of an egg, drawn at intervals of about two
| |
| or three minutes. The egg is viewed from the posterior pole and shows the complicated
| |
| forms taken by the yellow crescent during the division, also the enlargement of the area
| |
| of clear protoplasm and its extension toward the animal pole.
| |
| | |
| | |
| | |
| | |
| Plate III.
| |
| | |
| Living Eggs of Oynthia partita; First to Fourth Cleavage.
| |
| | |
| Figs. 25 and 26- Same egg a* the one shown in figs. 21-24; final stages in the first cleavage.
| |
| | |
| Fig. 27. Another egg at the close of the first cleavage ; seen from the posterior pole.
| |
| | |
| Fig. 28. End view of egg of same stage as preceding, showing the lateral limits of the yellow crescent, the clear protoplasm in the upper hemisphere and the yolk in the lower. The
| |
| anterior portion of the lower hemisphere is composed of light gray material ; this is the
| |
| gray crescent and gives rise to chorda and neural plate.
| |
| | |
| Fig. 2'.). Four-cell stage, viewed from the animal pole.
| |
| | |
| Fig. 30. Similar egg seen from the vegetal pole; the crescent covers about half of the posterior
| |
| blastomeres.
| |
| | |
| Fig. 31. Fight-cell stage: the crescent is limited entirely to the two posterior blastomeres at the vegetal pole : while under observation the furrow between B 4 ' and b 4-2 shifted from the position indicated by the faint line to that shown by the heavy line, thus giving rise to the
| |
| "cross furrow" shown in the next figure.
| |
| | |
| Fig. 32. Fight-cell stage, viewed from the right side, showing a small amount of yellow protoplasm
| |
| around all the nuclei.
| |
| | |
| Fig. 33. Same stage viewed from the vegetal pole, showing 1% yolk laden endoderm cells and the
| |
| crescent.
| |
| | |
| Fig. 34. Same stage viewed from the posterior-animal pdreTshowing the clear ectodermal cells and
| |
| the crescent.
| |
| | |
| Fig. 35. Same stage seen from the anterior-vegetal pole; yellow protoplasm around all the nuclei.
| |
| | |
| Fig. 36. -Fourth cleavage of the egg seen from the vegetal pole.
| |
| | |
| | |
| | |
| | |
| Plate IV.
| |
| | |
| Living Eggs of Cynthia partita ; Fourth Cleavage to Gastrula.
| |
| | |
| Fig. 37. Sixteeu-cell stage viewed from vegetal pole.
| |
| | |
| Fig. 38. Sixteen-cell stage, from the animal pole, yellow protoplasm around the nuclei.
| |
| | |
| Fig. 39. Twenty-two cell stage, from the vegetal pole; four mesoderm cells (yellow), ten endoderm,
| |
| | |
| chorda and neural plate cells (gray) and eight ectoderm cells (clear).
| |
| Fig. 40. Same stage viewed from the posterior pole.
| |
| | |
| Fig. 41. Egg passing into the 32-cell stage; postero-dorsal (vegetal pole) view.
| |
| Fig. 42. Thirty-two-cell stage, postero-dorsal view.
| |
| Fig. 43. Forty-four- cell stage; posterior view, showing separation of mesenchyme (m'ch) from muscle
| |
| | |
| cells (ms.).
| |
| Fig. 44. Same stage, dorsal view, showing subdivision of endoderm cells.
| |
| Fig. 45. Similar stage, posterior view, showing separation of another mesenchyme cell from a muscle
| |
| | |
| cell.
| |
| Fig. 4(j. Seventy-four cell stage, dorsal view, showing division of 4 chorda and 4 neural piatc cells;
| |
| | |
| there are 10 mesenchyme and 6 muscle cells, besides 10 endoderm cells.
| |
| Fig. 47. One hundred and sixteen cell-stage, showing the beginning of gastrulation, also the neural
| |
| | |
| plate, chorda, muscle and mesenchyme cells.
| |
| Fig. 48. Slightly older stage showing advancing gastrulation with inrolling of cells at edge of blastopore.
| |
| | |
| | |
| | |
| | |
| Plate V.
| |
| | |
| Living Embryo* of Oynthia partita; Gastrula to Tadpole.
| |
| | |
| Fig. 49. Gastrula showing neural plate, chorda and muscle cells; about 176 cells, 96 ectoderm, 32
| |
| | |
| mesoderm, 20 endoderm, 8 chorda, 8 dorsal neural plate and 12 ventral neural plate cells.
| |
| Figs. 50 and 51. Two stages of one embryo, the second drawn about ten minutes after the first; the
| |
| | |
| yellow cells in the mid-line (m'ch.) are mesenchyme cells, the others (ms. ) muscle cells.
| |
| Fig. 52. Posterior view of elongated gastrula, the blastopore reduced to a narrow slit.
| |
| Fig. 53. Dorsal view of similar stage, the blastopore a small opening at the posterior end of a groove
| |
| Fig. 54. An older embryo, the blastopore covered by the forward growth of the posterior lips.
| |
| Fig. 55. Embryo with inrolling neural plate and with muscle cells arranged in three rows.
| |
| Fig. 56. Embryo of about the same stage as that shown in figs. 52 and 53; seen from the left side
| |
| | |
| showing neural groove, three rows of muscle cells which run transversely to the long
| |
| | |
| axis, and a row of mesenchyme cells in the long axis.
| |
| Fig. 57. Embryo viewed from posterior end, showing blastopore-raphe with open groove above and
| |
| | |
| with three rows of muscle cells on each side, also a row of mesenchyme cells.
| |
| Fig. 58. Young tadpole seen from dorsal side, neural groove open in front and closed behind, smallcelled mesenchyme in front of large muscle cells.
| |
| Fig. 59. Same stage as preceding seen from the right side, showing neural groove, mesenchyme and
| |
| | |
| three rows of muscle, cells.
| |
| Fig. 60. Tadpole of slightly older stage viewed from the posterior (caudal) end showing on each
| |
| | |
| side three large muscle cells each of which belongs to a row of such cells (<;/'. tig. 59).
| |
| | |
| These muscle cells are connected across the mid-line at the posterior end by a few
| |
| | |
| small mesenchyme cells.
| |
| | |
| | |
| | |
| PLATE VI.
| |
| | |
| SeHitiits of Eggs of Cynthia partita; Maturation and Fertilization.
| |
| Figs. 617S5 magnified 5S5 diameters; Figs. 70-87 magnified :.>t!<! diameters.
| |
| | |
| Fig. 61. Ovarian egg fully formed, showing germinal vesicle surrounded by yolk, and peripheral
| |
| | |
| layer of protoplasm containing test cells and yellow granules (small spheres in figure).
| |
| Fig. 62. Free egg shortly after the dissolution of the nuclear membrane, showing in the middle of the
| |
| | |
| clear karyoplasm fragments of nucleolus, chromosomes and a granular mass from which
| |
| | |
| spindle fibres arise; the peripheral protoplasm contains yellow granules.
| |
| Fig. 63. Egg similar to the preceding, but with the spindle fibres more fully formed.
| |
| Fig. 64. Similar to preceding, spindle fibres radiate in all directions.
| |
| | |
| Fig. 65. The first polar spindle lies near the surface of the egg and its fibres are approximately paratangential ; the peripheral layer of protoplasm has streamed away from the animal pole
| |
| | |
| and the karyoplasm from the germinal vesicle has spread out here in a broad disk.
| |
| Fig. 66. Metaphase of first polar spindle which is nearly parallel with surface; no centrosomes present.
| |
| Fig. 67. Anaphase of first polar spindle which is turning into a radial position.
| |
| Fig. OS. Separation of first polar body.
| |
| | |
| Fig. 69. Metaphase of second polar spindle, which is paratangential in position.
| |
| Fig. 70. Anaphase of second polar spindle.
| |
| Fig. 71. Second polar spindle approaching a. radial position.
| |
| Fig. 72. Separation of second polar body.
| |
| | |
| Fig. 73. Fusion of chromosomal vesicles in egg to form egg nucleus.
| |
| Fig. 74. Vegetal pole of egg of the stage shown in figs. 65 and 79, showing the entrance of the sperm
| |
| | |
| into the egg and the collection of yellow granules around the sperm head.
| |
| Fig. 75. Later stage in the entrance of the sperm ; formation of sperm aster from the middle- piece.
| |
| Fig. 76. Free egg before the solution of the nuclear membrane but after the extrusion of the tesl
| |
| | |
| cells; the chromosomes at the periphery of the germinal vesicle.
| |
| Fig. 77. Egg after being laid but before fertilization ; chromosomes and granular substance which
| |
| | |
| forms spindle fibres in the center of the karyoplasm. The egg remains in this condition
| |
| | |
| until fertilized.
| |
| Fig. 78. Same as preceding, save that spindle fibres are forming and karyoplasm has moved nearer to
| |
| | |
| the animal pole.
| |
| Fig. 79. Egg showing the entrance of the spermatozoon near the vegetal pole and the spreading of
| |
| | |
| the karyoplasm into a thin cap at the animal pole.
| |
| Fig. 80. Slightly more advanced stage showing development of sperm aster and collection of yellow
| |
| | |
| granules at vegetal pole, spermatozoa have entered some of the test cells.
| |
| Fig. 81. First polar spindle assuming a radial position ; increase of cytoplasmic area surrounding the
| |
| | |
| sperm nucleus and aster, the latter are moving across the egg axis and hence in the
| |
| | |
| longest path toward the equator.
| |
| Fig. 82. Stage slightly more advanced than the preceding; sperm nucleus, aster, clear and yellow
| |
| | |
| protoplasm becoming eccentric toward the posterior side.
| |
| Fig. 83. First polar body formed ; prophase of second polar spindle.
| |
| Fig. 84. Metaphase of second polar spindle; yellow protoplasm collecting into crescent.
| |
| Fig. 85. Anaphase of second polar spindle, spermatozoa in some of the test cells.
| |
| Fig. 86. Telophase of second polar spindle.
| |
| Fig. 87. Movement of sperm nucleus and aster and of surrounding protoplasm to the posterior side
| |
| | |
| of the egg; approach of the germ nuclei.
| |
| | |
| | |
| | |
| PLATE VII.
| |
| | |
| Sections of Eggs of Cynthia partita; Fertilization and Early Cleavages.
| |
| | |
| Fig. 88. Division of sperm aster ; crescent substance at periphery on posterior side.
| |
| | |
| Fig. 89. Similar to preceding but showing egg nucleus.
| |
| | |
| Figs. 90 and 91. Sections at right angles to first cleavage plane but oblique to egg axis, showing the
| |
| union of pronuclei; clear and yellow protoplasm and sperm amphiaster in posterior half
| |
| of egg.
| |
| | |
| Fig. 92. Section in the plane of the first cleavage (future median plane) showing the union of the
| |
| germ nuclei, the clear protoplasm and crescent on the posterior side of the egg, and the
| |
| polar bodies above.
| |
| | |
| Fig. 93. Early prophase of the first cleavage, in the plane of the spindle axis.
| |
| | |
| Fig. 94 and 95. Two dispermic eggs; the first showing two sperm nuclei on the posterior side of the
| |
| egg with a single crescent ; the second, two sperm nuclei and one egg nucleus, with the
| |
| clear protoplasm and crescent about equally divided, by a tongue of yolk, between the
| |
| two sperm nuclei.
| |
| | |
| Fig. 96. Section in the plane of the first cleavage and transverse to the spindle, showing a cross section of the equatorial plate near the middle of the egg, surrounded by clear protoplasm,
| |
| also the crescent at the posterior border with some of the yellow spherules all around
| |
| the spindle.
| |
| | |
| Fig. 97. Metaphase of the first cleavage; equatorial section at right angles to the preceding.
| |
| | |
| Figs. 98 and 99. Early and late anaphases of the first cleavage; sections in equatorial plane.
| |
| | |
| Fig. 99. Constriction of cell body; chromosomal vesicles at the ends of the nuclear spindle. In both
| |
| | |
| figures the daughter centrosi is are elongating in the equatorial plane and at right
| |
| | |
| angles to the first spindle axis.
| |
| | |
| Fig. 100. Telophase of the first cleavage, showing the rotation of nuclei, eentrosomes and cytoplasm
| |
| toward the animal pole of the egg.
| |
| | |
| Fig. 101. Equatorial section, prophase of second cleavage; crescent substance at the posterior pole.
| |
| | |
| Fig. 102. Section at right angles to the preceding and through the axis of one of the spindles; nucleus
| |
| and cytoplasm lie above the equator, yolk and crescent below.
| |
| | |
| Fig. 103. Equatorial section showing precocious division of the eentrosomes in one of the blastomeres.
| |
| | |
| Fig. 104. Anaphase of second cleavage; equatorial section ; chromosomal vesicles at ends of nuclear
| |
| spindles.
| |
| | |
| Fig. 105. Telophase of second cleavage ; equatorial section, showing the bending of the middle of the
| |
| spindle toward the center of the egg, and a large amount of clear cytoplasm aud of
| |
| crescent substance in the posterior quadrants.
| |
| | |
| Figs. 106 and 107. Two sections through one and the same egg in the eight-cell stage ; the first
| |
| through the anterior blastomeres ; the second through the posterior. The cytoplasm is
| |
| most abundant in the cells at the animal pole, the crescent substance in the two posteriorvegetal cells, though it is also found around all the nuclei. The polar bodies, which are
| |
| shown in stippled outlines, are not in the plane of either of these sections, but in the
| |
| region between them.
| |
| | |
| | |
| | |
| PLATE VIII.
| |
| | |
| Surface Views of Entire Eggs of Cynthia partita; Eight to Forty-four Cells.
| |
| | |
| Fig. 108. Eight-cell stage ; left side of egg ; showing spindles of third cleavage.
| |
| | |
| Fig. 109. Anterior view of 8-cell stage, showing cytoplasm most abundant in the animal pole cells,
| |
| | |
| and the yolk largely collected in the anterior cell of the vegetal hemisphere.
| |
| Figs. 110, 111,112. Stages in the fourth cleavage; figs. 110 and 112 viewed from the animal pole, fig.
| |
| | |
| Ill from the. vegetal pole.
| |
| Fig. 113. Telophase of fourth cleavage, vegetal poje view ; caps of deeply staining protoplasm lie at
| |
| | |
| the hinder borders of the small posterior cells (B 5,2 ).
| |
| Figs. 114 and 115. Anterior and posterior views of the 16-cell -stage; fig. 115 showing caps of
| |
| | |
| deeply staining protoplasm at the posterior pole, which later go into the posterior mesenchyme cells (B' 6 , figs. 130, 131).
| |
| Figs, lit! and 117. Ventral and dorsal views of a 20-cell stage, showing the cells at the vegetal pole
| |
| | |
| dividing before those at the animal pole.
| |
| Fig. 118. Slightly older stage with some of the animal pole cells dividing.
| |
| Figs. 119-123. Five views of one and the same egg; fig. 119, ventral; 120, dorsal; 121, anterior;
| |
| | |
| 122, posterior; 123, right side; the latter shows in dotted outlines the great elongation
| |
| | |
| of the cells at the animal pole and the flattened shape of the cells at the vegetal pole ;
| |
| | |
| all the designations of cells in tig. 123 should be underscored ; 44 cells, 16 ectoderm, 10
| |
| | |
| endoderm, 10 mesoderm, 4 chorda and 4 neural plate cells.
| |
| Figs. 124-129. Six different views of one and the same egg in the 44-cell stage showing the divisions
| |
| | |
| of the ectodermal cells and the second cells of the crescent (B 6 ' 4 ) ; when these divisions
| |
| | |
| are completed there will be 62 cells. Fig. 124, ventral; 125, dorsal; 126, anterior; 127,
| |
| | |
| postero-dorsal.
| |
| | |
| | |
| | |
| | |
| | |
| Plate IX.
| |
| | |
| Surfact Views oj Entire Eggs of Cynthia partita; Forty-four to Tiro Hundred and Eighteen Cells,
| |
| | |
| Figs. 128, 129. Same egg as shown in figs. 124-127. Fig. 128 from the left side; the equator of the
| |
| egg (plane of the third cleavage) is the heavy line running between A ami 1' and
| |
| separating cells designated by lower case from those designated by capital letters. Fig.
| |
| 129. View from the posterior pole.
| |
| | |
| Figs. 130 and 131. Ventral and dorsal views respectively of one and the same egg; 64 cells, 32 in
| |
| each hemisphere, distributed as follows: Ventral hemisphere, 26 ectoderm, 6 neural plate
| |
| cells (a 7 -', a 7 -', a 7- ' 3 ) ; Dorsal hemisphere, 10 endoderra, 4 chorda (A 73 , A 77 ), 4 neural plate
| |
| (A'-, A 7 - 8 ), 10 mesenchyme (B 7 - 6 , B 7 5 , B", B 73 , A 7 ' 6 ) and 4 muscle (B 78 , B 7 ->).
| |
| | |
| Fig. 132. Postero-dorsal view of an egg in the same stage as the preceding showing direction of
| |
| division of mesenchyme cell (B 73 ).
| |
| | |
| Figs. 133 and 134. Ventral and dorsal views respectively of one and the same egg; 110 cells; Ventral hemisphere 64 cells, 52 ectoderm, 12 neural plate (a 8 ' 9 , a 8= , a 8 ' 7 , a 8 ' 8 , a 8,25 , a 8=6 ) ;
| |
| Dorsal hemisphere 46 cells, 10 endoderm, 8 chorda (A 85 , A 86 , A 8 - 13 , A 8 ' 4 ), 8 neural plate
| |
| (A 8 - 7 , A 8 - 8 , A 8 -' 3 , A 8 ' 6 ), 12 mesenchyme (B 7 - 6 , B", B 7 7 , B 8 ', B 86 , A 7 - 6 ), 8 muscle (B 8 - 7 , B 88 ,
| |
| B 8 ' 5 , B 8 " 6 ). Gastrulation has begun.
| |
| | |
| Fig. 135. Dorsal view of a slightly more advanced stage showing increasing gastrulation ; 118 cells,
| |
| ventral hemisphere 64 cells, dorsal hemisphere 54, viz., 10 endoderm, 16 mesenchyme, 12
| |
| muscle, 8 chorda, 8 neural plate; when the divisions indicated by spindles are completed
| |
| there will 4 additional endoderm and 2 additional mesenchyme cells.
| |
| | |
| Figs. 136-139. Four views of one and the same egg; fig. 136 dorsal, 137 posterior, 138 anterior, 139
| |
| ventral; gastrulation well advanced. 124 cells; Ventral hemisphere 64 cells, 52 ectoderm, 12 neural plate (a 8 -'", a 8 -*", a 8 " 7 , a 8 ' 8 , a 8 - 23 , a 6 - 56 ); Dorsal hemisphere 60 cells, 14
| |
| endoderm, 8 chorda, 8 neural plate, 18 mesenchyme, 12 muscle. Spindles are already
| |
| present for divisions, which, when completed, will lead to 178 cells, viz., 96 ectoderm, 12
| |
| neural plate of ventral hemisphere, 12 neural plate of dorsal hemisphere, 8 chorda. 211
| |
| mesenchyme, 12 muscle, 18 endoderm.
| |
| | |
| Figs. 140-143. Four views of one and the same egg; 140 dorsal, 141 posterior, 142 anterior, 143
| |
| ventral. Gastrulation is here far advanced. 180 cells; Ventral hemisphere 108 cells,
| |
| 96 ectoderm, 12 neural plate; Dorsal hemisphere 72 cells, 12 neural plate of dorsal hemisphere, 8 chorda, 20 mesenchyme. 12 muscle, 20 endoderm; when the divisions indicated
| |
| by spindles are completed there will be 4 additional neural plate cells.
| |
| | |
| Figs. 144 and 145. Two views of the same egg; 144 dorsal, 145 median optical section in sagittal
| |
| plane. In fig. 145 the polar body is not visible, but its supposed position is indicated by
| |
| the dotted outline at the animal pole; the dotted outlines at the lower pole indicate the
| |
| mesoderm cells which lie in the lateral lip of the blastopore and out of the plane of the
| |
| section ; the rolling in of the muscle cells in the lateral lips is well shown.
| |
| | |
| Fig. 146. Anterior view of an egg of about the same stage as the preceding, showing the division of
| |
| the 12 neural plate cells of the ventral hemisphere.
| |
| | |
| Fig. 147. Ventral view of a similar stage with the ectoderm omitted in order to show the endoderm
| |
| and mesoderm from the ventral side. At this stag.' all the ectoderm cells have passed
| |
| into the 9th generation, all the endoderm into the 8th or 9th, all the mesoderm except
| |
| IV" into the 8th or 9th, all the chorda and neural plate cells into the 9th. There
| |
| are 218 cells; Ventral hemisphere 128 cells, 104 ectoderm, 24 neural plate of ventral
| |
| hemisphere; Dorsal hemisphere 90 cells, viz., 16 mural plate, 16 chorda, 20 mesenchyme,
| |
| 12 muscle, 26 endoderm.
| |
| | |
| | |
| Plate X.
| |
| Entire Embryos of Cynthia partita; Gastrula to Tadpole.
| |
| | |
| Muscle cells shaded by vertical lines, mesenchyme by transverse line*, nerve plate and tube by fine
| |
| | |
| stipples, chorda by coarse stipples.
| |
| | |
| Figs. 148-151. Four view? of the same embryo; fig. 14.8 dorsal, 149 ventral -posterior, 150 same view
| |
| but deeper level, showing mesoderm and endoderm, 151 right side. Many of the ectoderm cells are passing into the 10th generation.
| |
| | |
| Figs. 152-154. Three views of : embryo ; fig. 152 dorsal, showing closure of the blastopore from in
| |
| | |
| front, 153 same view but deeper level, showing cells beneath the ectoderm, 154 ventral
| |
| view of mesoderm and endoderm below ectoderm.
| |
| | |
| Figs. 155 and 156. Two ventral views of an advanced gastrula, the first showing the superficial ectoderm, the latter the endoderm and mesoderm lying beneath the ectoderm.
| |
| | |
| Fig. 157. Left side of embryo, showing, in optical section, muscle cells, mesenchyme, chorda and nerve
| |
| plate.
| |
| | |
| Figs. 158 and 159. Two optical sections of the same embryo, the former (158) through the open blastopore at the posterior end of the embryo, the latter (159) through the region anterior to
| |
| the blastopore.
| |
| | |
| Figs. 160-162. Three views of the same embryo from the ventral side but drawn at different levels;
| |
| 160 surface view showing ectoderm ; 161 same view deeper level, showing mesoderm and
| |
| endoderm : 162 same view still deeper level, showing chorda, mesoderm and endoderm.
| |
| | |
| Fig. 163. Dorsal view of embryo of same stage as preceding showing nerve plate and tube, mesoderm
| |
| and chorda.
| |
| | |
| Figs. 164 and 165. Two views of same embryo from right side ; fig. 164 median optical section showing chorda, nerve tube and endoderm ; 165 more superficial view showing muscle cells
| |
| and mesenchyme.
| |
| | |
| Fig. 166. Older embryo from right side, showing in optical section nerve tube, chorda, mesoderm and
| |
| endoderm.
| |
| | |
| Fig. 167. Young tadpole from left side showing in optical section nerve tube, chorda and mesoderm.
| |
| | |
| | |
| | |
| PLATE XL
| |
| | |
| Eggs of Ciona intestinalis ; Ovocyte to Fourth Cleavage.
| |
| | |
| Figs. 168-179 actual section*; figs. 180-187 whole eggs. Crescent substance and clear protoplasm
| |
| stippled. Polar bodies are actually present where drawn.
| |
| | |
| Fig. 168. Half grown ovarian egg, showing test cells within the egg cytoplasm, which is composed of
| |
| | |
| a layer of yolk surrounding the nucleus and a peripheral layer of clear protoplasm ;
| |
| | |
| around the whole egg is a layer of small follicle cells which later develop into the very
| |
| | |
| large conical cells which surround the chorion.
| |
| Fig. 169. Older ovarian egg, showing the area of the yolk increased and the test cells limited to the
| |
| | |
| peripheral layer of protoplasm, which is sharply differentiated from the yolk.
| |
| Fig. 170. Still older ovarian egg, showing the test cells arranged in groups or "nests" at the periphery
| |
| | |
| of the egg.
| |
| Fig. 171. Ripe ovarian egg ready to escape from the ovary, showing the extrusion of the test cells and
| |
| | |
| the formation of a thick homogeneous membrane around the egg, which ultimately
| |
| | |
| becomes the chorion.
| |
| Fig. 172. Free hut unfertilized egg which lias shrunken away from the chorion, showing the first
| |
| | |
| maturation spindle, the peripheral layer of protoplasm collected over the lower pole of
| |
| | |
| the egg and the achromatic substance of the germinal vesicle (kp) spread in a broad
| |
| | |
| layer over the upper pole and around the central yolk.
| |
| Fig. 173. Extrusion of first polar body and entrance of spermatozoon ; the sperm nucleus lies in the
| |
| | |
| peripheral layer surrounded by clear protoplasm in which astral rays are developing.
| |
| Fig. 174. Nearly equatorial section of the egg showing sperm nucleus and amphiaster at the posterior
| |
| | |
| side of the egg; at the surface is the granular protoplasm of the peripheral layer.
| |
| Fig. 175. Section of egg in plane of first cleavage (median plane), showing the approach of the germ
| |
| | |
| nuclei and the movement of the peripheral layer of protoplasm from the lower pole to
| |
| | |
| the posterior side to form the crescent ; in the darkly staining substance of the crescent
| |
| | |
| is a clear triangular area which corresponds to the clear area surrounding the sperm
| |
| | |
| nucleus in fig. 173.
| |
| Fig. 176. Section of slightly older stage in plane of first cleavage, showing the union of the germ
| |
| | |
| nuclei ; polar body out of the plane of section.
| |
| Fig. 177. Anaphase of first cleavage, showing the complete separateness of the nuclear and astral
| |
| | |
| portions of the mitotic figure; the crescent substance shows at the ends of the spindle.
| |
| Fig, 178. Telophase of first cleavage, showing " zwischenkorper " (z), also the bending of spindle axis
| |
| | |
| and shifting of cytoplasm and nuclei toward the animal pole.
| |
| Fig. 179. Anaphase of second cleavage, showing separateness of nuclear and astral portions of mitotic
| |
| | |
| figure, also position of crescent on postero- dorsal side ; polar body out of the plane of
| |
| | |
| section.
| |
| Fig. 180. Entire egg of Ciona, showing formation of second polar spindle, peripheral layer of protoplasm, sperm nucleus and aster.
| |
| Fig. 181. Anaphase of first cleavage viewed from posterior pole.
| |
| Fig. 182. Two-cell stage seen from posterior pole, cytoplasm and nuclei lie near animal pole, crescent
| |
| | |
| near vegetal,
| |
| fig. 183. Four-cell -stage seen from vegetal pole ; the crescent covers about one half the surface of the
| |
| | |
| two posterior blastomeres.
| |
| Fig. 1S4. Eight cell-stage, left side, showing cap of deeply staining protoplasm at posterior pole of
| |
| | |
| cells (B 4-1 ), which later goes into the posterior mesenchyme cells (B 7 ' J , fig. 200.)
| |
| Fig. 185. Eight cells, posterior view; spindles of the fourth cleavage present.
| |
| Fig. 186. Fourth cleavage, vegetal pole.
| |
| Fig. 187. Fourth cleavage, animal pole; 16 cell.-.
| |
| | |
| | |
| | |
| | |
| | |
| PLATE XII.
| |
| | |
| Whole Eggs of Ciona intestinalis ; Sixteen Cells to Gastrula.
| |
| | |
| Crescent substance stippled in nil the figures; clear protoplasm also stippled in figs. 188-191.
| |
| Pobty bodies are actually present where drawn.
| |
| | |
| Fig. 188. Sixteen cell stage, anterior pole.
| |
| | |
| Fig. 189. Sixteen cells seen from vegetal pole; all but two of the cells are passing from the 5th to the
| |
| 6th generation.
| |
| | |
| Fig. 190. Similar stage viewed from animal pole.
| |
| | |
| Fig. 191. Animal pole view, 5th to the 6th generation; 20 cells; when the divisions are completed,
| |
| which are here indicated by spindles, there will be 32 cells.
| |
| | |
| Figs. 192-195. One and the same egg in the 32-cell-stage. Fig. 192. From animal pole. Fig. 193.
| |
| From vegetal pole, spindles present in many of the cells, which are about to pass into the
| |
| 7th generation. Fig. 194. From posterior pole; all nuclei at the animal pole in a
| |
| resting condition. Fig. 195. Antero-vegetal view.
| |
| | |
| Figs. 196 and 197. One and the same egg seen from the animal (ventral) and from the vegetal (dorsal) poles; most of the cells at the vegetal pole have just passed into the 7th generation ;
| |
| spindles are present in all the cells at the animal pole preparatory to their division into
| |
| the 7th generation ; 44 cells, 20 of the 6th and 24 of the 7th generation.
| |
| | |
| Figs. 198-204. With the exception of fig. 199, all these figures represent views of one and the same
| |
| egg. Fig. 198. View from animal pole showing all the ectoderm cells in process of
| |
| division. Fig. 199. Another egg of the same stage as the preceding, but seen from a
| |
| more anterior point of view. Fig. 200. Vegetal pole showing all the cells at this pole
| |
| in a resting condition. Fig. 201. Same egg seen from a more anterior point of view.
| |
| Fig. 202. Same egg viewed from the anterior pole. Fig. 203. Viewed from the right
| |
| side showing in dotted outline the great elongation of the cells of the vegetal pole and a
| |
| corresponding flattening of the cells at the animal pole. Fig. 204. Viewed from the
| |
| posterior pole. 76 cells, viz., 32 ectoderm, !< endoderm, 18 mesoderm, 8 chorda, 8 neural
| |
| plate; when the divisions indicated in the ectoderm cells are completed there will be 64
| |
| ectoderm cells and 108 cells in all.
| |
| | |
| Fig. 205. Gastrula corresponding in stage to fig. 140 of the Cynthia egg; 178 cells, vis., 108 ectoderm
| |
| and neural plate (ventral), 20 endoderm, 18 mesenchyme, 12 muscle, 8 chorda, 12 neural
| |
| plate (dorsal).
| |
| | |
| Fig. 206 and 207. Older gastrula with blastopore much restricted by the growth of the anterior lip
| |
| Fig. 206. View of superficial ectoderm and mesoderm. Fig. 2(17. Deeper view of
| |
| same embryo showing endoderm and mesoderm.
| |
| | |
| | |
|
| |
|
|
| |
|