Paper - The Organization and Cell-Lineage of the Ascidian Egg
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Conklin EG. The Organization and Cell-Lineage of the Ascidian Egg (1905) J. Acad., Nat. Sci. Phila. 13, 1. Conklin 1905 TOC: I. The Ovarian Egg | II. Maturation and Fertilization | III. Orientation of Egg and Embryo | IV. Cell-Lineage | V. Later Development | VI. Comparisons with A.mphioxus and Amphibia | VII. The Organization of the Egg | Summary | Literature Cited | Explanation of Figures
| Online Editor |
|---|

See also - Conklin EG. Mosaic Development In Ascidian Eggs (1905)
|
| Historic Disclaimer - information about historic embryology pages |
|---|
|
The Organization and Cell-Lineage of the Ascidian Egg


By Edwin G. Conklin.
Professor of Zoology, University of Pennsylvania,
With Plates I-XII.
Philadelphia - 1905
Contents
- A. Organization of the Egg
- B. Ascidian Embryology
- C. Methods and Material
- Development of the Ovocyte
- Test Cells and Chorion .
- Structure of Fully Formed Ovocyte
II. Maturation and Fertilization
- A. Maturation
- Disappearance of Nuclear Membrane
- Chromosomes
- Nucleolus
- Spindle Formation
- Movements of Spindle and Nuclear Plasm ; Formation of
- B. Fertilization
- Entrance of Spermatozoon
- Movements of Ooplasm
- Localization of Yellow Protoplasm
- Localization of Clear Protoplasm and Yolk
- Development of Sperm Nucleus and Aster
- Path of Spermatozoon within the Egg .
- The Egg Nucleus and its Movements
- Sperm Amphiaster and First Cleavage Spindle
- Dispermy
- Polar Bodies
III. Orientation of Egg and Embryo
- Van Beneden and Julin's System of Orientation
- Seeliger's System
- Samassa's System
- Castle's System
- Evidences in Favor of Van Beneden and Julin's System
A. Nomenclature
B. Cleavage of the Egg ; First to Seventh Generation of Cells
- First Cleavage ; 1-2 Cells
- Second Cleavage ; 2-4 Cells .
- Third Cleavage; 4-8 Cells .
- Fourth Cleavage; 8-16 Cells
- Fifth Cleavage ; 16-32 Cells
- Sixth Cleavage ; 32-64 Cells
C. Gastrulation ; Seventh to Ninth Generation of Cells
7. Seventh Cleavage; 64-76 Cells, 76-112 Cells .
8. Eighth Cleavage ; 112-132 Cells, 132-218 Cells
- Closure of Blastopore
- Development of Larva
VI. Comparisons with A.mphioxus and Amphibia
- Axial Relations of Egg and Embryo
- Entrance of Spermatozoon
- Cleavage
- Blastula and Gastrula
- Closure of Blastopore
- Neural Plate
- Chorda
- Origin of Mesoderm
VII. The Organization of the Egg
- A. Polarity
- B. Symmetry
- C. Cytoplasmic Localization
- Localization in the Cleavage Stages Cytoplasmic Organization and the Nuclear
- Localization before Cleavage
- D. Genesis of the Organization of the Egg
- Role of the Nucleus in Differentiation - Inheritance Theory
- Factors of Localization
- Cytoplasmic Movements
- Cell Division as a Factor of Localization
- Types of Germinal Localization ; Evolution of Types
- Annelid-Mollusk Type
- Ctenophore Type
- Echinoderm Type
- Ascidian Type
- Ascidian Embryology
- Cytological Results
- Organization of the Egg

Plate I

Plate II

Plate III

Plate IV
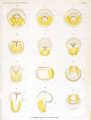
Plate V

Plate VI

Plate VII

Plate VIII

Plate IX

Plate X

Plate XI

Plate XII











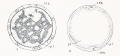
Fig 1-2
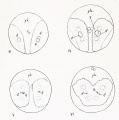
Fig 3-6
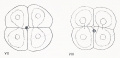
Fig 7-8
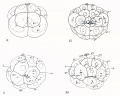
Fig 9-12
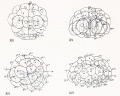
Fig 13-16
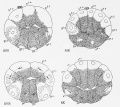
Fig 17-20
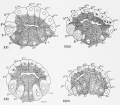
Fig 21-24
Fig 25-26

Fig 27-33
Fig 34-35










Introduction
A. Organization of the Egg
Recent years have witnessed a revival of the ancient controversy as to the nature and contents of the germ cells. On the one hand are those who with Weismann maintain that the egg must contain the elements or determinants of very many structures which will appear in the course of development; on the other hand are ranged the modern epigenesists who find in the egg cell only complex chemical substances which have the capacity under certain outer conditions of undergoing regular transformations into other substances which incidentally have peculiar forms, just as crystals have.
But while this modern controversy recalls the ancient one between the
adherents of evolution and those of epigenesis, it does so chiefly because it proceeds
from the same temper of mind, and not because anyone today is ready to defend
the views of either the evolutionists or the epigenesists of a century ago. No one
now expects to find in the egg or sperm a predelineated germ with all adult parts
present in miniature, neither can anyone now maintain that the egg is composed
of unorganized and non-living material. Everyone now admits that the truth is
somewhere between these two extremes ; the real problem is how much or how
little of organization is present, and not whether the germ is organized at all.
Though the controversy as to evolution and epigenesis has thus been narrowed within relatively small limits, and has therein- lost much of its startling and
picturesque character, it is none the less a real and important controversy today.
In general the attitude of physiologists and those who deal with the processes of
development has ever been to jdace emphasis upon the epigenetic character of
development and the extremely simple structure of the germ; whereas those who
are concerned chiefly with organic structures are prone to seek for antecedent
structures in earlier and earlier stages of development and so finally in the
unsegmented egg itself.
It is not many years since all embryological studies were dominated by the
eerm-laver theory, since the time when germ layers were considered to be the
earliest appearing differentiations which could be profitably compared and homologized. More recently it has Been shown that such differentiations appear at a
stage much earlier than the formation of the germ layers, ami that many of
the early cleavage cells of different animals show such fundamental resemblances that they can be homologized with one another. May not these differentiations
appear at still earlier stages and " organ-forming germ regions" be marked out in
the egg before cleavage begins'.' Finally does the organization of the egg arise
de novo in the ovary, or ma\ a certain part of it be carried over from generation to
generation, and is this early organization of the egg in any way different from the
organization of any cell? These are problems of profound importance which lie at
the basis of any thorough study of development, inheritance and evolution.
B. Ascidian Embryology
Anyone who has observed the ascidian egg will understand why it has been such a favorite object of study. The cleavage of the egg is so beautifully regular and can be observed so readily in life that it is not surprising that ascidians were among the first animals to which the "cell-lineage" method was applied. It is surprising, however, to find such diversity of opinion with regard to the development of these animals ; even in some of the most important points in the early development, such as the relation of the poles of the egg to those of the gastrula, or the cell origin of the germ layers, scarcely two authors agree, in spite of the fact that these eggs are perhaps as favorable as any others in the whole animal kingdom for the stud}' of these problems. Under the circumstances it is impossible to avoid the conclusion that a large amount of very careless work has been done in this field. Accuracy is ever the avowed aim of science, and when one finds scientific work unpardonably inaccurate he may justly feel indignant. For some reason or other ascidian embryology has ever been a field of conflict and confusion. In some cases controversy has raged for years around a mere blunder which has thus gained a sort of immortality rarely attained by accurate work .
But ascidian embryology furnishes illustrations not only of inaccurate work; it also affords some of the most classical examples of accurate and substantial research. Considering the time when the}- were written, Kowalevsky's papers (1866, 1871) are models of accuracy. The beautiful studies of Van Beneden and Julin on the segmentation of the ascidian egg and on the morphology of the tunicates (1884, 1886) surpass in excellence anything which had been done up to that time, and in some respects they have not been equalled by an}' more recent work on the development of these animals. Chabry's (1887) classical paper on the normal and teratological embryology of ascidians is a masterpiece, one of the first and best illustrations of the application of the experimental method to the study of embryology. After this, the most important work dealing with the early development of ascidians is that of Castle (1894, 1896), although it is marred by some fundamental mistakes; he has applied in detail the method of cell lineage to the study of the development of Ciona intestinalis, and has followed the history of the individual cleavage cells farther than had any of his predecessors. Other work which deserves mention has been done by Kupffer (1870), Seeliger (1885), Davidoff (1889), Samassa (1894) and others, and will be referred to later in the body of this work.
In the face of all this antecedent work, it may seem superfluous to devote yet
another paper to this subject, and nothing was further from my purpose when I began. It was in the attempt to find out the manner in which the ascidian conies to form its polar bodies at the endodermal pole, as described by Castle, that I
was led to conclusions radically different from his, as will be described later, and this
induced me to make a detailed study of the cell-lineage of three different genera of
simple ascidians. In such a Held it may lie expecting too much to hope that m\
observations will meet with general acceptance; hut perhaps it may he proper for
me to say that I have spared no pains or labor to make them accurate.
C. Material and Methods
Early in July. 1903, while working at the Marine Biological Laboratory, Woods Holl, Mass.. I began the study of the maturation and fertilization of the egg of Ciona intestinalis (L.) Flemming, with the aim mentioned in the preceding paragraph. Only a small number of these animals was to be found at that time at Woods Holl. though they occurred more abundantly later in the summer. I therefore turned my attention to two other simple ascidians. Molgula manhattensis Verrill and Cynthia (Styeld) partita Stimpson, both of which occur in considerable numbers in the Woods Holl region. The verj first lot of the living eggs of Cynthia which I examined showed a most remarkable phenomenon and one which modified the whole course and purpose of my work ; for there on many of the unsegmented eggs, which were of a slate-gray color, was a brilliant orange-yellow spot, which in other eggs appeared in the form of a crescent or band. Further observation showed that this crescent became divided into two equal parts at the first cleavage and that it could be followed through the later cleavages and even into the tadpole stage. I thereafter, for a considerable portion of the summer, devoted myself to the study of the living eggs of Cynthia, and a record of these observations will be found in the body of this work and in plates I-V. Afterward I took up also the living eggs of Ciona and Molgula, and finally I fixed and prepared for microscopical examination, both as whole objects and as serial sections, the eggs and embryos of all three of these genera.
Castle (1896) has described in considerable detail the time and manner of egg
laying in these three genera, and his observations lean entirely confirm. The eggs
of Ciona and Molgula are laid in the early morning, a little before daybreak, while
those of Cynthia are laid in the late afternoon, a little before sunset. These
ascidians rarely lay eggs the first da\ they are in the laboratory. Since the
yellow pigment of the egg of Cynthia is difficult to observe by artificial light, it
was necessary to take eggs from the ovary or oviducts and artifically fertilize
them in the morning in order to lie able to study by daj light the later stages in the
development. A lame proportion of such eggs never develop, though the eggs
seem ripe and the spermatozoa are active; however, some of them develop into
normal embryos and tadpoles, and from such I have obtained material for the study
of the later stages of the living egg. Whenever possible, however. 1 have relied
upon eggs which were normally laid and fertilized, inasmuch as all such develop
normally. In Ciona and Molgula it is very eas} to artificial!} fertilize the eggs; in fact, almost every ripe egg of Ciona will develop if fertilized with sperm from
another individual, hut if fertilized with sperm from the same animal the eggs
rarely if ever develop, as Castle has shown. This is due to the fact that such
spermatozoa never enter the egg, though they may be quite active. Morgan (1904) has recently discussed this interesting fact in a suggestive manner.
The method which I employed in studying the living eggs of these ascidians
was very simple ; they were placed in several drops of fresh sea water upon a
glass plate and were covered by a large cover glass, which was supported by
pieces of thin glass about 200 n thick. In such condition the eggs can be rolled
over at will by pushing on the cover glass, and, if drops of water are occasionally
added at the edge of the cover, the eggs will continue to develop normally for
two hours or more. Inasmuch as the entire development of Cynthia up to the formation of the free-swimming tadpole normally occupies not more than eight to
twelve hours, depending upon the temperature, it will be seen that a considerable
portion of the development can be followed on a single egg. I do not doubt that
with proper precautions the entire development might be followed on a single egg;
however, since eggs which have been along time under a cover glass develop slowly
and may become abnormal, and since there was nothing to be gained for my purposes by the observation of a single egg through the whole development rather than
of several eggs through consecutive portions of it, I chose the latter and easier
method.
All my studies of the living eggs of these ascidians were made with a dry lens,
the 4 mm. Apochromat of Zeiss which, with the No. 4 ocular, gives a magnification
of about 260 diameters. Even with a magnification of 50 diameters or less the
yellow crescent of the Cyiitliia egg is plainly visible. In order to see this crescent
to the best advantage, especially with high powers, it is necessary to use wide angle
lenses with open diaphragm and clear white light. The fact that Castle studied
the development of this species but makes no mention of this yellow crescent is
difficult" to explain. I can only account for it by supposing that he obtained the
eggs in the evening and studied them by yellow artificial light.
Preserved material was fixed in various fluids, Perenyi's,. Kleinenberg's, PicroAcetic, Sublimate and Sublimate-Acetic. For the study of entire eggs and embryos
Kleinenberg's fluid followed by the Picro-Ha?matoxylin, which I have used with
success on molluscan eggs, gave incomparably better results than any other method.
Eggs so stained were mounted in balsam under thin cover glasses without supports
of any kind, and were studied with an oil immersion lens, the 3 mm. Apochromat
of Zeiss. Pyy occasionally applying a drop of xylol to the edge of the cover glass
the balsam remains sufficiently soft so that the eggs can be rolled into any position
desired. For serial sections, material iixed in Boveri's Picro-Acetic gave the most
satisfactory results. Such material was stained on the slide in Delafield's Hematoxylin and Eosin or in Iron Hematoxylin and Bordeau red.
Castle states that he found it necessary to remove the egg envelopes by drawing the egg into a pipette through an opening so small that the egg alone could puss in, after the manner recommended by Chabry. In most cases I have
found that the presence of the egg envelope's (iocs not seriously interfere with clear
seeing, possibly owing to the fact that in the study of preparations I have used an immersion lens in which the depth of locus is relatively slight. In late stages,
however, the test cells are sometimes confusing, and in the case of Ciona I found
that these, together with the other envelopes, could he removed by simply rolling the eggs under the cover glass. In Cynthia the envelopes may sometimes he removed in the same way, though not so easily as in Ciona.
Conklin EG. The Organization and Cell-Lineage of the Ascidian Egg (1905) J. Acad., Nat. Sci. Phila. 13, 1.
Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - The Organization and Cell-Lineage of the Ascidian Egg. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_The_Organization_and_Cell-Lineage_of_the_Ascidian_Egg
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G