Paper - Address Upon The Development Of The Brain
| Embryology - 23 Jun 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
His W. Address upon the development of the brain. (1897) Trans. Royal Acad. Medicine Ireland.
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
Address Upon The Development Of The Brain

of Leipzig.
Delivered in the Theatre of the Royal Dublin Society to the Bellows of the Royal Academy of Medicine in Ireland, on the 11th June, 1897, during the Meeting of the Anatomical Society of Great Britain and Ireland in Dublin. (Reprinted from the Transactions of the Royal Academy of Medicine in Ireland, 1897)
Dublin: Printed By John Falconer, 53 Upper Sackville St.
Mr. President and Gentlemen,
It may be a little bold to deliver before you an English address, as I shall scarcely be able to express myself conveniently in your language. I must place all the responsibility on my dear friend, Mr. Cunningham ; he has induced me to accept the honoured invitation of the Royal Academy of Medicine, and also to speak on the development of our brain, a theme which he could have treated himself as one of our best authorities.
It cannot be my task to enter into the high architectural complexities of the noblest of our organs ; but, proceeding from embryologies! observation, I shall try to obtain some general points of view on the fundamental principles of its organisation.
Since the researches of C. E. v. Baer, and of R. Remak, we know that the nervous centres of vertebrates arise from he primary ectoderm as a canal of an epithelial character. it also well-known that the brain-part of the neural canal forms, in the first instance, three enlargements - the forebrain, the mid-brain, and the hind or rhombic brain.
These three parts can be distinguished very early, even before the canal is closed. From the time of their first formation they follow each other in a curve, or rather in a broken line. Owing to this curve the extremity of the brain canal is turned ventrally, and it extends beyond the extremity of the notochord and of the alimentary canal (Fig. 1).

|
Fig. 1. Photographic reproduction of black-board drawing made by Prof. His for his Lecture.
B. Diagrammatic representation of early condition of the neural tube, showing the manner in which it becomes bent on itself. A. Diagrammatic representation of a mesial longitudinal section through the head and neck of a human embryo. The brain-flexures and the optic vesicle are seen, and the manner in which the fore-brain is bent around the anterior extremity of the notochord and foregut is indicated. The early heart is shown bulging out in front of the foregut. The different sub-divisions of the brain are marked off from each other by dotted lines. In both diagrams (A as well as B) a dotted line running in the long axis of the neural tube indicates the separation of the alar from the basal lamina of the lateral wall. |
The primary formation of the head is the same in all cramote vertebrates. It shows a ventrally free, and a fixed part - the fore-head and the back-head ; the latter encloses the heart. The fore-head is formed by a frontal and a facial piece. The frontal portion contains only the fore-brain, while the facial piece includes the blind end of the foregut and the extremity of the notochord. There is a small field between the frontal and the facial pieces of the head, where the base of the brain, the extremity of the notochord, and the extremity of the foregut join together. This field is of great morphological importance, and it shows the place where the hypophysis is afterwards to be formed. So long as the junction between the foregut, the brain, and the notochord exists there is no room for a hypophysis. But this junction is a temporary one, and when it has been interrupted the ectodermal pouch of the hypophysis enters into the gap.
The next advance in the development of the brain brings about the separation of the eyeball from the forebrain, the division between the hemispherical and the thalamic parts of it, and also the segmentation of the hindbrain from the medulla oblongata. The boundary between the two last-mentioned parts is marked by a flexure called the pontine flexure (Fig. 6, p. 11 ; Fig. 14, PL II.).
Before following further changes, I have to say some words about the histological development of the neural canal. The wall of the neural canal shows already, before it is closed, two different kinds of elements - (1) cells extending through the whole thickness of the wall, and forming a columnar epithelium ; and (2) round germinal cells placed on its inner side.
The columnar epithelium undergoes a special transformation. Between the inner parts of its cells arises a regular succession of open clefts whilst the tops of the single columns remain united by a fine limiting membrane. The peripheral parts of the cells form a sponge-like network with irregular meshes. The whole formation is called the myelosponge, and its elements are termed the spongioblasts (Plate I., Fig. 10 and Fig. 11).
The round germinal cells are dispersed through the clefts of the internal columnar part of the myelosponge. Many of these cells are in full process of multiplication, and show the different stages of karyokinesis. Others grow out in the form of pear-shaped corpuscles (Plate I., Figs. 10 and 11 ; Plate II., Fig. 12). Each of these sends out a tapering protoplasmic prolongation, directed outwards towards the surface of the medulla. These pear-shaped corpuscles are the neuroblasts, and their prolongations constitute the origin of nerve-fibres. The formation of neuroblasts is confined to the two thick lateral walls of the neural canal. The roof and the floor of the tube are thinner than the side-walls, and they furnish only epithelial cells, or spongioblasts.
The primary neuroblasts have merely neural processes ; the dendrites or ramified processes arise later than the nerves, and it seems that these attain very slowly their full development. A certain number of the newlv-formed nerves leave the neural canal to reach muscles, whilst others remain in the thickness of its wall. We find therefore motor fibres and internal or intra-medullary fibres. The motor fibres all come from the ventral half of the canal; the internal fibres start partly from its dorsal, partly from its ventral half. Some of them are commissural fibres, and cross the mid-line ; others enter into the longitudinal columns of the medulla.
All sensory nerves, also the fibres of the olfactory, the optic, and the auditory nerves arise outside the neural canal. The sensory nerves spring from the bipolar cells of the spinal ganglia; the fibres of the olfactory nerve from the epithelial cells of the olfactory groove, and the true optic fibres from the neuroblasts of the retina. All these nerves reaching the neural canal from without apply themselves to its surface, and form longitudinal bundles. The dorsal column of the spinal cord, the spinal roots of the trifacial nerves, the auditory and the optic nerves are all in their first appearance in loose connection with the neural canal.

Fig. 2. Photographic reproduction of a black-board drawing made by Prof. His for his Lecture. It gives a diagrammatic representation of a transverse section through the spinal portion of the early neural tube. It is sufficiently described in the text.
A section through the brain or the spinal cord of a human embryo of about four weeks gives the plainest representation of the typical architecture of nervous centres. We find the motor fibres starting from the ventral half of the canal (Plate II h, Fig. 10), the sensory fibres reaching the surface of its dorsal half, and internal fibres forming connecting arches (Fig. 2). These three kinds of fibres represent the groundwork of all central organisation : they supply the paths from the periphery to the centre, those from the centre to the periphery, and the paths from one part of the centre to another. Every fibre coming from a single cell forms with its cell a histological and physiological unit, which we now call a neuron.
I return to the history of the brain. The most striking feature in its morphological development is the general increase of its flexures ; that increase shows many variations in the different classes and orders of vertebrates. Generally the two limiting flexures of the mid-brain precede the others in their progression. The base of the fore-brain is brought near to that of the hind-brain. A narrow cleft between these two parts leaves the place for the dorsum sellte (Fig. 6, p. 11; Plate If., Fig. 14); this cleft persists even in the adult brain. By the new nomenclature it is called the interpeduncular groove, and it shows an anterior and a posterior recess, corresponding to the two primary limits of the mid-brain.
The flexures of the rhombic brain follow a little later in their development. The pontine flexure at the superior, and the neck-flexure at the inferior end of the medulla oblongata become very considerable, and the neural canal assumes thenceforth a zigzag-like conformation (Plate V., Fig. 25). There is also a constant but feeble flexure in the isthmus or narrow part of the rhombic brain. At the same time the neural canal loses its primary simplicity. Its parts begin to acquire more characteristic forms, and we can soon distinguish all the principal parts of the developed brain.
Before entering into any details it will be necessary to follow the transformation of the primary longitudinal Jaminee. If we leave aside the purely epithelial or ependymal laminee of the neural canal, its roof and its floor, we find its wall consisting of a ventral or basal, and of a dorsal or alar lamina. As we have already seen the basal lamina contains the motor centres, whilst the dorsal lamina receives the sensory nerves. The boundary- line between these two laminae of the wall is generally well marked in the brain canal; its terminal end can be followed through

Fig. 3. Photographic reproduction of a black-board drawing made by Prof. His for his Lecture. It gives a diagrammatic representation of a transverse section through the rhombic or hind-brain (medulla oblongata), and is specially intended to show the alar and basal longitudinal laminae of the lateral wall of the tube. Into the alar lamina the sensory fibres from the cells of the ganglion of the tenth nerve (vagus) are entering, so as to form a longitudinal column of fibres cut transversely and lying close to the surface. This is the - solitary tract - which later becomes deeply buried within the substance of the medulla. The neuroblasts of the nuclei of origin of the twelfth (hypoglossal) and of the eleventh or spinal accessory nerves are seen sending their motor fibres out from the medulla.
[In this drawing Professor His makes use of the English word zone, and he thus follows his distinguished American friend, Professor. C. S. Minot, of Boston. (See Minot’s Human Embryology, p. 606). But zone is altogether an inappropriate word to apply to either of the two the third ventricle to the optic stalk, or optic recess. The further development of these two laminae is disproportionate in the different parts of the brain. In man and in the higher vertebrates the basal lamina diminishes as we ascend from the lower to the higher parts ; the motor centres begin to be discontinuous and to fail, and the last of them - the oculomotor centre - belongs to the mid-brain. The basal lamina of the fore-brain has no motor centres and is very reduced. It contains the mammillary body and the infundibulum, and is separated from the thalamus by a shallow furrow. We call it now the hypothalamus.
The alar lamina of the brain canal remains feeble in the medulla oblongata, but its growth is mighty in the higher parts, and results in the development of the cerebellum and of the cerebral hemispheres. In lower vertebrates the basal or hypothalamic lamina of the fore-brain is of more importance than in the higher ones, whilst the alar or hemispheric part remains less developed.
In the rhombic brain the alar lamina furnishes the restiform and the olivary bodies, the cerebellum and the superior peduncles, whilst the basal lamina gives the floor of the fourth ventricle. In the embryo of four weeks the rhombic brain when seen in transverse section has a pentagonal form, the epithelial roof being broad and the basal and alar laminae forming together an obtuse angle. The motor centres lie near to the surface of the basal laminae, and their fibres leave the canal wall
portions of the neural tube in question. The term zone could only be properly given to an area or strip of the neural wall which is placed at right angles to the long axis of the tube and which extends, girdle-like, around its circumference. The districts referred to, however, so far from presenting this relation, run parallel to the long axis of the tube. Still further, by using the word - zone - we do not give a correct translation of the German term (riattc) employed by Professor His himself. 1'liipelplaUc (alar lamina) and Gmndplatte (basal lamina) are the names used by Professor His to designate the two thick portions of each lateral wall of the neural canal. In the text the term zone has been changed to lamina.) in two lines - a mesial one and a lateral. Near the lateral motor line is the place where the sensory fibres come from without to the surface of the medulla, to form longitudinal bundles of fibres. As typical instances we can cite the two motor lines of the hypoglossal and the spinal accessory nerves, and close by the sensory roots of the pneumogastric nerves (Fig. 3). The longitudinal bundle receiving the sensory fibres of pneumogastric and of glossopharyngeal nerves is called the solitary tract ; its primary position is very superficial. In the fully developed medulla the solitary tract, and also the nuclei of the hypoglossal and accessory nerves, are separated from the ventral surface by a broad layer of tissue containing nerve-cells and also nerve-fibres running in all directions. How is this layer formed? The dorsal border of the alar lamina stands straight and erect during its primary stages, but it afterwards begins to bend outwards in the form of a longitudinal fold, which projects in a lip-like way along the entire length of the rhombic brain (Fig. 4). This rhombic lip (Rautenlippe) is the origin of the cerebellum and of the olivary bodies. (Plate II., Fig. 13.)

Fig. 4. Photographic reproduction of a black-board drawing by Prof. His to explain the folding over of the rhombic lip of the rhombic or hindbrain in the region of the fourth ventricle. It represents a diagrammatic transverse section through the medulla oblongata.
The formation of the cerebellum is not very difficult to understand, but that of the olivary bodies presents a greater amount of complexity, and is one of the most interesting processes in the history of the brain. The rhombic lip becomes adherent to the remaining part of the alar lamina, and sends out a multitude of neuroblasts which wander towards the middle line and pass on the ventral side of the basal lamina, superficial to the solitary tract, and the motor nuclei. These neuroblasts and their arched fibres are supported by the outgrowing myelospouge of the basal lamina, and they form a thick layer containing together with other nuclei those of the olivary bodies. Afterwards longitudinal fibres of different origins and new arch-like fibres join them to produce the final organisation.

Fig. 5. Photographic reproduction of a black-board drawing by Prof. His to show the migration of neuroblasts from the region of the rhombic lip - a developmental process by means of which the solitary tract becomes deeply buried within the substance of the medulla. This figure represents a later stage of the medulla than that depicted in Fig. 3, and, as in the latter, it shows the neuroblasts which give origin to the motor fibres of the hypoglossal and spinal accessory nerves.
The medulla gives a good example ot the relation between the local and the chronological succession ol the layers of the brain. The oldest layer of the medulla is the floor of the fourth ventricle, with its nuclei. It is followed in the first instance by the reticular formation, and afterwards by the layer containing the olivanes and other nuclei. Last of all come the pyramids and the outer arcuate fibres (I ig. bA).

Fig. 6. Figure obtained of the head and neck of a human embryo of five weeks from a series of consecutive sections by the reconstruction method. It exhibits the different sub-divisions of the brain at this period, and shows particularly well the thalamic brain with the early outgrowth from it which ultimately forms the cerebral hemisphere.
In the pons Yarolii similar relations between the chronological and local succession of layers are to be established. For instance, we can cite the primitive position of the motor nuclei of the trifacial nerve and of its spinal sensory root, and compare it with the later position, which shows these parts covered by thick layers of transverse and longitudinal fibres. In the mid-brain the tegmental part is the first formed. It presents itself as a thick prominence of the basal canal wall. In this prominence we early distinguish the nucleus of the oculomotor nerve and the posterior longitudinal bundle ; then comes also a reticular formation, followed by the substantia nigra. The fibres of the crusta are partly very late formations.
I beg leave to limit my remarks concerning the thalamic brain to the demonstration of photographs, and to pass immediately to the primitive history of the cerebral hemispheres. The cerebral hemispheres are formed by the terminal piece of the alar brain lamina, and this place can be determined very early as an angular field situated between the optic vesicle, the terminal, and the dorsal border of the neural canal (big. 1, p. 2). This field has two free borders and one fixed; the free borders being the terminal and the dorsal ones, the fixed border remaining in continuity with the thalamic brain and with the stalk of the optic vesicle.
The primary hemispheric brain is undivided. I he separation of two hemipheres by a longitudinal fissure begins in the human embryo at the end ol the first mouth. From this time we distinguish also the place of the nasal brain or rhinencephalon of Sir W. Turner (Fig. fiA). The rhinic brain is situated immediately beneath the terminal border of the brain tube. Between its two halves appears the thin lamina cinerea. It represents the terminal section of the alar brain lamina; the suture of the basal lamina is enclosed in the optic chiasma and in the anterior vail of the infundibulum.
The nasal brain is limited by Sir William Turners rhinic fissure, and it is divided by another fissure, the fissura prima, into an anterior and a posterior part. The anterior part furnishes the olfactory bulb, tract, and trigone, and the field of Broca (area Brocae). The posterior part remains as the wall of the anterior perforated space, and as the gyrus subcallosus (peduncle of the callosum). In the developed brain the anterior part of the rhinencephalon is fixed to the frontal lobe, the posterior part to the temporal lobe; these relations will enable us to distinguish from the earliest times the position of the hemispheric lobes. It is easy to see that the frontal lobe is directed partly towards the anterior, partly towards the dorsal border of the hemispheres. The temporal lobe is represented by the inferior part of the fixed border, the occipital lobe by its superior part, and the parietal lobe by the posterior part of the dorsal border (Fig. (iA).
The rhinic fissures and the fissura prima are folds of the brain-wall - and on its internal side they form three prominences, the first traces of a striate body. This part of the brain-wall increases very quickly, and forms the well-known prominence in the floor of the lateral ventricle. During many months the striate body shows its three branches, and remains in close relation to the lobes of the nasal brain.
I he problem of brain formation has its histological as well as its morphological side, and we have to consider both. To a large extent the changes in the form of the brain depend upon the increase of its histological elements ; but the increase of these elements is again ruled by the iorm of the surrounding parts, inasmuch as cells and nerves need Iree room to extend themselves.
To understand the complicated interlacement of the different processes, it is necessary to follow their chronological succession. Time is the mightiest factor in every development. As we have seen before, the local position of the layers of the brain depends on it ; but it possesses tar, even much more, influence on the physiological organisation of the neural system. Some examples may explain this: - In the spinal cord, the myelosponge is the first formed part of the tissues, its inner clefts furnishing the room for the germinal cells and for the neuroblasts, the meshes of the outer network are only pervious to nerve-fibres, but not to nerve-cells. On this account the different density of the myelosponge in its inner and outer layers determines the separation of the grey and white matter of the spinal cord. In the brain things seem to be more complicated. We find there evident cell-emigrations from the inner to the outer side of the canal wall.
The formation of nerves begins with the motor fibres ; the sensory fibres follow soon afterwards. These two kinds of fibres are not only formed very early, but this formation is also soon brought to an end. All the later generations of neuroblasts furnish only internal connecting fibres. The cells are formed during the first two or three months of the embryonic life, but the fibres going out from them can grow indefinitely long, before they reach their destination. Also the dendritic or ramified processes of central nerve-cells and the collateral fibres grow very slowly, and we can scarcely say how long their formation and their increase can continue. The regeneration of sectioned nerves shows that even during later life nerves can continue to grow.
In the whole neural canal, especially in the brain, the alar lamina is slower in its development than the basal lamina. But as it grows continually through long periods of time enormous organs like the cerebral hemispheres may be produced. In every case the new formation of neural elements must necessarily be a process of superaddition. The cells and the nerve-fibres once formed persist : they are joined by otter cells and fibres formed afterwards. The older and the younger cell formations become connected together ; and, by the succession of different generations, the whole system will become more and more complicated. In all these things time will multiply the most intimate connections between the different parts. The medulla oblongata is the first formed of the brain parts. All the later formed parts enter into connection with it, and finally it is the most central and most complicated of all these parts. We may, therefore, make the generalisation, that during the earlier periods of development the single elements of the neural system in their rough connections are formed, and that only during the later developments the intimate organisation of the whole system and the dependence of all its parts upon each other is established.
All the higher functions of our brain depend upon the anatomical and physiological superposition of neural centres. By physiologists such superpositions have been presumed many years ago, but only the inquiries of recent times have given full evidence of their anatomical disposition. The iron methods of Weigert, the silver methods of Golgi and of Cajal, and the methyline blue methods of Ehrlig have armed anatomical workers with strong weapons. We can now trace the nerve-fibres from their beginning to their final terminations ; we can distinguish the fibres entering into a neural centre from the fibres issuing from it. The rich and very unexpected system of collateral fibres has been uncovered before our eyes - a system which enables a single fibre to enter into relations with long series of terminal points. By the new anatomical methods applied to the undeveloped brain Flechsig and Held could, for instance, follow the auditory paths from their first origin in the cochlear ganglion to the nuclei of the medulla oblongata. From thence new paths are established to the upper olivary bodies, and to the lower quadrigeminal body, and at last come the paths from these centres of the mid-brain to the cerebral cortex of the temporal lobe. There are at least four different cell stations, every one being a centre with many particular connections, and all united together in a harmoniously working apparatus.
But the brightest victory of anatomical methods is the analysis of the cerebral cortex given in these last years by Prof. Flechsig. The controversies concerning the localisation or non-localisation of cerebral functions are not new. C4all on the one side, Flourens on the other, were in former periods the champions of the two parties. Since that time the physiological inquiries of Hitzig, Ferrier, Horsley, Schafer, and many others, have given the decision for the localisation, and the experiments of Mr. Horsley have taught us how precisely the centres of different muscles can be isolated in the cortex. Prof. Flechsig on his side pursuing, for more than twenty years, by anatomical methods the course of central nerve-bundles, has at last succeeded in limiting in a sharp way the different special sensory centres of the cortex - the centres of the optic, the auditory, the olfactory nerves, and those of the nerves of general sensibility. Each of these centres has its own particular field on the brain surface - the optic centre in the occipital, the auditory centre in the temporal, the olfactory centre in the frontal lobe, while the centres of general sensibility are situated partly in the frontal and partly in the parietal lobe. All the centres receive projection fibres from the internal capsule. They are separated one from the other by large fields of cortex, which are without any connection with projection fibres. The sensory cortex-centres show medullated fibres from the, time of birth, or soon afterwards.
In the rest of the cortex the medullated fibres appear later, and they develop from the side of adjacent sensory centres. These parts of the cortex are regarded by Prof. Flechsig as centres of association, and it may be remarked that of the two principal centres of association the parietal one is placed between the optic, the auditory, and the tactile centres, whilst the other, the frontal one, has only few direct connections with the sensory centres.
I cannot go farther in following the consequences of these new discoveries, but I think that we anatomists feel profoundly satisfied by the fact that discoveries of such general importance can be made by our science. We rejoice not only in what we know, but even much more in the perspective of coming knowledge. Anatomy is not a closed science, as it was asserted some time ago. It is a science full of life, and one of the most promising branches of general biology.
Explanation of Plates
The figures in the following five plates are reproductions of some of the more important lantern slides which were shown by Professor His during his Lecture. Figures 7-17 (Plates I.-III.) are photographs from the original preparations. Figures 20-27 are photographs from wax models.
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Plate I


Fig 7. Mesial longitudinal section through the neural tube of an early chick embryo. The aperture or neuropore (n.p.) at the anterior end of the tube is still open.
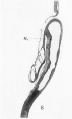
Fig. 8. Mesial longitudinal section through the head-end of a slightly older chick embryo, showing the intimate connection between the anterior end of the notochord (n.) and the hinder part of the floor of the fore-brain. Note that the neuropore is closed.
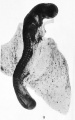
Fig. 9. Early embryo of a dog-fish. The upper end of the figure shows ie enlargement and bending of the brain-part of the neural tube.

Fig. 10. Micropliotograph of a portion of a transverse section through the neural tube. The inner ends of the spongioblastic columns are observed to be flattened out and joined together to form a fine limiting membrane. This is seen in the upper part of the section. Between them three germinal cells, two on the left with karyokinetic formations, may be seen. Human embryo of four weeks.
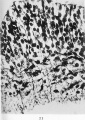
Fig. 11. In this microphotograph the neuroblastic cells are darkcoloured, and in connection with many of these the tapering axis cylinder processes are seen. It will be observed that these processes are growing towards the surface ( i.e the lower part of the figure) through the mesh-work of the myelosponge. Human embryo of four weeks.




Plate II


Fig. 12. Microphotograph of a portion of a transverse section through the whole thickness of the neural tube. The arrangement of the neuroblasts and of the spongioblasts is well seen. The limiting membrane on the inner aspect of the tube is uppermost in the figure. Human embryo of four weeks.
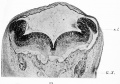
Fig. 13. Transverse section through the early medulla showing the rhombic lip. r.l. Rhombic lip. G.x. Ganglion of the vagus.
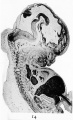
Fig. 14. Mesial section through the head of a human embryo of the seventh week. It shows the great expansion of the cavity of the brain-part of the neural tube as well as the strongly-marked brain flexures.

Fig. 15. Photograph of a human embryo of six weeks. The different subdivisions of the brain shining through the thin head-coverings are well seen, and the cervical, pontine and mid-brain flexures observed, in section, in Fig. 14 are prominently brought out. Compare with Fig. 6 in the text (page 11).




Plate III

Fig. 16. Transverse section through the spinal part of the neural tube of a human embryo of four weeks. It shows the early condition of the roots of a spinal nerve. A diagrammatic representation of this is given in Fig. 2 in the text.
c.c. Central canal of neural tube.
m.r. Motor fibres from the neuroblasts in the wall of the tube, growing out to form the anterior nerve-root.
S.G. Spinal ganglion, from the cells of which the fibres grow into the wall of the neural tube and thus form the posterior nerve-root.
Fig. 17. Section through a portion of the wall of the brain tube at the level of the origin of the fifth cranial nerve. It has been found necessary to turn this figure round on its side so as to make it fit into the plate.
w. Wall of the brain-tube.
G.G. Gasserian ganglion. x.r. Sensory root, formed of fibres growing out from the cells of the ganglion and entering the wall of the neural tube. a.T. The same fibres as represented by s.r. cut transversely owing to their having turned downwards to form the spinal root of the fifth nerve.
m.n. Neuroblasts of the motor nucleus of the fifth nerve giving origin to the motor fibres m.r . which are seen growing out from the neural wall. m.r. Fibres of the motor root of the fifth nerve.
Human embryo of four weeks.
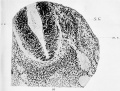
Fig. 16. Transverse section through the spinal part of the neural tube of a human embryo of four weeks. It shows the early condition of the roots of a spinal nerve. A diagrammatic representation of this is given in Fig. 2 in the text.
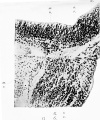
Fig. 17. Section through a portion of the wall of the brain tube at the level of the origin of the fifth cranial nerve.


Plate IV

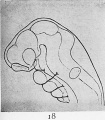
Fig. 18. Reconstruction drawings of the head and brain of early human embryo to show the brain flexures.
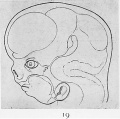
Fig. 19. Reconstruction drawings of the head and brain of later human embryo to show the brain flexures.

Fig. 20. Interior view of the brain-tube of a human embryo of four weeks (model).
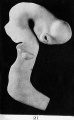
Fig. 21. Outer surface of the brain-tube of a human embryo of four weeks (model).
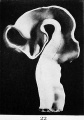
Fig. 22. Interior of the brain-tube of a human embryo of four and a half weeks (model).
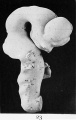
Fig. 23. Outer surface of the brain-tube of a human embryo of four and a half weeks (model).






Plate V

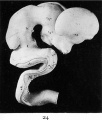
Fig. 24. Outer surface of the brain of a five weeks human embryo (model).
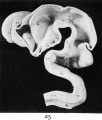
Fig. 25. Interior surface of the brain of a five weeks human embryo (model).
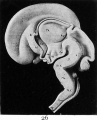
Fig. 26. Mesial section of the brain of a human embryo of the third month (model).
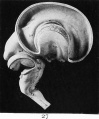
Fig. 27. The same brain seen from the outside, with the lateral ventricle opened (model).
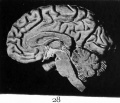
Fig. 28. Mesial section of the adult brain to compare with Fig. 26, and to show how similar the basal part of the thalamic brain is in both. The basal portion of the wall of the third ventricle is shaded more lightly than the rest.





{kind=link}
Cite this page: Hill, M.A. (2026, Haziran 23) Embryology Paper - Address Upon The Development Of The Brain. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_Address_Upon_The_Development_Of_The_Brain
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G