Book - The Emergence of Experimental Embryology in the United States
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Robert Hilfer RS. The emergence of experimental embryology in the United States. (1990) National Library of Medicine Bethesda, Maryland.
| Online Editor |
|---|

|
| Historic Disclaimer - information about historic embryology pages |
|---|
|
The Emergence of Experimental Embryology in the United States

by S. Robert Hilfer, Ph.D.
National Library of Medicine, Bethesda, Maryland 1990
US. Department Of Health And Human Services • Public Health Service • National Institutes of Health
This booklet was prepared in conjunction with an exhibit of the same title at the National Library of Medicine, Bethesda, Maryland, June 1 - August 31, 1990.
Cover Illustration: Chicken embryo, from M. Malp Dissertatio epistolica de formatione pulli in ovo (London: Joannem Martyn, 1673).

This booklet is dedicated to the memory of Professor Oscar Schotte of Amherst College.
Single copies of this publication may be obtained without charge by writing: Chief, History of Medicine Division National Library of Medicine 8600 Rockville Pike Bethesda, Maryland 20894
Background

For many centuries embryological thought was dominated by a controversy commonly called the Preformation-Epigenesis argument. In the fourth century B.C. Aristotle recognized that development was progressive, so that organs formed in sequence within the embryo and not all at once. This is the first statement of epigenesis. Aristotle belonged to a Greek tradition that tried to explain life by observation, believing that explanations for phenomena that were not understood could be found by studying nature rather than by invoking the supernatural. Similar thoughts were expressed by the Greek physician, Galen, in the second century A.D. Little progress was made for the next 1000 years. From that time through medieval times, the writings of earlier workers were accepted as fact and there was little attempt to correct inaccuracies.
During the Middle Ages, a point of view arose that all structures required for the production of a new individual were preformed within the egg and that development required only their unfolding and growth in size. With the discovery that the egg and sperm are the agents responsible for reproduction, this argument for preformation took the form of the egg or the sperm as containing the entire body of the new organism. The proponents of this theory became known as ovists or animiculists, depending upon whether they believed the egg or the sperm to contain the intact individual. The preformationists became convinced that each new generation had to exist preformed within the egg or sperm of the preceding generation. The ridiculous nature of this argument was pointed out in the 1720's by the physicist Hartsoeker, when he calculated the size that the fiftieth generation would have been within an egg. This turned out to be a negative number. However, his argument was not accepted by the preformationists for many years. Hartsoeker himself (1694, Fig. 1) had apparently accepted the preformationist view earlier in his career.
With the emergence of the Renaissance, the naturalist tradition became reestablished. The renewed interest in the examination of embryos provided additional descriptions of development, for instance by DaVinci, Fabricius, Harvey, Spallanzani, and Malpighi (1673, Fig. 2). Most of these observations were interpreted as supporting preformation and it took the discoveries of cellular structure generated by development of the microscope for real progress to be made. The careful observations on the progressive nature of organ formation by Wolff in the second half of the eighteenth century were instrumental in ending rigid preformationist viewpoints.

|
| Figure 2. Illustrations from M. Malpighi (1673) of developing chicken embryos from the time of laying to 36 hours of incubation. This plate is representative of the keen observations done on early embryos during the 17th century. |
Beginnings of Experimental Embryology
The early nineteenth century was marked by great progress in the development of techniques to study cells and tissues as well as by the development of concepts important to embryological thought. The importance of the egg for embryonic development was established by von Baer (1827, Fig. 3), the cell theory was advanced, and it was recognized that the embryo developed from cell sheets. About the mid- 19th century, Darwin's writings on evolution began to appear and the concept that developing embryos went through stages that recapitulated their ancestors was given an evolutionary interpretation. An early proponent of recapitulation was the Swiss naturalist, Louis Agassiz, who believed (Agassiz, 1859, p. 174) that "the phases of development of all living organisms corresponds to the order of succession of their extinct representatives in past geological time." In the next few decades, Haeckel popularized the concept and introduced several other embryological ideas, such as the gastrea hypothesis and the theory of germ layers. The century was also a period of intensive work on comparative embryology by descriptive techniques.

|
Figure 3. Plate from C.E. von Baer (1827) to illustrate his discovery of the human ovum. He was the first to show that the ovum lies within the follicular structure described by de Graaf, which generally was recognized as too large to be an egg. |

|
Figure 4. Surface views of frog embryos from W. Roux (1888). His Figures 5 and 6 show normal embryos at early and late stages of formation of the neural tube. His Figures 7 to 10 show embryos in which one blastomere was killed at the two-cell stage. In Figure 7, the ectoderm has covered much of the surface of the dead blastomere whereas much less repair has occurred in the other cases and it is clear that only partial embryos have formed. He interpreted these results as proving that development is mosaic in character, each blastomere giving rise to a restricted part of the individual. |
Wilhelm Roux, the German embryologist, frequently is given credit for founding experimental embryology with his classical experiment published in 1888 (Roux, 1888). Actually, experimental intervention with development was tried sporadically at earlier times, but it was not until the 1850's that experiments were done with a clear purpose in mind. Newport, in England, began testing the effects of various chemicals on fertilization of frog eggs and devised techniques to keep the eggs in a particular position.
His careful observations led to the demonstration that the plane of the first cleavage as well as the point of sperm entry into the egg usually mark the axis of the embryo. Other lines of investigation in the latter half of the century were concerned with the way embryos inherited the characteristics of their species. Several theories were advanced, which became the bases for experimental work that led to modern concepts.
Roux was a dynamic writer and speaker and his forceful personality probably played a large part in the reputation he received for originating the experimental approach. Based upon theoretical arguments of the time, he used an experimental approach to attack a fundamental problem of development. The question Roux asked was whether the embryo contains all of the information necessary for development or if it is influenced by the environment. He was concerned with differentiation of parts of the organism from the seemingly unspecialized egg. His (1874) had earlier proposed that different regions of the cytoplasm of a fertilized egg contain different materials needed to make specific parts of the developing embryo. Roux tested this hypothesis by killing one cell of a two-celled frog embryo (Fig. 4). He reasoned that if different regions of the egg formed different parts of the embryo, the organism resulting from his manipulation should be defective, whereas it should be normal if controlled by the environment. The result he obtained was a partial embryo and gave rise to a point of view that can be considered a modified expression of preformationism.

|
Figure 5. Illustrations from H. Driesch (1892) showing effects of manipulating blastomeres of sea urchin eggs. His Figure 1 shows a normal 16-cell stage (copied from Selenka). Figure 2 shows a half-embryo from an 8-cell stage and Figures 3 and 4 half-embryos from a 16-cell stage. Figure 5 is a half-embryo at the blastula stage. These half-embryos developed into complete individuals. Figures 7 through 9 show blastulae in the process of dividing. These formed conjoined twins as shown in Figures 10 through 12. These results were interpreted as demonstrating regulative development. (Courtesy of the Marine Biological Laboratory.) |
It was shown in subsequent experiments that Roux misinterpreted his results. At the time, however, his experiments renewed interest in the idea that formation of an embryo results from mechanical partitioning of a specialized egg. This modified view of preformation became known as determinate development and Roux compared the structure of the egg to a mosaic in which small regions of the cytoplasm constructed specific parts of the adult, just as small parts of the mosaic construct a picture. In contrast, experiments by Driesch (1892, 1902) on sea urchin embryos gave the opposite result (Fig. 5). Driesch isolated blastomeres of two-cell stages by shaking the embryos in sea water. Each individual blastomere was able to develop into a complete individual, thus showing that the embryo could regulate to replace the missing parts that normally would have been formed by the missing cytoplasm. If the blastomeres did not separate completely, double embryos were formed. This developmental pattern was called indeterminate and corresponds to epigenetic development. Thus, the theoretical argument had shifted from preformation vs epigenesis to mosaic vs regulative development. On the one hand, the search for preformed individuals within egg or sperm was replaced by attempts to recognize specialized cytoplasmic domains within the fertilized egg. On the other hand, the epigenetic approach gradually became shaped into studies on the structure of cytoplasm and the establishment of pattern by interactions among cells of the embryo or by external forces as development proceeds.

|
Figure 6. Photograph of Louis Agassiz giving a lecture. Agassiz was a dynamic speaker and attracted large audiences at popular lectures in the United States. He married an American and settled at Harvard University. He did much to popularize biology and was responsible for training a number of embryologists in summer classes as well as at Harvard. (Courtesy of the Marine Biological Laboratory.) |
The Role of Marine Stations in Experimental Embryology
In the mid 1800's, a naturalist tradition was well established in the United States. Most biologists were teachers and there was little opportunity for graduate training. The first significant American training centers were established by Agassiz, who had come to Harvard from Switzerland, and by W. K. Brooks, who established the Darwinian tradition at Johns Hopkins. Interest in the experimental approach was generated by contact with European laboratories and writings. C. O. Whitman broke with American tradition and went to Germany for graduate study. He returned to assume a professorship at the University of Chicago, bringing the new interest in experimental embryology with him. Four students of Brooks - E. G. Conklin, E. B. Wilson, T. H. Morgan, and R. G. Harrison - also joined the ranks of the new experimental biologists and made major contributions to the establishment of experimental embryology in this country. All worked at times in European laboratories; for instance, Conklin, Wilson and Morgan were introduced to marine organisms at the recently established Naples Zoological Station.
The embryos of marine organisms became favored material for embryological research because they were easier to observe and manipulate than amphibian or bird embryos. European marine stations were established as centers of experimental research and many European workers such as the Hertwigs, Boveri, Driesch, and Herbst used their facilities. In the United States, the first marine stations were established as teaching facilities. Agassiz opened a small summer school for teachers on the island of Penikese off Woods Hole, Massachusetts in 1873 (Fig. 6). Brooks at Johns Hopkins established the Chesapeake Zoological Laboratory in Virginia in 1878 and one of Agassiz's students, Hyatt, founded a school at Annisquam, north of Boston, in 1880.
The Marine Biological Laboratory in Woods Hole, Massachusetts grew out of the Annisquam school. It was founded with a research mission as well as that of teaching and consequently played a significant role in the development of experimental embryology in this country. Another of the students in Agassiz's first summer class, Whitman, was appointed director and the first building was constructed in 1888 for classes that summer (Fig. 7). The faculty were involved in research programs as well as in teaching and tended to return year after year to do research. The teaching and research missions overlapped and researchers and students were exposed to the latest ideas in embryological theory, as well as other subjects, in the series of evening lectures that were held each summer. The speakers ranged from Whitman discussing preformation vs epigenesis, through Wilson's and Morgan's arguments on heredity, to lectures on geology, evolution, and other branches of science. Visitors came from Europe, including well established embryologists as well as younger ones.

|
Figure 7. Photograph of a laboratory class at the Marine Biological Laboratory, circa 1900. Leonard W. Williams (standing, center) was the instructor. From its inception, courses at the Marine Biological Laboratory have played an important role in training teachers and scientists in modern experimental techniques, a role that continues today. (Courtesy of the Marine Biological Laboratory.) |

|
Figure 8. Diagrams of early cleavage stages of the tunicate, Cynthia partita, from Conklin (1905). The four cells at the bottom of the upper left figure are traced through succeeding cleavages into a series of blastomeres that give rise to segmental muscles and other mesodermal derivatives. This is an example of the painstaking care that was needed to trace regionalization of egg cytoplasm through development. |
Cell Lineage and the Attack on Haeckel's Germ Layer Theory
Some embryologists, such as Harrison, did not participate in research on marine organisms; the questions they formulated were better studied on vertebrates. They also found it disruptive to work on material that was available during only a part of the year and did not want to move their laboratories to a summer station. This did not diminish their contributions to experimental embryology, as will be described below.

|
Figure 9. Diagrams of cleavage stages of a polyclad worm (A and C), a mollusc (D) and annelid worms (B and E) from Wilson (1892). Wilson was impressed with the similarity in the formation of mesoderm by these different groups of organisms. The shaded cells are the precursors of mesoderm. Wilson interpreted studies of cell lineage as showing that development is mosaic. |
Haeckel's theory of the germ layers proposed that organization of the embryo occurred only after the three germ layers of ectoderm, mesoderm, and endoderm became established. This represented an epigenetic position that was attacked by the proponents of determinate development. The first approach that was taken to discover if early cell divisions could be related to developmental pattern, was the tracing of cell lineages, which became known as mapping cell fates. This procedure allowed the descendants of early blastomeres to be traced to the organs they formed during normal development. Organisms were chosen for study that had distinctive cleavage patterns or pigments to provide natural markers which could be followed during successive cleavages.
The Woods Hole workers were active in this field. An early study was done by Whitman (1878) in Europe on the leech Clepsine (now Glossiphonia). He showed that the mesoderm, nervous system, and excretory organs arise at distinct cleavages from pairs of cells that he called teloblasts. This type of ordered development was shown for the worm, Nereis, by Wilson (1892), the mollusc, Crepidula, by Conklin (1897) and the ascidian, Cynthia (now Styela), also by Conklin (1905, Fig. 8). All of these organisms were similar in that entire organ systems could be traced back to one or a pair of blastomeres at an early developmental stage. Furthermore, blastomeres having similar positions in different organisms were responsible for similar organs (Fig. 9). Thus, development of different organisms showed homology that was assumed to relate to their evolutionary origins. These studies did much to discredit the germ layer theory and strengthened the ideas that specific regions of the zygote (fertilized egg) are set aside (or determined) early in development to form specific organs.
Fate mapping continues to be a useful technique to the present time. Techniques have changed to match the organism being studied. The German embryologist W. Vogt (1925) introduced the use of vital dyes to follow cleavages and movements of amphibian blastomeres. Dyes were not appropriate for the small cells of bird embryos and Spratt (1946) introduced the technique of marking surfaces with fine carbon particles. In more recent times bird and mammalian fate maps have been prepared by using new fluorescently labeled dyes that can be injected into single cells, by transplanting groups of cells labeled with radioactive precursors of DNA synthesis, or by making chimeras between species having distinctive nuclear structure. These studies have all been important in tracing the movements of cells during normal development.
Studies on Epigenesis the Physicochemical Nature of Cytoplasm

|
Figure 10. Experiments by J. Loeb, as in this paper from 1893, gave additional support to the argument that sea urchin eggs do not contain specialized areas of cytoplasmic determinants. Treatment of fertilized eggs with hypotonic sea water resulted in the formation of cytoplasmic blebs (upper left) that remained attached to the main embryo during cleavage (upper right) and resulted in doubled embryos at the blastula stage (lower left). These embryos developed into Siamese twins (lower right). The egg cytoplasm was able to form most of the parts of more than one individual. |
The alternate concept to that of determinate development was that blastomeres become specialized as a result of the position they come to occupy in the embryo. This represents the extreme regulative position as opposed to mosaicism. Proponents of this viewpoint believed that answers to how blastomeres become specialized lie in the physicochemical characteristics of cytoplasm, which appears homogeneous in the zygote. Experiments had been done to distort the organization of egg cytoplasm as a way of testing for mosaic development. In England, Jenkinson (1909) subjected eggs to pressure, while other investigators placed zygotes or early embryos in centrifugal fields. In spite of these insults, the embryos developed normally. Jaques Loeb was a strong proponent of this viewpoint and as a result of his studies much was learned about the physical properties of cytoplasm. One of his early experiments was to cause rupture of the cell membrane by placing fertilized eggs in hypotonic sea water. Blebs formed that either separated completely from the cleaving cytoplasm or remained partially attached. The result was duplication, either as complete embryos or as Siamese twins or triplets (Fig. 10). Loeb concluded that "the number of embryos which come from one ovum is not determined by preformation of germ regions in the protoplasm, or nucleus, but by the geometric shape of the ovum and the molecular condition of the protoplasm" (Loeb, 1893, p. 58). Loeb's main contributions were on the physical chemistry of proteins. His interest in the chemistry of cytoplasm led him to work with fertilization and as a result he discovered the cause of artificial parthenogenesis.
Other proponents of a physicochemical interpretation of development included Matthews, Lillie, and Just. All three had serious disagreements with Loeb and held to old interpretations of colloid chemistry while Loeb's ideas were continuously changing. Matthews worked on the effects of salt on the properties of cytoplasm, using some embryological material, but in the latter part of his career he concentrated on nerve cells. Lillie was a student of Whitman, whose primary interest was in fertilization; he is best known for his fertilizin-antifertilizin theory of fertilization. He served as the second director of the Marine Biological Laboratory and was instrumental in its expansion.

|
Figure 11. Illustrations from E. B. Wilson (1904), showing the effect of removing the polar lobe in the mollusc, Dentalium. |
Figures 4 to 9 show normal early cleavages. Regions of cytoplasm have different densities and shadings; a special cytoplasmic region becomes segregated in the polar lobe (1, and 1 2 ). When the polar lobe was removed, major parts of the larval body were missing (Figures 32 to 37) as compared with normal development (Figures 29 to 31).
These results suggested that the polar lobe contains special cytoplasmic determinants.
Experimental Analysis of Cytoplasmic Determinants
Just was a student of Lillie's and also was greatly influenced by Matthews. As a black biologist, he was confronted by the prejudices of the time and his work was accomplished under the serious disadvantages of not gaining an appointment at a research oriented university; thus he had no graduate students, had access to colleagues with similar interests only during the summer, and had an excessive teaching load during the rest of the year. He has been described as a brilliant observer and a perfectionist. He became the outspoken proponent of Lillie and Matthews against Loeb and was subjected to personal as well as scientific attacks by Loeb because of his stand. Just's major contribution to the analysis of cytoplasm was in establishing the role of the egg cortex during fertilization. He also provided some of the early experimental support for Lillie's theory. The later work of Just included analyses of all of the chemical components of the egg cell, including nucleic acids. In his later unpublished writings he may have been on the verge of discovering the role of nucleic acids as the genetic material, although to the end of his career he remained a strong proponent of regulative development.
Investigations on the properties of cytoplasm also led to discoveries on cell motility and relationships of cells to their substrate. Ross Harrison (1910) originated procedures for growing cells in isolation from the animal body, thus establishing the techniques of tissue culture. His early work was directed at solving problems of nerve development. He showed that nerve cells produce axons by outgrowth and not by fusion of several cells. S. Ramon y Cajal in Spain discovered the growth cone and Paul Weiss showed that nerve fibers become oriented by the substratum. At the same time, Warren and Margaret Lewis were successful in growing chick and mammalian cells in culture and studied their migrations, while Holtfreter isolated amphibian cells and studied their behavior.
A position intermediate between indeterminate and determinate development was expressed by Wilhelm His (1874). He believed that the cytoplasm of the egg contains germinal regions that are responsible for forming organs as a result of physiological interactions during the course of development. His believed that development was not truly mosaic nor was it totally regulative. The fate mapping experiments of Whitman, Wilson, and Conklin supported His's argument. It appeared that organ-forming regions could be related to specific regions of cytoplasm in the zygote. Constituents of these cytological germ regions of His later became known as cytoplasmic determinants.
The idea of cytoplasmic determinants was tested by several experimental approaches. Separation of blastomeres of Amphioxus by Wilson (1893), of a ctenophore by Driesch and Morgan (1895), and of tunicates by Conklin (1905) suggested that some regulation could occur but that eventually during development defects were produced when blastomeres were deleted. Similarly, removal of polar lobes from eggs of spirally cleaving molluscs and worms resulted in defective larvae (Fig. 11). Eventually Wilson evolved an explanation which stated that development starts as regulative and that different organisms became mosaic at different points during embryogenesis.
The interest in and search for cytoplasmic determinants continues to the present time. This field has seen a merging of ideas from the other lines of embryological research. As advances were made in the newly emerging discipline of genetics, it became clear that the composition of cytoplasm is controlled by active genes and that all genes need not be active in all cells. It also was established that genetic information is stored in the egg during maturation and is used during early embryogenesis. Identification of stored proteins and messenger RNA that are differentially distributed within the zygote's cytoplasm and in emerging embryos gives promise of leading to an understanding of developmental control mechanisms at a genetic level. The recent molecular biological approaches are confirming the theories proposed by His, Wilson and their contemporaries.
Establishment of Developmental Genetics
He modified preformationist argument of the late 1800's implied that embryogenesis occurs during cleavage through distribution of inherited information to specialized regions of the body. His's argument was that inheritance occurs through distribution of cytoplasmic factors. In contrast, August Weismann suggested that inheritance occurs through the cell nucleus and that the developing organism is a mosaic of nuclei having specialized potentials. This was part of the Germ Plasm Theory, which postulated that hereditary determinants were carried only in the cell lineage giving rise to gametes. Roux set out to test the hypothesis that nuclei become specialized with his experiments in which blasomeres of frog embryos were killed; he interpreted the results as support for Weismann. However, it soon became apparent that Roux's original experiment was flawed. The presence of the killed blastomere influenced the attached living one, whereas the first two blastomeres when separated from each other both developed normally (McClendon, 1910). Nevertheless, as in other so-called regulative organisms, blastomeres of later stages were not able to form complete embryos.
In the early twentieth century, Spemann tested the potential of nuclei at later developmental stages by a clever procedure. He reasoned that if nuclei become specialized during cleavage, they should not be able to support complete development if introduced at a later time into uncleaved cytoplasm. He used a fine hair to constrict frog eggs so that nuclei could not enter part of the cytoplasm. After the fourth cleavage when 1 6 nuclei were present, he released the constriction and a nucleus entered the uncleaved zygote cytoplasm. In spite of the delay in arrival of nuclei until a stage when isolation experiments gave partial embryos, these embryos were normal. It was not until much later that it became possible to transplant nuclei of older embryos into the cytoplasm of an uncleaved egg (Briggs and King, 1952). By this method it was shown that nuclei from blastula through early neurula stages are able to support development of a complete embryo. More recently, it has been demonstrated that nuclei from even mature tissues when transplanted to an egg can support normal development of a fertile adult organism. These experiments have proved that specialization of nuclei during development does not involve loss of genetic information and support the alternate hypotheses of Weismann's contemporaries that inheritance occurs through differential nuclear activity rather than by loss of genetic information.
The rediscovery of Mendel's writings did much to stimulate interest in inheritance in the early 1900's. Wilson became interested in the cytological basis of inheritance after performing his experiments on cytoplasmic determinants. He came to view the nucleus as playing a crucial role in controlling development and devoted much of his later career to these problems. His book on "The Cell in Development and Heredity" (1925) is a classic which is still worth reading because it is so modern in its approach.

T.H. Morgan was the most influential scientist in establishing genetics as a separate field of biology. In doing so, he changed from a strong supporter of totally regulative development to a proponent of nuclear control of development. He also gave up at least temporarily his interests in embryology to pursue the establishment of the genetic basis of inheritance. He was instrumental in causing a split between embryology and genetics that lasted for many years. In fact, O.E. Schotte pictured the way an embryologist and geneticist looked upon the cell during this time as either mostly cytoplasm or mostly nucleus (Fig. 12).
Morgan was concerned primarily with establishing the gene as the basis of genetics. He did this by examining mutants and established Drosophila as the organism of choice for these studies. He was responsible for describing many characteristics and showing that they could be related to sequential arrays on specific chromosomes. Nevertheless, Morgan retained a concern for with the problem of how genetic control might be exercised during development. In his Nobel Prize address, delivered in 1934, Morgan (1935, p. 15) remarked that "It is conceivable that different batteries of genes come into action one after the other, as the embryo passes through its stages of development. This sequence might be assumed to be an automatic property of the chain of genes." In discussing the genetic control of development, he stated (Morgan, 1935, p. 16):
We have also come to realize that the problem of development is not as simple as I have so far assumed to be the case, for it depends, not only on independent cell differentiation of individual cells, but also on interactions between cells, both in the early stages of development and on the action of hormones on the adult organ systems. At the end of the last century, when experimental embryology greatly flourished, some of the most thoughtful students of embryology laid emphasis on the importance of the interaction of the parts on each other, in contrast to the theories of Roux and Weismann that attempted to explain development as a progressive series of events that are the outcome of selfdifferentiating processes or, as we would say today, by the sorting out of genes during the cleavage of the egg. At that time there was almost no experimental evidence as to the nature of the postulated interaction of the cells. The idea was a generalization rather than an experimentally determined conclusion, and, unfortunately, took a metaphysical turn.
Today this has changed, and owing mainly to the extensive experiments of the Spemann. school of Germany, and to the brilliant results of Horstadius of Stockholm, we have positive evidence of the farreaching importance of interactions between cells of different regions of the developing egg. This implies that original differences are already present, either in the undivided egg or in the early formed cells of different regions. From the point of view under consideration results of this kind are of interest because they bring up once more, in a slightly different form, the problem as to whether the organizer acts first on the protoplasm of the neighboring region with which it comes in contact, and through the protoplasm of the cells on the genes; or whether the influence is more directly on the genes. In either case the problem under discussion remains exactly where it was before. The conception of an organizer has not as yet helped to solve the more fundamental relation between genes and differentiation, although it certainly marks an important step forward in our understanding of embryonic development. Answers to the questions raised above are becoming available only today with the advent of molecular genetics and a renewed merging of interest in developmental biology and genetics.
Another line of research that contributed basic information for the eventual merging of embryology and genetics was the physiological approach espoused by Goldschmidt. Goldschmidt came to this country to escape Nazism and found it difficult to gain acceptance in the American scientific community. Part of this came from the unpopular point of view he took of attempting to unite embryology and genetics into a common field of developmental genetics. He stimulated work on genetic analysis of development through use of mutants rather than through surgical manipulation. Embryology to him was subservient to genetics and simply a way of examining the time that mutations had their effect on development. His point of view was expressed most forcefully in his book, "Physiological Genetics" (1938). Goldschmidt's position that the genes in the nucleus are the most important factor in development gained support from Hammerling's experiments on the alga, Acetabularia. In the 1930's, Hammerling showed that the structure of the spore-forming body is controlled by the nucleus. The cap-like structure differs in different species and upon transplanting the nucleus of one species into the cytoplasm of another, the structure was changed to that of the species from which the nucleus came. Goldschmidt's work eventually resulted in renewed interest by geneticists in the embryo. Current work on pattern forming genes is an outgrowth of a long line of studies in developmental genetics, primarily on Drosophila, but on mammals as well. The renewal of a combined interest in genetics and development, however, depended upon the writings of other workers. One of the most influential was Waddington in England, who in "Organizers and Genes" (1940) played an important role in the process of reuniting the two disciplines.
Embryonic Induction and Control of Differentiation
In the early 1900's embryologists increasingly turned away from the mosaic-regulative argument and became more concerned with the problems of differentiation of organs. One of the prime movers in this new interest was the German, Hans Spemann. By using hairloops to constrict embryos, he showed that duplications of the head region could be produced by pinching along the axis of the egg. As a result, he questioned how different organs arose and what controlled the order with which they formed. The first step was to discover what part of the embryo controlled duplications; presumably this event occurred before gastrulation was completed because duplications were not obtained from older embryos. Furthermore, the dorsal edge of the blastopore, where cells initially move from the surface to the interior of the embryo during gastrulation, seemed to be most important. Only constrictions that went through the dorsal "lip" caused duplications. Testing the role of the dorsal lip was accomplished by transplanting this region of the embryo to different sites in a host embryo. Hilde Mangold (nee Proscholdt), a Ph. D. student to whom he assigned this problem, discovered that the dorsal lip of the blastopore, when placed into the future abdominal region, produced duplications (Spemann & Mangold, 1924). These always occurred as a duplication of the body axis (Fig. 13), and Spemann coined the term "organizer" to describe the role and "induction" to describe the action of the dorsal lip. The extra tissue resulted in the formation of a "secondary axis". Even before this publication Spemann, as well as W. Lewis in the United States, had demonstrated interactions among different parts of the embryo as necessary for normal development, the best known being the requirement of the eye primordium for development of a lens.

|
Figure 13. The classical experiments of Spemann and H. Mangold (1924) demonstrated induction in amphibian embryos. A graft of the dorsal lip of the blastopore was made from an unpigmented species to a pigmented species. The graft was placed in the future ventral region of the body. At the neurula stage, the normal neural plate is found on the dorsal surface (left) and a second neural plate, containing the graft on the ventral surface (center). The result was the formation of a secondary embryonic axis which produced Siamese twinning (right). |
Spemann had a number of talented students who in the next few decades examined the properties of what became known as embryonic induction. The organizer was shown to correspond with the presumptive notochord region and to have a differential effect at different levels along the axis of the embryo (O. Mangold, 1933). Although cells could be induced to form new structures, these were limited to the genetic information in the responding cells and not to the inducing tissue. A prime example was the work done on induction of mouth parts in amphibians (Spemann & Schotte, 1932). Transplantation of ectoderm from the region that was fated to become belly skin in a frog to the mouth region of a salamander produced mouth parts but had the organization belonging to structures of the frog mouth region. The reverse experiment produced salamander mouth parts in the frog mouth region. Thus, the mouth region was able to induce belly ectoderm to make mouth structures, but they conformed to the genetic information provided by the organism from which the donor ectoderm came.
Finally, it was shown by Holtfreter, another student of Spemann's, that exogastrulation, in which the cells move outside instead of into the embryo, results in an absence of induction. The endodermal and mesodermal mass formed some internal organs, but there was a failure of axial organs, including the nervous system and somites. Holtfreter also was instrumental in providing a new method of testing induction. He realized that transplants to a host embryo, or insertion into the blastocoel cavity as was used in later Spemann experiments, did not isolate the responding ectoderm from organismal influences. As a result he searched for a way to test the responding tissue in isolation. This was accomplished by devising a medium of physiological salt solution in which isolated amphibian cell layers remained healthy and responsive to inductive influences. He showed that combinations such as ventral ectoderm and chordamesoderm would form neural structures just as in the embryo. This system also served to study other sources of induction, such as chemicals.
Within a decade, the role of induction came into question because of the discovery that various killed tissues could cause induction of limited neural structures. The response to these findings took two forms. Holtfreter stimulated a line of research that showed these "evocations," as he called them, to result from limited proteolysis of the responding cells. He concluded that potential in the cells was released, or as Needham and Waddington in England proposed, evoked by limited damage caused by the killed tissue or chemical agents. This line of research has resulted currently in a better understanding of the capabilities for self differentiation of groups of cells within different regions of the developing embryo. The other line of research concerned attempts to elucidate the chemical nature of the organizer, especially of the neural "inducer" after evocation was discovered.
There was great interest in the chemical control of development at the time. It had been demonstrated, for instance, that different regions of the embryo have different respiratory rates. This was incorporated by Child into a theory that development can be explained by gradients existing within different planes of the embryo. For the amphibian, he proposed that one gradient exists from animal to vegetal pole and that a second dominant gradient is established at the dorsal lip at the onset of gastrulation. Attempts to document gradients within the embryo as well as to isolate active inducing substances led to confusing results. The work was done in England, Sweden, and Germany as well as by Barth and his coworkers in the United States. It appeared that these experiments supported the notion that a variety of substances could release the ability of competent ectoderm to undergo only limited development. It is only in the last few years that progress has again begun to be made in understanding the way the cells of the dorsal lip establish the axis of the developing embryo.

|
Figure 14. Summary diagram of experiments by S. Horstadius (1928) on regulative development in sea urchins. These experiments, combining animal hemispheres, (closed semicircles) with macromeres (large circles) and micromeres (small circles), showed that normal development results from a balance of animal and vegetal blastomeres. Too few macromeres and/or micromeres reduces the completeness of endodermal and mesodermal differentiation. Note also that skeleton (rods), normally formed from micromeres, can form in the absence of micromeres if the macromere population is sufficiently large. |
Another line of investigation that provided evidence that cells of one part of an organism exert an influence upon another part concerned the sea urchin. This work was an outgrowth of the experiments of Driesch, followed by the work of the American embryologists. It became apparent that even sea urchin embryos are not completely regulative. It was recognized that the vegetal pole cells, the micromeres formed at the fourth cleavage, are different from the rest of the blastomeres in other ways besides their smaller size. In a series of brilliant experiments the Swedish embryologist, Horstadius, demonstrated that the micromeres play an important role in the control of morphogenesis. It had been shown earlier that a variety of chemical treatments produced two classes of embryos, those in which the formation of either dorsal or ventral structures was inhibited. Horstadius (1928) was able to separate and combine layers of blastomeres into hybrid embryos. He showed that either the central layers or the animal and vegetal ends, when combined, could regulate to produce a normal larva (Fig. 14). Furthermore, addition of micromeres to a layer of animal tier blastomeres was enough to permit normal development. As a result, Horstadius proposed that development of the sea urchin is regulated by gradients of an inductor released by the animal pole and an opposing inductor released by the vegetal pole. This scheme was similar to that proposed for amphibian induction and the parallel between the two organisms was heightened by the ability of extra micromeres implanted in the animal region to induce a second embryonic axis consisting of a duplicated digestive tract. As with amphibian embryos, the search for chemical inducers has resulted in marginal success.
Tissue Interactions and Differentiation
Interest in tissue interactions during embryogenies were pursued along lines other than studies on induction. One of the leaders in this field was Ross Harrison, already mentioned above, who occupies as prestigious a niche in American scientific history as Spemann does in Germany. As early as 1904 Harrison showed an interest that was to occupy most of his later career. He tested the role of nerves in muscle development by removing the neural tube from an early embryo and showed that limb development still was normal. From 1920 on, Harrison devoted most of his efforts to explore the origin of axial organization and symmetry in the embryo. He was particularly concerned with the establishment of symmetry in the limb and used the method of transplantation to ask what controls the establishment of the limb axes (Harrison, 1921, Fig. 15).
Holtfreter also was interested in tissue interactions other than induction. He combined different layers of the embryo in his cultures and showed that they behaved in isolation the same way as they did in the embryo, endoderm taking an internal position when combined with mesoderm and ectoderm becoming external. The study of cell interactions as opposed to tissue interactions became a reality when Holtfreter discovered (as often occurs in science, based upon preliminary experiments by others) that lowered calcium concentration in the culture medium resulted in the cells of a tissue becoming separated, or dissociated. He then was able to reassemble masses of cells from different tissue origins and to test their interactions. His discovery that the cells separated into groups based upon their origin was a revolutionary finding. It had been anticipated in earlier studies by H. V. Wilson and Galtsoff on sponges, who showed that dissociated cells from different species will segregate or "sort out" when they are intermixed. Holtfreter proceeded to catalogue the positions that cells took in different combinations, and concluded that they conformed to the normal relationships in the embryo (Townes & Holtfreter, 1955, Fig. 16). This work led to current investigations on cell adhesion molecules, cell communication through specialized junctions, and interest in the role of extracellular matrix in morphogenetic movements.
Holtfreter's work was done on amphibian embryos. It took many more years for methods to be devised that allowed the cells of birds and mammals to be separated while retaining their viability. Two important contributions were made by Moscona, who showed that trypsin, and Zwilling, who showed that a calcium chelating agent, would effect dissociation in bird embryos. Moscona's experimental work has contributed significantly to the understanding of cell interactions in a variety of organs, while Zwilling concentrated on interactions occurring among cells of the limb.

|
Figure 15. Illustrations from a paper by R. G. Harrison (1921) on the establishment of symmetry within the embryo. Limb primordia were transplanted in various orientations to the flank region or to the normal position of the limb. This series of illustrations shows an embryo in which a left limb bud was implanted into the same location after being rotated 180°. Figures 49 & 50 are dorsal and lateral views at 4 days, 51 & 52 at 7 days, 53 & 54 at 10 days, and 55 a dorsal view at 17 days after the operation. Duplication of
distal limb parts occurred. |
Immigration and Embryological Research
Finally, an area of cell interaction that has had a tremendous influence on current embryological research is of recent origin. Clifford Grobstein (1953) demonstrated that the mesenchyme surrounding an epithelial organ plays an important role in controlling its shape. He used trypsin to separate the mesenchymal capsule from the epithelial surface of organ primordia and combined mesenchyme from one organ with epithelium of another. He showed that the initiation of shape changes in some organs depends upon a highly specific association with their own mesenchyme and that the mesenchymal influence can be transmitted through a semiporous filter. This research also contributed to the current active field of investigations on the roles of extracellular matrix, membrane receptors, and cytoplasmic filaments in changes in cell shape that occur during organ formation.
The scientific well being of the United States has been enhanced by continued immigration to this country. Agassiz came from Switzerland to settle at Harvard in 1847 and Loeb came from Germany to teach first at Bryn Mawr in 1891 and later at the University of Chicago and University of California. Although some immigration occurred during World War I, the greatest influx of scientists occurred with the rise of Nazism. Schotte, who was stranded as a student in Switzerland at the time of the Russian revolution, left Spemann's laboratory to be a Silliman Fellow at Yale under Harrison. In 1933 he obtained a teaching post at Amherst College. Other embryologists who came from Germany included D. Bodenstein, V. Hamburger, J. Holtfreter, R. Goldschmidt, P. Weiss, and Salome Waelsch. Migrations also occurred from Asia.
|
|
Figure 16. Illustration from Townes and Holtfreter (1955) demonstrating sorting of dissociated cells from different embryonic layers in amphibians. Cells from different germ layers sorted into clumps and tended to take a position resembling that of normal embryonic development. |
Dissemination of Information - Establishment of Societies and Journals
The expansion of biological research at various institutions in the United States beginning in the 1880's resulted in desires to communicate orally and in writing. At this time, the American Association for the Advancement of Science (AAAS) was the only national scientific society and was considered too broad by the biologists. Those interested in biology became served by a loose federation, the American Society of Naturalists, that separated from the AAAS. The Society of Naturalists of the Eastern United States was founded in 1883 and changed its name to the American Society of Naturalists in 1886. The motivating force was provided by A. Hyatt, who played a role in the founding of the Marine Biological Laboratory, and S.F. Clark of Williams College, who was among the first to receive a degree in the new Ph. D. program at Johns Hopkins. The society served as a source for the establishment of smaller societies that tended to develop along narrow disciplinary lines; those which had some interest in embryology or were founded by the new experimental embryologists included the Association of American Anatomists (later changed to the American Association of Anatomists) in 1888 and the American Morphological Society (later changed to the American Society of Zoologists) in 1890. The Association of American Anatomists was founded as an offshoot of the American Medical Association and at first included primarily teachers of anatomy; only around 1900 did membership become limited to trained investigators. Whitman provided the initiative for founding the American Morphological Society. These societies were not limited to embryology and it was not for many years that a society dedicated to the discipline was established.
The Society for the Study of Growth and Development, now named the Society for Developmental Biology, was founded in 1940 after a symposium on Growth and Development was held at Truro on Cape Cod in August, 1939. It was organized by the editors of Growth, a journal founded by the Lankenau Hospital Research Institute's Marine Research Station in Truro. The meeting generated enthusiasm for the establishment of a continuing symposium series sponsored by the newly organized society. The symposia were designed to allow much time for discussion in small, informal groups in addition to the discussion periods after formal lectures. This format was maintained for many years and the program was deliberately diverse, with speakers recruited from fields other than embryology where the work was related to development and growth. Speakers at the first symposium included Lewis, Schott, Glaser, and
Sinnott from the United States and Waddington, Needham, and Stern from overseas. The second symposium was concerned entirely with physical and chemical aspects of growth and development. The topics of the society continued to be at the forefront of embryological knowledge. This, plus the inclusion of speakers at the periphery of embryology and the encouragement of informal discussion, made the meetings an exciting time. The relatively small size and informal atmosphere also encouraged interactions between students and established investigators, adding to its value as a stimulator of research. The society celebrated its 50th anniversary in 1989.
A number of journals were founded in the same time period. Whitman founded the Journal of Morphology in 1887, which became unofficially affiliated with the Morphological Society. The American Journal of Anatomy ( 1901) and Anatomical Record (1906), both started with private funds, served the Anatomists. The Journal of Experimental Zoology was founded in 1904 by Harrison at Yale, and the Marine Biological Laboratory launched its own journal, the Biological Bulletin, in 1898. Prior to that, summer lectures at the laboratory were published in bound volumes, beginning in 1893. All of these journals exist today and publish papers on developmental biology.
Sources Of Research Funding
Funding of research by government agencies is a relatively recent development in American science. Europe has a much longer tradition of institutes established and funded by the State. In the 1800's most embryologists paid for their own research materials and attendance at research stations in the summer. Those who were lucky received support from their institutions. The first substantial grants were provided by private foundations, such as the Rockefeller Foundation and the Carnegie Foundation. The larger grants were made to institutions but some small grants also were awarded to individuals.
Support targeted for embryology through Federal grants began in 1952, with the establishment of the National Science Foundation (NSF). An average of 11 awards per year were made in the first four years, but the numbers climbed to average 100 in the period from 1961 to 1964. During that same time period the budget climbed from an average of approximately $98,200 in the first four years to an average of approximately $3,450,000 in the early 1960's. Currently the budget for Developmental Biology is approximately $18,000,000 for all programs with approximately 60 new grants funded each year at a cost of between $2.0 and 2.5 million. New grants represent approximately one third of the total number of research grants that are supported.
Funding of grants specifically in developmental biology started at the National Institutes of Health in 1966. In that year, the National Institute of Child Health and Human Development awarded approximately $3,400,000 to approximately 150 research grantees. A decade later, 246 research awards were made for approximately $13,400,000 while 371 awards were made in 1989 for $50,200,000. These figures represent new and continuing awards and are not comparable to the NSF figures. Over the years, the number of awards as well as the dollar amount of individual grants has increased dramatically. The increase in funding in more recent years reflects not only the rate of inflation but also the increased costs for the more sophisticated equipment that is used.
Figures

Fig 1. by N. Hartsoeker 1694

Fig 2. by M. Malpighi 1673
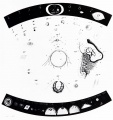
Fig 3. by C.E. von Baer 1827
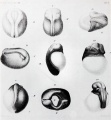
Fig 4. by W. Roux 1888

Fig 5. by H. Driesch 1892

Fig 6. Louis Agassiz

Fig 7. Leonard W. Williams c1900
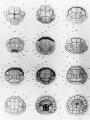
Fig 8. by Conklin 1905
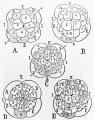
Fig 9. by Wilson 1892
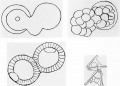
Fig 10. by Loeb 1893

Fig 11. by E. B. Wilson 1904
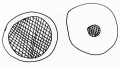
Fig 12. by O. E. Schotte
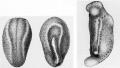
Fig 13. by Spemann and H. Mangold 1924

Fig 14. by S. Horstadius 1928

Fig 15. by R. G. Harrison 1921
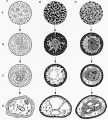
Fig 16. by Townes and Holtfreter 1955

{kind=link}
{kind=link}
Bibliography
General References
Goldschmidt, R.B. 1938 Physiological Genetics. McGraw-Hill, New York.
Horder, T.J., J.A. Witkowski, and C.C. Wylie, eds. 1986 A History of Embryology. Cambridge University Press, Cambridge. Contains papers by the editors and others on Harrison, Spemann, and Morgan as well as other interesting topics.
Just, E.E. 1939 The Biology of the Cell Surface. Blakiston, Philadelphia.
Maienschein,J. 1989 100 Years Exploring Life, 1888-1988, The Marine Biological Laboratory at Woods Hole. Jones and Bartlett, Boston.
Moore, J.A. 1987 "Science As A Way Of Knowing. IV. Developmental Biology." Am. Zool. 27:415-573.
Rainger, R, K.R Benson, and J.Maienschein, eds. 1988 The American Development of Biology. University of Pennsylvania Press. Chapters by Maienschein, Gilbert, Appel are especially pertinent.
Spemann, H. 1938 Embryonic Development and Induction. Yale University Press, New Haven. Reprinted by Hafner, New York, 1962.
Willier, B.F. and J. M. Oppenheimer 1974 Foundations of Experimental Embryology. Hafner, New York. Contains translated reprints of papers by Roux, Driesch, Child, Spemann & Mangold, Holtfreter, and others.
Works Cited
Agassiz, L. 1859 An Essay on Classification. Longmans et al., London. Reprinted by Harvard University Press, Cambridge, 1962.
Baer, C.E. von 1827 De ovi mammalium et hominis genesi. L. Vossius, Lipsiae.
Barth L.G. 1939 "The chemical nature of the amphibian organizer. III. Stimulation of the presumptive epidermis of Ambystoma by means of cell extracts and chemical substances." Physiol. Zool. 12:22-30.
Briggs, R. & T.J. King 1952 "Transplantation of living nuclei from blastula cells into enucleated frog's eggs." Proc. Natl. Acad. Sci. 38:455-463.
Child, CM. 1929 "The physiological gradients." Protoplasma 5:447-476.
Conklin, E.G. 1897 "The embryology of Crepidula, a contribution to the cell-lineage and early development of some marine gastropods." J. Morph. 13:1-226.
Conklin, E.G. 1905 "The organization and cell lineage of the ascidian egg." J. Acad. Nat. Sci. Phila. 2nd ser. 13:1-119.
Driesch, H. 1892 "Entwicklungsmechanische Studien. I." Zeit. Wissenschaft. Zool. 53:160-178.
Driesch, H. 1902 "Neue Erganzungen zur Entwickelungsphysiologie des Echinidenkeimes." Arch. Entwick. Organ. 14:500-531.
Driesch, H. & T.H. Morgan 1895 "Zur Analyse der ersten Entwicklungsstadien der Ctenophoreneies. I. Von der Entwickelung einzelner Ctenophorenblastomeren." Roux Arch. 2:204-215.
Grobstein, C. 1953 "Epithelio-mesenchymal specificity in the morphogenesis of mouse sub-mandibular rudiments in vitro." J. Exp. Zool. 124:383-414.
Harrison, RG. 1910 "The outgrowth of the nerve fiber as a mode of protoplasmic movement." J. Exp. Zool.
Harrison, R.G. 1921 "On the relations of symmetry in transplanted limbs." J. Exp. Zool. 32:1-136, p. 36. Hartsoeker, N. 1694 Essay de Dioptrique. Jean Anisson, Paris.
His, W. 1874 Unser Korperform und das Physiologische Problem ihrer Entstehung. Vogel Verlag, Leipzig.
Holtfreter, J. 1934 "Der Einfluss thermischer mechanischer und chemischer Eingriffe auf die Induzierfahigkeit von Triton-keimteilen." Roux Arch. 132:225-306.
Horstadius, S. 1928 "Uber die Determination des Keimes bei Echinodermen." Acta Zool. 9:1-191.
Jenkinson, J.W. 1909 Experimental Embryology. Clarendon, Oxford.
Loeb, J. 1893 "On some facts and principles of physiological morphology." In Biological Lectures Delivered at the Marine Biological Laboratory of Wood's Holl [sic] in the Summer Session of 1893, pp. 37-61. Ginn and Company, Boston.
Malphigi, M. 1673 Dissertatio epistolica de formatione pulli in ovo. Joannem Martyn, London.
O. Mangold, 1933 "Uber Induktionsfahigkeit der verschiedenen Bezir ke der Neuru la von Urode len." Naturwiss. 21:76 1-766.
McClendon, J. F. 1910 "The development of isolated blastomeres of the frog's egg." Am. J. Anat. 10:425-430.
Morgan, T.H. 1935 "The relation of genetics to physiology and medicine." Sci. Monthly 41:5-18.
Roux, W. 1888 "Beitrage zur Entwicklungsmechanik des Embryo." Arch. Path. Anat. Physiol. Klin. Med. 114:1 13-153.
Spemann, H. & H. Mangold 1924 "Uber Induktion von Embryonalanlagen durch Implantation artfremder Organisatoren." Arch. Mikroskop. Anat. Entwick. 100:599-638.
Spemann, H. & O.E. Schotte 1932 "Uber xenoplastische Transplantationals Mittel zur Analyse der embryonalen Induktion." Naturwiss. 20:463-467.
Spratt, N.T. 1946 "Formation of the primitive streak in the explanted chick blastoderm marked with carbon particles." J. Exp. Zool. 103:259-304.
Townes, P.L. & J. Holtfreter 1955 "Directed movements and selective adhesion of embryonic amphibian cells." J. Exp. Zool. 128:5 3-118.
Vogt, W. 1925 "Gestaltungsanalyse Amphibienkeim mit ortlicher Vitalfarbung. II. Teil. Gastrulation und mesodermbildung bei Urodelen und Anuren." Roux Arch. 120:384-706.
Waddington 1940 Organizers and Genes. Cambridge University Press, Cambridge.
Weiss, P. 1934 "In vitro experiments on the factors determining the course of the outgrowing nerve fiber." J. Exp. Zool. 68:393-448.
Whitman, CO. 1878 "Embryology of Clepsine". Quart. J. Micros. Sci. 18:215-315.
Wilson, E.B. 1892 "The cell-lineage of Nereis. A contribution to the cytogeny of the annelid body." J. Morph. 6:361-480.
Wilson 1893 "Amphioxus and the mosaic theory of development." J. Morph. 8:579-639.
Wilson, 1904 "Experimental studies on germinal localization. I. The germ-regions in the egg of Dentalium." J. Exp. Zool. 1:1-72.
Wilson 1925 The Cell in Development and Heredity. MacMillan, New York.
| Historic Disclaimer - information about historic embryology pages |
|---|
|
External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
- American Association for the Advancement of Science
- American Society of Naturalists
- Carnegie Foundation
- Journal of Experimental Zoology
- Marine Biological Laboratory
- National Science Foundation
- National Institute of Child Health and Human Development
- Society for Developmental Biology
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2024, April 27) Embryology Book - The Emergence of Experimental Embryology in the United States. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Book_-_The_Emergence_of_Experimental_Embryology_in_the_United_States
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G