Book - Human Embryology and Morphology 15
| Embryology - 28 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Keith A. Human Embryology and Morphology. (1902) London: Edward Arnold.
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
Chapter XV. The Brain and Spinal Cord
Formation of the Central Canal
The medullary plates of epiblast, which form the spinal cord and brain, rise up, meet, and enclose a canal — the central canal of the spinal cord and brain (Figs. 158 and 69, p. 90). The lips of the medullary plates meet and fuse together in the cervical region first (Fig. 1 5 8), the process of union spreading forwards and backwards, the last parts to be enclosed being the cephalic and caudal extremities. The optic vesicles start to grow out from the medullary plates before they have united to enclose the cavity of the fore-brain. The canal is completely closed by the middle of the 3rd week.

Fig. 158. Medullary Folds uniting to form the Neural Tube in a Human Embryo of about 14 days. (After Graf Spee.)
Divisions of the Neural Canal (Fig. 159).— At the end of the 3rd week the neural tube is divided into four parts. They are:
- An anterior dilatation, the fore-brain, which forms the rd and lateral ventricles and their walls.
- The mid-brain, which becomes transformed into the aquelct of Sylvius and crura cerebri.
- The hind-brain, the basis of the 4th ventricle, pons, cereilium and medulla.
- The central canal and spinal cord.

Fig. 159. Diagram of the Four Primary Divisions of the Neural Tube.
Cerebro-spinal fluid fills the canal. We do not know how it secreted or absorbed, but under certain abnormal conditions it y collect and give rise to a cystic condition of the neural tube. 3 expansion may involve the whole tube or only a part of it.
Chick embryos, hatched under abnormal conditions as regards temperature, frequently show a cystic condition of the neural tube, which is also accompanied by a dropsical state of the mesoblastic tissues. If the cystic condition occurs at an early stage it may dilate the fore-brain, or even the whole cephalic part of the tube until it bursts. This is probably the pathology of the condition seen in anencephalic foetuses, children born with all the parts of the body developed except the brain and roof of the skull, which are represented merely by a broken mass of tissue. A cystic condition of the lateral ventricles, which are formed as diverticula of the fore-brain, occasionally occurs towards the end of foetal life, and gives rise to the condition known as hydrocephalus.
Encephalocele and Spina-bifida
Localised dilatations of the neural tube may occur. The most common, spina bifida, occurs in the lumbar region, where the medullary plates are last to close. The dilatation may affect the arachnoid and dura mater only ; or the neural tube may be also distended. Another site is at the anterior or cephalic end of the tube, where the medullary plates are also late in closing. The encephalocele, or it may be only a meningocele, formed in this site, projects at the root of the nose or within the nasal cavity. A meningocele may also occur at the roof of the 4th ventricle ; it projects at the occipital fontanelle (Fig. 132, p. 166).
The Spinal Cord
The Spinal Cord at first extends the whole length of the spinal column. After the 4th month the spinal column and canal grow more rapidly than the cord, and at birth its lower end has become withdrawn to the level of the 3rd lumbar vertebra. By the third year it only reaches the disc between the 1st and 2nd lumbar vertebrae. The results of this inequality of growth, are — (1) The roots of the lumbar and sacral nerves become enormously elongated, forming the cauda equina ; all the nerves are more or less drawn up, except the 1st and 2nd cervical ; the origins of the lower cervical nerves are drawn up 2 vertebrae (as indicated by the position of their spines) ; the upper dorsal, 3 ; the lower dorsal, 4 ; the lower lumbar, 5 ; the coccygeal, 10. These statistics represent a broad expression of the observations made by Professor E. W. Reid. j .
(2) As the caudal termination of the neural canal is never parated from the epiblast over the coccyx, the posterior end of e cord with its pial covering (mesoblast) is pulled out into a le string — the filum terminate. The neural canal extends for me way into the filum terminale, and in the foetus shows there dilatation or ventricle. The arachnoid and dura mater, mesoastic sheaths of the cord, do not suffer in the retraction of the rd ; they remain widely open to the level of the 2nd sacral rtebra.
Neurenteric Canal. — The caudal ends of the medullary plates Id over and include within the neural canal the blastopore Fig. 158). The blastopore occurs at the anterior end of the primitive streak and marks the point at which the cavity of e hypoblast (that part which becomes the hind-gut) opens i the epiblast. There is thus set up a communication between e neural canal and the gut to which the name of neurenteric ,nal is given (see p. 121). The neurenteric canal also opens to the posterior end of the canal of the notochord.
Differentiation of the Spinal Part of the Neural Tube. — when the medullary plates close in about the 14th day to form the neural tube, they are composed of a layer of elongated epithelial piblastic) cells. By the 6th week these cells have • undergone ie following changes (Fig. 160) :
(1) Some become neuroglia, elongated and branched supporting cells ; in the outer zone of the tube wall they form a meshwork ; in the inner zone, round the central canal, they form the ependyma; in the middle zone their interstices are filled with neuroblasts.
(2) Some of the cells become differentiated into neuroblasts, These are produced from germinal cells lying in or near the inner zone (Fig. 160).

Fig. 160. Diagrammatic Section showing the three Zones of the Spinal Neural Tube at the Gth week.
Two points should be noted (Fig. 160) —
(a) That the cells in the roof and floor scarcely undergo any proliferation — they retain for a considerable time the simple embryonic form.
(b) That there are two lateral centres of proliferation — a dorsal centre or lamina, the cells of which are connected afterwards with the ingrowing fibres of the dorsal root, and a ventral centre or lamina, at which the cells of the anterior horn are produced and from which the fibres of the anterior root arise.

Fig. 161. Diagrammatic Section o£ the Spinal Cord to show the Parts formed in the three Zones of the Embryonic Spinal Cord.
The changes which occur in the Spinal Cord after the 6th week (see Figs. 160 and 161) are: A. In the Outer Neuroglial Zone :
- Postero-mesial and postero-lateral columns are produced the fibres of the posterior roots. They become myelinated out the 6 th month.
- To the inner side and to the outside of the anterior horn ires are produced from cells in the anterior horn, which conct together neighbouring spinal segments (association fibres).
- Two tracts of fibres grow out from cells in the grey matter the cord and go to the cerebellum. They are the direct cereliar tract (Flechsig's) from the cells of Clarke's column and Dwer's tract from cells at the base of the anterior horn.
- Fibres produced by the cells of the cerebellar cortex grow iwnwards outside the anterior horn (intermediate tract). All ese tracts are medullated before birth.
- The pyramidal tracts (crossed and direct) grow down from e cells of the motor cortex. They are not medullated until r e months after birth.
The white matter of the cord is thus entirely produced by the growth of nerve fibres within the neuroglial network of the iter zone. The cerebral motor cortex, through the pyramidal acts, comes to dominate the cord ; sensory and cerebellar paths e formed, and intercommunications set up between the spinal gments.
The Anterior and Posterior Median Fissures are produced by the owth of the white matter. It is possible that the posterior edian fissure is formed in part by the inclusion of the dorsal gment of the central canal of the neural tube.
B. The Middle Zone — filled with neuro-blasts, forms the horns id grey matter of the cord.
C. The Inner Zone becomes the ependyma which lines the ntral canal and the gelatinous tissue which surrounds it. ie columnar cells which line the central canal are ciliated. The ntral canal retains the embryonic calibre while the wall creases enormously in thickness.
The Hind Brain
That part of the neural tube which forms the hind brain (Fig. 159) becomes transformed into:
- The Medulla Oblongata.
- The Pons Varolii.
- The Cerebellum.
The Fourth Ventricle. — The cavity or neural canal of the hind brain becomes the fourth ventricle. In its floor are developed, out of the ventral and dorsal parts or laminae (Fig. 162) of the medullary plates, the pons and medulla. In its roof are developed the cerebellum, superior and inferior medullary vela.

Fig. 162. Section across the Hind Brain of a Human Embryo in the 5th week.
Inferior Medullary Velum. — When a section is made across the posterior half of the hind brain of a fifth week human embryo (Fig. 162), the same parts are seen as in a section of the cord. The only difference is that the roof plate, which in the cord is narrow, is here very wide and thin. It will also be noticed that each medullary plate, shows, as in the spinal cord, a dorsal or alar lamina and a ventral or basal one. The two laminae of each side meet at an angle. In the roof plate over the anterior half of the 4th ventricle is developed the cerebellum and superior medullary velum ; over the posterior half, the roof plate forms the inferior medullary velum.
As shown in Fig. 163, the velum is continuous with the cerebellum above and the roof of the central canal of the cord below. In the posterior margin of the cerebellar rudiment are developed : (1) the nodule, (2) the flocculus, (3) the peduncle of the flocculus between 1 and 2 (Fig. 164). Hence the inferior medullary velum ends above in these structures.
The obex and ligula, thickenings found on the margins of the lower angle of the 4th ventricle, mark the attachment of the roof plate or velum to the alar laminae of the medullary plates, hey represent the attached margin of the velum. The velum is Iso attached to the restiform body which is developed in the pper margin of the alar lamina.

Fig. 163. Lateral view of the Cephalic Part of the Neural Tube in a 5th week human embryo. (After His.)
The velum is to be regarded as a specialized part of the neural ibe, which has been arrested at an early embryonic stage and serialized for the secretion (and absorption ?) of cerebro-spinal uid. The foramen of Majendie and openings at the lateral scesses of the 4th ventricle, where the velum is produced into loroid villi or cornucopia, may be caused by the absorption of a art of the velum or in some cases they may be produced in the imoval of the brain from the skull. In the early embryonic mdition, at least, the ventricle is quite closed. As will be sen from Fig. 163, the neural tube is bent with its convexity irwards at the pons. His has suggested that this bend may ive something to do with the production of the wide roof plate ' the 4th ventricle, for if a piece of tubing be bent so, the part . the concavity of the bend becomes widened out.
Cerebellum. — The condition of the cerebellum in a 4th week jman foetus resembles that of the Frog (Figs. 165 and 166). . is then merely a thickened transverse band in the anterior irt of the roof plate of the 4th ventricle. The vermis or median be is the first part to be developed, the lateral lobes form in the .

Fig. 164. Diagram of the Attachments of the Inferior Medullary Velum in a 5th month foetus. (After Kollmann.)
Only in the higher primates are the lateral lobes well developed. In the 2nd and 3rd months the simple transverse roof plate is transformed by the outgrowth from its surface of five transverse ridges. As these ridges or foliae grow out (Fig. 166) the anterior two of them come to occupy the upper surface, while the three posterior ones are thrust to the lower surface. The folding of the simple cerebellar plate, so that it comes to present an upper and lower surface, is caused by its growth. The secondary and tertiary foliae are produced in the last three months of foetal life.

Fig. 165. Median Section of the Cerebellum and 4th Ventricle of a Frog.
The three Peduncles of the cerebellum are produced thus — probably during the latter half of intra-uterine life :
- The Superior Peduncles Fibres grow from the cells of the dentate nucleus of the cerebellum — probably also from the cortex — to the red nucleus and optic thalamus on the opposite side of the brain. Some of the ascending antero-lateral cerebellar fibres from the cord probably enter the cerebellum by the superior peduncles. The Superior Medullary Velum is part of the roof plate of the 4th ventricle which remains between the superior peduncles. The vestigial laminae which cover it form the lingula (Fig. 166).
- The Middle Peduncles are formed by processes which grow from the cortical cells of the cerebellum to the nuclei of the Pons and also by processes from the cells of the Pontine nuclei to the cerebellum. They are probably connected indirectly with the frontal lobes through the fronto-cerebellar fibres which lie in the inner third of the crusta.
- The Inferior Peduncles are formed by :
- Processes from the cerebellar cortex to the cord (descending cerebellar tract)
- Processes which end in the opposite olive ;
- Processes which grow in from Clarke's column (direct cerebellar) ;
- Processes from the cells of the sensory nuclei of the postero-mesial and postero-lateral tracts (nucleus gracilis et cuneatus).

Fig. 166. Diagrammatic Section of the Cerebellum of a 3rd month Human Foetus showing the folding of the Cerebellar Plate.
The central canal of the mid-brain forms the aqueduct of Sylvius (Fig. 167). In its roof are developed the corpora quadrigemina. The dorsal and ventral laminae of its medullary plates form the tegmentum and crusta of the crura cerebri.
The Three Neural Flexures (see Fig. 163). — The pontine flexure, a convexity forwards of the pons, has already been mentioned ; the nuchal flexure is concave forwards and occurs between the medulla and cord. Both of these are of small import, but the anterior flexure, whereby, in the third week of foetal life, the fore-brain is bent downwards and forwards until it comes to lie on the ventral aspect of the cephalic end of the notochord, leads to a great alteration in the form and relationships of the fore and mid brains and is of great importance. The mid-brain, by this flexure, is brought to be, for a short time, the most anterior part of the neural canal; the fore-brain is doubled back under the notochord. Eound the projecting end of the notochord — projecting between the mid and fore brains — is developed the posterior clinoid processes and dorsum sellae (Fig. 163). The tentorium cerebelli is developed between the mid-brain and fore-brain, and lies at first at right angles to the axis of the mid-brain, but the great subsequent growth of the cerebrum forces it backwards and downwards until it becomes a horizontal partition between the cerebral and cerebellar chambers of the skull.
The Fore-Brain
The Third Ventricle and Structures derived from its Walls. — The Third Ventricle (see Fig. 167) is the cavity of the forebrain and represents the anterior dilated end of the neural canal. From its walls many structures are derived. We have already seen that the optic vesicles are produced from its ventro-lateral walls; from its floor is produced the hypopophysis cerebri — the ancient mouth of the alimentary canal if G-askell's views are right. From the posterior part of its roof plate is produced the pineal body — an ancient median eye ; from its an tero- superior part is produced a bifid hollow outgrowth — the cerebral vesicles — which come in time to dominate the whole nervous system.
The Pituitary Body is formed from two elements (Fig. 167): (1) An epiblastic hollow bud from the stomodaeuni (Fig 22 p. 30). (2) A neuroblastic bud from the floor plate of the 3rd ventricle (fore-brain).

Fig. 167. A schematic figure to show the parts derived from the walls of the fore-brain. (After Wilhelm His (1831-1904))
The union of the two processes takes place at the anterior extremity of the notochord. The epiblast of the stomodaeum and the floor of the neural tube are in contact from the very beginning ; subsequently the mesoblast grows in between the forebrain and the epiblast of the stomodaeum, but the parts which form the pituitary adhere. The posterior or neuroblastic bud becomes solid ; its structure is that of neuroglia into which many vessels have grown carrying mesoblastic tissue with them.
The anterior or stomodaeal bud embraces the posterior. While its posterior wall remains quiescent, its anterior throws out solid processes between which a network of vessels lies. This forms the glandular part of the pituitary — reticulated rows of cells surrounding blood channels — similar in structure to the medullary part of the supra-renals, carotid bodies and parathyroids. The trabeculae cranii form round the pituitary. When the basi- and pre-sphenoids are developed in the trabeculae, the position of the stalk of the stomodaeal process is seen in the later months of foetal life between these two bones and forms the canalis craniopharyngeus (Fig. 3).
Pineal Body (see Fig. 167) grows as a hollow bud from the dorsal plate of the hinder part of the fore-brain during the 6 th week. In fossil reptiles and in some forms still living it forms a median eye which perforates and appears on the dorsum of the head between the parietal bones. It differs from the lateral eyes which grow from the third ventricle as the optic vesicles in this, that it produces the lens as well as the retina and optic stalk. The retina is inverted — i.e. the apices of the rods and cones point towards the vitreous chamber. The ganglion of the habenula, situated on the dorsal and inner aspect of the optic thalamus, represents its terminal ganglion. In man and mammals its development is arrested at an early stage. It produces a number of diverticula which are filled up by a proliferation of the cells which form the walls of the diverticula. In man it appears to be merely vestigial. It lies in the velum interpositum, which is forced down on it by the growth backwards of the cerebral hemispheres.

Fig 168. Transverse Section of the brain of a Human Foetus at the commencement of the 3rd month to show the Cerebral Vesicles overlapping the Thalamencephalon (schematic).
The Lamina Cinerea or lamina terminalis (Fig. 167) represents the anterior end of the neural tube. In the adult it stretches between the optic chiasma, which is developed on the floor of the 3rd ventricle and the rostrum of the corpus callosum. Its development will be described later, but it retains with little alteration its early simple structure. The inter-peduncular space, which forms the floor of the 3rd ventricle, also retains in the adult to a considerable extent the simple embryonic form. In it are developed the corpora albicantia and posterior perforated space.
The Optic Thalami are formed in the lateral walls of the forebrain (thalamencephalon) and in the adult occupy the whole extent of this wall. At first they are completely exposed on the outer aspect of the brain (see Fig. 154), but after the cerebral vesicles grow out from the antero-lateral angles of the fore-brain (3rd ventricle) they are prolonged backwards and downwards over the optic thalami and thus bury them (Figs. 173, 174, and 175). As may be seen from Fig. 169, the optic thalami, with the internal capsule, are simply the enlarged upward continuations of the tegmentum and crusta of the crura cerebri or mid-brain. From Fig. 167, it will be seen that the optic thalamus is an upward continuation of the alar lamina of the mid-brain, and the same may also be said of the caudate nucleus. The sulcus of Monro (Fig. 167), which runs from the opening of the aqueduct of Sylvius to the foramen of Munro, on the lateral wall of the 3rd ventricle, marks off the alar from the basal lamina of the thalamencephalon. A section across the 3rd ventricle and optic thalami, at the end of the second month is shown diagrammatically in Fig. 168. The internal capsule has not yet appeared. The cerebral vesicles already overlap and bury the optic thalami.

Fig. 169. Diagrammatic Section across the 3rd Ventricle of the Adult to show the ' Structures formed ill its Walls.
The Cerebral Hemispheres.
In the 3rd week very soon after the medullary plates have closed, a hollow bud, which almost immediately divides into a right and left half, grows out from the anterior superior extremity of the fore-brain. These two processes form the cerebral hemispheres. It will be seen that the cerebrum represents a super-addition to the neural-tube system. The lateral ventricles with their horns represent the cavity of the cerebral vesicles ; the foramina of Monro, which have a common entrance to the third ventricle, represent the position at which the primitive cerebral outgrowth took place. The walls of the vesicles thicken and form the mantle of the brain. The anterior horn represents the anterior extremity of the vesicle — the point at which the olfactory lobe is produced ; the descending horn represents the real posterior extremity of the vesicle ; the posterior horn, although found in all mammalian brains, is a later diverticulum formed in connection with the growth of the occipital lobe.
The primitive simple relationship of the cerebral vesicle which holds for low vertebrates (see Figs. 154 and 155) and for the first two months of foetal life, becomes obscured in the third by the vesicles growing over the optic thalami and burying them. The wall of the vesicle, which comes in contact with the optic thalamus, adheres to the outer surface of that body (Figs. 168 and 169). Hence the optic thalamus comes to form part of the floor of the body of the lateral ventricle and enters into the roof and inner wall of the descending horn.
Divisions of each Cerebral Vesicle. — A prolonged study of the adult vertebrate brain has led Elliot Smith to divide the wall of each cerebral vesicle into three primary parts :
- Rhinencephalon (defined on p. 22 ; see Figs. 18 and 172). It represents the oldest part of the cerebral vesicle and composes nearly the whole of the cerebrum of fishes, amphibians, reptiles, and birds.
- The Corpus Striatum.
- The Neo-pallium. This comprises the remainder of the cerebral vesicle. With the evolution of the higher mammals the neo-pallium became bigger and bigger until in man it constitutes by far the greater part of the cerebrum. "With its development the corpus striatum also became increased, while the rhinencephalon became more and more reduced.
The Velum Interpositum. — We have seen that the roof plate of the 4th ventricle (hind-brain) forms the cerebellum in front, while its posterior half becomes the inferior medullary velum— a secretory membrane. The roof plate of the third ventricle, from the foramina of Monro backwards, becomes modified in a similar manner. It merely forms the ependymal covering of the lower surface of the velum interpositum, also a secretory membrane (Fics. 168 and 169). The anterior part of the roof plate is produced into the cerebral vesicles over the foramina of Monro, and covers the apex of the velum interpositum (Fig. 170). The mesial wall of each cerebral vesicle from the foramen of Monro back to the posterior extremity of the vesicle (Fig. 170), which becomes the tip of the descending horn, is also inflected and becomes a secretory ependyma, covering the velum interpositum and choroid plexus within the lateral ventricles. Into this inflection of the embryonic neural wall spreads the mesoblast, carrying vessels with it. The velum interpositum is thus composed of a basis of mesoblast and its intraventricular parts have a covering of the ependyma of the neural wall.

Fig. 170. A dorsal view of the Fore and Mid-brain at the 5th week of development to show the formation of the Velum Interpositum. The Cerebral Vesicles are laid open and the inflection of their mesial walls shown on the ingrowing Velum. The Roof Plate of the 3rd Ventricle is also exposed. (Modified from His.)
The ependymal covering of the entire velum is derived from :
- The roof plate of the 3rd ventricle (lower surface) ;
- The roof plate of the foramen of Monro ;
- An inflection of the mesial wall of the cerebral vesicle. The choroid plexus, which fringes the velum in the adult,
completely fills the cavities of the lateral ventricles, which for the first five months are relatively very large and the containing walls thin. The velum and choroid plexus must play an important part in the development of the cerebral vesicle in the early period of growth. The spread of the vesicles backwards and downwards over the optic thalami (Fig. 173) obscures the original simple relationship of the velum to the brain ; but, when withdrawn from the transverse fissure, the velum is seen to rest on the optic thalami and project within the ventricle from the foramen of Monro to the tip of the descending horn, and that stretch marks the line at which the choroidal inflection took place.
The fibrous substance of the velum interpositum is continuous with the pial covering of the brain, and also with the edge of the tentorium cerebelli. The veins of Galen are developed in the velum and join the straight sinus in the tentorium. Pressure applied to the veins causes dropsy of the lateral ventricles.
Development of Commissures

(1) The Anterior Commissure; (Fig. 171) is developed in the lamina terminalis — the primitive anterior wall of the fore-brain. The commissure passes between the temporo-sphenoidal lobes. These lobes represent the posterior ends of the cerebral vesicles. At first they are mere dilatations behind the foramen of Monro. The commissure crosses in the lamina terminalis below and rather anterior to the foramen of Monro. This is the earliest and most primitive of the cerebral (pallia!) commissures (Elliot Smith).
(2) The Corpus Callosum, the great commissure between the cerebral hemispheres, forms late ; except in the higher mammals it is smaller in size than the anterior commissure. It reaches its fullest development in man, and in him it commences to form at the end of the third month.
To understand its development the student must be familiar with the mesial aspect of the brain during the third and fourth months. On this aspect he should note —
(1) The sulcus arcuatus (Fig. 171), which indents the mesial wall of the fore-brain. This fissure in the adult brain becomes (a) the callosal sulcus between the corpus callosum and callosal gyrus (gyrus fornicatus) (Fig. 172); (b) the hippocampal fissure, which indents the posterior extremity (descending horn) of the cerebral vesicle and causes the hippocampus major (Fig. 172). The calcarine fissure may also be a derivative of it (Elliot Smith).
(2) The Choroid or Transverse Fissure (Fig. 171), caused by the inflection of the mesial wall of the vesicle on the velum interpositum and choroid plexuses.

Fig. 172. Diagram to show the structures formed in tlie Lamina Terminalis and Primitive Callosal Gyrus. (After Elliot Smith.)
(3) The primitive callosal gyrus 1 (see Figs. 171 and 172) is that part of the mesial wall of the cerebral vesicle which lies between the arcuate and transverse (choroid) fissures. In the lower edge of this marginal gyrus, the edge which bounds the transverse fissure, and therefore overlies the velum interpositum, is developed the Fornix with the fimbria, its posterior continuation, a longitudinal commissure which connects the optic thalamus with the hippocampal (uncinate) gyrus. The corpus callosum is developed in the lamina terminalis above the foramen of Monro about the end of the 3rd month. It afterwards extends backwards, encroaching on and displacing the primitive callosal gyrus.
The grey matter of the primitive callosal gyrus becomes reduced to — (1) The vestigial supra-callosal gyrus, lying on the upper surface of the corpus callosum (Fig. 172); (2) the gyrus dentatus ; (3) the gyrus, which unites 1 and 2 round the splenium of the corpus callosum ; (4) the cortex of the hippocampus buried in the hippoeainpal fissure. The longitudinal •iae, the thin layer of grey matter on the upper surface of the rpus callosum, and the grey matter on the fornix, are also rived from the primitive callosal gyrus.
- 1 This gyrus forms part of the Rhinencephalon, and the name is proposed merely as a provisional one, until comparative anatomists agree as to its proper designation. At present, Elliot Smith proposes the term "hippocampal formations " for the parts of the brain derived from it in the adult.
(4) The lamina terminalis is seen in section. It is the rminal anterior wall of the fore-brain (Fig. 171). The lower part becomes the lamina cinerea ; in the upper part, the anterior mmissure is developed, and the anterior pillars of the fornix ig. 172). Its dorsal extremity bounds anteriorly the foramina Monro. It connects the mesial walls of the cerebral vesicles 'ig. 171), and becomes thickened and enlarged. In its dorsal ,rt, where it is continuous with the primitive callosal gyrus, e callosal commissure (corpus callosum) commences. The irt of the lamina terminalis which lies between the fornix id corpus callosum (Fig. 172) forms the septum lucidum. In is septum a cavity appears — the 5 th ventricle. Cases are lown of people with normally functional brains in which the rpus callosum was found afterwards to be absent.
The corpus callosum connects the cortex of one hemisphere ith the basal ganglia and cortex of the other. It is the mmissure of the neo-pallium. Its fibres are probably collaterals srived from the pyramidal and other fibres of the cortical Us. Each fibre grows out and crosses the great median sure of the brain in the lamina terminalis.
The Fornix is developed in the inner margin of the primitive llosal gyrus, which bounds the transverse or choroid fissure Fig. 171).
The Corpus Striatum. — As soon as the cerebral vesicle grows it, the corpus striatum appears as a thickening in its wall at e outer side of the foramen of Monro (Fig. 173). With the isterior development of the vesicle it comes to lie in the venieular floor, from the foramen of Monro to its posterior tremity.
The anterior extremity of the corpus striatum is continuous ith the olfactory lobe (Fig. 173). Hence in the adult brain e anterior end of the corpus striatum appears at the base of the ain in the anterior perforated space, a lamina of grey matter rived from the olfactory lobe (Fig. 18, p. 22). As the cerebral side grows over the optic thalamus (Fig. 173) the corpus striatum comes to be applied to that body. The posterior extremity of the cerebral vesicle, in which the tail of the caudate nucleus is situated (Fig. 173), develops downwards, behind and below the optic thalamus, forming the descending horn of the lateral ventricle. Hence the occurrence of the tail of the caudate nucleus and the amygdaloid nucleus on the roof of the descending horn, both being intrinsic parts of the corpus striatum. The corpus striatum is imperfectly separated, during the 4th and 5th months of foetal life, into two parts, the caudate and lenticular nuclei, by the downgrowth, from the cerebral cortex of the fibres which form the internal capsule, crusta, and pyramidal tracts.

Fig. 173. Showing the Development of the Corpus Striatum in the floor and outer wall of the Cerebral Vesicle.
Formation of the Island of Reil and Fissure of Sylvius. — The formation of the corpus striatum, the Island of Reil, and Fissure of Sylvius are part and parcel of the same process.
When the lateral wall of the cerebral vesicle is examined at the 5th month (Fig. 175) an area of cortex is seen to be rapidly becoming submerged by the overgrowth of the surrounding cortex. The submerged area is the Island of Reil ; it covers that part of the wall of the cerebral vesicle which is thickened by the corpus striatum (Fig. 168). The submerged area becomes triangular in shape, the apex being directed backwards ; it is bounded by three limiting sulci — an anterior, superior, and inferior. The rising lips of cortex, which bound the limiting sulci, form the temporal, fronto-parietal, and orbital opercula, and ultimately meet over the submerged area (Fig. 176). The fissure of Sylvius separates the opercula. It will be readily grasped that the development of the corpus striatum prevents the expansion of the insular part of the vesicle, whereas the thin-walled mantle, out of which the other lobes of the brain are developed, expands readily and overwhelms the thickened area. The corpus striatum begins to form during the 2nd month, hence as earJy as that date the insular depression is visible on the lateral wall of the hemisphere (Fig. 174).

Fig. 174. Lateral Aspect of the Cerebral Hemisphere during the 2nd month.

Fig. 175. The same Aspect during the 5th month.
The lower end of the stem of the Sylvian fissure also indents the Ehinencephalon, separating the uncinate gyrus from the parts derived from the olfactory lobe.
The student is already familiar with the fact that the Island of Reil forms a cortical cap to the corpus striatum. The structures between the islandic cortex and the foramen of Monro represent a section of the thickened wall of the cerebral vesicle (Fig. 169). Convolutions appear on it at the 7th month, when the rest of the cortex also becomes convoluted. Further, the larger the area of cerebral cortex in any primate, the larger is the Island of Reil; the more convoluted the cortex, the more convoluted the Island. Flechsig has shown that the cortex of the Island is joined to all the cortical areas of the mantle by bands of association fibres. Hence the Island must be regarded as playing a highly important part in co-ordinating the functions of the brain.

Fig. 176. The same Aspect during the 7th month.
The Opercula. — Three opercula grow up and cover the Island of Reil (see Figs. 175 and 176): (1) the temporal, (2) the fronto-parietal, (3) the orbital. Cunningham, whose researches into this region of the brain have become classical, found that during the later months (7-9) of foetal life, the orbital operculum in quite 50°/ o of brains shows a subdivision into two, an upper, the pars triangularis, and a lower, the pars orbitalis (Figs. 176 and 177). The subdivision occurs more frequently on the left side than on the right, probably owing to the centres for speech being situated on the left side. The temporal operculum rises first (5th month) the others a month later. The opercula which bound the posterior horizontal limb of the fissure of Sylvius are the first to come in contact. By the end of the first year after birth all three opercula meet over the Island and completely hide it. At birth there is still a part of the Island exposed behind the orbital operculum and in lower human races this is frequently the condition throughout life. The orbital opercula (pars triangularis and pars orbitalis) become part of the centre of speech and represent the latest additions to the human brain. If the orbital operculum is not subdivided, which is commonly the condition on the right hemisphere, then the anterior limb of the fissure of Sylvius is not subdivided into anterior horizontal and ascending parts (Fig. 177 A and B).

Fiq. 177. Diagram of the Opercula and Fissure of Sylvius. In A the orbital operculum is undivided ; in B it is subdivided. (After Cunningham.)
Comparative Anatomy of the Opercula and Island. — The Island of Reil and its opercula are only well developed in the higher primates. Figs. 178, 179 and 180 represent the stages in its evolution. In Fig. 178 the condition in dog-like apes is represented. Only the upper and lower limiting sulci of the Island are present. The Island, which is small, is continuous anteriorly with the frontal lobe. In anthropoids (gorilla, etc.) the Island is larger ; the upper and lower limiting sulci are present ; an imperfect anterior limiting sulcus (fronto-orbital fissure) is present and partially separates the Island from the orbital surface of the frontal lobe. In man all three limiting sulci are present and completely isolate the Island, and occasionally this is the condition (Fig. 179 E) in the higher anthropoids, but it is in man only that the orbital operculum grows up and meets with the other opercula. This can be the more easily understood when it is remembered that the orbital part of the 3rd frontal convolution is connected with speech.

Fig. 178. The Island of Reil and Fissures on the lateral Aspect of the Brain of a dog-like Ape.
Temporary Fissures. — In the 3rd month of foetal life the cerebral vesicles have thin walls, and when extracted from the head show several fissures. These fissures, unlike those which appear at the sixth and seventh months on the cortex of the brain, are really inflections of the whole thickness of the cerebral wall and are variable in number and position. It is possible that some of them are post-mortem products, for Hochstetter found that when the brain of a 3rd month foetus, which had been just removed from the mother, was hardened in situ, these temporary fissures were absent. But to this there are at least two exceptions. (1) The choroid fissure caused by the ingrowth of the velum interpositum and choroid plexus (Fig. 170).
(2) The arcuate fissure on the mesial aspect, which has been already described (Fig. 171). The hippocampal fissure, the callosal, and probably the calcarine too, are remnants of this fissure.

Fig. 179A. The more common Condition of the Island of Reil in Anthropoids.
Fig. 179B. The complete isolation of the Island of Reil, the condition seen constantly in the Human Brain and occasionally in the Anthropoid.
The Sylvian depression is also visible from the second month onwards, but it is in no sense a fissure comparable either to the temporary or even to the permanent.
The Fissures of the Brain. — The Principal Fissures of the Brain include: (1) The Sylvian, (2) the Calcarine, (3) the Hippocampal, (4) the 1st Temporal or Parallel, (5) the Parietooccipital, (6) Eolandic, (7) the Intra-parietal, (8) the Inferior Pre-central, (9) the Collateral, (10) the Calloso-marginal. These, with the exception of the first three (which have been already described), appear at the commencement of the 6 th month. At this date the human brain presents a marked resemblance in the arrangement of its fissures to that of the dog-like ape (Figs. 178 and 176).
The fissures and sulci are caused by a rapid increase in number and size of the cortical cells ; the increase of the area of the cortex leads to a crumpling up of its surface. The increased rate of growth appears to affect certain definite areas, hence the fairly constant forms into which the surface of the brain is thrown.
Affenspalte or Simian Fissure (Fig. 178). — In all ape brains the anterior margin of the occipital lobe grows upwards and forwards as an operculum, which covers the posterior margin of the parietal lobe. The sulcus between the occipital operculum and parietal lobe is the simian fissure or affenspalte. In the human brain it is never developed owing to the great growth of the posterior area of the parietal lobe ; the area which forms the floor of the sulcus in apes is spread out on the surface of the human brain. The ramus occipitalis of the intra-parietal fissure lies in the floor of the simian fissure ; in the human brain the ramus occipitalis is raised to the surface of the brain.
Sensori-motor Areas of the Brain. — The fissure of Eolando, which begins by two depressions — an upper and a lower — appears in the fifth month ; it divides the sensori-motor areas into anterior and posterior parts. Sherrington and Grunbaum found, however, in their experiments on the brains of anthropoids, that the posterior area gave no reaction when excited artificially, and that the fissure of Eolando formed the posterior boundary of the motor cortex. The anterior area is further subdivided by two fissures (Fig. 176), the inferior pre-central, an L-shaped fissure which commences as soon or even before the fissure of Eolando, and the superior pre-central, quite a late development, and evidently an isolated part of the superior frontal sulcus. The post-Eolandic area of the sensori-motor cortex is limited behind by the inferior and superior parts of the post-central fissure. The superior part is of late origin. The inferior limb of the post-central fissure is an intrinsic part of the intra-parietal fissure (Fig. 178). From the pyramidal cells of the sensori-motor cortex, processes grow out and form the middle part of the internal capsule. They reach the spinal cord during the 4 th and 5 th months of foetal life and become myelinated about the 5th and 6th months after birth. As the processes grow downwards in the lateral wall of the cerebral vesicle they pierce the corpus striatum, dividing it into the caudate and lenticular nuclei.
The Secondary Sulci, which divide the superior and middle frontal convolutions, the calloso-marginal, the parietal and occipital gyri, appear in the 9 th month. They are for the greater part peculiar to the human brain.
The Cranial Nerves
The differentiation of the simple neural tube of the embryo has thus far been followed into the complicated central nervous system of the adult. It is now necessary to make a short survey of the arrangement of the cranial nerves and see what evidence they afford of a segmental arrangement of the cephalic part of the neural tube.
The Cranial Nerves. — The segmental arrangement of the nerves of the body has been already discussed (page 158). Even although the human trunk is highly specialized the 33 or more segments of which it is made up can still be recognised from the arrangement of the spinal nerves ; each segment is constituted on a similar principle, and it becomes increasingly difficult to deny that man and the whole kingdom of vertebrates are derived from a form in which all the segments of the body were identical.
The head has become even more highly specialized than the trunk, and in it evidence of segmentation is accordingly more difficult to detect. Most of the evidence at present at our disposal indicates the presence of nine segments in the head. That is to say that the mammalian head is the derivative of a structure which was made up of nine segments, every one of which was originally constituted very much alike. Each had a similar arrangement of nerves and muscles, a similar arrangement of vessels, and provided with a similar pair of appendages.
In Fig. 180 is diagrammatised the relationship of the cranial nerves to the nine segments of the head. The olfactory and optic nerves arise as processes of the neural tube, and are not comparable to the remaining cranial nerves. Primarily each segmental cranial nerve appears to have contained sensory and motor fibres. The sensory fibres, like those of the spinal nerves, are developed from ganglionic cells derived from the neural crest, and are of two kinds, somatic and visceral. The motor fibres are developed from cells in the neural tube and are also of two kinds, somatic and visceral. In the evolution of the vertebrates there has been much reconstruction in the arrangement of the segmental fibres, the sensory fibres of several segments having become grouped together in the 5th nerve, and the motor fibres of others in such nerves as the 10th and 12th.

Fig. 180. A Diagram to show the Relationship of the Cranial Nerves to the Primitive Segments of the Head.
The Segments to which the Cranial Nerves belong.
1st Cranial Segment. — The motor nerve is the 3rd or oculo-motor. The ciliary ganglion, a derivative of the Gasserian, represents the sympathetic ganglion. Ganglion cells representing a vestigial posterior root may be found on the trunk of the nerve. The ophthalmic division of the 5th appears to represent its posterior or sensory root.
2nd Cranial Segment. — The motor nerve is the 4th. The sensory is represented by the superior maxillary division of the fifth. Meckel's ganglion represents the sympathetic ganglion. It is known to be derived from the same group of nerve cells as the Gasserian ganglion.
3rd Cranial Segment. — The motor nerve is the 6th and motor fibres of the fifth. The sensory root is represented by the inferior maxillary division of the 5th. The otic and submaxillary represent its sympathetic ganglia.
4th Cranial Segment. — The motor nerve is the 7th. The sensory root is represented by the chorda tympani and great superficial petrosal, which are developed from the geniculate ganglion (Dixon). The eighth nerve and its ganglia also belong to the sensory system of this segment. The great superficial petrosal represents a splanchnic nerve, the chorda tympani the nerve on the anterior margin of the 1st visceral cleft (see p. 34).
5th Cranial Segment. — The motor fibres of this segment have probably been scattered. Some may still remain in the 9th cranial nerve (glosso-pharyngeal) which is the chief nerve of the segment. The ganglia on the trunk of the glosso-pharyngeal represent the posterior root ganglion. The tympanic branch and small superficial petrosal represent an afferent (sensory) splanchnic branch.
6th, 7th, 8th and 9th Cranial Segments. — It has been already mentioned (pages 152 and 161) that the four posterior cranial segments are probably trunk segments which have become modified and added to the head. The anterior or motor nerve roots of these four segments are combined in the 12th nerve. Motor visceral fibres, which issue by the anterior roots of spinal nerves, here issue by the vagus and bulbar part of the spinal accessory (all of which are properly designated vagal fibres — Sherrington) and represent the visceral motor fibres of the four posterior cranial segments. The ganglia on the root and trunk of the vagus represent part of a posterior root ganglion. From these ganglia are developed the sensory visceral fibres connected with the fore gut and all the structures derived from the fore gut or splanchnopleure of the fore gut. A vestigial posterior root ganglion may occur on the 12th nerve.
The circuitous course of the spinal accessory is probably due to the migration of the trapezius caudalwards from a cephalic to its present position.
Chapter Figures
- Brain and Spinal Cord: Fig. 158. Human Neural Tube about 14 days | Fig. 159. Neural Tube Four Primary Divisions | Fig. 160. Zones of the Spinal Neural Tube Gth week | Fig. 161. Zones of the Embryonic Spinal Cord. | Fig. 162. Hind Brain 5th week | Fig. 163. Lateral Neural Tube 5th week | Fig. 164. Inferior Medullary Velum 5th month | Fig. 165. Frog Cerebellum and 4th Ventricle | Fig. 166. Human Cerebellum 3rd month | Fig. 167. walls of the fore-brain | Fig. 168. Human Cerebral Vesicles and Thalamencephalon 3rd month | Fig. 169. Adult 3rd Ventricle | Fig. 170. Fore and Mid-brain 5th week | Fig. 171. Fetal Brain 4th month | Fig. 172. Lamina Terminalis and Primitive Callosal Gyrus | Fig. 173. Corpus Striatum in Cerebral Vesicle | Fig. 174. Cerebral Hemisphere 2nd month | Fig. 175. Cerebral Hemisphere 5th month | Cerebral Hemisphere 7th month | Fiq. 177. Opercula and Fissure of Sylvius | Fig. 178. Ape Island of Reil and Fissures | Fig. 179A. Anthropoid Island of Reil | Fig. 180. Cranial Nerves and Primitive Segments | Figures
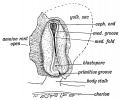
Fig. 158. Medullary Folds uniting to form the Neural Tube in a Human Embryo of about 14 days. (After Graf Spee.)
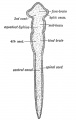
Fig. 159. Diagram of the Four Primary Divisions of the Neural Tube.
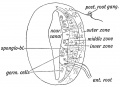
Fig. 160. Diagrammatic Section showing the three Zones of the Spinal Neural Tube at the Gth week.
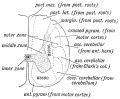
Fig. 161. Diagrammatic Section o£ the Spinal Cord to show the Parts formed in the three Zones of the Embryonic Spinal Cord.
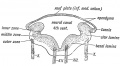
Fig. 162. Section across the Hind Brain of a Human Embryo in the 5th week.
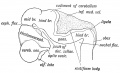
Fig. 163. Lateral view of the Cephalic Part of the Neural Tube in a 5th week human embryo. (After His.)
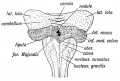
Fig. 164. Diagram of the Attachments of the Inferior Medullary Velum in a 5th month foetus. (After Kollmann.)
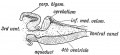
Fig. 165. Median Section of the Cerebellum and 4th Ventricle of a Frog.
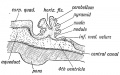
Fig. 166. Diagrammatic Section of the Cerebellum of a 3rd month Human Foetus showing the folding of the Cerebellar Plate.
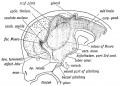
Fig. 167. A schematic figure to show the parts derived from the walls of the fore-brain. (After His.)
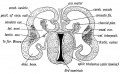
Fig. 168. Transverse Section of the brain of a Human Foetus at the commencement of the 3rd month to show the Cerebral Vesicles overlapping the Thalamencephalon.
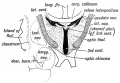
Fig. 169. Diagrammatic Section across the 3rd Ventricle of the Adult to show the Structures formed in its Walls.
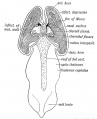
Fig. 170. A dorsal view of the Fore and Mid-brain at the 5th week of development to show the formation of the Velum Interpositum.
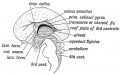
Fig. 171. Mesial Aspect of the human Foetal Brain during the 4th month. (After Minot.)
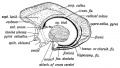
Fig. 172. Diagram to show the structures formed in the Lamina Terminalis and Primitive Callosal Gyrus. (After Elliot Smith.)
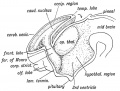
Fig. 173. Showing the Development of the Corpus Striatum in the floor and outer wall of the Cerebral Vesicle.
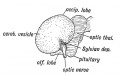
Fig. 174. Lateral Aspect of the Cerebral Hemisphere during the 2nd month.
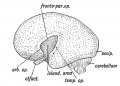
Fig. 175. The same Aspect during the 5th month.
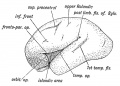
Fig. 176. The same Aspect during the 7th month.
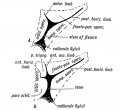
Fiq. 177. Diagram of the Opercula and Fissure of Sylvius. In A the orbital operculum is undivided ; in B it is subdivided. (After Cunningham.)
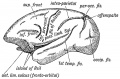
Fig. 178. The Island of Reil and Fissures on the lateral Aspect of the Brain of a dog-like Ape.
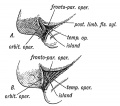
Fig. 179A. The more common Condition of the Island of Reil in Anthropoids. Fig. 179B. The complete isolation of the Island of Reil, the condition seen constantly in the Human Brain and occasionally in the Anthropoid.
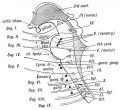
Fig. 180. A Diagram to show the Relationship of the Cranial Nerves to the Primitive Segments of the Head.
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Human Embryology and Morphology (1902): Development or the Face | The Nasal Cavities and Olfactory Structures | Development of the Pharynx and Neck | Development of the Organ of Hearing | Development and Morphology of the Teeth | The Skin and its Appendages | The Development of the Ovum of the Foetus from the Ovum of the Mother | The Manner in which a Connection is Established between the Foetus and Uterus | The Uro-genital System | Formation of the Pubo-femoral Region, Pelvic Floor and Fascia | The Spinal Column and Back | The Segmentation of the Body | The Cranium | Development of the Structures concerned in the Sense of Sight | The Brain and Spinal Cord | Development of the Circulatory System | The Respiratory System | The Organs of Digestion | The Body Wall, Ribs, and Sternum | The Limbs | Figures | Embryology History
Reference
Keith A. Human Embryology and Morphology. (1902) London: Edward Arnold.
Cite this page: Hill, M.A. (2024, April 28) Embryology Book - Human Embryology and Morphology 15. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Book_-_Human_Embryology_and_Morphology_15
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G