Paper - A presomite human embryo showing an early stage of the primitive streak
| Embryology - 26 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Lewis BV. and Harrison RG. A presomite human embryo showing an early stage of the primitive streak. (1953) J Anat. 87(2):124-9. PMID: 13044724
| Online Editor |
|---|
 Embryo Liverpool II was later described in - Lewis BV. and Harrison RG. A presomite human embryo showing an early stage of the primitive streak. (1953) J Anat. 87(2):124-9. PMID: 13044724 |
| Historic Disclaimer - information about historic embryology pages |
|---|
|
A Presomite Human Embryo showing an early stage of the Primitive Streak
By R. G. Harrison And T. N. A. Jeffcoate
Departments of Anatomy and Obstetrics and Gynaecology, University of Liverpool
Introduction
Although many human embryos of a presumed age of 15-17 days showing early stages of the primitive streak have been reported in the literat11re, most of them show specific characteristics (compare, for example, Mollendorff (1925) and Florian & Hill (1935)), while others have suffered distortion during fixation and preparation for histological examination (Bryce, 1925). When an embryo of this age group in a moderately well-preserved state was discovered by one of us (T.N.A.J.), it was decided to describe it as a contribution to the literature on such embryos. We propose to designate the specimen ‘Liverpool I’.
Clinical History of the Embryo
The embryo was obtained from a woman aged 37 years, married 12 years, who has four children, the youngest being 2 years of age. She has also had two abortions, the last one occurring in 1942. In June 1951, she sought medical advice on account of epimenorrhagia, her menstrual cycle for the previous few months having consisted of rather heavy bleeding lasting 7-8 days and commencing every 21-28 days.
Uterine curettage was performed on 21 June 1951, and the embryo found during routine histological examination of the curettings. The last menstrual flow before operation commenced on 24 May 1951 and was more normal than previously, lasting only 5 days; coitus occurred on 1, 5 and 13 (or 14) June. From the histological features of the embryo, it appears most likely that conception occurred on 5 June. Curettage was carried out on the day when menstruation might have been expected to begin, and the evidence suggests that conception took place 16 days previously, that is, on the twelfth day of the cycle. The curettings were fixed in Bouin’s fluid, and one edge of the blastocyst was discovered in the first section cut and examined. Fortunately, the embryo itself was intact, and 7 ,u sections of the remainder of the parafiin block were taken and stained with Ehrlich’s haematoxylin and eosin. The embryo is found in twenty-three sections (section numbers 24-46), the sections being transverse to the antero-posterior axis of the embryo at an angle of 32° from the vertical.
The Blastocyst
It is only possible to measure the dimensions of the chorion in two planes, the external dimensions (including trophoblastic villi) being 1-86 x 1-47 mm., and the internal dimensions of the cavity 1-50 x 0-84 mm. The blastocyst forms an elevation on the decidua, and a closing plug consisting of fibrin whose maximum diameter is 1-05 mm. is present. The villi consist almost entirely of cytotrophoblast, with very little syncytiotrophoblast, and well-marked cores of chorionic (extraembryonic) mesoderm; they are only beginning to show evidence of branching (Pl. 1, fig. 1), and are more well-developed at the embryonic pole of the blastocyst. Lacunae have formed and the syncytiotrophoblast is most clearly seen in relation to their walls. The extraembryonic mesoderm lines the trophoblast and forms a connecting stalk at the caudal end of the embryo (Text-fig.‘ 1) where it is clearly continuous with cells being formed from the primitive streak. Mitotic activity is present in all tissues of the chorion.

Text-fig. 1. A drawing of a reconstruction model of the embryo. Note the slight unevenness of the inner surface of both anmiotic cavity and yolk-sac. The line AB on the figure indicates the plane of sectioning the embryo.
The Embryo
Yolk-sac and amniotic cavity are present and the embryonic disc is bilaminar, except at the caudal end of the embryo where cells produced from the primitive streak are interposed between ectoderm and endoderm. A reconstruction model of the embryo was made at a magnification of 680 x from T3; in. thick Perspex. By direct measurement, and by estimation from the reconstruction model, the size of the embryonic disc is found to be 0-161 x 0-199 x 0-033 mm., the disc being wider in its transverse diameter. The volume of the amniotic cavity is slightly larger than that of the yolk-sac, and a smaH allanto-enteric diverticulum, 0-021 mm. in length, is present.
At the anterior end of the embryonic plate one unusual feature is the presence of a thickening of the endoderm, bulging into the cavity of the yolk-sac (Text-fig. 1, and P1. 1, fig. 2); this appearance, for which it is difficult to provide an explanation, is seen in four sections. Owing to the oblique sectioning of the embryo, the amniotic cavity is not seen until the sixth section (Pl. 1, fig. 3). In this section and the next succeeding one, extraembryonic mesoderm cells are arranged over the apex of the amniotic sac to simulate an amniotic duct which, however, has no communication with the amniotic cavity itself. The endodermal cells of the embryonic disc at this level are arranged quite irregularly, in sharp contrast to the ectodermal cells which have a clearly marked basement membrane. More posteriorly, at the ninth section, the width of the endoderm in the embryonic disc is clearly less than that of the ectoderm (Pl. 2, fig. 1); the transition from the tall columnar cells of disc ectoderm to the flattened cells of amniotic ectoderm is quite abrupt at this level, and the extraembryonic mesoderm is beginning to increase in quantity around the yolk-sac from the single layer of cells seen more anteriorly. In the eleventh section (Pl. 2, fig. 2) there are six layers of extraembryonic mesoderm cells in places at the lateral margins of the yolk-sac, which itself is smaller in size. In the succeeding section (Pl. 2, fig. 3) the yolk-sac can be seen to have narrowed considerably from side to side, so that the area of contact of endoderm with ectoderm is reduced to about one-fifth of the area of the latter. The yolk-sac proper is now restricted to the upper part of the endodermal cavity; the lower part of this cavity is continuous with the allantoenteric diverticulum (Pl. 2, fig. 4) which continues through the next three sections embedded in the connecting stalk mesoderm.
By the sixteenth section no evidence of endodermal tissue is to be found, but interesting changes can be observed in the ectoderm of the embryonic disc, which are even more pronounced in the following section (Pl. 2, fig. 5); the basement membrane is indistinct, particularly in the axial region of the disc, and cells are passing from the ectoderm into the mesodermal tissue of the connecting stalk. There is also evidence of considerable cellular activity in this region, as shown by the more irregular arrangement of cells and occasional mitotic figures. This appearance is also seen in the eighteenth section (Pl. 2, fig. 6), and extends therefore over three sections (0-021 mm.). These histological features are taken to represent an early stage of the primitive streak.
The remainder of the sections show clearly the major part of the connecting stalk between embryo and chorion, and the gradual diminution in size of the amniotic cavity.
Discussion
Of the already reported embryos of a similar age and stage of development, this specimen most resembles the embryo of Schlagenhaufer & Verocay (1916), the OP (Mollendorff, 1921) and W0 (Mollendorff, 1925) embryos, the BiI (Florian, 1927)
embryo and Edwards-Jones-Brewer ovum (Brewer, 1938). All of these specimens have measurements very similar to those of our embryo, and have many other features in common. The embryo of Schlagenhaufer & Verocay (1916), for example, possesses an allanto—enteric diverticulum but no amniotic duct; there is no primitive streak, however. The primitive streak in M6llendorfl"s (1921) OP embryo is longer (50 ,u), and the allanto-enteric diverticulum more well—developed, while Mt')llendorff’s (1925) W0 embryo is younger in having a solid allanto-enteric diverticulum and no primitive streak, although the measurements of the embryonic disc in the W0 embryo are slightly larger than in our specimen. Florian’s (1927) Bil embryo exhibits similar histological features, but is slightly larger in size, and possesses an amniotic duct. The Edwards-Jones-Brewer ovum has a primitive streak which is larger (40 p), and no allanto-enteric diverticulum, but the embryonic disc is almost the same size as in our specimen, and the endodermal cells similarly do not extend to the margins of the disc. The histological features of our specimen also resemble those of the Carnegie 7801 embryo (Heuser, Rock & Hertig, 1945), except for the greater degree of histological development of the primitive streak in the latter.
The primitive streak in this embryo occupies an area at the extreme posterior end of the ectoderm of the embryonic plate (Text-fig. 1). Immediately ventral to this is a collection of cells’ which shows characteristics not previously reported in other embryos of a similar age, and which may therefore indicate some abnormality. Although the allanto-enteric diverticulum is enclosed by these cells, which must consequently be regarded as belonging to the connecting stalk, at least in part, the transition between them and the tissue of the connecting stalk proper at the posterior end of the embryo is abrupt (compare, for example, the disposition of these cells in P1. 2, fig. 5 and the disposition of cells in the connecting stalk proper in P1. 2, fig. 6), since the cells in the latter structure are much less compactly arranged. This suggests that the origin of the mass of cells ventral to the primitive streak is not from extraembryonic mesoderm, but from primitive streak, since cells are actively proliferating from the ventral aspect of this structure. The unusual feature of this mass of cells, however, is that it extends to a level anterior to the primitive streak, ventral to a part of the ectoderm which has an intact basement membrane (Pl. 2, fig. 4). The cells in this region are presumably mainly of primitive streak origin, although they are continuous still further anteriorly with extra-embryonic mesoderm cells at the lateral margins of the yolk-sac (Pl. 2, figs. 2 and 3). If this embryo is normal, therefore, the condition represented may be an early stage of the primitive streak, which has differentiated in a small area at the caudal end of the embryonic plate, and is proliferating cells anteriorly, ventral to a part of the ectoderm with an intact basement membrane which will also eventually form the definitive primitive streak, before this structure is able to grow in length by the addition of new ectodermal cells at its caudal end.
There are signs in this specimen that the yolk-sac may not be quite normal. Although the shape of this structure does not exactly conform to that in the Carnegie 7801 embryo, it is very similar to the yolk-sac in the BiI embryo. There is also some evidence in the more anterior sections (as shown in P1. 1, fig. 3) of a shrinkage of endodermal cells of the embryonic plate, so that they are becoming detached from overlying ectoderm.
The characteristics of this specimen are therefore quite specific as regards size and degree of development. Its resemblance, in many features, to previously reported embryos, confirms the age of 16 days estimated from the clinical history.
Summary
A presomite human embryo of an estimated age of 16 days has been recovered from uterine curettings. It shows an early stage of the primitive streak, which is 21 /1. in length. The embryo also has many features previously reported in embryos of a similar age.
This work was aided by a grant from the Medical Research Council. We wish to thank Mr L. G. Cooper for his technical assistance.
References
Brewer JI. A human embryo in the bilaminar blastodisc stage (The Edwards-Jones-Brewer ovum). (1938) Contrib. Embryol., Carnegie Inst. Wash. Publ. 496, 27: 85-93.
Bryce TH. Observations on the early development of the human embryo. (1925) Trans. Roy. Soc. Edinb. 53, 533-567.
Florian J. Uber zwei junge menschliche Embryonen. (1927) Anat. Anz. Erg. 63: 184-192.
Florian J. & Hill JP. (1985). An early human embryo (No. 1285, Manchester Collection), with capsular attachment of the connecting stalk. J. Anat., Lond., 69, 399-411.
Hausan, C. H., Rocx, J. & Hanna, A. T. (1945). Two human embryos showing early stages of the definitive yolk sac. Contr. Embryol. Cameg. Instn, 31, 85-99.
M6u.ENDonFF, W. VON (1921). Uber einem jungen, operativ gewonnen menschlichen Keim (Ei OP). Z. ‘ges. Anat. 1. Z. Anal. EntwGesch. 62, 406-432.
NIOLLENDORFF, W. VON (1925). Das menschliche Ei WO(lf1-ing). Implantation, Verschluss der Implantationsoffnung und Keirnesentwicklung beim Menschen vor Bildung des Primitivstreifens. Z. ges. Anat. 1. Z. Anal. Em-wGesch. 76, 16-42.
SCHLAGENHAUFER, F. & VEROCAY, F. (1916). Ein junges menschliches Ei. Arch. Gynaelc. 105, 151-168.
Explanation of Plates
Plate 1

Fig. 1. A low-power photomicrograph of the blastocyst (section 35). The chorionic villi are only beginning to divide, and the lacunae are well developed. The closing plug is clearly shown in the lower part of the figure. ( x 65.)
Fig. 2. A section (number 27) of the anterior end of the embryo, showing the thickening of the endoderm (at a in the figure) bulging into the cavity of the yolk-sac. Since the embryo was cut obliquely, only the disc ectoderm and not the amniotic cavity is seen in this section. Part of the wall of the yolk-sac was torn during histological sectioning (just below the pointer in the figure), and was displaced into the blastocyst cavity. ( x 180.)
Fig. 3. Photomicrograph of section 29, showing the amniotic cavity and the arrangement of extraembryonic mesoderm cells over its apex to simulate an amniotic duct (s.a.d.) which does not communicate with the amniotic cavity. ( x 180.)
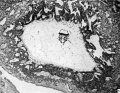
Fig. 1.
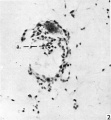
Fig. 2.
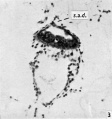
Fig. 3.



Plate 2
(All figures are at a magnification of x 180.)

Fig. 1. The embryo at a more posterior level (section 32) showing the diminution in width of disc endoderm. In this, and the next four succeeding figures, the transition from the taH columnar cells of disc ectoderm to the flattened amniotic ectodermal cells is clearly marked.
Fig. 2. Photomicrograph of section 34 showing the increased number of extraembryonic mesoderm cells at the lateral margins of the yolk-sac. The ectodermal cells in the centre of the disc are slightly irregular, simulating the formation of neural folds.
Fig. 3. The narrowing of the yolk-sac in its transverse diameter as seen in section 35. There is also an indication of the division of the yolk-sac into yolk-sac proper, and the allanto-enteric diverticulum (in the lower part of the endodermal cavity).
Fig. 4. Photomicrograph of section 36 to show the allanto-enteric diverticulum (a.e.d. in the figure) embedded in connecting stalk mesoderm.
Fig. 5. The region of the embryo (section 40) in which the disc ectoderm cells are clearly dividing (the primitive streak; 12.3. in figure) to contribute cells to the connecting stalk. A mitotic figure can be seen in this region.
Fig. 6. Section 42 of the embryo at the posterior end of the primitive streak in which there is no clear distinction between ectodermal cells and cells of the connecting stalk. No endodermal tissue is present at this level, and only the amniotic cavity can be seen. The part of the connecting stalk between embryo and chorion is seen in the top right-hand corner of the figure.
Cite this page: Hill, M.A. (2024, April 26) Embryology Paper - A presomite human embryo showing an early stage of the primitive streak. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_A_presomite_human_embryo_showing_an_early_stage_of_the_primitive_streak
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G