|
|
| (40 intermediate revisions by the same user not shown) |
| Line 1: |
Line 1: |
| {{Arey1924 Header}} | | {{Arey1924 Header}} |
|
| |
|
| | [[File:Arey1924 Title-page.jpg|thumb]] |
| =Developmental Anatomy - A Text-Book And Laboratory Manual Of Embryology= | | =Developmental Anatomy - A Text-Book And Laboratory Manual Of Embryology= |
|
| |
|
| Line 6: |
Line 7: |
| By | | By |
|
| |
|
| Leslie Bleainerd Arey | | Leslie Brainerd Arey (1891-1988) |
|
| |
|
|
| |
|
| Professor Of Anatomy At The Northwestern University Medical School, Chicago . | | Professor Of Anatomy At The Northwestern University Medical School, Chicago. |
|
| |
|
|
| |
|
| Line 24: |
Line 25: |
|
| |
|
| ==Preface== | | ==Preface== |
| | {| width=100%| |
| | | This book has been prepared for the use of medical students and others whose interests center primarily on man and mammals. The emphasizing of structural rather than functional aspects of Embryology is reflected in the title; such presentation is consistent both with the practical demands of modern courses and with the meagre information existant as to the physiological factors in developmcnt. |
|
| |
|
| This book has been prepared for the use of medical students and others whose interests center primarily on man and mammals. The emphasizing of structural rather than functional aspects of Embryology is reflected in the title; such presentation is consistent both with the practical demands of modern courses and with the meagre information existant as to the physiological factors in developmcnt.
| |
|
| |
|
|
| |
|
| The volume contains three sections. In the first i>art the early stages are treated comparatively and the fulCcourse of prenatal and postnatal development is outlined. The second section traces the origin and differentiation of the human organ-systems, grouped according to their germlayer derivations. The third division comprises a laboratory manual for the study of chick and pig embryos. | | The volume contains three sections. In the first part the early stages are treated comparatively and the full course of prenatal and postnatal development is outlined. The second section traces the origin and differentiation of the human organ-systems, grouped according to their germlayer derivations. The third division comprises a laboratory manual for the study of chick and pig embryos. |
| | |
|
| |
|
|
| |
|
| Many illustrations are from the earlier Prentiss-Arey text and discontinuous fragments of description have likewise been retained. Yet, in plan and content the work is essentially new. It is hoped that the developmental story has been told in an orderly and clear, but concise fashion, and that it records accurately the present state of the subject. | | Many illustrations are from the earlier Prentiss-Arey text and discontinuous fragments of description have likewise been retained. Yet, in plan and content the work is essentially new. It is hoped that the developmental story has been told in an orderly and clear, but concise fashion, and that it records accurately the present state of the subject. |
| | |
| | |
| | |
|
| |
|
|
| |
|
| Line 37: |
Line 43: |
|
| |
|
| Chicago, ill., September, 1924. | | Chicago, ill., September, 1924. |
| | | [[File:Leslie Arey.jpg|thumb|Leslie Brainerd Arey (1891-1988)]] |
| | |} |
|
| |
|
| ==Contents== | | ==Contents== |
| | | {| |
| PART I. GENERAL DEVELOPMENT | | | valign=top|PART I. GENERAL DEVELOPMENT |
| | | * Introduction |
| Introduction | | * General Features of Development |
| | | * Fundamental Conceptions |
| General Features of Development | | * The Vertebrate Groups |
| | | * Titles for Collateral Reading and Reference |
| Fundamental Conceptions | | * [[Book - Developmental Anatomy 1924-1|Chapter I. - The Germ Cells and Fertilization]] |
| | | ** The Germ Cells |
| The Vertebrate Groups | | ** Spermatogenesis, Oogenesis and Maturation |
| | | ** Ovulation and Insemination |
| Titles for Collateral Reading and Reference | | ** Fertilization |
| | | ** Heredity and Sex |
| [[Book - Developmental Anatomy 1924-1|Chapter I. - The Germ Cells and Fertilization]] | | * [[Book - Developmental Anatomy 1924-2|Chapter II. - Cleavage and the Origin of the Germ Layers]] |
| | | ** Cleavage |
| The Germ Cells | | ** The formation of Ectoderm and Entoderm (Gastrulation) |
| | | ** Origin of the Mesoderm, Notochord and Neural Tube |
| Spermatogenesis, Oogenesis and Maturation | | * [[Book - Developmental Anatomy 1924-3|Chapter III. - Implantation and Fetal Membranes]] |
| | | ** The Fetal Membranes of Reptiles and Birds |
| Ovulation and Insemination | | ** The Fetal Membranes of Mammals |
| | | ** The Fetal Membranes of Man |
| Fertilization | | ** Implantation and Early Mucosal Relations |
| | | ** The Decidual Membranes |
| Heredity and Sex | | ** The Placenta |
| | | ** Parturition |
| [[Book - Developmental Anatomy 1924-2|Chapter II. - Cleavage and the Origin of the Germ Layers]] | | * [[Book - Developmental Anatomy 1924-4|Chapter IV. - Age, Body Form and Growth Changes]] |
| | | ** Age, Size and Weight of Embryos |
| Cleavage | | ** An Outline of Prenatal Development |
| | | ** The Establishment of External Form |
| The formation of Ectoderm and Entoderm (Gastrulation) | | ** Growth Changes |
| | | | valign=top|PART II. ORGANOGENESIS |
| Origin of the Mesoderm, Notochord and Neural Tube | |
| | |
| [[Book - Developmental Anatomy 1924-3|Chapter III. - Implantation and Fetal Membranes]] | |
| | |
| The Fetal Membranes of Reptiles and Birds | |
| | |
| The Fetal Membranes of Mammals | |
| | |
| The Fetal Membranes of Man | |
| | |
| Implantation and Early Mucosal Relations | |
| | |
| The Decidual Membranes | |
| | |
| The Placenta | |
| | |
| Parturition | |
| | |
| [[Book - Developmental Anatomy 1924-4|Chapter IV. - Age, Body Form and Growth Changes]] | |
| | |
| Age, Size and Weight of Embryos | |
| | |
| An Outline of Prenatal Development | |
| | |
| The Establishment of External Form | |
| | |
| Growth Changes | |
| | |
| | |
| PART IV. ORGANOGENESIS | |
|
| |
|
| Entodermal Derivatives | | Entodermal Derivatives |
| | * [[Book - Developmental Anatomy 1924-5|Chapter V. - The Digestive System]] |
| | ** The Mouth |
| | ** The Pharynx |
| | ** The Digestive Tube |
| | ** The Liver |
| | ** The Pancreas |
| | * [[Book - Developmental Anatomy 1924-6|Chapter VI. - The Respiratory System]] |
| | ** The Larynx |
| | ** The Trachea |
| | ** The Lungs |
|
| |
|
| [[Book - Developmental Anatomy 1924-5|Chapter V. - The Digestive System]]
| | Mesodermal Derivatives |
| | | * [[Book - Developmental Anatomy 1924-7|Chapter VII. - The Mesenteries and Coelom]] |
| The Mouth
| | ** The Mesenteries |
| | | ** The Primitive Mesentery |
| The Pharynx
| | ** Differentiation of the Dorsal Mesentery |
| | | ** Differentiation of the Ventral Mesentery |
| The Digestive Tube
| | ** The Ccelom |
| | | ** The Primitive Coelom |
| The Liver
| | ** The Septum TransveRsum |
| | | ** The Pleuro-pcricardial and Pleuro-peritoncal Membranes |
| The Pancreas
| | ** The Pericardium and Diaphragm |
| | | * [[Book - Developmental Anatomy 1924-8|Chapter VIII. - The Urogenital System]] |
| [[Book - Developmental Anatomy 1924-6|Chapter VI. - The Respiratory System]]
| | ** The Urinary Organs |
| | | ** The Pronephros |
| The Larynx
| | ** The Mesonephros |
| | | ** The Metanephros |
| The Trachea
| | ** Differentiation of the Cloaca |
| | | ** The Genital Organs |
| The Lungs
| | ** Indifferent Stage |
| | | ** Internal Sexual Transformations |
| MeSodermal Derivatives
| | ** The External Genitalia |
| | | ** Homologies of Internal and External Genitalia |
| [[Book - Developmental Anatomy 1924-7|Chapter VII. - The Mesenteries and Coelom]] | | * [[Book - Developmental Anatomy 1924-9|Chapter IX. - The Vascular System]] |
| | | ** Origin of the Blood Vessels and Blood Cells |
| | | ** Hemopoiesis |
| The Mesenteries | | ** Development of the Heart |
| | | ** The Primitive Vascular System |
| The Primitive Mesentery | | ** Development of the Arteries |
| | | ** Development of the Veins |
| Differentiation of the Dorsal Mesentery | | ** Fetal Circulation and the Changes at Birth |
| | | ** The Lymphatic System |
| Differentiation of the Ventral Mesentery | | | valign=top| |
| | |
| The Ccelom | |
| | |
| The Primitive Coelom | |
| | |
| The Septum TransveRsum | |
| | |
| The Pleuro-pcricardial and Pleuro-peritoncal Membranes | |
| | |
| The Pericardium and Diaphragm | |
| | |
| [[Book - Developmental Anatomy 1924-8|Chapter VIII. - The Urogenital System]] | |
| | |
| The Urinary Organs | |
|
| |
| The Pronephros | |
| | |
| The Mesonephros | |
| | |
| The Metanephros | |
| | |
| Differentiation of the Cloaca | |
| | |
| The Genital Organs | |
| | |
| Indifferent Stage | |
| | |
| Internal Sexual Transformations | |
| | |
| The External Genitalia | |
| | |
| Homologies of Internal and External Genitalia | |
| | |
| | |
| [[Book - Developmental Anatomy 1924-9|Chapter IX. - The Vascular System]] | |
| | |
| Origin of the Blood Vessels and Blood Cells | |
| | |
| Hemopoiesis | |
| | |
| Development of the Heart | |
| | |
| The Primitive Vascular System | |
| | |
| Development of the Arteries | |
| | |
| Development of the Veins | |
| | |
| Fetal Circulation and the Changes at Birth | |
| | |
| The Lymphatic System | |
| | |
| [[Book - Developmental Anatomy 1924-10|Chapter X. - The Skeletal System]]
| |
| | |
| Histogenesis of the Supporting Tissues
| |
| | |
| Connective Tissue
| |
| | |
| Cartilage
| |
| | |
| Bone
| |
| | |
| Morphogenesis of the Skeleton
| |
| | |
| The Axial Skeleton
| |
| | |
| The Appendicular Skeleton
| |
| | |
| [[Book - Developmental Anatomy 1924-11|Chapter XI. - The Muscular System]]
| |
| | |
| The Histogenesis of Muscle
| |
| | |
| Morphogenesis of the Muscles
| |
| | |
| Ectodermal Derivatives
| |
| | |
| [[Book - Developmental Anatomy 1924-12|Chapter XII. - The Integumentary System]]
| |
| | |
| The Skin
| |
| | |
| The Nails
| |
| | |
| The Hair
| |
| | |
| Sebaceous Glands
| |
| | |
| Sweat Glands
| |
| | |
| Mammary Glands
| |
| | |
| [[Book - Developmental Anatomy 1924-13|Chapter XIII. - The Central Nervous System]]
| |
| | |
| Histogenesis of the Nervous Tissues
| |
| | |
| Morphogenesis of the Central Nervous System
| |
| | |
| The Spinal Cord
| |
|
| |
| The Brain
| |
| | |
| [[Book - Developmental Anatomy 1924-14|Chapter XIV. - The Peripheral Nervous System]]
| |
| | |
| The Spinal Nerves
| |
| | |
| The Cranial Nerves
| |
| | |
| The Sympathetic Nervous System
| |
| | |
| The Chromaffin Bodies and Suprarenal Gland
| |
| | |
| [[Book - Developmental Anatomy 1924-15|Chapter XV. - The Sense Organs]]
| |
| | |
| General Sensory Organs
| |
| | |
| The Gustatory Organ 292
| |
| | |
| The Nose
| |
| | |
| The Eye
| |
| | |
| The Ear
| |
| | |
| | |
| Part III. A LABORATORY MANUAL OF EAIBRYOLOGY
| |
| | |
| | |
| [[Book - Developmental Anatomy 1924-16|Chapter XVI. - The Study of Chick Embryos]]
| |
|
| |
| The Unincubated Ovum and Embryos of the First Day
| |
| | |
| Embryo of Five Segments (Twenty-Three Hours)
| |
|
| |
|
| Embryo of Seven Segments (Twenty-five Hours) | | * [[Book - Developmental Anatomy 1924-10|Chapter X. - The Skeletal System]] |
|
| | ** Histogenesis of the Supporting Tissues |
| Embryo of Seventeen Segments (Thirty-eight Hours) | | ** Connective Tissue |
| | ** Cartilage |
| | ** Bone |
| | ** Morphogenesis of the Skeleton |
| | ** The Axial Skeleton |
| | ** The Appendicular Skeleton |
| | * [[Book - Developmental Anatomy 1924-11|Chapter XI. - The Muscular System]] |
| | ** The Histogenesis of Muscle |
| | ** Morphogenesis of the Muscles |
| | ** Ectodermal Derivatives |
| | * [[Book - Developmental Anatomy 1924-12|Chapter XII. - The Integumentary System]] |
| | ** The Skin |
| | ** The Nails |
| | ** The Hair |
| | ** Sebaceous Glands |
| | ** Sweat Glands |
| | ** Mammary Glands |
| | * [[Book - Developmental Anatomy 1924-13|Chapter XIII. - The Central Nervous System]] |
| | ** Histogenesis of the Nervous Tissues |
| | ** Morphogenesis of the Central Nervous System |
| | ** The Spinal Cord |
| | ** The Brain |
| | * [[Book - Developmental Anatomy 1924-14|Chapter XIV. - The Peripheral Nervous System]] |
| | ** The Spinal Nerves |
| | ** The Cranial Nerves |
| | ** The Sympathetic Nervous System |
| | ** The Chromaffin Bodies and Suprarenal Gland |
| | * [[Book - Developmental Anatomy 1924-15|Chapter XV. - The Sense Organs]] |
| | ** General Sensory Organs |
| | ** The Gustatory Organ 292 |
| | ** The Nose |
| | ** The Eye |
| | ** The Ear |
| | | valign=top| Part III. A LABORATORY MANUAL OF EMBRYOLOGY |
| | * [[Book - Developmental Anatomy 1924-16|Chapter XVI. - The Study of Chick Embryos]] |
| | ** The Unincubated Ovum and Embryos of the First Day |
| | ** Embryo of Five Segments (Twenty-Three Hours) |
| | ** Embryo of Seven Segments (Twenty-five Hours) |
| | ** Embryo of Seventeen Segments (Thirty-eight Hours) |
| | ** Embryo of Twenty-seven Segments (Two Days) |
| | ** Embryos of Three to Four Days |
| | * [[Book - Developmental Anatomy 1924-17|Chapter XVII. - The Study of Pig Embryos]] |
| | ** The Anatomy of a Six Mm. Pig Embryo |
| | ** The Anatomy of Ten to Twelve Mm, Pig Embryos |
| | ** The Anatomy of an Eighteen Mm. Pig Embryo |
| | ** The Anatomy of a Thirty-five Mm. Pig Embryo |
| | ** Methods for the Dissection of Pig Embryos |
| | |} |
|
| |
|
| Embryo of Twenty-seven Segments (Two Days)
| | ==Part I. General Development== |
|
| |
|
| Embryos of Three to Four Days
| | ===Introduction=== |
|
| |
|
| [[Book - Developmental Anatomy 1924-17|Chapter XVII. - The Study of Pig Embryos]]
| | ====The Scope of Embryology==== |
| | Developmental anatomy, or embryology, traces the formative history of the individual from the origin of the germ cells to the adult condition. Although the most striking changes in human development occur while the young (called an embryo or fetus) is still inside its mother's womb, yet development by no means ceases at birth. Birth is a mere incident which occurs when the new individual is sufficiently advanced to allow its transference from a protected riterine environment to one in the external world. Some vertebrates, like fishes and amphibia, are capable of an active and independent existence at very immature stages; these free-living larvae, as they are termed, then gradually progress to adults. The human newborn, although far more complete anatomically, is still utterly dependent for food and care: many years of infancy and childhood must elapse before it becomes self-maintaining in human society. During all this period, postnatal development continues. Birth, itself, initiates anatomical changes of profound influence on the body. Throughout the entire growth period, with its uneven but steadily slowing growth rate, come the completion of some organs and a gradual remoulding of the shape of the body and its parts. Only at the age of twenty-five are these progressive changes complete. |
|
| |
|
| The Anatomy of a Six Mm. Pig Embryo
| |
|
| |
|
| The Anatomy of Ten to Twelve Mm, Pig Embryos | | All vertebrate, or backboned, animals are organized upon a common anatomical plan, and even many of their structural details are comparable, though superficially disguised. Similarly, their fundamental mode of development is essentially identical. The minor variations that do occur are caused by such secondary modifying factors as the crowding yolk content of the egg or adaptations to development inside or outside the mother's body. While the comparative viewpoint is indispensable for gaining a broad understanding of embryology, it has been of especial importance in supplying missing parts of the human developmental story and in interpreting many perplexing conditions. For, the earliest human embryos known are about two weeks old and have the three primary germ layers already formed. Even invertebrate material is highly useful for demonstrating such early stages as maturation, fertilization, cleavage, and the formation of blastula and gastrula. |
|
| |
|
| The Anatomy of an Eighteen Mm. Pig Embryo | | ====The Value of Embryology==== |
| | A general conception of how man and other animals develop from a single cell by orderly and logical processes should share in the cultural background of every educated mind. To the medical student, embryology is of primary importance because it affords a comprehensive understanding of the intricacies and variations of human anatomy, and thus is essential to sound surgical training. It also explains many anomalies and 'monstrous - conditions, and the origin of certain tumors and other pathological changes in the tissues. Obstetrics is essentially applied embryology. From the theoretical side, it is the key with which we may unlock the secrets of heredity, the determination of sex, and, in part, of organic evolution. |
|
| |
|
| The Anatomy of a Thirty-five Mm. Pig Embryo
| |
|
| |
|
| Methods for the Dissection of Pig Embryos
| | ====Historical==== |
| | [[File:Arey1924 fig001.jpg|thumb|Fig. 1. - Human sperm cell containing a miniature organism, according to Hartsoeker (1694).]] |
| | The science of modern embryology is comparatively new, originating with the use of the compound microscope and advancing with the improvement of microscopical technique. [[Book_-_Fathers_of_Biology#Aristotle|Aristotle]] (384-322 B. c.), however, centuries before the introduction of magnifying lenses had followed the general development of the chick, day by day. The popular belief that slime and decaying matter is capable of giving rise to living animals, as also asserted by Aristotle, was disproved by Redi (1668). |
|
| |
|
|
| |
|
| PART I. GENERAL DEVELOPMENT
| | A few years after Harvey and Malpighi had published their fundamental studies on the chick embryo, Leeuwenhoek reported the discovery of the human spermatozoon by Ham in 1677 - At this period, it was believed either that fully formed animals existed in miniature in the egg, needing only the stimulus of the spermatozoon to initiate development, or that similarly preformed bodies, male and female, constituted the spermatozoa and that these merely enlarged within the ovum. According to this doctrine of preformation, all future generations were likewise encased, one inside the sex cells of the other, and serious computations were made as to the probable number of progeny (200 millon) thus present in the ovary of Mother Eve, at the exhaustion of which the human race would end! Dalenpatius (1699) and others even believed they had observed a minute human form in the spermatozoon (Fig. 1). |
|
| |
|
|
| |
|
| INTRODUCTION
| | The preformation theory was strongly combated by [[:File:Theoria Generationis 1774.jpg|Wolff (1759)]], who saw that the organs of the early chick embryo were differentiated gradually from unspecialized living substance. This theory, known as epigenesis, was proved correct when von Baer discovered the mammalian ovum in 1827, and later demonstrated the germ-layer composition of all embryos. |
|
| |
|
|
| |
|
| The Scope of Embryology. - Developmental anatomy, or embryology, traces the formative history of the individual from the origin of the germ cells to the adult condition. Although the most striking changes in human development occur while the young (called an embryo or fetus) is still inside its mother - s womb, yet development by no means ceases at birth. Birth is a mere incident which occurs when the new individual is sufficiently advanced to allow its transference from a protected riterine environment to one in the external world. Some vertebrates, like fishes and amphibia, are capable of an active and independent existence at very immature stages; these free-living larvee, as they are termed, then gradually progress to adults. The human newborn, although far more complete anatomically, is still utterly dependent for food and care: many years of infancy and childhood must elapse before it becomes self-maintaining in human society. During all this period, postnatal development continues. Birth, itself, initiates anatomical changes of profound influence on the body. Throughout the entire growth period, with its uneven but steadily slowing growth rate, come the completion of some organs and a gradual remoulding of the shape of the body and its parts. Only at the age of twenty-five are these progressive changes complete.
| | About twenty years after Schleiden and Schwann (1839) had shown the cell to be the structural unit of the organism, the ovum and spermatozoon were recognized as true cells. [[:File:Oskar Hertwig.jpg|O. Hertwig]], in was the first to observe and appreciate the events of fertilization. Henceforth, all multicellular organisms were believed to develop each from a single fertilized ovum. This conception is expressed in the famous aphorism: ''ornne vivum ex ovo'' . |
|
| |
|
| | ====Modern Embryology==== |
| | As an organized and definite science, began with [[:File:Francis Balfour.jpg|Balfour (1874)]], who reviewed, digested, and made accessible the earlier scattered facts. Throughout this period, the experimental method of investigation has been used increasingly; without it many structural and physiological aspects of development would remain unsolved. |
|
| |
|
| All vertebrate, or backboned, animals are organized upon a common anatomical plan, and even many of their structural details are comparable, though superficially disguised. vSimilarly, their fundamental mode of development is essentially identical. The minor variations that do occur are caused by such secondary modifying factors as the crowding yolkcontent of the egg or adaptations to development inside or outside the mother - s body. While the comparative viewpoint is indispensable for gaining a broad understanding of embryology, it has been of especial importance in supplying missing parts of the human developmental story and in interpreting many perplexing conditions. For, the earliest human embryos known are about two weeks old and have the three primary germ layers already formed. Even invertebrate material is highly useful for demonstrating such early stages as maturation, fertilization, cleavage, and the formation of blastula and gastrula.
| | ===General Features Of Development=== |
| | |
| | |
| The Value of Embryology. - A general conception of how man and other animals develop from a single cell by orderly and logical processes should share in the cultural background of every educated mind. To the medical student, embryology is of primary importance because it affords a comprehensive understanding of the intricacies and variations of human anatomy, and thus is essential to sound surgical training. It also explains many anomalies and 'monstrous - conditions, and the origin of certain tumors and other pathological changes in the tissues. Obstetrics is essentially applied embryology. From the theoretical side, it is the key with which we may unlock the secrets of heredity, the determination of sex, and, in part, of organic evolution.
| |
| | |
| | |
| Histoucal. - The science of modern embryology is comparatively new, originating with the use of the compound microscope and advancing with the improvement of microscopical technique. Aristotle (384-322 B. c.), however, centuries before the introduction of magnifying lenses had followed the general development of the chick, day by day. The popular belief that slime and decaying matter is capable of giving rise to living animals, as also asserted by Aristotle, was disproved by Redi (1668).
| |
| | |
| .A few years after Harvey and Malpighi had published their fundamental studies on the chick embryo, Leeuwenhoek reported the discovery of the human Sp ermatozoon by Ham in 1677 - At this period, it was believed either that fully formed animals existed in miniature in the egg, needing only the stimulus of the spermatozoon to initiate development, or that similarly preformed bodies, male and female, constituted the spermatozoa and that these merely enlarged within the ovum. According to this doctrine of preformation, all future generations were likewise encased, one inside the sex cells of the other, and serious computations were made as to the probable number of progeny (200 millon) thus present in the ovary of Mother Eve, at the exhaustion of which the human race would end! Dalenpatius ( 1699) and others even believed they had observed a minute human form in the spermatozoon (Fig. i).
| |
| | |
| | |
| | |
| The preformation theory was strongly combated by Wolff ( 175 9), who saw that the organs of the early chick embryo were differentiated gradually from unspecialized li ving substance. This theory, known as epi genesis, was proved correct wEen von Baer discQvered the mammalian ovurn in 1 827, and later demonstrated the germ-layer composition of all embryos.
| |
| | |
| | |
| Fig. I. - Human sperm cell containing a miniature organism, according to Hartsoeker (1694).
| |
| | |
| | |
| About twenty years after Schleiden and Schwann (1839) had shown the cell to be the structural unit of the organism, the ovum and spermatozoon were recognized as true cells. O. Hertwig , in was the first to
| |
| observe and appreciate the events of fertilizat ion . Henceforth, all multicellular organisms were believed to develop each from a single fertilized ovum. This conception is expressed in the famous aphorism: - ornne_ vivum ex ovo . -
| |
| Modern embryology , as an organized and definite science, began with Balfou r (1874 ), who reviewed, digested, and made accessible the earlier scattered facts. Throughout this period, the experimental method of investigation has been used increasingly; without it many structural and physiological aspects of development would remain unsolved.
| |
| | |
| | |
| | |
| GENERAL FEATURES OF DEVELOPMENT
| |
| | |
|
| |
|
| A multicellular embryo results from the division of the fertilized ovum to form daughter cells. These are at first quite similar in structure, and, if separated, in some animals each may become a complete embryo (sea urchin; certain vertebrates). In general, the development of an embryo depends: (i) upon the multiplication of its cells by division; (2) upon the growth in size of the individual cells; (3) upon changes in their form and structure. | | A multicellular embryo results from the division of the fertilized ovum to form daughter cells. These are at first quite similar in structure, and, if separated, in some animals each may become a complete embryo (sea urchin; certain vertebrates). In general, the development of an embryo depends: (i) upon the multiplication of its cells by division; (2) upon the growth in size of the individual cells; (3) upon changes in their form and structure. |
|
| |
|
|
| |
|
| Cell Division. - All cells arise from pre-existing cells by division. There are two methods of cell division - amitosis and mitosis. | | Cell Division - All cells arise from pre-existing cells by division. There are two methods of cell division - amitosis and mitosis. |
| | |
| | |
| Amitosis. - Cells may divide directly by the simple fission of their nuclei and cytoplasm. This rather infrequent process is called amitosis. Amitosis is said by many to occur only in specialized or moribund cells. It is the type of cell division demonstrable in the epithelium of the bladder.
| |
| | |
| | |
| Mitosis - In the reproduction of typically active somatic cells and in all germ cells, complicated changes take place in the nucleus. These changes give rise to thread-like structures, hence the process is termed mitosis (thread) in distinction to amitosis (no threadk Mitosis is divided for convenience into four phases (Fig. 2) :
| |
| Prophase. - 1. The centrosome divides and the two minute bodies resulting from the division move apart, ultimately occup^dng positions at opposite poles of the nucleus (I-uI).
| |
| | |
| | |
| | |
| 2. Astral rays appear in the cytoplasm about each centriole. They radiate from it, and the threads of the central or achromatic spindle are formed between the two asters, thus constituting the amphiastcr (u).
| |
| | |
| | |
| 3. The nuclear membrane and nucleolus disappear, the karyoplasm and cytoplasm becoming confluent.
| |
| | |
|
| |
|
| 4. During the above changes the chromatic network of the resting nucleus resolves itself into a skein, or spireme, which soon shortens and l;)reaks up into distinct, heavily-staining bodies, the chromosomes (u, uI). x\ definite number of chromosomes is always found in the cells of a given species, the chromosomes may be block-shaped, rod- shaped, or bent in the form of a U or V.
| |
|
| |
|
| | Amitosis - Cells may divide directly by the simple fission of their nuclei and cytoplasm. This rather infrequent process is called amitosis. Amitosis is said by many to occur only in specialized or moribund cells. It is the type of cell division demonstrable in the epithelium of the bladder. |
|
| |
|
| | [[File:Arey1924 fig002.jpg|thumb|Fig. 2. Diagrams of the phases of mitosis (Schafer).]] |
| | ====Mitosis==== |
| | In the reproduction of typically active somatic cells and in all germ cells, complicated changes take place in the nucleus. These changes give rise to thread-like structures, hence the process is termed mitosis (thread) in distinction to amitosis (no thread). Mitosis is divided for convenience into four phases (Fig. 2) : |
|
| |
|
| Fig. 2. - Diagrams of the phases of mitosis (Schafer).
| | Prophase |
| | # The centrosome divides and the two minute bodies resulting from the division move apart, ultimately occupying positions at opposite poles of the nucleus (I-III). |
| | # Astral rays appear in the cytoplasm about each centriole. They radiate from it, and the threads of the central or achromatic spindle are formed between the two asters, thus constituting the amphiaster (II). |
| | # The nuclear membrane and nucleolus disappear, the karyoplasm and cytoplasm becoming confluent. |
| | # During the above changes the chromatic network of the resting nucleus resolves itself into a skein, or spireme, which soon shortens and breaks up into distinct, heavily-staining bodies, the chromosomes (II, III). The definite number of chromosomes is always found in the cells of a given species, the chromosomes may be block-shaped, rod- shaped, or bent in the form of a II or V. |
| | # The chromosomes arrange themselves in the equatorial plane of the central spindle (IV). If U- or V-shaped, the angle of each is directed toward a common center. The amphiaster and the chromosomes together constitute a mitotic figure, and at the end of the prophase this is called a monaster. |
|
| |
|
|
| |
|
| | Metaphase - The longitudinal splitting of the chromosomes into exactly similar halves constitutes the metaphasc (IV). The aim of mitosis is thus accomplished, an accurate division of the chromatin between the nuclei of the daughter cells. |
|
| |
|
| 5. The chromosomes arrange themselves in the equatorial plane of the central spindle (IV). If U- or V-shaped, the angle of each is directed toward a common center. The amphiaster and the chromosomes together constitute a mitotic figure, and at the end of the prophase this is called a monaster.
| |
|
| |
|
| | Anaphase - The two groups of daughter chromosomes separate and move up along the central spindle fibers, each toward one of the two asters. Hence this is called the diaster stage (V, VI). Each centriole may divide in preparation for the next diviSion of the daughter cells. |
|
| |
|
| Metaphase. - The longitudinal splitting of the chromosomes into exactly similar halves constitutes the metaphasc (IV). The aim of mitosis is thus accomplished, an accurate division of the chromatin between the nuclei of the daughter cells.
| | Telophase - i. The daughter chromosomes resolve themselves into a reticulum and daughter nuclei are formed (Vu, VuI). |
|
| |
|
|
| |
|
| Anaphase. - The two groups of daughter chromosomes separate and move up along the central spindle fibers, each toward one of the two asters. Hence this is called the diaster stage (V, VI). Each centriole may divide in preparation for the next diviSion of the daughter cells.
| | 2. The cytoplasm divides in a plane perpendicular to the axis of the mitotic spindle (VIII). Two complete daughter cells have thus arisen from the mother cell. |
|
| |
|
| Telophase. - i. The daughter chromosomes resolve themselves into a reticulum and daughter nuclei are formed (Vu, VuI).
| |
|
| |
|
| | The number of chromosomes is constant in the cells of a given species. The smallest assortment, two, occurs in Ascaris megalocephala univaleus, a round worm parasitic in the intestine of the horse. The largest number known is found in the brine shrimp, Artemia, where 168 have been counted. The chromosome enumeration for the human cell has been variously stated but the results of Winiwarter (1912), Grosser (1921), and Painter (1923) now agree on a relatively high number, which Painter establishes as 48 for whites and negroes of both sexes. |
|
| |
|
| 2. The cytoplasm divides in a plane perpendicular to the axis of the mitotic spindle (VuI). Two complete daughter cells have thus arisen from the mother cell.
| | ===The Germ Layers=== |
| | [[File:Arey1924 fig003.jpg|thumb|Fig. 3. Mesenchyme from a chick embryo (Prentiss). X 495.]] |
| | The first changes in the form and arrangement of the cells establish three definite plates, the primary germ layers, which are termed from their positions the ectoderm (outer skin), mesoderm (middle skin) and entoderm (inner skin) (Fig. 4). Since the ectoderm covers the body, it is primarily protective in function, but it also gives origin to the nervous system, through which sensations are received from the outer world. The entoderm, on the other hand, lines the digestive canal and is from the first nutritive. The mesoderm, lying between the other two layers, naturally performs the functions of circulation, of muscular movement, and of excretion; it also gives rise to the skeletal structures which support the body. While all three germ layers form definite sheets of cells known as epithelia, the mesoderm takes also the form of a diffuse meshwork of cells, the mesenchyme (Fig. 3). |
|
| |
|
| The number of chromosomes is constant in the cells of a given species. The smallest assortment, two, occurs in Ascaris megalocephala univaleus, a round worm parasitic in the intestine of the horse. The largest number known is found in the brine shrimp, Artemia, where 1 68 have been counted. The chromosome enumeration for the human cell has been variously stated but the results of Winiwarter (1912), Grosser (1921 ), and Painter (1923) now agree on a relatively high number, which Painter establishes as 48 for whites and negroes of both sexes.
| |
|
| |
|
| |
|
| |
| The Germ Layers. - The first changes in the form and arrangement of the cells establish three definite plates, the primary germ layers, which are termed from their positions the ectoderm (outer skin), mesoderm (middle skin) and entoderm (inner skin) (Fig. 4). Since the ectoderm covers the body, it is primarily protective in function, but it also gives origin to the nervous system, through which sensations are received from the outer world. The entoderm, on the other hand, lines the digestive canal and is from the first nutritive. The mesoderm, lying between the other two layers, naturally performs the functions of circulation, of muscular movement, and of excretion; it also gives rise to the skeletal structures which support the body. While all three germ layers form definite sheets of cells known as epithelia, the mesoderm takes also the form of a diffuse meshwork of cells, the mesenchyme (Fig. 3).
| |
|
| |
|
| |
|
| |
| Fig. 3. - Alesenchyme from a chick embryo (Prentiss). X 495.
| |
|
| |
|
|
| |
|
| Line 388: |
Line 258: |
| This series of changes - an embryonic (undifferentiated) stage; progressive functional s])ecialization ; gradual degeneration; death and removal - which tissue cells experience is designated by the term cytomorphosis. | | This series of changes - an embryonic (undifferentiated) stage; progressive functional s])ecialization ; gradual degeneration; death and removal - which tissue cells experience is designated by the term cytomorphosis. |
|
| |
|
| Derivatives of the Germ Layers. - The tissues of the adult are derived from the primary germ layers as follows: . | | ====Derivatives of the Germ Layers==== |
| | |
| | The tissues of the adult are derived from the primary germ layers as follows: |
| | |
| | {| |
| | |-bgcolor="CEDFF2" |
| | ! width=30%|Ectoderm |
| | ! width=30%|Mesoderm |
| | ! width=30%|Entoderm |
| | |- |
| | | I. Epidermis and derivatives. |
|
| |
|
| | Hair; nails; glands. |
|
| |
|
| Ectoderm
| | Lens of eye. |
|
| |
|
| Mesoder
| | 2. Epithelium of: |
|
| |
|
| | Organs of special sense. Cornea. |
|
| |
|
| Entoderm Epithelium of:
| | Mouth; enamel organ. |
| I. Pharynx and derivatives. Auditory tube.
| | |
| | Oral glands; hypophysis. |
|
| |
|
| Tonsils.
| | Anus. |
|
| |
|
| Thymus.
| | Amnion; chorion. |
|
| |
|
| Thyroid.
| | 3. Nervous tissue. |
|
| |
|
| | Neuroglia. |
|
| |
|
| I. Epidermis and derivatives. Hair; nails; glands.
| | Chromaffin tissue. |
|
| |
|
| Lens of eye.
| | 4. Smooth muscle of; Iris. |
|
| |
|
| A. Mesothelium. | | Sweat glands. |
| | | A. Mesothelium. |
|
| |
|
| 1. Pericardium. | | 1. Pericardium. |
| Line 418: |
Line 303: |
| 3. Peritoneum. | | 3. Peritoneum. |
|
| |
|
| 4. LTrogenital epithelia. | | 4. Urogenital epithelia. |
|
| |
|
| 5. Striated muscle. | | 5. Striated muscle. |
|
| |
|
| | B. Mesenchyme. |
|
| |
|
| | 1 . Smooth muscle. |
|
| |
|
| 2. Epithelium of: | | 2. Notochord. |
|
| |
|
| Organs of special sense. Cornea.
| | 3. Connective tissue; cartilage; bone. |
|
| |
|
| | 4. Blood; bone marrow. |
|
| |
|
| Ectoderm
| | 5. Endothelium of blood vessels and lymphatics. |
|
| |
|
| | 6. Lymphoid organs. |
|
| |
|
| Mesoderm
| | 7. Suprarenal cortex. |
| | | Epithelium of: |
|
| |
|
| | I. Pharynx and derivatives. Auditory tube. |
|
| |
|
| Entoderm
| | Tonsils. |
|
| |
|
| | Thymus. |
|
| |
|
| Mouth; enamel organ. Oral glands; hypophysis. Anus.
| | Thyroid. |
| | |
| Amnion; chorion.
| |
| | |
| | |
| B. Mesenchyme.
| |
| | |
|
| |
|
| Parathyroid. | | Parathyroid. |
| Line 450: |
Line 336: |
| 2. Respiratory tract. | | 2. Respiratory tract. |
|
| |
|
| | | Larynx; trachea. |
| 1 . Smooth muscle.
| |
| | |
| 2. Notochord.
| |
| | |
| 3. Connective tissue; .
| |
| | |
|
| |
|
| Lungs. | | Lungs. |
| Line 462: |
Line 342: |
| 3. Digestive tract. | | 3. Digestive tract. |
|
| |
|
| Larynx; trachea.
| | Liver; pancreas. |
| | |
| 3. Nervous tissue. Neuroglia. Chromaffin tissue.
| |
| | |
| | |
| cartilage; bone.
| |
| | |
| 4. Blood; bone marrow.
| |
| | |
| 5. Endothelium of blood .
| |
| | |
|
| |
|
| Yolk sac; allantois. | | Yolk sac; allantois. |
| Line 478: |
Line 348: |
| 4. Bladder (except trigone). | | 4. Bladder (except trigone). |
|
| |
|
| 5. LTrethra (except prostatic). | | 5. Urethra (except prostatic). |
|
| |
|
| 6. Prostate. | | 6. Prostate. |
| | |} |
|
| |
|
| Liver; pancreas.
| | ===Primitive Segments - Metamerism=== |
| | | [[File:Arey1924 fig004.jpg|thumb|Fig. 4. Diagrammatic transverse section of a vertebrate embryo (Minot-Prentiss).]] |
| 4. Smooth muscle of; Iris.
| |
| | |
| Sweat glands.
| |
| | |
| vessels and lymphatics.
| |
| | |
| 6. Lymphoid organs.
| |
| | |
| 7. Suprarenal corte.
| |
| | |
| | |
| | |
| | |
| | |
| Primitive Segments - Metamerism. - A prominent feature of vertebrate embryos are the primitive segments, or metameres (Fig. 59). These segments are homologous to the serial divisions of an adult earth-worm - s body, divisions which, in the earth worm, are identical in structure, each containing a ganglion of the nerve cord, a muscle segment, or myotome, and pairs of blood vessels and nerves. In vertebrate embryos, the block like primitive segments lie next the neural tube and are known as mesodermal segments, or somites (Fig. 4). Each pair gives rise to a vertebra, to two myotomes, or muscle segments, and to paired vessels; each set of mesodermal segments is supplied by a pair of spinal nerves: consequently, the adult vertebrate body is segmented like that of the earth worm. As a worm grows by the formation of new segments at its tail-end, so the metameres of the vertebrate embryo begin to form in the head and are added tailward. There is this difference between the segments of the worm and the vertebrate embryo; the segmentation of the worm is complete, while that of the vertebrate is incomplete ventrally. | |
| | |
| | |
| | |
| Fig. 4. - Diagrammatic transverse section of a vertebrate embryo (Minot-Prentiss). | |
|
| |
|
| | A prominent feature of vertebrate embryos are the primitive segments, or metameres (Fig. 59). These segments are homologous to the serial divisions of an adult earth-worm's body, divisions which, in the earth worm, are identical in structure, each containing a ganglion of the nerve cord, a muscle segment, or myotome, and pairs of blood vessels and nerves. In vertebrate embryos, the block like primitive segments lie next the neural tube and are known as mesodermal segments, or somites (Fig. 4). Each pair gives rise to a vertebra, to two myotomes, or muscle segments, and to paired vessels; each set of mesodermal segments is supplied by a pair of spinal nerves: consequently, the adult vertebrate body is segmented like that of the earth worm. As a worm grows by the formation of new segments at its tail-end, so the metameres of the vertebrate embryo begin to form in the head and are added tailward. There is this difference between the segments of the worm and the vertebrate embryo; the segmentation of the worm is complete, while that of the vertebrate is incomplete ventrally. |
|
| |
|
|
| |
|
| Somatopleure and Splanchnopleure. In early embryos the mesoderm splits into two layers, the somatic (dorsal) and splanchnic (ventral) mesoderm (Fig. 4). The ectoderm and somatic mesoderm constitute the lu)dy wall, which is termed the somatopleure. In the same way, the entoderm and splanchnic mesoderm combine as the splanchnopleure; it forms the mesenteries and the walls of the gut, heart, and lungs. | | ===Somatopleure and Splanchnopleure=== |
| | In early embryos the mesoderm splits into two layers, the somatic (dorsal) and splanchnic (ventral) mesoderm (Fig. 4). The ectoderm and somatic mesoderm constitute the body wall, which is termed the somatopleure. In the same way, the entoderm and splanchnic mesoderm combine as the splanchnopleure; it forms the mesenteries and the walls of the gut, heart, and lungs. |
|
| |
|
|
| |
|
| Coelom. -The space between the somatopleure and splanchnopleure is the ccclom, or body cavity. At the first splitting of the mesoderm, isolated clefts are produced. These unite on each side and eventually form one cavity - the coelom. With the extension of the mesoderm, the cot'lom surrounds the heart and gut ventrally (Fig. 4). Later, it is subdivided into the pericardial caznty about the heart, the pleural cavity of the thorax, and the peritoneal cavity of the abdominal region. The ci)ithelia lining the several body cavities are termed mesothelia. | | ===Coelom=== |
| | The space between the somatopleure and splanchnopleure is the coelom, or body cavity. At the first splitting of the mesoderm, isolated clefts are produced. These unite on each side and eventually form one cavity - the coelom. With the extension of the mesoderm, the coelom surrounds the heart and gut ventrally (Fig. 4). Later, it is subdivided into the pericardial cavity about the heart, the pleural cavity of the thorax, and the peritoneal cavity of the abdominal region. The epithelia lining the several body cavities are termed mesothelia. |
|
| |
|
|
| |
|
| The Nephrotome. - The bridge of cells connecting the primitive segment with the unsegmented somatic and splanchnic layers is the nephrotome, or intermediate cell mass (Fig. 4). From these will develop the urogenital glands and ducts. | | ===The Nephrotome=== |
| | The bridge of cells connecting the primitive segment with the unsegmented somatic and splanchnic layers is the nephrotome, or intermediate cell mass (Fig. 4). From these will develop the urogenital glands and ducts. |
|
| |
|
| | | ===Developmental Processes=== |
| D evelopmental Processes. - The developing embryo exhibits a ])rogressively comjilex structure, the various steps in the production of which occur in orderly sequence. There may be recognized in development a number of component mechanical processes which are used repeatedly by the embryo. The general and fundamental process conditioning ilifferentiation is cell multiplication, and the subsequent growth of the daughter cells. The more important of the specific developmental ])rocesses are the following: ( i) cell migration; (2) localized growth, resulting in eidargements and constrictions; (3) cell aggregation, forming (a) cords, (b) sheets, [c] masses; (4) delamination, that is, the splitting of single sheets into separate layers; (5) folds, including circumscribed folds which produce ia) evaginations, or out-pocketings, (b) invaginations, or in-pocketings.
| | The developing embryo exhibits a progressively complex structure, the various steps in the production of which occur in orderly sequence. There may be recognized in development a number of component mechanical processes which are used repeatedly by the embryo. The general and fundamental process conditioning ilifferentiation is cell multiplication, and the subsequent growth of the daughter cells. The more important of the specific developmental processes are the following: ( 1) cell migration; (2) localized growth, resulting in eidargements and constrictions; (3) cell aggregation, forming (a) cords, (b) sheets, [c] masses; (4) delamination, that is, the splitting of single sheets into separate layers; (5) folds, including circumscribed folds which produce (a) evaginations, or out-pocketings, (b) invaginations, or in-pocketings. |
|
| |
|
|
| |
|
| The production of folds, including evaginations and invaginations, due to unequal rapidity of growth, is the chief factor in moulding the organs and hence the general form of the embryo. | | The production of folds, including evaginations and invaginations, due to unequal rapidity of growth, is the chief factor in moulding the organs and hence the general form of the embryo. |
|
| |
|
| | ===Fundamental Conceptions=== |
| | ====The Anlage==== |
| | This German word, which lacks an entirely satisfactory English equivalent, is a term applied to the first discernible cell, or aggregation of cells, which is destined to form any distinct jiart or organ of the embryo. In the broad sense, the fertilized ovum is the anlage of the entire adult organism; furthermore, in the early cleavage stages of certain embryos it is possible to recognize single cells or cell groups from which definite structures will indubitably arise. The term anlage, however, is more commonly applied to the primordia that differentiate from the various germ layers. Thus the epithelial thickening over the optic vesicle is the anlage of the lens. |
|
| |
|
| | ====The Law of Genetic Restriction==== |
| | As development advances, there is a constantly increasing restriction in the kind of differentiation open to the various parts. Each emerging tissue or organ is more rigidly bound to its particular type of differentiation than was the generalized material from which it came. A line of specialization, once begun, cannot be abandoned for another type. The parent tissue, likewise, is limited by losing the capacity for duplicating anlages already formed. Thus, the primitive thyroid can never become anything but a thyroid, whereas the gut that formed it also buds off, at other levels, the lungs, liver, and pancreas. Yet if the embryonic thyroid were destroyed, the pharynx would never replace it. From mesenchyme arise connective tissue, blood cells, and smooth muscle; when once the specialization begins, there can be no retraction or transformation to another type. |
|
| |
|
| FUNDAMENTAL CONCEPTIONS
| | ====Continuity of the Germ Plasm==== |
| | According to this important conception of Weismann, the body-protoplasm, or soma, and the reproductive-protoplasm differ fundamentally. The germinal material is a legacy that has existed since the beginning of life, from which representative portions are passed on intact from one generation to the next. Around this germ plasm there develops in each successive generation a shortlived body, or soma, which serves as a vehicle for insuring its transmission and perpetuation. The reason, therefore, why offspring resembles parent is because each develops from portions of the same stuff. |
|
| |
|
| The Anlage. - This German word, which lacks an entirely satisfactory English equivalent, is a term applied to the first discernible cell, or aggregation of cells, which is destined to form any distinct jiart or organ of the embryo. In the broad sense, the fertilized ovum is the anlage of the entire adult organism; furthermore, in the early cleavage stages of certain embryos it is possible to recognize single cells or cell groups from which definite structures will indubitably arise. The term anlage, however, is more commonly applied to the primordia that differentiate from the various germ layers. Thus the epithelial thickening over the optic vesicle is the anlage of the lens.
| | ====The Law of Biogenesis==== |
| | | Of great theoretical interest is the fact, constantly observed in studying, embryos, that the individual in its development repeats hastily and incompletely the evolutionary history of its own species. This law of recapitulation was first stated clearly by Muller in 1863, and was termed by Haeckel the law of biogenesis. In accordance with it, the fertilized ovum is compared to a unicellular organism like the Ameba: the blastula is supposed to represent an adult Volvox type; the gastrula, a simple sponge; the segmented embryo, a worm-like stage ; and the embryo with gill slits may be regarded as a fishlike stage. Moreover, the blood of the human embryo in development passes through stages in which its corpuscles resemble in structure those of the fish and reptile; the heart is at first tubular, like that of the fish, and the arrangement of blood vessels is equally primitive; the kidney of the embryo is like that of the amphibian, as are also the genital ducts. Many other examples of this law may readily be observed. |
| | |
| The Law of Genetic Restriction. - As development advances, there is a constantly increasing restriction in the kind of differentiation open to the various parts. Each emerging tissue or organ is more rigidly bound to its particular type of differentiation than was the generalized material from which it came. A line of specialization, once begun, cannot be abandoned for another type. The parent tissue, likewise, is limited by losing the capacity for duplicating anlages already formed. Thus, the primitive thyroid can never become anything but a thyroid, whereas the gut that formed it also buds off, at other levels, the lungs, liver, and pancreas. Yet if the embryonic thyroid were destroyed, the pharynx would never replace it. From mesenchyme arise connective tissue, blood cells, and smooth muscle; when once the specialization begins, there can be no retraction or transformation to another type. | |
| | |
| | |
| Continuity of the Germ Plasm. - According to this important conception of Weismann, the body-protoplasm, or soma, and the reproductive-protoplasm differ fundamentally. The germinal material is a legacy that has existed since the beginning of life, from which representative portions are passed on intact from one generation to the next. Around this germ plasm there develops in each successive generation a shortlived body, or soma, which serves as a vehicle for insuring its transmission and perpetuation. The reason, therefore, why offspring resembles parent is because each develops from portions of the same stuff.
| |
| | |
| | |
| The Law of Biogenesis - Of great theoretical interest is the fact, constantly observed in studying, embryos, that the individual in its development repeats hastily and incompletely the evolutionary history of its own species. This law of recapitulation was first stated clearly by Muller in 1863, and was termed by Haeckel the law of biogenesis. In accordance with it, the fertilized ovum is compared to a unicellular organism like the Ameba: the blastula is supposed to represent an adult Volvox type; the gastrula, a simple sponge; the segmented embryo, a worm-like stage ; and the embryo with gill slits may be regarded as a fishlike stage. Moreover, the blood of the human embryo in development passes through stages in which its corpuscles resemble in structure those of the fish and reptile; the heart is at first tubular, like that of the fish, and the arrangement of blood vessels is equally primitive; the kidney of the embryo is like that of the amphibian, as are also the genital ducts. Many other examples of this law may readily be observed.
| |
|
| |
|
|
| |
|
| Some apparently useless structures appear during development, perfunctorily reminiscent of ancestral conditions; certain other parts, of use to the embryo alone, are later replaced by better-adapted, permanent organs. Representatives of either type may eventually disappear or they may persist throughout life as rudimentary organs; more than a hundred of the latter have been listed for man. Still other ancestral organs abandon their provisional embryonic function, yet are retained in the adult and utilized for new purposes. | | Some apparently useless structures appear during development, perfunctorily reminiscent of ancestral conditions; certain other parts, of use to the embryo alone, are later replaced by better-adapted, permanent organs. Representatives of either type may eventually disappear or they may persist throughout life as rudimentary organs; more than a hundred of the latter have been listed for man. Still other ancestral organs abandon their provisional embryonic function, yet are retained in the adult and utilized for new purposes. |
|
| |
|
| | | ===The Vertebrate Groups=== |
| THE VERTEBRATE GROUPS
| |
|
| |
|
| There are five vertebrate classes, the higher characterized by the possession of an enveloping embryonic membrane, called the amnion, and another embryonic appendage, known as the allantois: | | There are five vertebrate classes, the higher characterized by the possession of an enveloping embryonic membrane, called the amnion, and another embryonic appendage, known as the allantois: |
|
| |
|
| (R) Anamniota (amnion absent). | | (A) '''Anamniota''' (amnion absent). |
| | | : 1. ''Fishes'' - lamprey; sturgeon; shark; bony fishes; lung fish. |
| 1. Fishes - lamprey; sturgeon; shark; bony fishes; lung fish. | | : 2. ''Amphibia'' - salamander; frog; toad; etc. |
|
| |
|
| 2. Amphibia - ^salamander; frog; toad; etc.
| | (B) '''Amniota''' (amnion present). |
| | : 3. ''Reptiles'' - lizard; crocodile; snake; turtle. |
| | : 4. ''Birds.'' |
| | : 5. ''Mammals''. Characterized by hair and mammary glands. |
| | :: (a) Monotremes - duck-bill; primitive mammals that have a cloaca and lay eggs with shells. |
| | :: (b) Marsupials - oppossum; kangaroo; etc. The young are born immature and are sheltered in an integumentary pouch. |
| | :: (c) Placentalia. All other mammals whose young are nourished in the uterus by a placenta. |
| | ::: Ungulate series. Hoofed mammals (cattle; sheep; pig; deer; horse; etc.). |
| | ::: Unguiculate series. Clawed mammals (mole; bat; rat; rabbit; cat; dog; etc.). The highest order is the Primates (lemur; monkey; ape, man). |
|
| |
|
| {B) Amniota (amnion present).
| | ====The Vertebrate Body Plan==== |
| | All vertebrate animals are constructed in accordance with a common body plan. The distinctive characteristics of the vertebrate type include: . |
|
| |
|
| 3. Reptiles - lizard; crocodile; snake; turtle.
| | # A tubular central nervous system, dorsally placed (Fig. 4). |
| | # A notochord, between the neural tube and gut (Fig. 4). This cellular |3rimitive-axis is replaced, wholly or in part, by the vertebral column. |
| | # A pharynx, which develops paired pouches and clefts that determine the positions of important nerves, muscles and blood vessels (Fig. 91). |
| | # The position of the mouth. Unlike the condition in many invertebrates, it is not surrounded by a circumoral ring of nervous tissue which connects a dorsal - brain - with a ventral chain of ganglia. |
| | # The limbs, Two pairs, with an internal skeleton (Fig. 227). |
| | # A coelom, which is divided into a dorsal, segmental part (cavities of the somites), and a ventral, unsegmented part, partitioned by the septum transversum (diaphragm) into thoracic and abdominal portions (Fig. 4). |
|
| |
|
| 4. Birds.
| | ==Titles for Collateral Reading and Reference== |
| | |
| 5. hlammals. Characterized by hair and mammary glands, (a) Monotremes - duck-bill; primitive mammals that have a
| |
| cloaca and lay eggs with shells.
| |
| | |
| | |
| (C) Marsupials - oppossum; kangaroo; etc. The young are born immature and are sheltered in an integumentary pouch, (r)Placentalia. All other mammals whose young are nourished in the uterus by a placenta.
| |
| | |
| Ungulate series. Hoofed mammals (cattle; sheep; pig; deer; horse; etc.).
| |
| | |
| Unguiculate series. Clawed mammals (mole; bat; rat; rabbit; cat; dog; etc.). The highest order is the Primates (lemur; monkey; ape, man).
| |
| | |
| The Vertebrate Body Plan. - All vertebrate animals are constructed in accordance with a common body plan. The distinctive characteristics of the vertebrate type include: .
| |
| | |
| 1. A tubular central nervous system, dorsally placed (Fig. 4).
| |
| | |
| 2. A notochord, between the neural tube and gut (Fig. 4). This cellular |3rimitive-axis is replaced, wholly or in part, by the vertebral column.
| |
| | |
| 3. A pharynx, which develops paired pouches and clefts that determine the positions of important nerves, muscles and blood vessels (Fig. 91).
| |
| | |
| 4. The position of the mouth. Unlike the condition in many invertebrates, it is not surrounded by a circumoral ring of nervous tissue which connects a dorsal - brain - with a ventral chain of ganglia.
| |
| | |
| 5. The limbs. Two pairs, with an internal skeleton (Fig. 227).
| |
| | |
| 6. A coelom, which is divided into a dorsal, segmental part (cavities of the somites), and a ventral, unsegmented part, partitioned by the septum transversum (diaphragm) into thoracic and abdominal portions
| |
| (Fig. 4 • .
| |
| | |
| | |
| TITLES FOR COLLATERAL READING AND REFERENCE
| |
|
| |
|
| Broman. Normale und abnorme Entwicklung des Menschen. | | Broman. Normale und abnorme Entwicklung des Menschen. |
| Line 589: |
Line 428: |
| Duval. Atlas D - Embryologie. | | Duval. Atlas D - Embryologie. |
|
| |
|
| Hertwig. Handbuch der Entwicklungslehre der Wirbeltiere. | | Hertwig. [[Book - Text-Book of the Embryology of Man and Mammals|'''Handbuch der Entwicklungslehre der Wirbeltiere''']]. |
|
| |
|
| Keibel and Mall. Human Embryology. | | Keibel and Mall. [[Book - Manual of Human Embryology|'''Human Embryology''']]. |
|
| |
|
| Kellicott. A Textbook of General Embryology. | | Kellicott. [[Book - Outlines of Chordate Development|'''A Textbook of General Embryology''']]. |
|
| |
|
| Kollmann. Handatlas der Entwicklungsgeschichte des Menschen. | | Kollmann. [[Atlas_of_the_Development_of_Man_1|'''Handatlas der Entwicklungsgeschichte des Menschen''']]. |
|
| |
|
| Lillie. The Development of the Chick. | | Lillie. The Development of the Chick. |
|
| |
|
| Minot. A Laboratory Text-book of Embryology. | | Minot. [[Book - A Laboratory Text-Book of Embryology (1903)|'''A Laboratory Text-book of Embryology''']]. |
|
| |
|
| McMurrich. The Development of the Human Body. | | McMurrich. The Development of the Human Body. |
|
| |
|
| Patten. The Early Embryology of the Chick. | | Patten. [[Book - The Early Embryology of the Chick|'''The Early Embryology of the Chick''']]. |
|
| |
|
| Wilson. The Cell in Development and Inheritance. | | Wilson. The Cell in Development and Inheritance. |
|
| |
|
|
| |
|
| | | {{Arey1924 Footer}} |
| | | [[Category:1920's]] |
| | |
| CHAPTER I . THE GERM CELLS AND FERTILIZATION THE GERM CELLS .
| |
| | |
| | |
| All multicellulai' animals, except a few invertebrates, result from the union of two ripe sex cells. These are re])resentative portions of the germ plasm stored in the male and female sex glands, and are termed spermatozoon and ovinn respectively. In form and function they are quite unlike, for each is adapted to a specific purpose. It will be simplest first to describe these elements fully-formed, and then to show how they develop, mature, meet, and unite.
| |
| | |
| | |
| | |
| The Ovum. - The female germ cell, or ovum, is a typical animal cell produced in the ovary. Although always large, its exact size is correlated with the amount of stored food substance. The smallest eggs are those of the mouse and deer (about 0.07 mm.). The largest have a diameter measurable in inches (birds; a shark). Most ova are nearly spherical in form and posSess a nucleus with nucleolus, chromatin network, and nuclear membrane (Figs. 5 and 7). The nucleus is essential to the life, growth, and reproduction of the cell. The function of the nucleolus is unknown; the chromatin bears the hereditary qualities. The cytoplasm is distinctly granular and contains more or less numerous yolk granules, mitochondria, and rarely a minute centrosome.
| |
| | |
| | |
| | |
| Fig. 5. - Ovum of monkey (Prentiss). X 430.
| |
| | |
| | |
| | |
| The yolk, or deutoplasm, containing a fatty substance termed lecithin, furnishes nutriment for the developing embryo. It is doubtful if any ovum is totally devoid of yolk, yet it is useful as a basis for classifying eggs. Those ova which contain relatively little yolk, uniformly distributed, are termed isolecithal. Examples are found among various invertebrates and in all placental mammals, for such embryos either attain an independent existence quickly or are sheltered and nourished within the uterine wall of the mother. If the yolk collects at one end (called the vcgdal pole in contrast to the more jmrely jirotoplasmic animal pole) the ova are said to he telolecuhal. Many invertebrates and all vertebrates lower than the Placentalia illustrate this type. The so-called yolk of the hen - s egg (Fig. 6) is the ovum proper and its yellow color is due to the large amount of lecithin it contains. Finally, among the arthropods the yolk is centrally located and surrounded by a peripheral shell of clear cytoplasm; such eggs are centrolccithal.
| |
| | |
| | |
| | |
| | |
| | |
| Fig. 6. - Diagrammatic longitudinal section of a hen - s egg (Thomson in Heisler).
| |
| | |
| | |
| Fig. 7. - . 4 , Human ovum, approaching maturity, examined fresh in the liquor folliculi (Waldeyer). X 415. The zona pellucida appears as a clear girdle surrounded by the cells of the corona radiata. Yolk granules in the cytoplasm enclose the nucleus and nucleolus. 5 , A human spermatozoon correspondingly enlarged.
| |
| | |
| | |
| | |
| Most ova become enclosed within protective membranes, or envelopes. The vitelline membrane, secreted by the egg itself, is a primary membrane (Fig. 5). The follicle cells about the ovum usually furnish other secondary membranes, such as the zona pellucida. In lower vertebrates tertiary membranes may be added as the egg passes through the oviduct and uterus ; the albumen and shell of the hen - s egg (Fig. 6) or the jelly of the frog - s egg are of this sort.
| |
| | |
| | |
| The Human Ovum. - This is relatively of small size, measuring about 0.2 mm. in diameter (Fig. 7). It conforms closely to the isolecithal mammalian type, but has fine yolk granules somewhat condensed centrally. There is apparently a very delicate vitelline membrane, and outside it a thick, radially-striate membrane, the zona pellucida. The striate appearance is said to be due to fine canals through which nutriment is transferred from smaller follicle cells during the growth of the ovum within the ovary.
| |
| | |
| | |
| The Spermatozoon. - -In a few instances only, does the mature male element, or spermatozoon, resemble a typical cell. Most are slender, elongate structures which develop a flagellum to accomplish the active swimming that characterizes the cell. Unlike the ovum, which is the largest cell of an organism, the spermatozoon is usually the smallest. The extremes of size range from 0.018 mm. in Amphioxus to 2.0 mm. in an amphibian. The commonest shape is that of an elongate tadpole, with an enlarged head, short neck (and connecting piece), and thread-like tail (Fig. 8).
| |
| | |
| | |
| Fig. 8. - .1, Diagram of a human spermatozoon, surface view (Meves). B, Human spermatozoa, from life, in edge and surface view. X 700.
| |
| | |
| | |
| | |
| The Human Spermatozoon. - The sperm of man is of average size (0.055 mm.) and shape (Fig. 8). Compared to the ovum its volume is as i : 200,000 (Fig. 7). The /zcah is about 0.005 mm. in length. It appears oval in surface view, pear-shaped in profile. When stained, the anterior two-thirds of the^head may be seen to constitute a cap, and the sharp border of this cap is the so-called perforatorium. The head contains the nuclear elements of the sperm cell. The disc-shaped neck includes the anterior centrosonial body. The tail begins with the posterior centrosonial body and is divided into a short connecting piece, a chief piece, or flagellum, which forms about four-fifths of the length of the sperm cell, and a short end piece, or terminal filament. The connecting piece is marked off from the chief piece by the annulus. The connecting piece is traversed by the axial filament {fdvLm principale), and is surrounded: (i) by the sheath common to it and to the flagellum; (2) by a sheath containing a spiral filament; and (3) by a mitochondrial sheath. The chief piece is composed of the axial filament, surrounded by a cytoplasmic sheath, while the end piece comprises the naked continuation of the axial filament.
| |
| | |
| | |
| Atypical spermatozoa occur in some individuals. These include giant and dwarf forms, and elements with multiple heads or tails.
| |
| | |
| | |
| Comparison of the Ovum and Spermatozoon. - The dissimilar male and female sexual cells are admirably adapted to their respective functions, and illustrate nicely the modifications that accompany a physiological division of labor. Each has the same amount of chromatin, although in the sperm it is more compactly stored. The cells thus participate equally in heredity. The egg contains an abundance of cytoplasm (but nO' centrosome), and often a still greater supply of stored food. As a result, it is large and passive, yet closely approximates the typical cell. On the contrary, the sperm is small, and at casual inspection bears slight resemblance to an ordinary cell. Its cytoplasm is reduced to a bare minimum and contains no deutoplasm. Structurally, all is subordinated to a motile existence. Correlated with small size is an extraordinary increase in numbers, for the greater the total liberated the more surely will the ovum be found. Hence, apart from its role in heredity, the chief function of the spermatozoon is to seek the ovum and activate it to divide.
| |
| | |
| SPERMATOGENESIS, OOGENESIS AND MATURATION
| |
| | |
| | |
| In becoming specialized germ cells, the ovum and spermatozoon pass through parallel stages. The general process of sperm formation is designated spermatogenesis; that of egg formation, oogenesis. An essential feature of lioth is a component process, termed maturation, which is important for the following reason. Since reproduction in vertebrates depends upon the union of male and female germ cells, it is manifest that without special provision this union would necessarily double the number of chromosomes at each generation. Such progressive increase is prevented by the events of maturation. This may be defined as a form of cell division during which the number of chromosomes in the germ cells is reduced to one-half the number characteristic for the species. Its significance in the mechanism of inheritance is discussed on p. 28.
| |
| | |
| | |
| Spermatogenesis. - The spermatozoa originate in the epithelial lining of the testis tubules. Two types of cells are recognizable : the sustentacular cells (of Sertoli), and the male germ cells (Fig. 9). All the latter are descendants of primordial germ cells, which, by division, first form spermatogonia. These in turn |3roliferate and produce numerous generations of like cells. Ultimately the spermatogonia enter a growth period, at the end of which they are termed primary spermatocytes. Each contains the full number of chromosomes typical for the male of the species. Next ensues the process of maturation. This comprises two cell divisions, each primary spermatocyte producing two secondary spermatocytes, and these in turn four cells known as spermatids. During these cell divisions the number of chromosomes is reduced to half the original number in the spermatogonia.
| |
| | |
| | |
| | |
| | |
| Fig. 9. - Stages in the spermatogenesis of man arranged in a composite to represent a portion of a seminiferous tubule sectioned transversely. X 900.
| |
| | |
| | |
| | |
| The spermatids now attach to Sertoli cells, from which they appear to receive nutriment, and become transformed into mature spermatozoa (Fig. 10). The nucleus forms almost all the head; the centrosome divides, the resulting particles passing to the extremities of the neck. The posterior centrosome differentiates the annulus and is prolonged to become the axial filament. The cytoplasm forms the sheaths of the neck and tail, whereas the spiral filament of the connecting piece is derived from cytoplasmic mitochondria. When the transformation is complete, the spermatozoa detach from the sustentacular cells and are set free in the lumen of the seminiferous tubule.
| |
| | |
| | |
| | |
| Maturation in Ascaris. - The way the number of chromosomes is reduced may be seen in the spermatogenesis of Ascaris (Fig. 1 1). Four chromosomes are typical for Ascaris megalocephala hivalens, and each spermatogonical cell contains this number. In the early prophase of the primary spermatocyte there appears a spireme thread consisting of four parallel rows of granules (B). This thread breaks in two and forms two quadruple structures, known as tetrads (D-F) ; each is equivalent to two original chromosomes, paired side by side and split lengthwdse to make a bundle of four. At the metaphase {G}, a tetrad divides into its two original chromosomes which already show evidence of longitudinal fission and are termed dyads. One pair of dyads goes to each of the daughter cells, or secondary spermatocytes (G-I). Without the formation of a nuclear membrane, the second maturation spindle appears at once, the two dyads split into four monads, and each daughter spermatid receives two single chromosomes (monads), or one-half the number characteristic for the species. The tetrad, therefore, represents a precocious division of the chromosomes in preparation for two rapidly succeeding cell divisions which occur without the intervention of the customary resting periods. The easily understood tetrads are not formed in most animals, although the outcome of maturation is identical in either case. A diagram of maturation is shown in Fig. 12. The first maturation division in Ascaris is probably reductional, each daughter.
| |
| | |
| | |
| | |
| | |
| | |
| Fig. 10. Diagrams of the development of spermatozoa (Meves in Lewis and Stohr). a.c, Anterior centrosome; a.f., axial filament; c.p., connecting piece; ch.p., chief piece; g.c., cap; «., nucleus; nk., neck; p., cytoplasm; p.c., posterior centrosome.
| |
| | |
| | |
| | |
| Fig. 11. - Reduction of chromosomes in the spermatogenesis of Ascaris megalocephala bivalens (Brauer in Wilsonj. X about i loo. A-G, Successive stages in the division of the primary spermatocyte. The original reticulum undergoes a very early division of the chromatin granules which then form a quadruply split spireme (B, in profile). This becomes shorter (C, in profile), and then breaks in two to form two tetrads (D, in profile), (E, on end). F, G, H, first division to form two secondary spermatocytes, each receiving two dyads. I, Secondary spermatocyte. J, K, The same dividing. L, Two resulting spermatids, each containing two monads or chromosomes.
| |
| | |
| | |
| | |
| nucleus receiving two complete chromosomes of the original four, whereas in the second maturation division, as in ordinary mitosis, each daughter nucleus receives a half of each of the two chromosomes, these being split lengthwise. The latter division is equauonal and the daughter nuceli receive chromosomes bearing similar hereditary qualities.
| |
| | |
| Some animals reverse the sequence of events, reduction occurring at the second maturation division.
| |
| | |
| | |
| | |
| Maturation in Man - All spermatogonia, like the somatic cells, contain 48 chromosomes. The primary spermatocytes form tetrads and their division separates the mated chromosomal pairs into 24 single chromosomes of the secondary spermatocyte. Hence, this mitosis is reductional. The secondary spermatocytes then divide equationally into spermatids, each of which also contains 24 single chromosomes. Transformation into spermatozoa ensues (Figs. 9 and 10). Those details of maturation which pertain to sex determination are explained on p. 29.
| |
| | |
| | |
| | |
| | |
| | |
| Fig. 12. - Diagrams of maturation in spermatogenesis and oogenesis (Boveri).
| |
| | |
| | |
| | |
| Oogeneiss. - The ova, like the male elements, arise from the multiplication of primordial germ cells in the ovary (cf. p. 156). At birth, or shortly after, human ova cease forming. The number at this time in both ovaries has been placed between 100,000 and 800,000. Cellular degeneration reduces this supply until, at 18 years, the total is from 35,000 to 70,000 and several years after the menopause no more are to be found.
| |
| | |
| | |
| Late in fetal life, indifferent cells, by surrounding the young ova ioogonia) of the cortex, produce primordial follicles (Fig. 13 A). Some begin growth at once, others are quiescent until childhood or adult life is attained. During the slow growth period, the small, nutritive follicle cells increase in number and the oogonium gains greatly in size. When the follicle cells are several layers deep, a cavity appears between them. This enlarges, and there reSults a sac, the vesicular, or Graafian follicle, filled with fluid, the liquor folliculli (Fig. 13 5 ). As growth continues, the oogonium becomes located more and more eccentrically until it lies at one side of the follicle, buried in a mound of follicular cells termed the cumulus odphorus (egg-bearing hillock) (Fig. 14). Around the stratified follicle cells, now designated the stratum granulosum, there is differentiated from the stroma of the ovary the theca folliculi. This is composed of an inner, vascular tunica interna, and an outer, fibrous and muscular tunica externa.
| |
| | |
| | |
| | |
| | |
| Fig. 13. A, Two primordial human follicles and one early in growth (De Lee). X200. B, Section of a human ovarian cortex with ten primordial follicles and one young Graafian follicle (Piersol). X 90.
| |
| | |
| | |
| .At the end of the growth period, the follicle has enlarged from a structure 0.04 to 0.06 mm. in diameter to one 5 to 12 mm. (Fig. 16 A); similarly, the primordial ovum measured 0.04 to 0.05 mm. whereas it now has a diameter of about 0.2 mm. In harmony with the terminology for the male cell, the grown oogonium is designated a primary oocyte. The final stages of oogenesis are maturative. As in spermatogenesis, two cell divisions take place, but with this difference: the cytoplasm is divided unequally, and instead of four cells of equal size resulting, there are formed one large ripe ovum, or ootid, and three rudimentary or abortive ova, known as polar bodies, or polocytes (Fig. 15). The number of chromosomes is reduced in the same manner as in the male, so that the ripe ovum and each polar cell contain one-half the number of chromosomes found in the oogonium or primary oocyte.
| |
| | |
| | |
| | |
| Fig. 14. - An advanced Graafian follicle and ovum from a girl of fifteen (Prentiss). X 30.
| |
| | |
| | |
| Fig. 15. - . 4 , Formation of the first polar cell in the mouse ovum (Sobotta). X 1500. B, Separation of the second polar cell in the bat ovum (after Van der Stricht).
| |
| | |
| | |
| During maturation the ovum and first polocyte are termed secondary oocytes (comparable to secondary spermatocytes) ; the mature ovum (ootid) and second polocyte, with the daughter cells of the first polocyte. are comparable to the spermatids (Fig. 12). Each spermatid, however may form a mature spermatozoon, but only one of the four daughter cells of the primary oocyte becomes functional. The ovum develops at the ex])ense of the three ])olocytes which are abortive and degenerate eventually, though it has been shown that in some insects the polar cell may be fertilized and segment several times like a normal ovum. In most animals, the actual division of the first polocyte into two daughter cells is suppressed (cf. Fig. 1 5 B). The nucleus of the ovum after maturation is known as the jcniale pronndcus.
| |
| | |
| | |
| Maturation in the Mouse. - Typical maturation occurs in the mouse. 1 'he first jiolocyte is formed while the ovum is still in the Graafian follicle. Neither astral rays nor typical centrosomes have been observed; the chromosomes are V-shaped. The finst polar cell is constricted from the ovum and lies beneath the zona pellucida as a spherical mass about 25 micra in diameter (Fig. 15 A). Both ovum and polar cell (secondary oocytes) contain 20 chromosomes, or half the number normal for the mouse. The first maturation division is the reductional one and the chromosomes take the form of tetrads.
| |
| | |
| | |
| After ovulation has taken place, the ovum lies in the ampulla of the uterine tube. If fertilization occurs, a second polocyte is cut off, the nucleus of the ovum not having regained its membrane between the production of the first and second polar bodies (Figs. 15 A and 17 A, D). The second maturation spindle and second polar cell are smaller than the first. Immediately after the appearance of the second polar cell, the chromosomes resolve themselves into a reticulum and the female pronucleus is complete [Mig. 17 D).
| |
| | |
| | |
| Maturation in Man. - -The only observations are those of Thompson (1919), who believes to have identified stages in the formation of all three polar cells prior to ovulation or fertilization. The evidence presented, however, can hardly be accepted as conclusive. Yet, in Tarsius, a low primate, both polar cells have been observed.
| |
| | |
| | |
| OVULATION AND INSEMINATION
| |
| | |
| The ripe germinal products are next released from their respective sex glands and then brought together.
| |
| | |
| Ovulation. - The discharge of the ovum from its follicle comprises ovulation. A few animals breed continuously, but commonly there is a seasonal or annual spawning period. The several mammalian groups show various gradations between an almost continuous breeding period (oestrus) and an annual one. In man ovulation is periodic, at intervals of four weeks, beginning at puberty and ending with the menopause. However, fully formed Graafian follicles appear in the ovary during the second year of infancy, and, in some individuals, even before birth.
| |
| | |
| | |
| Ovulation may occur at this time, but usually these precociously formed follicles degenerate with their contained ova. Generally, only one follicle and ovum mature each month, the ovaries roughly alternating. Yet, ordinary multiple births depend on the rupture of two or more follicles. Rarely in man, but frequently in the monkey, follicles contain more than one egg. Thus, from the thousands of potential ova, only about 200 ripen in each ovary during the 30 years of sexual activity.
| |
| | |
| | |
| The completed follicle is from 5 to 12 mm. in diameter. It makes a bud-like protuberance from the surface of the ovary, and at this point the ovarian wall is very thin (Fig. 16 A). Internally, the follicle contains fluid, probably under vascular and muscular tension. The precise factors which cause rupture are not positively known, but they doubtless include mechanical pressure, perhaps combined with a weakening of the follicular wall by the digestive influence of the contained fluid (Schochet, 1920).
| |
| | |
| | |
| Fig. 16. A , Human uterine tube and ovary with mature Graafian follicle] (RibemontDessaignes). B, Sectioned human ovary with a corpus luteum verum and two corpora albicantia. X 1.5.
| |
| | |
| | |
| | |
| When the follicle bursts, the fluid gushes out, carrying with it the ovum torn loose from its cumulus oophorus. The adhering follicular cells, immediately investing the ovum, constitute the corona radiata (Fig. 6). The ovum is swept into the uterine tube by inwardly stroking cilia of the tubal Ambriae. Although the ovum is now ready to be fertilized, it is not yet technically - mature, - for the last polar division awaits the stimulus of fertilization.
| |
| | |
| | |
| The Corpus Luteum - After ovulation, a blood clot, the corpus liemorrhagicum, forms within the empty follicle. The follicle cells of the stratum granulosum proliferate, enlarge, and produce a yellow pigment. The w'hole structure, composed of lutein cells and connective-tissue strands, is termed the corpus luteum, or yellow body (Fig. 16 B). If pregnancy does not supervene, the corpus luteum spurium reaches its greatest development within two weeks and then gradually is replaced by fibrous tissue ; the resultant white scar is known as the corpus albicans. In pregnancy the corpus /zhtvuKuer/oH continues its growth until, at the thirteenth week, it reaches a maximal diameter of 1 5 to 30 mm. ; at term it is still a prominent structure in the ovary. The corpus luteum is believed to produce an important internal secretion, for if removed the ovum fails to attach to the wall of the uterus, or if the ovum is already embedded, development ceases (Fraenkel). An influence in retarding ovulation and stimulating the mammary gland function has also been shown experimentally (L. Loeb; O - Donoghue).
| |
| | |
| | |
| | |
| Relation of Ovulation and Menstruation. - Since human ovulation and menstruation both begin with puberty, recur at about twenty-eight day intervals, and discontinue during pregnancy and at the menopause, a close relation has long been inferred. The cessation of the menses after ovarian removal further indicates dependence. For many years the two processes were supposed to be synehronous. This belief was based upon clinical oliservations by Leopold, Ravano and others who tried to correlate the ages of corpora lutea with known menstrual histories. Since then, Meyer, Ruge, Schroder, Fraenkel, and Halban, utilizing better standardized corpora lutea, have presented convincing evidence that ovulation occurs most often between the fourth and fourteenth day after the menstrual onset. While correct as a generalization, this correlation is not rigid and often ova are liberated at other times. Moreover, in young girls ovulation may precede the inception of menstruation and it may occur in women during pregnancy and lactation or after the menopause.
| |
| | |
| | |
| | |
| Coitus and Insemination. - In most aquatic animals the eggs and sperm are discharged externally at about the same time and place. Their meeting depends largely upon chance, enhanced by the production of immense numbers of spermatozoa. Some animals increase the certainty of such cell union by a pscudocopulation ; thus, the male frog clasps the female and jiours his milt over the eggs as they are extruded. Many invertebrates and all amniote vertebrates have their sex cells unite inside the female - s body. This is effected by the sexual embrace termed copulation, or coitus. In general, those animals whose offspring reach maturity with reasonable surety (as the result of internal fertilization and postnatal care) produce fewer germ cells, especially ova, than those that leave fertilization to chance and development to hazard. The codfish produces 10,000,000 eggs in a breeding period, a sea urchin 20,000,000; in certain birds and mammals only a single egg is matured, yet the stock of each remains constant.
| |
| | |
| | |
| The purpose of coitus is to introduce spermatozoa into the vagina. The completed human sperm detach from the Sertoli cells, and clusters are moved along the efferent ductules into the epididymis. Here they become separate and motile, due to a secretion of the duct epithelium. The seminal fluid accumulates about the ampulla of the ductus deferens; its storage in the seminal vesicles is much questioned. At the climax of coitus ejaculation occurs and the spermatozoa, suspended in seminal fluid, are forcibly ejected. The seminal fluid, or semen, is a mixture chiefly of the secretions of the seminal vesicles, prostate, and bulbourethal glands, in which occur the spermatozoa. The volume of the ejaculate is about 3 c.c. and in it swim over 200,000,000 spermatozoa.
| |
| | |
| | |
| The outstanding functional feature of spermatozoa is their flagellate swimming. Because of this they were once regarded as parasites living in the seminal fluid. Forward progress is at the rate of about 2.5 mm. a minute, which, length for length, compares with the ordinary gait of man. An acid environment, such as the vagina, is deleterious or fatal; an alkaline medium, as furnished by the uterus, is favorable. Spermatozoa tend always to swim against feeble currents. This is important, as the outwardl^^ stroking cilia of the uterine tubes and uterus direct the spermatozoa by the shortest route to the ovum. They probably reach the ampulla of the uterine tube two hours or more after coitus.
| |
| | |
| | |
| Spermatozoa have been found motile in the uterine tube nine days after the admission of a patient to the clinic, and, according to her statement, three and one-half w^eeks after coitus. They have been kept alive eight days outside the body. It is not known for how long spermatozoa are capable of fertilizing ova. Keibel holds that this would certainly be more than a week. However, Lillie (1915) has shown with sea urchins that the ability to fertilize is lost long before vitality or motility is impaired, and Mall (1918) concludes that the duration of the fertilizing power of human spermatozoa is safely less than the corresponding period in the ovum, which is probably for fully 24 hours after ovulation. In the hen, spermatozoa remain functional three weeks; in bats six months; in bees five years.
| |
| | |
| | |
| | |
| FERTILIZATION
| |
| | |
| The formation, maturation, and meeting of the male and female germ cells are all preliminary to their actual union which definitely marks the beginning of a new individual. This penetration of ovum by spermatozoon and the fusion of their - pronuclei - constitute the process oi jertilization. In practically all animals, fertilization also starts the ovum dividing and thus initiates development in the ordinary sense. A few invertebrates, however, can develop without the aid of fertilization; this method is styled parthenogenesis, and in such eggs there is usually but one polar cell and hence no chromosome reduction.
| |
| | |
| Random movements of the sperm bring them in contact with ova. It is very doubtful whether there is any chemical attraction. In some forms, as for example fishes, tactile response keeps -the spermatozoa in contact with anything touched. In mammals, amphibia, and many invertebrates, the ovum is either naked or surrounded by a delicate vitelline membrane. Spermatozoa can enter such eggs at any point. Ova that are invested with heavy membranes usually have a definite funnel-shaped aperture, the micro pyle, through which the male cell must enter. Only motile spermatozoa are able to attach to the surface of an egg; it is probable that forces allied to phagocytosis, rather than vibrational energy, accomplish the actual - penetration. -
| |
| In general, only one spermatozoon normally enters an egg; how others, endeavoring to penetrate, are thereafter excluded is not entirely clear. If accident or im]iaired vitality admits more than one sperm, development is abnormal and soon ends. On the contrary, some sharks, amphibia, reptiles, and birds normally exhibit such polyspermy. In all these cases, however, only one spermatozoon unites with the female pronucleus.
| |
| | |
| | |
| The fertilized ovum derives its nuclear substance equally from both parents, the cytoplasm (and yolk) almost entirely from the mother, the centrosome probably from the father.
| |
| | |
| | |
| The fundamental results of fertilization are: (i) the union of male and female pronuclei to form the cleavage nucleus (thus restoring the original number of chromosome pairs); (2) the initiation of cell division, â– or cleavage, in which all male and female chromosomes take part.
| |
| | |
| | |
| These two factors are separate and independent phenomena. It has been shown by Boveri and others that fragments of sea urchin - s ova containing no part of the nucleus may be fertilized by spermatozoa, segment, and develop into larvae. The female chromosomes are thus not essential to the process of cleavage. Loeb, on the other hand, proved that the ova of invertebrates may be made to develop by chemical and mechanical means without the cooperation of the spermatozoon {artificial parthenogenesis). Even adult frogs have been reared from mechanically stimulated eggs. These facts show that the actual union of the male and female pronuclei is not the means of initiating the development of the ova. In all vertebrates it is, nevertheless, the end and aim of fertilization.
| |
| | |
| | |
| Lillie maintains that the cortex of a sea urchin - s ovum produces a substance, fertilizin. This he regards as an amboceptor essential to fertilization, with one side chain which agglutinates and attracts the spermatozoa, and another side chain which activates the cytoplasm and initiates the cleavage of the ovum. According to Loeb, agglutination is proved in but few forms and Lillie - s interpretation fails to meet all the facts. Loeb holds that the spermatozoon actually activates the ovum to develop by increasing its oxidations and by rendering it immune to the toxic effects of oxidation.
| |
| | |
| | |
| Fertilization in the Mouse. - Normally, a single spermatozoon enters the ovum six to ten hours after coitus. While the second polar cell is forming, the spermatozoon penetrates the ovum and loses its tail (Fig. 17 A-C). Its head enlarges and is converted into the male projiuclcits (D). The pronuclei, male and female, approach (E) and resolve first into a spireme stage (F), then into two groups of 20 chromosomes (G). A centrosome, possibly that of the male cell (cf. Fig. 15 B), appears between them, divides into two, and soon the first cleavage spindle is formed (F-H). The 20 male and 20 female chromosomes arrange themselves in the equatorial plane of the spindle, thus making the original number of 40 (H). Fertilization is now complete and the ovum divides in the ordinay way ( 7 , /), the daughter cells each receiving equal numbers of maternal and paternal chromosomes.
| |
| | |
| | |
| Fertilization in Man. - The union of the human germ cells is believed usually to take place in the ampulla of the uterine tube, although it never has been observed in any primate except Tarsius. This conclusion is supported by direct observations on other mammals and by the frequency of tubal pregnancies at this site. Rarely ova become fertilized before entering the tube, but the possibility of fertilization after they have reached the uterus is usually denied.
| |
| | |
| | |
| | |
| Fig. 17. - Fertilization of the ovum of the mouse (Sobotta). X 500. A-D, Entrance of the spermatozoon and formation of the polar cells; D-E, development of the pronuclei ; F - J, union of chromosomes and the first cleavage spindle.
| |
| | |
| | |
| | |
| To be fruitful, the time of coitus and ovulation must roughly agree (p. 22), and, on the average, about one day is supposed to elapse between insemination and fertilization. Most conceptions occur during the week or ten days following menstruation; this is in harmony with the known data on ovulation time (p. 24).
| |
| | |
| | |
| While there are no direct observations on fertilization in man, the ]irocess has been studied throughly in several mammals. In all essentials it undoubtedly follows the common course as described for the mouse.
| |
| | |
| | |
| Superfetation.- If an ovum is liberated by a pregnant woman and fertilized at a later coitus, it may develop into a second, younger fetus. This rare condition, called sit perjctation, is often denied, yet in the early weeks of ])regnancy it is theoretically possible. Superfetation should not be confused with strikingly unequal twin development, due to nutritional or other inequalities.
| |
| | |
| | |
| | |
| HEREDITY AND SEX
| |
| | |
| | |
| The Significance of Mitosis and Maturation. - The complicated processes of mitosis serve the purpose of dividing accurately the chromatic substance of the nucleus in such a way that the self-pcrjietuating chromosomes of each daughter cell may be the same, both quantitatively and ciualitatively. This is important since it is believed by most students of heredity that chromatin particles, or genes, in the chromosomes bear the hereditary characters, and that these are arranged in definite linear order in particular chromosomes. At maturation there is a side by side union of like chromosomes, one member of each pair having come from the father, the other from the mother of the preceding generation; each member, however, carries the same general set of hereditary charaeters as its mate. At this stage of chromosomal conjugation there may be an interchange, or - crossing over, - of corresponding genes, resulting in new hereditary combinations. The reducing division of maturation separates whole chromosomes of each pair, but chance alone governs the actual assortment of paternal and maternal members to the daughter cells; this mitosis obviously halves the chromosome number characteristic for the species. The significance of the ecjuational maturation mitosis, beyond accomplishing mere cellular multiplication, is obscure.
| |
| | |
| | |
| | |
| Mendel’s Law of Heredity. - Experiments show that hereditary characters fall into two opposing groups, the contrasted pairs of which are termed allelomorphs. As an example, we may take the hereditary tendencies for dark and blue eyes. It is believed that there are paired chromatic particles, or genes, which are responsible for these hereditary tendencies, and that paired spermatogonial chromosomes bear one each of these genes. Each chromosome pair in separate germ cells may possess similar genes, both bearing dark-eyed tendencies or both blue-eyed tendencies, or opposing genes, bearing the one dark-, the other blue-eyed tendencies. It is assumed that at maturation these paired genes are separated along with the chromosomes, and that one only of each pair is retained in each germ cell.
| |
| | |
| | |
| In our example, either a blue-eyed or a dark-eyed tendency-bearing particle would be retained. At fertilization, the segregated genes of one sex may enter into new combinations with those from the other sex. Three combinations are possible. If the color of the eyes be taken as the hereditary character: (i) two - dark - germ cells may unite; (2) two - blue - germ cells may unite; (3) a - dark - germ cell may unite with a - blue - germ cell. The offspring in (i) will all have dark eyes, and, if interbred, their progeny will likewise inherit dark eyes exclusively. Similarly, the offspring in (2), and if these are interbred their progeny as well, will include nothing but blue-eyed individuals. The first generation from the cross in (3) will have dark eyes solely, for black in the present example is dominant, as it is termed. Such dark-eyed individuals, nevertheless, possess both dark- and blueeyed bearing genes in their germ cells; in the progeny resulting from the interbreeding of this class, the original condition is repeated - pure darks, impure darks which hold blue recessive, and pure blues will be formed in the ratio of 1:2:1 respectively. It is thus seen that blue-eyed children may be born of dark-eyed parents, whereas blue-eyed parents can never have dark-eyed offspring. Many such allelomorphic pairs of hereditary characters are known.
| |
| | |
| | |
| Cytoplasmic Inheritance. - Certain eggs show distinct cytoplasmic zones which cleavage later segregates into groups of cells destined to form definite organs or parts. In a sense this represents a refined sort of preformation, but prelocalization is a more exact term. From these facts Conklin and Loeb argue that the cytoplasm is really the embryo in the rough, the nucleus, through Mendelian heredity, adding only the finer details. Morgan, among others, refuses to admit the validity of this interpretation.
| |
| | |
| | |
| The Determination of Sex. - The sex-determining power lies in a chromosome that can be identified in many animals. This chromosome is termed the accessory, X, or sex chromosome. According to Painter (1923), humair obgonia contain 46 ordinary chromosomes and two X -chromosomes. At maturation the number is halved, and all oocytes and polocytes contain 23 -f X. The spermatogonia, on the contrary, contain 46 ordinary chromosomes, one X-chromosome and its diminutive mate, called the Y-chromosome. After maturation, therefore, half the spermatids have 23 -j- X, the remaining half have 23 -p y. When a spermatozoon with 23 + X fertilizes an ovum, the number is restored to 46 -p 2X and a female results. When a spermatozoon with 23 -p Y fertilizes, the outcome is 46 -p X -p Y and a male results.
| |
| | |
| | |
| Many animals lack the Y, and the male cells contain an odd number of chromosomes. Reduction then forms two classes of spermatozoa, those with the extra chromosome being female producing. In certain birds and moths the system is the exact reverse, inasmuch as the spermatozoa are all alike in chromosomal constitution while the eggs are of two sorts.
| |
| | |
| | |
| | |
| | |
| | |
| | |
| ++++++++++++++++++++++++++++++++.
| |
| | |
| Chapter II Cleavage And The Origin Of The Germ Layers
| |
| | |
| | |
| Cleavage
| |
| | |
| The fertilized ovum promptly begins to form the new, multicellular individual by a process termed cleavage, or segmentation . This comprises orderly and rapid successions of mitoses which result in an aggregate of smaller cells, called blastomercs. Every blastomere receives the full assortment of chromosomes, half from each parent (Fig. 17 F-J).
| |
| | |
| The abundance and distribution of yolk in the egg so influences mitosis as to allow the following classification of cleavage:
| |
| (A) Total. Entire ovum divides; holoblastic ova.
| |
| | |
| 1. Equal. In isolecithal ova; blastomeres are of equal size; e.g., amphioxus and mammals.
| |
| | |
| | |
| 2. Unequal. In moderately telolecithal ova; yolk accumulated at vegetal pole retards mitosis, and fewer but larger blastomeres form there; e.g., lower fishes and amphibia.
| |
| | |
| | |
| {B) Partial. Protoplasmic regions alone cleave; meroblastic ova.
| |
| | |
| 1. Discoidal. In highly telolecithal ova; mitosis restricted to anim^d ]Jole; e.g., higher fishes, reptiles, and birds.
| |
| | |
| 2. Superficial. In centrolecithal ova; mitosis restricted to the peri])heral cytoplasmic investment ; arthropods.
| |
| | |
| | |
| | |
| | |
| Cleavage in Amphioxus. - The early processes of development are easily understood in a primitive, fish-like form, Amphioxus. About one hour after fertilization, its essentially isolecithal ovum divides vertically into two nearly equal blastomeres (Fig. 18, 2). Within the next hour the daughter cells again cleave in the vertical plane, at right angles to the first division, thus forming four cells (3). Fifteen minutes later a third division takes place in a horizontal plane (4). As the yolk is somewhat more abundant at the vegetal poles of the four cells, the mitotic spindles lie nearer the animal pole. Consequently, in the eight-celled stage the upper tier of four cells is slightly smaller than the lower four. By successive cleavages, first in the vertical, then in the horizontal ]3lane, a 16- and 3 2 -celled embryo is formed (5, 6). The upper two tiers are now smaller, and a cavity, the blastocode, is enclosed by the cells. The embryo at this stage is sometimes called a morula because of its resemblance to a mulberry. In subsequent cleavages, as development proceeds, the size of the cells is diminished, while the cavity enlarges (7, 8). The embryo is now a blastnla, nearly spherical in form and about four hours old. The cleavage of the holoblastic Amphioxus ovum is thus total and nearly equal.
| |
| | |
| | |
| Fig. 18. - Cleavage in Amphioxus, viewed laterally (Hatschek). X 200. i. Mature egg, with one polar body (P. 5 .) ; the other missing. 2. Ovum partly divided into two blastomeres. 3. Four blastomeres. 4. Eight blastomeres. 5. Sixteen blastomeres. 6. Thirty-two blastomeres, hemisected to show the blastocoele, B. 7, 8. Total and hemisected blastul®.
| |
| | |
| | |
| | |
| | |
| Cleavage in Lower Fishes and Amphibia. - These ova contain enough yolk so that the nucleus and most of the cytoplasm lie nearer the upper, or animal pole. The first cleavage spindle appears eccentrically in this cytoplasm. The first two cleavage planes are vertical and at right angles, and the four resulting cells are equal. The spindles for the third cleavage are located near the animal pole, and the division takes place in a horizontal plane. As a result, the upper four cells are much smaller than the lower four (Fig. ig A). The large, yolk-laden cells divide more slowly than the upper, small cells (B-D). At the blastula stage, the cavity is small, and the cells of the vegetal pole are many times larger than those of the animal pole {E, F). The cleavage is thus total but unequal vertical but the inert yolk does not cleave. The segmentation is thus partial and discoidal. In the bird - s ovum, the cytoplasm is divided by successive vertical furrows into a mosaic of cells, which, as it increases in size, forms a cap-like structure upon the surface of the yolk (Fig. 20 A). These cells are separated from the yolk beneath by horizontal cleavage.
| |
| | |
| | |
| | |
| Fig. 20. - Cleavage of the pigeon - s ovum (redrawn from Blount). A, Blastoderm in surface view; B, in vertical section.
| |
| | |
| | |
| | |
| Fig. 19. - Cleavage and gastrulation in the frog. X 12. A-D, Cleavage stages; E, blastula; F, hemisection of E; G, early gastrula; u, hemisection of G. an., Animal cells; arch., archenteron; b'c., blastocoele; b - p., blastopore; ect., ectoderm; ent., entoderm; v - g., vegetal cells.
| |
| | |
| | |
| | |
| Cleavage in Higher Fishes, Reptiles and Birds. - The ova of these vertebrates contain a large amount of yolk. There is very little pure cytoplasm except at the animal pole, and here the nucleus is located (Fig. 6). When segmentation begins, the first plane of separation is furrows, and successive horizontal cleavages give rise to several layers of cells (Fig. 20 B). The space between cells and yolk mass may be compared to the blastula cavity of Amphioxus and the frog (Fig. 22). The cellular cap is termed the germinal disc, or blastoderm. The yolk mass, which forms the floor of the blastula cavity and the greater part of the ovum, may be compared to the large, yolk-laden cells at the vegetal pole of the frog - s blastula. The main yolk mass never divides but is gradually used up in supplying nutriment to the embryo which is developed from the cells of the germinal disc. At the periphery of the blastoderm, new cells form progressively until they enclose the yolk (Fig. 22 C).
| |
| | |
| | |
| | |
| Fig. 21. - Diagrams of cleavage ami the blastodermic vesicle in the raut^it (Thomson, after van Beneden). X 200.
| |
| | |
| | |
| | |
| Cleavage in Mammals. - The ovum of all the higher mammals, including man, is isolecithal and nearly microscopic in size. Its cleavage has been studied in several forms, but the rabbit - s ovum will serve as an example. The cleavage is complete and nearly equal (Fig. 21), a cluster of approximately uniform cells being formed within the zona pellucida. This corresponds to the morula stage of Amphioxus. Next, an inner mass of cells is formed that is equivalent to the germinal disc, or blastoderm of the chick embryo. The inner cell mass is overgrown by an outer layer which is termed the trophectoderm, because it later supplies nutriment to the embryo from the uterine wall. Fluid then appears between the outer layer and the inner cell mass, thereby separating the two except at the animal pole. As the fluid increases in amount, a hollow blastodermic vesicle results, its walls composed of the single-layered trophectoderm, except where this is in contact with the inner cell mass. It is usually spherical or ovoid in form, as in the rabbit, and probably such is the form of the human ovum at this stage. In the rabbit, the vesicle is 4.5 mm. long before it becomes embedded in the wall of the uterus; among ungulates, or hoofed animals, the vesicle is greatly elongated and attains a length of several centimeters, as in the pig.
| |
| | |
| | |
| | |
| Fig. 22 . - Diagrams of blastula homologies (Prentiss). . 4 , Amphioxus; B, frog; C, chick; D, mammal.
| |
| | |
| | |
| | |
| | |
| Comparing the mammalian blastodermic vesicle with the blastula stages of Amphioxus, the frog, and the bird, it will be seen that it is to be homologized with the bird - s blastula, not with that of Amphioxus (Fig. 22). In each case there is an inner cell mass of the germinal disc. The trophectoderm of the mammal represents a ])recocious development of cells, which, in the bird, later enveloji the yolk. The cavity of the vesicle is to be compared, not with the blastula cavity of Amphioxus and the frog, but with the yolk mass pins the cleft-like blastocoele of the bird's ovum. The higher mammalian ovum, although almost devoid of yolk, thus develops a - blastula - resembling that attained by the yolk-laden ova of reptiles and birds. That this similarity has an evolutionary significance is attested by discoidal cleavage in the highly telolecithal eggs of present-day monotreme mammals.
| |
| | |
| | |
| In the low primate Tarsius, cleavage and the blastodermic vesicle are well known. A four-celled Macacus ovum, with blastomeres nearly equal and oval in form, is the only cleavage stage yet observed among higher jirimates. In all placental mammals, segmentation of the ovum occurs during its passage down the uterine tube.
| |
| | |
| | |
| | |
| | |
| | |
| | |
| THE FORMATION OF ECTODERM AND ENTODERM (GASTRULATION)
| |
| | |
| | |
| The blastula and early blastodermic vesicle show no differentiation into layers. Such differentiation next takes place, giving rise first to the ectoderm and entoderm, and finally to the mesoderm. From these three primary germ layers all tissues and organs of the body are derived.
| |
| | |
| The processes of gastridation, by which ectoderm and entoderm arise, and of mesoderm formation will be treated separately.
| |
| | |
| Amphioxus and Amphibia. - The larger cells at the vegetal pole of the Amphioxus blastula fold inward (Fig. 23 A, B). Eventually, these invaginating cells obliterate the blastula cavity and come in contact with the outer layer (Fig. 23 C). The new cavity, thus formed, is the primitive gut, or archenteron, and its narrowed mouth is the blastopore. The outer layer of cells is the ectoderm, the inner, newly formed layer is the entoderm. The entodermal cells are henceforth concerned in the nutrition of the body. The embryo is now termed a gastrula (little stomach).
| |
| | |
| | |
| In amphibia, invagination begins at the junction of animal and vegetal cells (Fig. ig G). Externally, the blastopore appears as a crescentic groove. Since the vegetal cells are large and the blastocoele is relatively small, simple invagination fails. Hence, archenteron formation is aided by a lip-like overgrowth of rapidly dividing cells from the animal pole (Fig. ig H).
| |
| | |
| | |
| Fig. 23. - Gastrulation in Amphioxus. X 200.
| |
| | |
| | |
| | |
| Reptiles and Birds. - ^The germinal disc, or blastoderm, in these animals lies like a cap on the surface of inert yolk (Fig. 6). Since the enormous amount of yolk makes gastrulation as in Amphioxus and amphibians impossible, the process exhibits marked modifications.
| |
| | |
| | |
| There appears caudally on the blastoderm of reptiles a pit-like depression. From this invagination, a proliferation of cells forms a layer which spreads beneath the ectoderm. The inner layer, originating in this manner, is the entoderm, and the region of the pit, where ectoderm and entoderm are continuous, is the blastopore. In Fig. 27 A these changes are complete.
| |
| | |
| | |
| | |
| Fig. 24. - Gastrulation in the pigeon, as shown by a longitudinal section of the blastoderm (redrawn after Patterson). X 50.
| |
| | |
| | |
| In birds, the caudal portion of the blastoderm is rolled or tucked under, the inner layer formed in this way constituting the entoderm (Fig. 24). The marginal region, where ectoderm and entoderm meet, bounds the blastopore, while the space between entoderm and yolk is the archenteron.
| |
| | |
| | |
| | |
| Mammals. - Cells on the under surface of the inner cell mass become arranged in a definite sheet, the entoderm (Fig. 2^ A). It is usually said to arise by s])litting, or delamination, although there are attempts to prove ingrowth from a - blastopore. - In most mammals, the entoderm spreads rapidly and lines the blastodermic vesicle (Fig. 38) but in Tarsius, the entoderm forms a much smaller sac (Fig. 25 B, C). The youngest human embryos known (Fig. 40) indicate a previous origin of entoderm much as in Tarsius.
| |
| | |
| | |
| | |
| | |
| Fig 23. - Gastrulation in the low primate, Tarsius, as demonstrated by sections of the blastodermic vesicle (redrawn after Hubrecht). X 260.
| |
| | |
| | |
| | |
| ORIGIN OF THE MESODERM, NOTOCHORD AND NEURAL TUBE .
| |
| | |
| | |
| Amphioxus and Amphibia. - The dorsal portion of the inner sheet, which forms the roof of the archenteron in Amphioxus, gives rise to paired, lateral diverticula, the ccdonuc pouches (Fig. 26). These separate both from a mid-dorsal plate of cells (the future notochord), and from the entoderm of the gut, and become the primary mesoderm. The mesodermal pouches grow ventrad and their cavities form the coelom, or body cavity. Their outer layers, with the ectoderm, constitute the body wall, or somatopleure; their inner layers, with the gut entoderm, form the intestinal wall, or splanchnopleure. In the meantime, a dorsal plate, cut off from the ectoderm, folds into the neural tube (anlage of the nervous system), and the notochordal plate becomes a cord, or cylinder, of cells (axial skeleton) extending the length of the embryo. In this simple fashion the ground plan of the chordate body is attained.
| |
| | |
| | |
| Fig. 26. - Origin of the mesoderm in Amphioxus (Hatschek). X 425. a/, Lumen of gut; r//., notochord : ai\, ccelom; crrl.p., coelomic pouch; ect., ectoderm; ent., entoderm; n.c., neural canal; n.g., neural groove.
| |
| | |
| | |
| | |
| Fig. 27. - Longitudinal sections of the snake - s blastoderm, at various stages, to show the origin of the notochordal plate (adapted after Hertwig).
| |
| | |
| | |
| | |
| In amphibia, solid mesodermal plates arise in a similar location and extend laterally between the ectoderm and entoderm. Later, these plates split into two layers and the cavity so formed is the coelom (cf .3S)' The notochord also originates as in Amphioxus.
| |
| | |
| | |
| Reptiles, - The same pocket-like depression in the caudal portion of the blastoderm, that gave rise to the cells of the entodermal layer, now invaginates more extensively and forms a pouch which pushes forward between ectoderm and entoderm (Fig. 27 A and B). The size of the invagination cavity varies in different species; in some it is elongate and narrow, lieing confined to the middle line of the blastoderm. The floor of this pouch soon fuses with the underlying entoderm, and the two thin, rupture, and disappear, thus putting the cavity of the pouch temporarily in communication with the space (archenteron) beneath the entoderm (Fig. 27 C). The cells of the roof persist as the notochordal plate, which later liccomes the notochord. The neural folds arise before the mouth of the pouch (blastopore) closes, and, fusing to form the neural tube, incorporate the blastopore into its floor. This temporary communication between the neural tube and the primitive enteric cavity is the neurenteric canal (cf. Fig. 27 C) \ it is found in all the vertebrate groups (cf. Fig. 58). A transverse section through the invaginated pouch, at the time of rupture of its floor, and through the underlying entoderm will make clear the lateral extent of these changes (Fig. 28).
| |
| | |
| | |
| | |
| | |
| | |
| Fig. 28. - Transverse section of a snake - s blastoderm, at a level corresponding to the middle of
| |
| | |
| | |
| Fig. 27 C (adapted after Hertwig).
| |
| | |
| | |
| | |
| | |
| From about the blastopore, and from the walls of the pouch, mesodermal plates arise and extend like wings between the ectoderm and entoderm (Fig. 28). As in amphibia, they later separate into outer (somatic) and inner (splanchnic) layers enclosing the Primitive groove coelom. The relation between notochordal plate, mesoderm, and entoderm, shown in Fig. 28, resembles strikingly the conditions in Amphioxus (Fig. 26 A).
| |
| | |
| | |
| Birds - Due to the modified gastrulation in reptiles, birds, and mammals through the influence of yolk, a structure known as the primitive streak becomes important. An account of its formation and significance, based on conditions found in the bird, may be introduced conveniently at this place.
| |
| | |
| | |
| Shortly after the formation of entoderm, an opaque band appears in the median line at the more caudal portion of the blastoderm (Fig. 29).
| |
| | |
| | |
| | |
| Fig. 29. - BlaStoderm of a chick cmliryo at the stage of the primitive streak. X 20.
| |
| | |
| | |
| Along this primitive streak, which is at first merely a linear ectodermal thickening, there forms a shallow primitive groove, and at its forward end the streak ends in a knob, the primitive knot, or node (of Hensen). The primitive streak becomes highly significant when interpreted in the light of the theory of concrescence, a theory of general application in vertebrate development. It will be remembered that the entoderm of birds arises by a rolling under of the outer layer along the caudal margin of the blastoderm. As the blastoderm expands, it is believed that a middle point on this margin remains fixed (Fig. 30 A) while the edges of the margin on eaeh side are carried caudad and brought together (B, C). Thus, a crescentic margin is transformed into a longitudinal slit. Since this marginal lip originally bounded the blastopore (p. 35), the longitudinal slit must also be an elongated blastopore whose direction has merely been changed. The lips of the slit fuse, forming the primitive streak (D). The teachings of comparative embryology support these conclusions, for the neurenteric canal arises at the cranial end of the primitive streak, the anus at its caudal end, while the primary germ layers fuse in its substance. All these relations exist at the blastopore of the lower animals.
| |
| | |
| | |
| | |
| | |
| Fig. 30. - Diagrams to illustrate the formation of the primitive streak according to the theorjof concrescence. The expanding blastoderm is indicated by dotted circles.
| |
| | |
| | |
| | |
| | |
| Fig. 31. - Median longitudinal section of a chick embryo at the stage of the primitive streak and head process. X 100.
| |
| | |
| | |
| From the thickened ectoderm of the primitive streak a proliferation of cells takes place, and there grows out laterally and caudally between the ectoderm and entoderm a solid plate of mesoderm which soon splits into somatic and splanchnic layers (Fig. 316). An axial growth, the head process, or notochordal plate, likewise extends forward from the primitive knot and fuses at once with the entoderm (Figs. 31 and 317). Since the primitive streak represents a modified blastopore, it is evident that this cranial extension, the head process, corresponds to the pouch-like invagination concerned in the formation of notochord and mesoderm in reptiles. In birds, the fusion of the head process with the entoderm, the relation of mesodermal sheets to it laterally, the formation of the notochord from its tissue and the occasional traces in it of a cavity continuous with the primitive pit (that is, a notochordal canal), all recall the conditions described for the less modified invagination in reptiles. The primitive groove is the visible result of mesoderm proliferation from the tissue of the streak.
| |
| | |
| | |
| Fig. 32. - . 1 , Embryonic disc of the Mateer human embryo, at the stage of the primitive streak (after Streeter). X 50. B, Embryonic disc of the Ingalls human embryo, with primitive streak and head process. X 26.
| |
| | |
| | |
| | |
| Mammals - A typical primitive streak appears on the blastoderm of mammals (Fig. 32 A). The under side of its ectodermal thickening proliferates mesodermal cells which grow laterally and caudally (Fig. 33). All three germ layers fuse in the primitive knot and from it a head process soon extends forward (Fig. 32 B).
| |
| | |
| | |
| | |
| Fig. 33. - Transverse section through the primitive streak of the Mateer human embryo (Fig. 32 ,1 ) to show the origin of mesoderm (redrawn after Streeter). X 185.
| |
| | |
| | |
| The head process of many mammalian embryos contains a cavity {notochordal canal), which in some cases is of considerable size, opening at the primitive pit (Fig. 34). As in reptiles, the floor of this cavity fuses with the entoderm, and the two rupture and disappear. Portions of the floor, still persistent, are shown in Fig. 34. Thus a canal, later enclosed by the neural folds, and then known as the neurenteric canal, puts the dorsal surface of the blastoderm in communication with the enteric cavity beneath the entoderm (Figs. 57 and 58). The roof of the head process, or notochordal plate, is for a time associated closely with the lateral mesoderm (compare these relations in reptiles. Fig. 28), but eventually it becomes the notochord.
| |
| | |
| | |
| | |
| | |
| Fig. 34. - Median sagittal section through the primitive knot and head process of the Ingalls human embryo (Fig. 32 5 ). X 200.
| |
| | |
| | |
| The mesoderm grows rapidly around the wall of the blastodermic vesicle, until Anally the two wings fuse ventrally. The single sheet then splits into two layers, the cavity between being the coelom, or body cavity (Fig. 35). The outer mesodermal layer (somatic), with the ectoderm, forms the somatopleure, or body wall; the inner splanchnic layer, with the entoderm, forms the intestinal wall, or splanchnoplcure. The neural tube having in the meantime arisen from the neural folds of the ectoderm, there is present the ground plan of the vertebrate body, the same in man as in Amphioxus (Fig. 35 A).
| |
| | |
| | |
| | |
| | |
| Fig. 35. - Diagrammatic transverse sections of mammalian embryos. . A , The origin and spread of mesoderm (Brjme); B, the further differentiation of mesoderm, and the body plan (Prentiss).
| |
| | |
| | |
| | |
| | |
| Mesoderm, but not a coelom, is already present in the youngest human embryo yet examined (Fig. 40 A). In Tarsius, a low primate, the mesoderm has two sources: (i) From the splitting of ectoderm at the caudal edge of the blastoderm; this constitutes the extra-embryonic mesoderm and takes no part in forming the body of the embryo. (2) The intra-embryonic mesodertu, which gives rise to body tissues, takes its origin from the primitive streak as in the chick and lower mammals. The origin in the human embryo is probably much the same as in Tarsius.
| |
| | |
| | |
| Homologies of Mesoderm and Notochord. - In Amphioxus and amphibia, transverse sections (Fig. 26) apparently show that the mesoderm and notochord are folded directly from dorsal gut-entoderm. Yet such is illusory, for the roof of the archenteron grows from the dorsal lip of the blastopore. Longitudinal sections prove that as the embryo elongates, this caudal, formative region progressively adds to the roof of the primitive gut. Hence, both notochord and mesoderm originate from the indifferent tissue at the blastopore where ectoderm and entoderm meet. In reptiles, birds, and mammals the mesoderm arises as lateral proliferations from the primitive streak, whereas the notochordal plate (head process) is a - growth - from its anterior end (cf. p. 40). But, as the primitive streak is a modified, fused blastopore (p. 39), their origin is fundamentally like that in Amphioxus and amphibia. From its external position and developmental relations the parent blastoporic tissue is often styled ectoderm; especially in embryos with a primitive streak this is convenient and unobjectionable. It will be evident, therefore, that although the ultimate source of both mesoderm and notochord is from an indifferent - ectoderm, - the notochord, once formed, is true mesoderm.
| |
| | |
| | |
| The Notochord or Chorda Dorsalis. - As the primitive streak recedes caudad during development, the head process is progressively lengthened at its expense. Ultimately, the primitive streak becomes restricted to the tail region and serves as a growth zone there, whereas the entire remainder of the body is built around the head process as an axis. The original position of the primitive knot corresponds to the junction of head and neck in the future body. In later stages, the rod-like notochord extends from head to tail in the midplane (Fig. 91). It becomes enclosed in the centra of the vertebrae and in the base of the cranium, and eventually degenerates. In Amphioxus, the notochord forms the only axial skeleton, and it is persistent in the vertebrae of fishes and amphibians. In adult man, traces are found as - pulpy nuclei - in the intervertebral discs.
| |
| | |
| | |
| Twinning. - LTsually but one human ovum is produced and fertilized at coitus. The simultaneous development of two or more embryos is due commonly to the ripening, expulsion, and subsequent fertilization of an equal number of ova. In such cases ordinary, or fraternal livins, triplets, and so on, of the same or opposite sex result; properly speaking, they are not twins at all. Identical, or duplicate twins, that is, those true twins always of the same sex and strikingly similar in form and feature, arise from two growing points on the embryonic cell mass, each of which develops as a separate embryo within the common chorion. The identical quadruplets of certain armadillos are known to result from the division of a single blastoderm into four parts. Separate development of the cleavage cells can also be produced experimentally in many of the lower animals.
| |
| | |
| Occasionally twins are conjoined. All degrees of union, from almost complete separation to fusion throughout the entire body-length, are known. If there is considerable disparity in size, the smaller is termed the parasite; in such cases the extent of attachment and dependency grades down to included twin (fetus in fetu) and tumor-like fetal inclusions. In some - monsters - the duplication is partial, as doubling of the head or legs. All of these terata, like identical twins, are the products of a single ovum, but variably fused in accordance with their original degree of separation on the blastodermic mass.
| |
| | |
| Stockard reduces the primary cause of all non-hereditary abnormal developments, including twins, to a single factor - developmental inhibition or arrest; the exact type of deformity that results depends solely on the precise moment when the interruption occurs. A slowing of the developmental rate at the critical moment (gastrulation) when one of several potential embryonic axes is about to assert its dominance, causes it to lose its original advantage and one or more neighboring points may then appear as additional axes. The direct cause of the arrest is referred to retarded oxidations.
| |
Arey LB. Developmental Anatomy. (1924) W.B. Saunders Company, Philadelphia.
| Historic Disclaimer - information about historic embryology pages
|
| Pages where the terms "Historic" (textbooks, papers, people, recommendations) appear on this site, and sections within pages where this disclaimer appears, indicate that the content and scientific understanding are specific to the time of publication. This means that while some scientific descriptions are still accurate, the terminology and interpretation of the developmental mechanisms reflect the understanding at the time of original publication and those of the preceding periods, these terms, interpretations and recommendations may not reflect our current scientific understanding. (More? Embryology History | Historic Embryology Papers)
|
Developmental Anatomy - A Text-Book And Laboratory Manual Of Embryology
By
Leslie Brainerd Arey (1891-1988)
Professor Of Anatomy At The Northwestern University Medical School, Chicago.
With 419 Illustrations Many In Color
Philadelphia And London, W. B. Saunders Company
1924
Copyright., 1924, by W. B. Saunders Company, Made in US.A .
Preface
| This book has been prepared for the use of medical students and others whose interests center primarily on man and mammals. The emphasizing of structural rather than functional aspects of Embryology is reflected in the title; such presentation is consistent both with the practical demands of modern courses and with the meagre information existant as to the physiological factors in developmcnt.
The volume contains three sections. In the first part the early stages are treated comparatively and the full course of prenatal and postnatal development is outlined. The second section traces the origin and differentiation of the human organ-systems, grouped according to their germlayer derivations. The third division comprises a laboratory manual for the study of chick and pig embryos.
Many illustrations are from the earlier Prentiss-Arey text and discontinuous fragments of description have likewise been retained. Yet, in plan and content the work is essentially new. It is hoped that the developmental story has been told in an orderly and clear, but concise fashion, and that it records accurately the present state of the subject.
L. B. Arey.
Chicago, ill., September, 1924.
|
 Leslie Brainerd Arey (1891-1988) |
Contents
| PART I. GENERAL DEVELOPMENT
|
PART II. ORGANOGENESIS
Entodermal Derivatives
Mesodermal Derivatives
- Chapter VII. - The Mesenteries and Coelom
- The Mesenteries
- The Primitive Mesentery
- Differentiation of the Dorsal Mesentery
- Differentiation of the Ventral Mesentery
- The Ccelom
- The Primitive Coelom
- The Septum TransveRsum
- The Pleuro-pcricardial and Pleuro-peritoncal Membranes
- The Pericardium and Diaphragm
- Chapter VIII. - The Urogenital System
- The Urinary Organs
- The Pronephros
- The Mesonephros
- The Metanephros
- Differentiation of the Cloaca
- The Genital Organs
- Indifferent Stage
- Internal Sexual Transformations
- The External Genitalia
- Homologies of Internal and External Genitalia
- Chapter IX. - The Vascular System
- Origin of the Blood Vessels and Blood Cells
- Hemopoiesis
- Development of the Heart
- The Primitive Vascular System
- Development of the Arteries
- Development of the Veins
- Fetal Circulation and the Changes at Birth
- The Lymphatic System
|
|
Part III. A LABORATORY MANUAL OF EMBRYOLOGY
- Chapter XVI. - The Study of Chick Embryos
- The Unincubated Ovum and Embryos of the First Day
- Embryo of Five Segments (Twenty-Three Hours)
- Embryo of Seven Segments (Twenty-five Hours)
- Embryo of Seventeen Segments (Thirty-eight Hours)
- Embryo of Twenty-seven Segments (Two Days)
- Embryos of Three to Four Days
- Chapter XVII. - The Study of Pig Embryos
- The Anatomy of a Six Mm. Pig Embryo
- The Anatomy of Ten to Twelve Mm, Pig Embryos
- The Anatomy of an Eighteen Mm. Pig Embryo
- The Anatomy of a Thirty-five Mm. Pig Embryo
- Methods for the Dissection of Pig Embryos
|
Part I. General Development
Introduction
The Scope of Embryology
Developmental anatomy, or embryology, traces the formative history of the individual from the origin of the germ cells to the adult condition. Although the most striking changes in human development occur while the young (called an embryo or fetus) is still inside its mother's womb, yet development by no means ceases at birth. Birth is a mere incident which occurs when the new individual is sufficiently advanced to allow its transference from a protected riterine environment to one in the external world. Some vertebrates, like fishes and amphibia, are capable of an active and independent existence at very immature stages; these free-living larvae, as they are termed, then gradually progress to adults. The human newborn, although far more complete anatomically, is still utterly dependent for food and care: many years of infancy and childhood must elapse before it becomes self-maintaining in human society. During all this period, postnatal development continues. Birth, itself, initiates anatomical changes of profound influence on the body. Throughout the entire growth period, with its uneven but steadily slowing growth rate, come the completion of some organs and a gradual remoulding of the shape of the body and its parts. Only at the age of twenty-five are these progressive changes complete.
All vertebrate, or backboned, animals are organized upon a common anatomical plan, and even many of their structural details are comparable, though superficially disguised. Similarly, their fundamental mode of development is essentially identical. The minor variations that do occur are caused by such secondary modifying factors as the crowding yolk content of the egg or adaptations to development inside or outside the mother's body. While the comparative viewpoint is indispensable for gaining a broad understanding of embryology, it has been of especial importance in supplying missing parts of the human developmental story and in interpreting many perplexing conditions. For, the earliest human embryos known are about two weeks old and have the three primary germ layers already formed. Even invertebrate material is highly useful for demonstrating such early stages as maturation, fertilization, cleavage, and the formation of blastula and gastrula.
The Value of Embryology
A general conception of how man and other animals develop from a single cell by orderly and logical processes should share in the cultural background of every educated mind. To the medical student, embryology is of primary importance because it affords a comprehensive understanding of the intricacies and variations of human anatomy, and thus is essential to sound surgical training. It also explains many anomalies and 'monstrous - conditions, and the origin of certain tumors and other pathological changes in the tissues. Obstetrics is essentially applied embryology. From the theoretical side, it is the key with which we may unlock the secrets of heredity, the determination of sex, and, in part, of organic evolution.
Historical
Fig. 1. - Human sperm cell containing a miniature organism, according to Hartsoeker (1694).
The science of modern embryology is comparatively new, originating with the use of the compound microscope and advancing with the improvement of microscopical technique. Aristotle (384-322 B. c.), however, centuries before the introduction of magnifying lenses had followed the general development of the chick, day by day. The popular belief that slime and decaying matter is capable of giving rise to living animals, as also asserted by Aristotle, was disproved by Redi (1668).
A few years after Harvey and Malpighi had published their fundamental studies on the chick embryo, Leeuwenhoek reported the discovery of the human spermatozoon by Ham in 1677 - At this period, it was believed either that fully formed animals existed in miniature in the egg, needing only the stimulus of the spermatozoon to initiate development, or that similarly preformed bodies, male and female, constituted the spermatozoa and that these merely enlarged within the ovum. According to this doctrine of preformation, all future generations were likewise encased, one inside the sex cells of the other, and serious computations were made as to the probable number of progeny (200 millon) thus present in the ovary of Mother Eve, at the exhaustion of which the human race would end! Dalenpatius (1699) and others even believed they had observed a minute human form in the spermatozoon (Fig. 1).
The preformation theory was strongly combated by Wolff (1759), who saw that the organs of the early chick embryo were differentiated gradually from unspecialized living substance. This theory, known as epigenesis, was proved correct when von Baer discovered the mammalian ovum in 1827, and later demonstrated the germ-layer composition of all embryos.
About twenty years after Schleiden and Schwann (1839) had shown the cell to be the structural unit of the organism, the ovum and spermatozoon were recognized as true cells. O. Hertwig, in was the first to observe and appreciate the events of fertilization. Henceforth, all multicellular organisms were believed to develop each from a single fertilized ovum. This conception is expressed in the famous aphorism: ornne vivum ex ovo .
Modern Embryology
As an organized and definite science, began with Balfour (1874), who reviewed, digested, and made accessible the earlier scattered facts. Throughout this period, the experimental method of investigation has been used increasingly; without it many structural and physiological aspects of development would remain unsolved.
General Features Of Development
A multicellular embryo results from the division of the fertilized ovum to form daughter cells. These are at first quite similar in structure, and, if separated, in some animals each may become a complete embryo (sea urchin; certain vertebrates). In general, the development of an embryo depends: (i) upon the multiplication of its cells by division; (2) upon the growth in size of the individual cells; (3) upon changes in their form and structure.
Cell Division - All cells arise from pre-existing cells by division. There are two methods of cell division - amitosis and mitosis.
Amitosis - Cells may divide directly by the simple fission of their nuclei and cytoplasm. This rather infrequent process is called amitosis. Amitosis is said by many to occur only in specialized or moribund cells. It is the type of cell division demonstrable in the epithelium of the bladder.

Fig. 2. Diagrams of the phases of mitosis (Schafer).
Mitosis
In the reproduction of typically active somatic cells and in all germ cells, complicated changes take place in the nucleus. These changes give rise to thread-like structures, hence the process is termed mitosis (thread) in distinction to amitosis (no thread). Mitosis is divided for convenience into four phases (Fig. 2) :
Prophase
- The centrosome divides and the two minute bodies resulting from the division move apart, ultimately occupying positions at opposite poles of the nucleus (I-III).
- Astral rays appear in the cytoplasm about each centriole. They radiate from it, and the threads of the central or achromatic spindle are formed between the two asters, thus constituting the amphiaster (II).
- The nuclear membrane and nucleolus disappear, the karyoplasm and cytoplasm becoming confluent.
- During the above changes the chromatic network of the resting nucleus resolves itself into a skein, or spireme, which soon shortens and breaks up into distinct, heavily-staining bodies, the chromosomes (II, III). The definite number of chromosomes is always found in the cells of a given species, the chromosomes may be block-shaped, rod- shaped, or bent in the form of a II or V.
- The chromosomes arrange themselves in the equatorial plane of the central spindle (IV). If U- or V-shaped, the angle of each is directed toward a common center. The amphiaster and the chromosomes together constitute a mitotic figure, and at the end of the prophase this is called a monaster.
Metaphase - The longitudinal splitting of the chromosomes into exactly similar halves constitutes the metaphasc (IV). The aim of mitosis is thus accomplished, an accurate division of the chromatin between the nuclei of the daughter cells.
Anaphase - The two groups of daughter chromosomes separate and move up along the central spindle fibers, each toward one of the two asters. Hence this is called the diaster stage (V, VI). Each centriole may divide in preparation for the next diviSion of the daughter cells.
Telophase - i. The daughter chromosomes resolve themselves into a reticulum and daughter nuclei are formed (Vu, VuI).
2. The cytoplasm divides in a plane perpendicular to the axis of the mitotic spindle (VIII). Two complete daughter cells have thus arisen from the mother cell.
The number of chromosomes is constant in the cells of a given species. The smallest assortment, two, occurs in Ascaris megalocephala univaleus, a round worm parasitic in the intestine of the horse. The largest number known is found in the brine shrimp, Artemia, where 168 have been counted. The chromosome enumeration for the human cell has been variously stated but the results of Winiwarter (1912), Grosser (1921), and Painter (1923) now agree on a relatively high number, which Painter establishes as 48 for whites and negroes of both sexes.
The Germ Layers
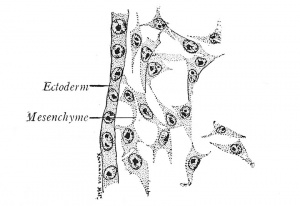
Fig. 3. Mesenchyme from a chick embryo (Prentiss). X 495.
The first changes in the form and arrangement of the cells establish three definite plates, the primary germ layers, which are termed from their positions the ectoderm (outer skin), mesoderm (middle skin) and entoderm (inner skin) (Fig. 4). Since the ectoderm covers the body, it is primarily protective in function, but it also gives origin to the nervous system, through which sensations are received from the outer world. The entoderm, on the other hand, lines the digestive canal and is from the first nutritive. The mesoderm, lying between the other two layers, naturally performs the functions of circulation, of muscular movement, and of excretion; it also gives rise to the skeletal structures which support the body. While all three germ layers form definite sheets of cells known as epithelia, the mesoderm takes also the form of a diffuse meshwork of cells, the mesenchyme (Fig. 3).
The cells of these layers are modified in turn to form tissues, such as muscle and nerve, of which the various organs are composed. The organs, associated as organ systenis, constitute the organism, or body, that of adult man containing 2 5 million million red blood cells alone. In every organ, one tissue, like the epithelial lining of the stomach, is predominately important; the others are accessory.
Histogenesis. -The cells of the germ layers are at first alike in structure. Thus, the evagination which forms the primordial arm is composed of a single layer of similar ectodermal cells, surrounding a central mass of diffuse mesenchyme (Fig. 406). Gradually the ectodermal cells multiply, change their form and structure, and give rise to the layers of the epidermis. By more profound structural changes the mesenchymal cells ahso are transformed into the elements of connective tissue, tendon, cartilage, bone, and muscle - aggregations of modified cells which are termed tissues. The development of modified tissue cells from the undifferentiated cells of the germ layers is known as histogenesis.
During histogenesis, the structure and form of each tissue cell are adapted to the ])erformance of some special function or functions. Cells which have once taken on the structure and functions of a given tissue cannot give rise to cells of any other type. In tissues like the epidermis, certain cells retain their ])rimitive embryonic characters throughout life, and, by continued cell division produce new layers of cells which are later specialized. In other tissues all of the cells are differentiated into the adult type, after which no new cells are formed: this takes place in the nervous elements of the central nervous system. Contrariwise, most tissue cells are undergoing retrogressive changes throughout life. In this way, the cells of certain organs like the thymus gland and mesonephros degenerate and largely disappear. The cells of the hairs and the surface layer of the epidermis become cornified and eventually are shed. Thus, normally, many tissue cells are continually being destroyed and replaced by new cells.
This series of changes - an embryonic (undifferentiated) stage; progressive functional s])ecialization ; gradual degeneration; death and removal - which tissue cells experience is designated by the term cytomorphosis.
Derivatives of the Germ Layers
The tissues of the adult are derived from the primary germ layers as follows:
| Ectoderm
|
Mesoderm
|
Entoderm
|
| I. Epidermis and derivatives.
Hair; nails; glands.
Lens of eye.
2. Epithelium of:
Organs of special sense. Cornea.
Mouth; enamel organ.
Oral glands; hypophysis.
Anus.
Amnion; chorion.
3. Nervous tissue.
Neuroglia.
Chromaffin tissue.
4. Smooth muscle of; Iris.
Sweat glands.
|
A. Mesothelium.
1. Pericardium.
2. Pleura.
3. Peritoneum.
4. Urogenital epithelia.
5. Striated muscle.
B. Mesenchyme.
1 . Smooth muscle.
2. Notochord.
3. Connective tissue; cartilage; bone.
4. Blood; bone marrow.
5. Endothelium of blood vessels and lymphatics.
6. Lymphoid organs.
7. Suprarenal cortex.
|
Epithelium of:
I. Pharynx and derivatives. Auditory tube.
Tonsils.
Thymus.
Thyroid.
Parathyroid.
2. Respiratory tract.
Larynx; trachea.
Lungs.
3. Digestive tract.
Liver; pancreas.
Yolk sac; allantois.
4. Bladder (except trigone).
5. Urethra (except prostatic).
6. Prostate.
|
Primitive Segments - Metamerism
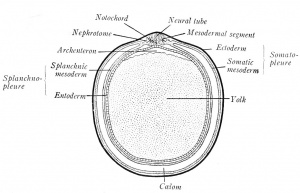
Fig. 4. Diagrammatic transverse section of a vertebrate embryo (Minot-Prentiss).
A prominent feature of vertebrate embryos are the primitive segments, or metameres (Fig. 59). These segments are homologous to the serial divisions of an adult earth-worm's body, divisions which, in the earth worm, are identical in structure, each containing a ganglion of the nerve cord, a muscle segment, or myotome, and pairs of blood vessels and nerves. In vertebrate embryos, the block like primitive segments lie next the neural tube and are known as mesodermal segments, or somites (Fig. 4). Each pair gives rise to a vertebra, to two myotomes, or muscle segments, and to paired vessels; each set of mesodermal segments is supplied by a pair of spinal nerves: consequently, the adult vertebrate body is segmented like that of the earth worm. As a worm grows by the formation of new segments at its tail-end, so the metameres of the vertebrate embryo begin to form in the head and are added tailward. There is this difference between the segments of the worm and the vertebrate embryo; the segmentation of the worm is complete, while that of the vertebrate is incomplete ventrally.
Somatopleure and Splanchnopleure
In early embryos the mesoderm splits into two layers, the somatic (dorsal) and splanchnic (ventral) mesoderm (Fig. 4). The ectoderm and somatic mesoderm constitute the body wall, which is termed the somatopleure. In the same way, the entoderm and splanchnic mesoderm combine as the splanchnopleure; it forms the mesenteries and the walls of the gut, heart, and lungs.
Coelom
The space between the somatopleure and splanchnopleure is the coelom, or body cavity. At the first splitting of the mesoderm, isolated clefts are produced. These unite on each side and eventually form one cavity - the coelom. With the extension of the mesoderm, the coelom surrounds the heart and gut ventrally (Fig. 4). Later, it is subdivided into the pericardial cavity about the heart, the pleural cavity of the thorax, and the peritoneal cavity of the abdominal region. The epithelia lining the several body cavities are termed mesothelia.
The Nephrotome
The bridge of cells connecting the primitive segment with the unsegmented somatic and splanchnic layers is the nephrotome, or intermediate cell mass (Fig. 4). From these will develop the urogenital glands and ducts.
Developmental Processes
The developing embryo exhibits a progressively complex structure, the various steps in the production of which occur in orderly sequence. There may be recognized in development a number of component mechanical processes which are used repeatedly by the embryo. The general and fundamental process conditioning ilifferentiation is cell multiplication, and the subsequent growth of the daughter cells. The more important of the specific developmental processes are the following: ( 1) cell migration; (2) localized growth, resulting in eidargements and constrictions; (3) cell aggregation, forming (a) cords, (b) sheets, [c] masses; (4) delamination, that is, the splitting of single sheets into separate layers; (5) folds, including circumscribed folds which produce (a) evaginations, or out-pocketings, (b) invaginations, or in-pocketings.
The production of folds, including evaginations and invaginations, due to unequal rapidity of growth, is the chief factor in moulding the organs and hence the general form of the embryo.
Fundamental Conceptions
The Anlage
This German word, which lacks an entirely satisfactory English equivalent, is a term applied to the first discernible cell, or aggregation of cells, which is destined to form any distinct jiart or organ of the embryo. In the broad sense, the fertilized ovum is the anlage of the entire adult organism; furthermore, in the early cleavage stages of certain embryos it is possible to recognize single cells or cell groups from which definite structures will indubitably arise. The term anlage, however, is more commonly applied to the primordia that differentiate from the various germ layers. Thus the epithelial thickening over the optic vesicle is the anlage of the lens.
The Law of Genetic Restriction
As development advances, there is a constantly increasing restriction in the kind of differentiation open to the various parts. Each emerging tissue or organ is more rigidly bound to its particular type of differentiation than was the generalized material from which it came. A line of specialization, once begun, cannot be abandoned for another type. The parent tissue, likewise, is limited by losing the capacity for duplicating anlages already formed. Thus, the primitive thyroid can never become anything but a thyroid, whereas the gut that formed it also buds off, at other levels, the lungs, liver, and pancreas. Yet if the embryonic thyroid were destroyed, the pharynx would never replace it. From mesenchyme arise connective tissue, blood cells, and smooth muscle; when once the specialization begins, there can be no retraction or transformation to another type.
Continuity of the Germ Plasm
According to this important conception of Weismann, the body-protoplasm, or soma, and the reproductive-protoplasm differ fundamentally. The germinal material is a legacy that has existed since the beginning of life, from which representative portions are passed on intact from one generation to the next. Around this germ plasm there develops in each successive generation a shortlived body, or soma, which serves as a vehicle for insuring its transmission and perpetuation. The reason, therefore, why offspring resembles parent is because each develops from portions of the same stuff.
The Law of Biogenesis
Of great theoretical interest is the fact, constantly observed in studying, embryos, that the individual in its development repeats hastily and incompletely the evolutionary history of its own species. This law of recapitulation was first stated clearly by Muller in 1863, and was termed by Haeckel the law of biogenesis. In accordance with it, the fertilized ovum is compared to a unicellular organism like the Ameba: the blastula is supposed to represent an adult Volvox type; the gastrula, a simple sponge; the segmented embryo, a worm-like stage ; and the embryo with gill slits may be regarded as a fishlike stage. Moreover, the blood of the human embryo in development passes through stages in which its corpuscles resemble in structure those of the fish and reptile; the heart is at first tubular, like that of the fish, and the arrangement of blood vessels is equally primitive; the kidney of the embryo is like that of the amphibian, as are also the genital ducts. Many other examples of this law may readily be observed.
Some apparently useless structures appear during development, perfunctorily reminiscent of ancestral conditions; certain other parts, of use to the embryo alone, are later replaced by better-adapted, permanent organs. Representatives of either type may eventually disappear or they may persist throughout life as rudimentary organs; more than a hundred of the latter have been listed for man. Still other ancestral organs abandon their provisional embryonic function, yet are retained in the adult and utilized for new purposes.
The Vertebrate Groups
There are five vertebrate classes, the higher characterized by the possession of an enveloping embryonic membrane, called the amnion, and another embryonic appendage, known as the allantois:
(A) Anamniota (amnion absent).
- 1. Fishes - lamprey; sturgeon; shark; bony fishes; lung fish.
- 2. Amphibia - salamander; frog; toad; etc.
(B) Amniota (amnion present).
- 3. Reptiles - lizard; crocodile; snake; turtle.
- 4. Birds.
- 5. Mammals. Characterized by hair and mammary glands.
- (a) Monotremes - duck-bill; primitive mammals that have a cloaca and lay eggs with shells.
- (b) Marsupials - oppossum; kangaroo; etc. The young are born immature and are sheltered in an integumentary pouch.
- (c) Placentalia. All other mammals whose young are nourished in the uterus by a placenta.
- Ungulate series. Hoofed mammals (cattle; sheep; pig; deer; horse; etc.).
- Unguiculate series. Clawed mammals (mole; bat; rat; rabbit; cat; dog; etc.). The highest order is the Primates (lemur; monkey; ape, man).
The Vertebrate Body Plan
All vertebrate animals are constructed in accordance with a common body plan. The distinctive characteristics of the vertebrate type include: .
- A tubular central nervous system, dorsally placed (Fig. 4).
- A notochord, between the neural tube and gut (Fig. 4). This cellular |3rimitive-axis is replaced, wholly or in part, by the vertebral column.
- A pharynx, which develops paired pouches and clefts that determine the positions of important nerves, muscles and blood vessels (Fig. 91).
- The position of the mouth. Unlike the condition in many invertebrates, it is not surrounded by a circumoral ring of nervous tissue which connects a dorsal - brain - with a ventral chain of ganglia.
- The limbs, Two pairs, with an internal skeleton (Fig. 227).
- A coelom, which is divided into a dorsal, segmental part (cavities of the somites), and a ventral, unsegmented part, partitioned by the septum transversum (diaphragm) into thoracic and abdominal portions (Fig. 4).
Titles for Collateral Reading and Reference
Broman. Normale und abnorme Entwicklung des Menschen.
Corning. Entwicklungsgeschichte des Menschen.
Duval. Atlas D - Embryologie.
Hertwig. Handbuch der Entwicklungslehre der Wirbeltiere.
Keibel and Mall. Human Embryology.
Kellicott. A Textbook of General Embryology.
Kollmann. Handatlas der Entwicklungsgeschichte des Menschen.
Lillie. The Development of the Chick.
Minot. A Laboratory Text-book of Embryology.
McMurrich. The Development of the Human Body.
Patten. The Early Embryology of the Chick.
Wilson. The Cell in Development and Inheritance.
| Historic Disclaimer - information about historic embryology pages
|
| Pages where the terms "Historic" (textbooks, papers, people, recommendations) appear on this site, and sections within pages where this disclaimer appears, indicate that the content and scientific understanding are specific to the time of publication. This means that while some scientific descriptions are still accurate, the terminology and interpretation of the developmental mechanisms reflect the understanding at the time of original publication and those of the preceding periods, these terms, interpretations and recommendations may not reflect our current scientific understanding. (More? Embryology History | Historic Embryology Papers)
|
Reference
Arey LB. Developmental Anatomy. (1924) W.B. Saunders Company, Philadelphia.
Cite this page: Hill, M.A. (2024, May 18) Embryology Book - Developmental Anatomy 1924. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Book_-_Developmental_Anatomy_1924
- What Links Here?
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G








{kind=link}
{kind=link}
{kind=link}