Book - Contributions to Embryology Carnegie Institution No.56-13
Chapter 13. Post-Mortem Intrauterine Changes
| Embryology - 26 Feb 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Mall FP. and Meyer AW. Studies on abortuses: a survey of pathologic ova in the Carnegie Embryological Collection. (1921) Contrib. Embryol., Carnegie Inst. Wash. Publ. 275, 12: 1-364.
- In this historic 1921 pathology paper, figures and plates of abnormal embryos are not suitable for young students.
1921 Carnegie Collection - Abnormal: Preface | 1 Collection origin | 2 Care and utilization | 3 Classification | 4 Pathologic analysis | 5 Size | 6 Sex incidence | 7 Localized anomalies | 8 Hydatiform uterine | 9 Hydatiform tubal | Chapter 10 Alleged superfetation | 11 Ovarian Pregnancy | 12 Lysis and resorption | 13 Postmortem intrauterine | 14 Hofbauer cells | 15 Villi | 16 Villous nodules | 17 Syphilitic changes | 18 Aspects | Bibliography | Figures | Contribution No.56 | Contributions Series | Embryology History
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
In discussing post-mortem changes, it is necessary to distinguish between the death of and changes in the cyema and the vesicles, that is, between inti acyemic and extracyemic changes. Such a distinction would be unnecessary if the death of the one were impossible without that of the other. It has been assumed in the past not only that the latter is not the case, but that the fetal vesicles may survive the cyema for some time. Under such circumstances, then, considerable post-mortem changes might take place within the cyema and perhaps within the amnion also, while the chorion might remain largely, even if not wholly, unaffected. Since such an assumption involves the implication that the fetal circulation is not indispensable for the life of the chorionic vesicle even after the usual time of the advent of the former, an assumption of independent survival of the entire vesicle would seem somewhat venturesome, although it is highly probable that certain constituents of the chorion nevertheless might survive death of the cyema.
That the amnion, which usually is non-vascular, and which fuses with the chorion quite early in development, may survive death of the embryo is entirely possible, not only because it is nutritionally more independent, especially in its early stages, but also because survival of the chorion would condition survival of the amnion after the latter fuses with the former. The belief, not only in the survival, but also in the growth of the fetal vesicles after death of the cyema, is relatively old and rests largely upon the disproportion in size between the embryo and the chorionic vesicles so frequently seen, and upon the existence of a similar disproportion between the placenta and the fetus in cases of retained abortions. According to Panum, von Baer, describing a rather firm, brownish-red, beanshaped lump, unlike an embryo of either bird or mammal, which was surrounded by an envelope and in relation to a vascular area, was said to have regarded it as a liver which had continued to grow after the death of the embryo. It seems that von Baer came to this conclusion, largely because of the color of the gross specimen and because he thought it possible that in poorly regulated incubators the "vegetative" portions of the body of the embryo very often continue to live, although the "animal" parts die. It must be added, however, that Panum (1860), from whose monograph this statement is taken, added that he thought von Baer only regarded these questions as propositions for investigation and proof, and not as established facts. However, Giacomini (1893) adduced experimental evidence in favor of such a belief, for he stated that the membranes also continued to grow in rabbits in which the cyema had been killed experimentally. He further stated that, although the embryo may be inhibited in growth or even be destroyed completely, the chorion and amnion not only may show no degenerative change, but actually may continue to grow. His (1891) spoke of evidence of an interrupted growth, and Giacomini (1894) also believed in the occurrence of an inhibition of growth before the death of the embryo occurred. According to Waldstein (1913), von Winckel also spoke of a slowing in development before the death of the embryo occurred. Taussig (1903) stated that both Breus and Gottschalk concluded that retained "ova" grow, and Graefe (1896) called attention to the opinion of Veit that "ova" may continue to grow after the death of the embryo, but added that the existence of bare areas and the bunching of villi in some of the retained specimens argue against the conception of His that "ova" with these characteristics have continued to grow for some time.
No one seems to have suggested that the entire embryo may continue to grow after cessation of the fetal circulation, but Wallenstein (1897), who made a careful microscopic examination of some early abortuses, not only believed that cellular proliferation can occur in the embryo after its death, but that either the lateral or the dorsal or ventral halves may continue to grow independently of each other after the death of the cyema. Wallenstein believed, even, that the cyema can become diseased after its death, and concluded that intravascular cells not only outlive the rest of the cyema, but that they proliferate and invade the dead or dying tissues of the latter.
Schaeffer (1898) also stated that the chorionic and amniotic vesicles continue to grow, provided death of the embryo occurs before the fourth month, and Mall (1900, 1903) also spoke in favor of growth in them after death of the embryo. Engel (1900) believed that the caudal half of the body of an embryo may continue to grow after the cephalic half has died. However, Engel's conclusion regarding the proliferation of cells was based largely upon the observation that the volume of the disintegrating central nervous system was too great to be accounted for merely by the cells which normally compose it.

Giacomini (1894) believed that even the isolated amnion may continue to grow and that the chorion may continue to live after all else has died. That the individual portions of very young conceptuses undoubtedly have considerable power of independent, though not necessarily post-mortem, growth is shown by such specimens as No. 1843 (fig. 7, plate 1, Chap. IV), a conceptus in which the chorion and yolk-sac have reached a considerable size, although the embryo and amnion both are absent, except perhaps in the merest rudiments, as shown in figure 172 (plate 16, Chap. XI). However, it is highly probable that this power of independent growth is far greater before than after the time when the fetal circulation has become established in the cyema and chorion. Soon after this time cessation of the embryonic circulation necessarily would seem to interfere effectively with growth of the cyema and increasingly with the nutrition of the chorion and more indirectly also with that of the amnion. However, before the fetal circulation has been established, it is highly probable that even the embryonic disk may be much more resistant to inhospitable surroundings, and one could pronounce these very young conceptuses dead only after correlated cellular proliferation had ceased entirely.
Although the chorions of older specimens may die a more gradual death than the cyema, one scarcely would expect any villi to be formed anew after the death of the latter. The syncytium and trophoblast, however, both might continue to grow for some time and apparently often do so, for, as is well illustrated by many things, they are much more independent, not only of the fetal circulation, but also of the conceptus, than the other components. Strahl and Henneberg (1902) also found the ectodermic elements of the placenta in guinea-pigs more resistant than the mesodermic. Some investigators (His, for example) expressed the opinion that, although survival of tissues, within both the cyema and the vesicles, is possible after death of the embryo, one really can not regard such tissues as truly living, although cell proliferation nevertheless may be present. Giacomini and His both spoke of cell proliferation, and Mall later used the term dissociation as including both migration and proliferation of cells. It may be recalled in this connection that Miiller (1847) also spoke of proliferation of the epithelium of the villi as being common in aborted ova, and that Giacomini (1888) stated that the cells of the central nervous system tend to become uniform and also seem to multiply because they no longer can be accommodated in the space provided for them. His, Giacomini, Grawitz, Mall, and Engel all believed that at least a proliferation of "round cells" occurs and that they are wandering or migratory in nature. Marchand (1895) also spoke of an infiltration of the conceptus by leucocytes, but did not believe, as did Grawitz, that they were autochthonous in origin. Although Daels (1908 b ) stated that maternal leucocytes can penetrate normal syncytium in order to reach a necrotic area beyond, Nattan-Larrier and Brindeau (1905 a , 1908) concluded that leucocytes never invade the stroma of the villi as long as the epithelium is intact. The conclusion of the latter investigators is wholly in accord with my own observations, and it would seem extremely difficult to determine whether leucocytes seen somewhere in the stroma of the villi or of the chorionic membrane really are fetal or maternal in origin. According to Windle (1893), the so-called round cells arise in the stroma of the villi, but Wallenstein believed that such an infiltration can occur only into dead or dying embryonic tissue. Berlin (1907) also spoke of the presence of extensive leucocytic infiltration in one of her cases, and Engel (1900) concluded that in one of the cases described by him the liver was completely destroyed by proliferating round cells which he apparently assumed to be phagocytic.
Microscopic evidence for the survival and growth of the cyema or certain parts thereof does not rest solely upon the transformation of various tissues and organs into "round cells," however. Giacomini and Wallenstein both stated that mitotic figures were observed by Chiarugi, but the latter seems to stand alone in regard to this observation. But, even if confirmed, the occurrence of mitoses locally would not establish the occurrence of a correlated proliferation which could rightly be designated as growth. Nor could the well-known independent survival or even growth of still-implanted villi or, more properly speaking, of the syncytium and stroma, in cases of hydatiform degeneration and chorio-epitheliomata, be regarded as establishing the occurrence of normal growth in the vesicles or the cyemata of conceptuses.
Post-mortem obliteration of the vessels would seem to fall more in this category, but the post-mortem occurrence of this phenomenon even is as yet unproved.

Besides, upon a priori grounds alone it would seem that the adverse conditions which cause the death of the embryo, and eventually also that of the entire conceptus, alone would make conditions of post-mortem growth rather unfavorable even for the most resistant of the dependent tissues. Other factors also would tend to make these conditions unfavorable for the occurrence of post-mortem proliferation, even for such growth as we have in recent years come to know as growth in vitro. I have rarely found appearances identical with those seen in certain artificial cultures, as figure 175 so well illustrates; but these appearances involved connective tissue only and always occurred in the interior of the chorionic membrane, or between the latter and the amnion when the two were separated somewhat. Nevertheless, the rareness of these appearances in itself throws much doubt on the occurrence of genuine growth after death, and in view of the fact that I have been unable to find any thickening of the chorionic and amniotic membranes at these places, I am more inclined to regard such appearances as these as due to a migration rather than to an actual proliferation of cells. Nor have I seen much evidence in favor of a post-mortem migration other than that of cells intravascular in origin. What has been termed lymphoid infiltration or transformation seems to be due rather to a degeneration of cells in loco, the degenerate forms simulating round cells and thus giving rise to misinterpretations. Indeed, the idea that a complete transformation of the entire body of the embryo into round cells can occur without the phagocytosis of the cells composing the various organs seems to imply that this so-called lymphoid infiltration really is a transformation in loco. That an apparent temporary increase in volume of the central nervous system has been noted is not surprising. But this does not postulate an actual increase in the number of cells any more than the old observation of maintenance in weight and volume of the central nervous system under conditions of inanition implied that the nerve-cells remained unchanged. That the latter is not the case was shown by Meyer (1917 h ).
It may be recalled that Merttens (1894) also reported finding obliteration of the blood-vessels due to proliferation of the intima. However, since Merttens found these obliterative vascular changes only in the chorion lave, his conclusion that these changes undoubtedly were post-mortem is open to very serious doubt. This doubt is greatly increased by the facts that Merttens suspected the existence of lues in a case in which these obliterative changes were present and that almost complete obliteration of the vessels was present in a specimen which was regarded as having been retained only 1 1 days. Since calcium deposits were present in this specimen, the latter estimate is open to grave doubt, however.
Berlin (1907), although aware of the fact that an ante-mortem obliterative endarteritis had been shown to occur in conceptuses, nevertheless concluded that the degree of obliteration found in the placental vessels roughly parallels the duration of the retention. She added that the processes found by her recall those observed in portions of a blood-vessel isolated by ligature and represented appearances within the placental vessels almost identical with those represented in figure 175. In spite of the similarity of these figures, it would seem unwarranted, however, to assume that phenomena identical with those observed in the chorionic vesicles could lead to obliteration of vessels. In the chorionic vesicle the picture presented is that of migration rather than that of proliferation, and it is indeed a long step from these things to complete obliteration of blood-vessels, especially when it is recalled that the latter involves the growth of new capillaries, as was reported by Merttens and as is the case in the normal postnatal obliteration of the umbilical vessels. Schickele (1907) also observed obliteration by endothelial proliferation in the various vessels, but insisted that they do not occur with the frequency reported by Merttens. Schickele further stated that obliteration occurs rapidly and that it is not necessarily post-mortem.
Waldstein, who made a very careful histologic examination of some abortuses, stated that, although the regressive changes had not yet been sufficiently studied, he did not believe in a continued growth of the embryo, but merely in a further differentiation of some of the constituent tissues, and also in a round-cell infiltration. Waldstein claimed that in some specimens examined by him the striations in the central musculature were maturer than they should have been, and hence concluded that post-mortem differentiation had taken place in them. He believed that the round cells had an intravascular origin. Boerma (1912), who described a young macerated embryo, also stated that the amniotic cavity contained cellmasses from the embryo which he believed had survived the parent mass.
His (1891), Giacomini (1894), Schaeffer (1898), Mall, and the writer occasionally found local thickenings of and adhesions between certain epidermal surfaces. These adhesions and thickenings, referred to in Chapter IV, group 7, were present in the lower extremities of the fetus shown in figures 64 and 65 (plate 5, Chap. IV). In this case, as in those of Lomer and Schaeffer, the extremities are glued together by epidermis. The appearance of the lower extremities at this point also suggests that there has been an accumulation of sloughed epidermis in the region of contact. To what extent this gluing is due to fusion of dying tissues and cells, rather than to growth, it is difficult to say. In other cases, as stated by His (1891), the extremities may be glued to the trunk, or the head to the chest, as observed also by Mall and as illustrated in figure 176. The mandibular region may adhere to the chest and the lips coalesce so completely that the epidermis is absolutely continuous from the chest to the chin and also across the lips, as represented in section in figure 180.
Giacomini (1889) further described invaginations of the epidermis into the mesoderm on the dorsum, lateral to the spine in an atrophic embryo 5 mm. long. Although not regularly symmetrical, these epidermal invaginations nevertheless were said to have a metameric arrangement, there being 6 on the right and 10 on the left side. Giacomini stated that these epidermal growths reminded him of the lateral-line organs of elasmobranchs, and added that he saw them at the caudal extremity of another specimen. Epidermal accumulations in various places were noticed also by Mall in long-retained cyemata, but nothing justifying Giacomini's interpretation was encountered either by Mall or myself. The accumulations noticed by us were small, irregular, and rather poorly preserved mounds of epidermis, as illustrated in figures 76 and 77.
That the placenta may continue to grow during retention after death of the cyema was once firmly believed. This idea had its origin in the disproportion frequently present between the size of the placenta and that of the cyema. At present, belief in the growth of the placenta after death of the cyema seems to have been abandoned, however. It was in connection with the placental changes, and especially with changes in the vessels of the villi, that the question of the survival and growth of cyemic tissues attracted attention. Among the placental changes which suggested the presence of growth was the proliferation of the ectoderm of the villi, indicated in so many specimens. It is interesting that Schickele (1907) regarded this apparent post-mortem proliferation as a direct cause of cyemic death. Schickele believed that proliferation of the ectoderm of the villi results in encroachment upon the placental circulation and ultimately in death of the cyema. It seems strange, however, that Schickele (1905) stated that proliferation of the syncytium usually does not occur, except in cases of long-standing retention. . Litthauer (1887) also had believed in the continued growth of the placenta as a result of retention, and had concluded that the death of the cyema was attributable to interference with its nutritive supply through proliferation of the endometrium.

Taussig (1903) stated that the vessels in the villi of retained "ova" may be preserved for a long time, as reported also by Davidson, and Berlin believed that obliteration of the villous vessels is a purely post-mortem phenomenon. As previously stated, Berlin affirmed the belief of Merttens (1894) that the degree of vascular obliteration somewhat parallels the duration of the retention. If this obliteration actually occurs post-mortem, this conclusion of Merttens may be correct, for it is well known that the degree of obliteration of the umbilical vessels is dependent upon the duration since birth. However, the occurrence of antemortem vascular changes established by numerous investigators can not be excluded. Nor is it necessarily an easy matter to differentiate the ante-mortem from the post-mortem proliferative vascular changes. If the villi can continue to grow after death of the fetus the rate of obliteration will be very much affected, no doubt, by this fact. It is interesting that Graefe, although granting the possibility of independent growth of the villi in retained uterine conceptuses, stated that he saw hypertrophy of the villi only once. Schaeffer also believed that the villi are preserved for a long time after death of the cyema, and Mall (1915) stated that in cases of tubal pregnancy the villi may continue to grow independently after death of the embryo.

Crosti (1896) stated that if the ovum is not aborted immediately after death of the embryo the villous capillaries gradually disappear. Crosti also believed that the villi are preserved longer than the rest of the conceptus, and that even the "appendici durate" may become hydropic. It must be borne in mind, however, that the disappearance of the villous capillaries in consequence of maceration is one thing, and the obliteration of the vascular lumen by proliferative processes is quite another. Obliteration such as that represented in figure 177 illustrates the former process, which is a purely post-mortem phenomenon as far as the particular villus is concerned, while that represented by No. 317, shown in figure 178, illustrates proliferative obliteration of the vascular lumen which undoubtedly was ante-mortem. However, it must be recalled that not all of a chorionic vesicle necessarily dies at the same time, and that implanted villi, or those in very young conceptuses which are not yet dependent upon the fetal circulation, may well continue to live and perhaps even continue to grow for some time. But such villi hardly can justly be designated as dead, and since death of the cyema does not necessarily result in simultaneous death of the entire conceptus, this distinction is not an unimportant one.
There seems to be no agreement among writers, however, as to which of the tissues of the cyema are most resistant to post-mortem changes. Litthauer (1887) found the muscles of certain fetuses well preserved, although the cartilages had lost all their normal structure. Virchow, Wyder, and Baumgarten, according to Litthauer, also found that the muscles are among the most resistant of structures. Giacomini (1888) thought the central nervous system very resistant, but Engel came to a contrary conclusion. Phisalix (1890) thought the epithelium the most easily affected by pathological processes, and while there is great difference of opinion, all investigators seem to be agreed that the cyema is affected before the vesicles.
Von Winckel (1904) spoke of three grades of maceration in the cyema, the first being that of bleb formation, the second that in which the epidermis is broken and shed in fragments, thus exposing the chorion, and the third stage that in which the joint capsules and ligaments are loosened, the joints consequently relaxed, and the extremities contorted. Von Winckel also set an approximate time-limit for the occurrence of these stages, the last stage being reached in three weeks. Nevertheless, von Winckel concluded that the degree of maceration could not be used as a criterion for the determination of the duration of intrauterine retention. Muller also was impressed by the post-mortem changes, and rightly stated that many embryos are deformed beyond recognition by maceration, bleb formation, and mummification.
Although it is incorrect to regard the changes which many conceptuses undergo after death and before abortion as maceration in the customary sense, no other word seems to be available. These changes undoubtedly are accompanied by softening, collapse, deformation, and no doubt also by autolysis. Whether imbibition and swelling are invariably present I do not know, but if they occur they usually are slight and their effects minor in character. Most of the changes enumerated here may occur under sterile conditions, and since maceration, even in this restricted sense, must effect changes, not only in the chemical composition, but also in the microscopic structure of cells, long before changes in external form of embryo or chorionic vesicle become apparent, a discussion of maceration changes really should begin with these. But the earliest modifications in microscopic structure remain very largely, and those in chemical composition almost wholly, unknown to us at present.
Although it undoubtedly is true, as stated by Miiller (1847), that very young embryos may retain their form surprisingly long after death if retained in sterile amniotic fluid, it does not therefore follow that they really remain structurally unchanged. However, I can not confirm the opinion of Herzog (1898) that chorionic villi degenerate with astonishing rapidity after the death of the embryo, which Herzog claimed was true especially in young placentae. According to Herzog, only the most intimate acquaintance enables one to recognize the villi after the embryo has been dead two or three weeks. Since the villi are usually the last thing to disappear and may survive the death of the embryo by months, even in the case of early conceptuses, it is difficult to understand what may have been responsible for Herzog's opinion. Leopold (1882) stated that it has been shown that a bare fetus in the peritoneal cavity will macerate and become disarticulated, but that a fetus surrounded by the intact membranes will become dry and leathery. Just what the basis for this statement is I do not know, but it would seem that comparatively prompt disarticulation of any except perhaps very young cyemata must imply the presence of putrefactive conditions, for the length of time which dead tissues can survive depends very largely upon the advent of putrefaction.

Microscopic changes no doubt appear quite promptly after death in all conceptuses, but, strange as it may seem, the most pronounced internal changes may sometimes fail to manifest themselves externally. In one instance (No. 962), for example, Mall had noted that the shape of the embryo had been preserved so perfectly that the specimen seemed normal in form. Yet the slightest jar on the containing vessel resulted in its complete disintegration. Apparently this specimen was somewhat farther advanced than embryo No. 2197 (figure 29, plate 4, Chap. IV). The same thing may be true of the villi, the shape of which may be preserved perfectly, although structurally they may have become mere gossamers, as illustrated by the villi of No. 606, shown in figure 11 (plate 1, Chap. IV).


Since very few abortuses are expelled promptly, practically all are macerated to a greater or lesser degree. This applies to specimens classed as normal, as well as to those classed as pathologic. Nor can the mere fact that a specimen was obtained in ute.ro at operation assure one that it is not macerated. No. 1224, for example, although obtained at operation, is a very greatly macerated, empty chorionic vesicle which was isolated completely in the uterine cavity. In another specimen (No. 1767), also obtained at hysterectomy, an abscess is found within the implantation site. No. 782, a third specimen of the same kind, and also other hysterectomy specimens, as previously stated, contain young hydatiform vesicles. Although the uterus and conceptus of No. 872 were placed in 10 per cent formalin after operation and kept in a thermostat for two days, the villi nevertheless are markedly macerated. They are also almost wholly non- vascular, remnants of the vessels being present in some of the villi only. The amnion is absent, the fibrous chorion also shows maceration changes, and the decidua is decided^ infiltrated. Hence it is important to remember that unless one is dealing with a normal implantation and a wholly normal uterus, the fact that the specimen was obtained at operation and preserved immediately with the best of care is not an absolute guarantee against the presence of early maceration changes. This is illustrated especially well by tubal specimens, which so frequently are embedded or isolated in blood-clot, as was the case with Nos. 1938 and 2035, shown in figures 179 and 181. The embryo and yolk-sac of the former are almost completely disintegrated, for nothing but scarcely recognizable remnants lie isolated in the chorionic cavity, which is moderately filled with an amorphous coagulum. The stroma of the chorionic membrane is edematous and degenerate, but contains some wellpreserved vessels, a few of which contain some blood-cells. The same thing is true of the stroma and of the vessels of the villi, which also are in process of dissolution. A moderate amount of trophoblast is present, but there is very little syncytium. The epithelium of some of the villi has undergone hyaline degeneration. The blood-cells in the large clot in which this chorionic vesicle, measuring 8 by 5 mm. in section, was embedded, are preserved fairly well, especially near the vesicle. Nevertheless, the whole conceptus is very apparently in a state of rapid disintegration and lysis. The tube-wall is very thin and the mucosa congested, hemorrhagic, and atrophic.



It is not difficult, as a rule, to identify maceration changes in the villi, for the appearances are rather characteristic, as figures 182 and 183 illustrate. But when other changes, such as the obliterative process shown in figure 184, are present at the same time, it is sometimes impossible to decide whether such changes as these were ante-mortem or post-mortem. In this case the lumina of the capillaries are plugged by what reminded Mall of "epithelial pearls," though he recognized that the cells were endothelial in origin. That such an obstruction of the lumina of blood-vessels can occur through a shedding, swelling, and clumping of the endothelial cells there can be little doubt, although proliferation of the endothelium can perhaps not be wholly excluded.

The earliest noticeable external post-mortem change in the color of the embryo is due to the occurrence of a greater opacity. The tissues lose their normal translucence, so that they look whiter and denser, and the surface also becomes less glistening. Later they also become yellowish and occasionally hemorrhagic, and the cutaneous surface becomes less smooth. Coincident with these changes, softening and some swelling also take place, as illustrated by No. 2146, shown in figure 187, and bleb formation occurs, especially on the abdominal extremity of the umbilical cord and elsewhere, as previously shown in figures 54, 70, 71, and 72 (plate 5, Chap. IV).
In consequence of the softening of the tissues, the upper extremities, if sufficiently developed, tend to gradually droop, as shown in figures 58 and 66 (plate 5, Chap. IV). Later, the caudal extremities also sag, and both pairs may gradually develop unusual curvatures. Changes occur also in the face, for the ocular margins become irregular and everted, or, if the lids have fused, they may prematurely open. But all these are relatively slight gross changes and merely herald the more profound modifications in form if retention continues under favorable conditions. What is now needed is a correlation between these early changes in external form and appearance and the histologic and cytologic changes. Although the advent of changes in color is quite prompt, it can safely be assumed from other evidence that the incipient structural changes are microscopic and can be detected long before changes in external form and color occur.

Since the chorionic vesicle almost invariably is surrounded by more or less blood and decidua in cases of abortion, it is naturally difficult to detect early gross post-mortem changes in it by inspection alone. However, the earliest noticeable external change in the villi is a decrease in translucency and a dulling of their surface. The villi of very young specimens also seem to become somewhat larger in caliber and look stiffer and slightly bulbous in places. They also frequently are somewhat matted, as was the case in No. 1878, a very young specimen, a portion of which is shown in figure 188, the chorionic vesicle of which measured but 10 by 12 mm. However, in No. 1843, which is a still younger specimen, measuring 5 by 6.5 mm. (figure 7, plate 1, Chap. IV) and shows more marked maceration changes upon microscopic examination, these changes, nevertheless, were not evident to the unaided eye, for the vesicle was unusually translucent. The chorionic and villous epithelium has sloughed over most of this vesicle and the remaining tissues stain but weakly. The cell-boundaries also are rather indistinct and the yolk-sac shows marked changes.


The first noticeable change in histologic structure seen in routine preparations seems to be a blurring of the structural detail in the tissues and a haziness in appearance of the cytoplasm. This early state is followed, if not preceded, by autolytic changes. As has so frequently been emphasized, evidences of disintegration, noticeable particularly in the central nervous system, supervene very early and the brain and cord become swollen and the folds become effaced, as shown in figure 185. These changes are accompanied and may be followed by complete dissociation of the constituent elements, with some shifting if not migration of cells. All the constitutent cells become rounded, the cytoplasm may disappear entirely, and the nuclei become more pycnotic. When this stage is reached the structure of the brain and cord is quite uniform and cells of various origins may all be quite properly designated as round cells, for all look alike, the changed nuclei only remaining. The entire central nervous system, for example, may be composed of these densely packed round cells and be characterized quite properly upon superficial appearances alone as lymphoid, as suggested by Giacomini. Finally, however, the nuclei also disintegrate and form a fine, granular material, spoken of by Mall as nuclear dust. Morever, not only the nervous system, but all the organs of small fetuses, may be so transformed, as illustrated in figure 186.
These things, however, do not necessarily or materially change the form of the cyema, and the effects produced by retention in utero after death will depend not so much upon the exact duration of the retention as upon the age of the cyema. A few weeks may suffice not only to change greatly the external form or the internal structure of young cyemata, but to effect complete absorption. In case of an approximately mature fetus, on the other hand, such a short period of retention after death might modify the form inappreciably and result merely in maceration of the epidermis.
One might expect that maceration, even under sterile conditions, especially in case of young specimens, would result in general distention, but this does not always seem to be the case. Edematous areas do, indeed, frequently form locally, as previously illustrated and as generally observed; but swelling due to imbibition, such as occurs when fresh young specimens are placed in formaldehyde, was observed only in a minor degree, as illustrated by No. 2146, shown in figure 187. Sometimes this swelling is local, as represented by the cephalic region of No. 1750. A subsequent reduction in size is much more common and may be due to absorption of salts from the amniotic fluid, with resultant concentration of it and of extraction of tissue fluid from the cyema itself. In very young specimens, such as No. 786 (fig. 189), an embryo 4 mm. long, the first noticeable shrinkage change in form seems to occur in the cephalic extremity, the frontal prominence of which recedes and the outline of the vertex of which becomes more and more rounded and also is reduced in size.
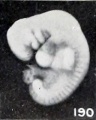
Figure 190
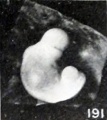
Figure 191
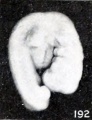
Figure 192



In consequence of these changes, the cephalic extremity becomes relatively too small, as is well illustrated by comparing this specimen with No. 1380, shown in figure 190, a cyema of the same development but 5.5 mm. long and in an excellent condition of preservation. This reduction in size of the cephalic region would, to be sure, result in a reduction in length were the measurement taken from tip to tip, and were it not for the fact that during this process the cephalic extremity, if indeed not the entire embryo, usually unbends and in becoming straighter compensates for, or even more than compensates for, the loss in length due to reduction in size of the cephalic extremity. This does not always occur, however, as Nos. 1299 and 2216 so well illustrate (see figs. 191 and 192). These changes make the trunk look disproportionately large and the features somewhat stunted. Previous to these changes, or coincident with them, the tissues lose their elasticity and also become softer, and the surface of the specimen becomes duller, more opaque, and finally more yellowish. Other changes noticeable in the specimen just referred to are the loss in detail of the surface relief, evident particularly in the branchial region, and in the effacement of the myotomes. In other instances, such as No. 2035 (a portion of a tubal specimen), a marked change is very evident in the caudal extremity, which has become shorter, blunter, and straighter, although upon close inspection other changes also are noticeable. Because of these things the actual soon may become less than the original length and only roughly indicate the true age of the cyema.
The change in the cephalic extremity, the decrease in the natural curve which accompanies erection of the specimen, and the loss in detail just spoken of are illustrated still better by No. 187a, which is a somewhat older embryo, 7 mm. in length. Young embryos which illustrate the early changes in form are Nos. 208 and 1296, shown in figures 193 and 194 embryos which are respectively 7 and 4 mm. long in their present state.
A somewhat similar, though slighter, effect of maceration upon relatively young specimens is illustrated by Nos. 1697 and 1477, shown in figures 195 and 196, cyemata 15 and 18.5 mm. long respectively. In the first of these two specimens the gaping mouth and blunted and rounded upper extremities are especially noticeable. That sagging of the upper extremities does not always occur, even in the presence of a more marked degree of maceration than present in the above specimen, is illustrated by No. 705, shown in figure 197, a somewhat older cyema with more marked maceration, and to a less degree also by No. 1477, both of which also show a rounding and shortening of the extremities.
A still more striking series of transformations is illustrated by the somewhat older specimens, Nos. 2244, 1891, 1655, 1260, 1333, 1926, and 1379, shown in figures 198 to 204 inclusive. Inspection of this series will show that the transition from one to the succeeding specimen is not very marked, although the difference in form between the first and last specimens is very great indeed, and could have been made more striking by accompanying each figure by a normal one of the same (or approximately the same) stage of development.
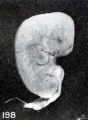
Figs. 198. Similar specimens, showing more pronounced changes. Nos. 2244 (X2.67).
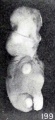
Figs. 199. Similar specimens, showing more pronounced changes. Nos. 1891 (X2).
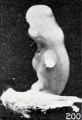
Figs. 200. Similar specimens, showing more pronounced changes. Nos. 1655 (X2.67).

Figs. 201. Similar specimens, showing more pronounced changes. Nos. 1260 (X2.67).
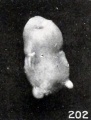
Figs. 202. Similar specimens, showing more pronounced changes. Nos. 1333 (X2.67).

Figs. 203. Similar specimens, showing more pronounced changes. In figure 203 the structure of the specimen is chaotic. 2244 (X2.67), 1891 (X2), 1655 (X2.67), 1260 (X2.67), 1333 (X2.67), 1379 (X2.67).
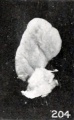
Figs. 204. Similar specimens, showing more pronounced changes. 2244 (X2.67), 1891 (X2), 1655 (X2.67), 1260 (X2.67), 1333 (X2.67), 1379 (X2.67).







Such oddly shaped bodies as those shown in figures 203 and 204 scarcely can be identified positively as cyemic remnants by the unaided eye, and even under some magnification give little indication of their internal structure. The same thing is true, though to a lesser degree, of Nos. 885 and 1333, shown in figure 202, and also in figure 22 (plate 3, Chap. IV). In these specimens the limbs are still present in such an abbreviated form that the latter suggests amelia. Hence the question at once arises whether these specimens show mere post-mortem deformations or true developmental anomalies. This question is presented still more forcibly by smaller, younger specimens such as Nos. 1226 and 2361, shown in figures 205 and 206. These cyemata may be truly anomalous, but they also may be macerated young normal embryos, and in case of young specimens the question can be decided only by a microscopic study of the sections, as was illustrated while considering the nodular group in Chapter IV. It must be borne in mind, however, that minor external deformities may be obliterated completely by post-mortem deformations, and that in such cases no positive conclusion can then be reached.

An unbending or erection of the cyema does not always occur, as is illustrated by No. 208 (fig. 191), which also is a 7-mm. specimen. Although this cyema is markedly macerated, there has been but little change in the total outline, save as a result of pitting, increase in flexure, and shrinkage. Whether or not failure to unbend on the part of this specimen is due to an early coagulation of the amniotic fluid, and the tissues of the embryo itself, I do not know, but that this might be an important factor does not seem improbable. Similarly, early coagulation of the cyema itself might postpone other changes in form, similar to those shown in No. 589 (fig. 207), a cyema of 10 mm. This cyema very apparently was not surrounded by a coagulum. In such soft specimens the whole surface may be wrinkled, the hand plates may look excavated, and the head may be sunken deeply upon the chest. Since this particular specimen remained extremely soft and pliable, the extremities also were rotated somewhat and the body became rather collapsed, as if compressed. Just why some cyemata remain relatively stiff, firm, and opaque, and others become extremely soft, swollen, and more translucent, remains unknown to me, but it is not impossible that the reaction of the amniotic fluid is the controlling factor.
In slightly older specimens the first marked changes in external form resulting from maceration sometimes are seen near the abdominal attachment of the umbilical cord, as illustrated in No 1523 (fig. 70, plate 5, Chap. IV), an embryo of 19 mm. Several blebs are sometimes located on opposite sides of the cord or surround it, as in the case of No. 1475, shown in figure 54 (plate 5, Chap IV). It is interesting that large blebs not infrequently are present in this location, even if the body of the cyema has suffered relatively little change. The size of these blebs, to be sure, may be considerable and, instead of local edema, the entire epidermis may be lifted and the entire cord may be swollen, but such a condition usually arises only later, during longer periods of retention. Sometimes the whole nuchal region is edematous, forming a marked prominence far above that normally present, or is occupied by a single large bleb. For some reason these blebs are common, especially on the head, the dorsum, and on the umbilical cord.

Sometimes a prominence on the dorsum is due to the swollen, macerated, and completely disintegrated central nervous system, which may form a marked ridge, as illustrated by No. 521/, shown in figure 208, a specimen in which lumbar spina bifida was present. Not infrequently, however, a marked prominence in the upper dorsal region is due to collapse of the brain and drooping of the head, as illustrated in No. 175, a cyema of 13 mm. This collapse of the central nervous system occurs most frequently in the brain of older specimens and is not infrequently followed by collapse of the calvarium in younger specimens. It may result in marked cutaneous depressions or sulci, as illustrated in No. 1250, a specimen 13 mm. in length (fig. 209). Rarely, these sulci or ridges may extend entirely down the dorsum of young specimens.

It is decidedly significant that some intrauterine changes in external form can be simulated by post-mortem extrauterine changes. This is splendidly illustrated by figures 210 and 211, which are photographs of a fresh and well-preserved and a poorly preserved cat embryo respectively. Similar changes have also been produced experimentally by incubation in sterile solutions, and are further illustrated by No. 1358/, shown in figures 212 and 213. The former shows the appearance of the somewhat macerated specimen before staining and preservation in alcohol, which very materially shrunk the tissues, as shown in the latter. Were the history of this case unknown, one would have been justified in calling it a rather decidedly soft, macerated specimen. Nor do two specimens of identical age, which existed under identical conditions, necessarily present exactly the same appearance. This is well illustrated by Nos. 2258a and 22586, single-ovum twins, shown in figure 214.
The relatively early loss in detail in the normal relief of the cyema is illustrated extremely well by No. 2014 (fig. 215), an embryo 17 mm. long. The normal relief of this specimen is replaced by a molding due to maceration. These changes, which are noticeable over the entire body, show particularly in the eyes and ears and in the gaping mouth. The effacement of the normal features in young specimens is sometimes very complete, but this is true also of older ones, so that the external form may suggest the amorphous group. An intermediate form between the latter and No. 2014 is represented by No. 1495d (fig. 216), a cyema 12.3 mm. long, but of approximately the same stage of development as the other two. A still more advanced change of this type is illustrated by No. 921, shown in figure 39 (plate 4, Chap. IV). The difference in length between some of these specimens of corresponding development is not infrequently due mainly to shrinkage accompanying maceration and retention. No. 1495d is interesting also because the placental site was marked internally by numerous subchorial hematomata (fig. 217). I do not wish to enter into a discussion of these so-called "Breus hematomatous moles" of which so much seems to have been made, but all the specimens which have come to my attention had been retained for a long time. It would seem that when, for some reason, the placental margins and also certain intermediate placental areas are more firmly adherent than the rest, the looser portions may be detached and forced inward by hemorrhage, thus producing a series of hummocks on the internal surface of the placenta. If the chorion and amnion over these areas rupture, blood may enter the amniotic fluid, and just in proportion as the latter is reduced in quantity the process of eversion of the placental site must be facilitated. It is, of course, entirely possible that a mixture of blood with the amniotic fluid, so common in these specimens, may help to preserve the cyema, but the abortuses with subchorial hematomata which I have seen did not give me the impression that their preservation was any better than that of the non-hematomatous type.
Blunting and rounding of the extremities of the cyema, with consequent shortening and loss of detail, are present in still more decided form than previously illustrated. Beginning sagging of the extremities is shown well in No. 1358, which is 18.3 mm. long, and in still older specimens, such as No. 1710 (fig. 66, plate 5, Chap. IV). The abnormal curvatures, and especially the blunting of the extremities, are exemplified still better, and the drooping mandible, gaping mouth, and locally edematous cord are also well shown in No. 797, a fetus 35 mm. long, represented in figure 218. Changes in curvature of the extremities are splendidly illustrated in No. 1860 (fig. 219), and become more pronounced the older the specimen, up to the period when skeletal development makes them extremely difficult or even impossible.
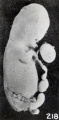
Figure 218

Figure 219
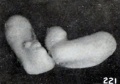
Figure 221

Figure 222
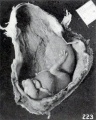
Figure 223




The changes in the extremities do not stop, however, with the production of abnormal curvatures, for not infrequently the hands and feet undergo an outward clubbing, shown in pronounced form in No. 1958 (fig. 220), a decidedly macerated specimen 41.5 mm. long. Separate views of the extremities of this specimen are given in figures 221 and 222. Earlier stages in this process are illustrated by No. 1751 (fig. 223) , with separate views of the extremities in figure 69 (plate 5, Chap. IV) .
Since the extremities in earlier stages of development are less resistant, they become soft and are more easily contorted ; but at a somewhat later stage of development, when they are more resistant, it is not uncommon to find the arms especially assuming more extended positions of gesture. Hence grotesque figures occur, such as exemplified in Nos. 1462 and 1309 (figures 224 and 225), which are purely accidental forms. As maceration becomes extreme, even without the presence of putrefaction, marked softening and distortion take place and the face may be drawn into grimaces, as illustrated by No. 1775 (fig. 226) and by other specimens previously represented. In these older specimens the epidermis, instead of falling off in flakes or hanging in streamers, especially from the extremities of the digits, may become rolled up and accumulate in welts, especially at points of contact of the extremities, as illustrated in No. 1859 (fig. 64, plate 5, Chap. IV).
If dehydration occurs under sterile conditions, the transition from such a specimen as No. 1859 to 1474a, shown in figure 61 (plate 5, Chap. IV.), is an easy one, and if continued beyond this stage it may eventually lead to typical mummification. If the amniotic fluid is finally completely or almost completely absorbed and the uterus then contracts upon the specimen, such rolled-up forms as Nos. 1525, 1976 (figures 227 and 228) and No. 1041 (figure 40, plate 4, Chap. IV.) may result. Sometimes the placenta and membranes of these long-retained, leathery specimens are found in the form of a firm covering with a small opening through which the fetus has escaped, as illustrated by No. 1850, a fetus of 17 mm., shown in figure 229.
In other instances the fetal tissues, instead of becoming dehydrated, as in case of the above and No. 1295a, shown in figure 230, remain soft, as represented in a comparatively early stage by No. 1350, 66 mm. long, shown in figure 61 (plate 5, Chap. IV).
A more advanced stage of this process is represented by Nos. 2034 and 1925, fetuses 96.5 and 147 mm. long respectively, shown in figure 234 and figure 48 (plate 5, Chap. IX). If this process of maceration becomes extreme, or if putrefaction supervenes, such disintegrated forms as No. 1515 (fig. 50, plate 5, Chap. IV) are finally produced, and the stage of disarticulation is at last reached.
That the advent of rigor mortis does not necessarily change the attitude of young embryos is suggested from occurrences observed personally under experimental conditions. As long as the amnion closely invests a mammalian embryo, the extremities of the latter can not become extended during rigor, and hence may retain the intrauterine position. The same thing holds, to a considerable extent at least, for the trunk also, for neither is it so free to extend as when the amnion has been opened or been removed. In the human conceptus the amnion soon becomes sufficiently large, however, but coagulum could have a similar influence.
Although little is known regarding intrauterine rigor mortis, Wolff (1903) stated that it is not rare, and held that an accumulation of waste products produced in consequence of circulating disturbances in the mother may be responsible for its early advent. In any case it probably is quite transitory, and the later rigidity of aborted cyemata undoubtedly must be due to something else.
At present we are not in position to accurately evaluate slight changes in bodily form of cyemata, for as long as the exact form of normal cyemata remains undetermined, it is inevitable that macerated specimens will be mistaken for and represented as normal in form, even in contemporary embryologies. Abnormal forms likewise have been described as normal, and macerated normal specimens also as pathologic, although no one would, I presume, defend the opinion that a dead conceptus may become pathologic.
Although His emphasized that the softness of aborted forms easily results in unusual folds and flexures, he, it seems, greatly reinforced, even if he did not introduce, the idea that embryos that are abnormal in form are pathologic. He apparently came to this conclusion because he did not believe that post-mortem changes could be responsible for modifications in form present in the specimens which came to his attention. Giacomini (1888) also believed that many of the deformities so common in embryos of the first months are due purely to regression after death, but added that he did not care to stress the idea that post-mortem changes can account for many of the deformities seen. Later Giacomini evidently became skeptical, however, and decided that it remained to be determined whether the changes in the embryo are primary or secondary. Furthermore, in 1894, he declared that one never meets with changes in abortive forms which one can attribute solely to softening, and concluded that embryos that die soften and disintegrate quickly, only those which merely are inhibited in growth being preserved for months. Waldstein (1913) also stated that von Winckel spoke of a slowing of development before death of the embryo occurred. While it is conceivable, and even probable, that inhibition in growth may occur, experiments now under way show that dead mammalian fetuses nevertheless may be preserved in sterile nutrient and non-nutrient solutions at body temperature for extended periods of time for months without indication of growth and with very little change in external form.
Although one must frankly recognize that it is extremely difficult to simulate intrauterine conditions closely, these experiments nevertheless very seriously question the conclusion of Giacomini. His (1891) also emphasized that dead embryos may be preserved in utero for months, and there is abundant evidence in the Carnegie Collection corroborating this conclusion. But it is conceivable that slight modifications in body-form might arise from gradual inhibition in growth, as well as from post-mortem changes of secondary origin. Panum (1860), in discussing what he termed monstruositates Males amorphoides occurring among chicks, also expressed the opinion that the changes in embryonic form noticed by him arose during life. He claimed that these changes could not have resulted from maceration, because adhesions of the membranes were present, because the specimens showed a total divergence from the normal form, and also because of the differences in size and consistency found to exist among them. Panum was led to the conclusion that all the deformities noticed by him in chicks were the result of the conditions under which development occurred and through the absence of evidence supporting the opinion of Bischoff that the cause of monsters was germinal. But Panum nevertheless believed that gases formed from putrefaction might compress the embryo and produce a fetus papyraceous.
When considering these greatly modified or even bizarre forms of human embryos, one also is reminded of the fact that Panum believed that monstrous chicks resulted directly from a lowering of the temperature during incubation.
However, since Panum found deformations especially common in the cephalic extremity, and further stated that many of his so-called "monstruositates totales cylindricse" were mummy-like, with features effaced, it seems more likely that he was dealing with maceration forms the death of which was due to the chilling. Just why some chick embryos, the heart-beat of which has been temporarily suspended by chilling, die quite promptly, and others only late in development, near the end of the normal incubation period, I do not know, but personal observations leave no doubt regarding this matter, and variation in vitality is all that I can suggest as an explanation at present.
It is only just to state that Panum considered the possibility that the anomalous forms were secondary in origin, and concluded that some were germinal in origin. His (1891), too, at first regarded certain abnormal forms of human embryos, such as the nodular, cylindrical, and flexed (geknickte), as germinal, but later inclined to the belief that they nevertheless arose from changes occurring after the death of the embryo, saying:
- "Seitdem ich mir aber Rechenschaft gegeben habe von den histologischeu Veranderungen welche eine Folge des Absterbens sind, von der Quellung des Gehirns und den Veranderungen der urspriinglichen Gewebe durch eine Zellenbrut, bin ich weit mehr geneigt, die abortiven Formen von Embryonen als secundar enstanden anzusehen. So werden speciell auch die so auffalligen Cylinderformen verstandlich, sowie man sich davon Rechenschaft giebt, dass von dem friiheren Embryo neben einem auffalligen Skelett fast nur noch die Haut ubrig geblieben ist. Als ausdehnbarer Sack kann sich dieselbe mit fremdem Material, mit Wanderzellen und zum Theil mit Fliissigkeit ausfiillen und nun giebt sie die alten Korperformen nur noch in den allergrobsten Ziigen wieder."
In spite of these words, His insisted that poorly preserved normal forms never show changes seen in so-called abortive forms, and emphasized that, although the cells in abortuses may not all be dead, we nevertheless can not speak of these abortive forms as living.
The mere fact that post-mortem changes may produce misleading modifications in form makes the identification of post-mortem deformations extremely important. If all conceptuses were aborted immediately after their death, the problem would be a far simpler one, for such an event would at once dispose of such difficult questions as, not only those of survival, but also of the independent growth of one or more members of the cyema, or of the whole of the vesicles, after the death of the cyema. That outward apparent stunting is a fairly common occurrence in young embryos is fully attested by the many specimens in the Carnegie Collection which illustrate this phenomenon, but I have not been able to find convincing proof indicating that it is also physiological. In the case of older fetuses with apparent brachydactyly, osseous development seems to have been normal and the modifications attributable to changes in the soft parts. Hence I am prompted to conclude that, although growth may be retarded in consequence of circulatory disturbances, it continues wholly normally, unless influenced by other things, until the heart stops.
It also may be difficult to tell whether a given condition arose before or after death of the embryo, merely because of the presence of maceration changes, even when these changes may have occurred in a sterile medium. The difficulty of distinguishing between ante-mortem and post-mortem changes, which impressed Phisalix and also others, is very great, if not insurmountable in some cases. This is true especially if it be assumed that an irregular or uncorrelated growth can occur under post-mortem, ante-partum conditions, for such growth could easily produce anomalous forms. But it must be admitted that the existence of a genuine postmortem growth is merety an inference at present.
In considering some of the strange forms to which attention has been called one can not disregard such instances in the newborn as that reported by Cowie (1914). The posture of this infant reminds one very strikingly of some of the fetuses in the accompanying illustrations. This seems very disconcerting at first thought, but there is, of course, no reason why a condition responsible for the occurrence of multiple intrauterine fractures, such as are present in this infant, should not result in very abnormal postures. Moreover, if fetal bone disease may begin very early, it is also probable that some of these strange, relatively .early fetal forms well may be genuine fetal anomalies and pathologic in addition.
The cases reported by de Lima (1915), especially cases 2 and 3, also belong among those which simulate some early fetal forms. Nevertheless, in these, and also in the other 2 cases reported by him, de Lima found bony defects. The same thing was true of the outwardly similar cases in infants and children reported by McKenzie (1897).
However, in view of the occurrence of these outward resemblances, it is not always possible to distinguish normal specimens which have suffered post-mortem deformations or changes in posture of the extremities from true developmental anomalies by outward inspection alone. This difficulty is particularly great in connection with changes in form and position of the hands, the feet, and the head and neck, and greatest of all, in connection with the knees. Even a cursory examination of some of the accompanying figures must also show that we at present are unable to decide whether any of these young specimens with pronounced deflections of the head and neck are cases of genuine torticollis and others are genuine cases of club-hand or club-foot, or genu varus, or valgum, for example. Indeed, this difficulty could be avoided only if these conditions arose only late in pregnancy, or if the cyemata showing them never were aborted before term. Waiving the exact definition of club-hand and club-foot, it must strike the attention of anyone that caput obstipum and apparent club-hands and club-feet so often are associated in these relatively early specimens, most of which are decidedly macerated. In 10 out of 21 cases of club-hand and club-foot listed as such among 3,000 accessions, excluding 2 cases with an embryonic length of 2.5 and 16 mm., respectively, clubbing was present in all the extremities. This is a wholly different relationship from that which obtains between these anomalies at the time of birth, when they seem to be associated but very rarely. However, in practically all of these cases of universal clubbing of macerated specimens the anomaly is associated with other developmental defects, as is not infrequently the case in the congenital condition.
Three of these 21 cases of clubbing were classed among the normal and 18 among the pathologic. With the exception of 3 fetuses which had a length of 100, 130, and 220 mm., respectively, all were less than 100 mm., the average length being 48.9mm. While it may be possible to recognize genuine clubbing, due to the absence of the bones or to defects in them so early in fetal life, by microscopic examination or by other special methods, this certainly can not be done by inspection alone. Hence it is clear that the term dub-foot, as used in the classification of this collection, is without special implication, except that it tells something about the shape of the periphery of the extremities.
It would also seem that if all these cases of deformed extremities in fetuses were true developmental anomalies, they should be encountered in well-preserved specimens, and more frequently in the older ones. That other anomalies should be found associated more frequently also seems to follow from our knowledge of congenital club-hand and club-foot. Among these genuine cases defective development of the bones seems to be more frequently noted, for congenital club-hand nearly always is accompanied by absence of either radius or ulna, while in the cases under consideration here such an absence has not been established. Nor is it without significance that the most pronounced forms of club-hand and clubfoot found in these fetuses always occurred in those longest retained and most macerated. Many of them also show quite general evidences of the presence of pressure defects, and unless it can be shown that a tendency to progressive elimination of specimens with double club-foot and hand exists, with survival of those suffering from club-foot alone, one can not harmonize the frequent association of these defects in fetuses with the conditions as known to exist at the time of birth. However, since certain forms of congenital club-foot are probably also the result of intrauterine pressure, the types found in fetuses if the latter survived would be quite similar or even identical, in a morphologic sense, with the condition in the newborn; yet in one case growth would have ceased before the pressure became effective, while in the other it continued. The fact that congenital clubfoot is relatively seldom due to bony defects would also seem to point to external factors as causes, but I have no final opinion on this matter and am calling attention to it merely to emphasize the fact that post-mortem intrauterine changes resulting from maceration and pressure, or from both, may easily cause confusion in young cyemata.
Description of Plates
Plate 17

Fig. 175. A portion of a chorionic vesicle, showing appearances identical with tissue cultures. No. 545. X135.
Fig. 176. Fetus showing gluing of hand to face. No. 316.
Fig. 177. Villi showing obliteration of vessels by maceration and disintegration. No. 640.
Fig. 178. Villi showing obliteration of vessels by proliferation. No. 317. X97.5.
Fig. 179. Macerated tubal specimen imbedded in clot and undergoing lysis. No. 1938. X0.75.
Fig. 180. Fetus showing continuity of epidermis across the mouth, with obliteration of the labial slit. No. 885. X 4.5.
Fig. 181. Macerated tubal specimen, only the chorionic vesicle remaining. No. 2035. X0.75.
Fig. 182. Slightly macerated villi. No. 275. X37.5.
Fig. 183. Greatly macerated villi. No. 7236.
Fig. 184. Macerated, long-retained villi, with lumina of capillaries plugged with coagulum. No. 286. X37.5.
Fig. 185. Fetus in sagittal section showing maceration, especially of the nervous system. No. 285. X4.5.
Fig. 186. Cross section of cyema showing homogeneous structure produced by maceration. No. 205. X 11.25.
- Plate 18: Figs. 187 | Figs. 188 | Fig. 189 | Fig. 190 | Fig. 191 | Fig. 192 | Fig. 193 | Fig. 194 | Fig. 195 | Fig. 196 | Fig. 197 | Fig. 198 | Fig. 199 | Fig. 200 | Fig. 201 | Fig. 202 | Fig. 203 | Fig. 204 | Fig. 205 | Fig. 206 | Fig. 207 | Fig. 208 | Fig. 209 | Fig. 210 | Fig. 211 | Fig. 212 | Fig. 213 | Fig. 214 | Fig. 215 | Fig. 216 | Chapter 13 Post-Mortem Intrauterine Changes

Fig. 175. A portion of a chorionic vesicle, showing appearances identical with tissue cultures. No. 545. X135.
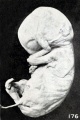
Fig. 176. Fetus showing gluing of hand to face. No. 316.
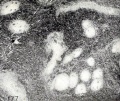
Fig. 177. Villi showing obliteration of vessels by maceration and disintegration. No. 640.
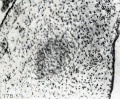
Fig. 178. Villi showing obliteration of vessels by proliferation. No. 317. X97.5.

Fig. 179. Macerated tubal specimen imbedded in clot and undergoing lysis. No. 1938. X0.75.
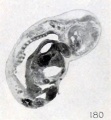
Fig. 180. Fetus showing continuity of epidermis across the mouth, with obliteration of the labial slit. No. 885. X 4.5.

Fig. 181. Macerated tubal specimen, only the chorionic vesicle remaining. No. 2035. X0.75.
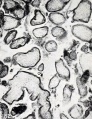
Fig. 182. Slightly macerated villi. No. 275. X37.5.

Fig. 183. Greatly macerated villi. No. 7236.
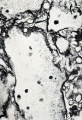
Fig. 184. Macerated, long-retained villi, with lumina of capillaries plugged with coagulum. No. 286. X37.5.
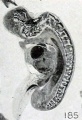
Fig. 185. Fetus in sagittal section showing maceration, especially of the nervous system. No. 285. X4.5.
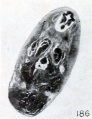
Fig. 186. Cross section of cyema showing homogeneous structure produced by maceration. No. 205. X 11.25.

Plate 18

Fig. 187. Slight swelling of fetus from brief maceration. No. 2146. X2.
Fig. 188. Matted, slightly macerated villi. No. 1878. X4.
Fig. 189. A macerated, disproportional cyema 4 mm. long, showing a development of 5.5 mm. No. 786. X4.
Fig. 190. A well-preserved cyema 5.5 mm., of the same development as the preceding. No. 1380. X4.
Figs. 191-192. Cyemata illustrating failure of extension of the body upon maceration. Nos. 1299 ( X2) and 2216 ( X4).
Figs. 193-196. Cyemata illustrating changes in form due to maceration. Nos. 208 (X2), 1296 (X2.67), 1697 (X2), and 1477 (X2).
Fig. 197. Illustrating beginning changes in form due to maceration. No. 705. X1.35.
Figs. 198-204. Similar specimens, showing more pronounced changes. In figure 203 the structure of the specimen is chaotic. Nos. 2244 (X2.67), 1891 (X2), 1655 (X2.67), 1260 (X2.67), 1333 (X2.67), 1379 (X2.67).
Figs. 205-206. Doubtful normally developed cyemata. Nos. 1226 (X2.67) and 2361 (X4).
Fig. 207. A cyema illustrating post-partum changes. No. 589. X2.67.
Fig. 208. Cyema showing maceration sulci and ridges, and drooping of the limbs. No. 52 If. X1.66.
Fig. 209. Cyema showing maceration sulci and ridges, and swelling of the cord. X2.
Fig. 210. Normal, well-preserved cat fetus.
Fig. 211. Normal, poorly preserved cat fetus of approximately the same length.
Fig. 212. Appearance of fetus before fixation. No. 1358. X2.
Fig. 213. The same specimen, showing wrinkling due to fixation and staining. X2.
Fig. 214. Macerated, distorted single-ovum twins. No. 2258. X2.67.
Fig. 215. Minor changes in relief, due to maceration. Swelling and constriction of cord. No. 2014. X2.
Fig. 216. An intermediate maceration form. No. 1495d. X2.67.
- Plate 18: Figs. 187 | Figs. 188 | Fig. 189 | Fig. 190 | Fig. 191 | Fig. 192 | Fig. 193 | Fig. 194 | Fig. 195 | Fig. 196 | Fig. 197 | Fig. 198 | Fig. 199 | Fig. 200 | Fig. 201 | Fig. 202 | Fig. 203 | Fig. 204 | Fig. 205 | Fig. 206 | Fig. 207 | Fig. 208 | Fig. 209 | Fig. 210 | Fig. 211 | Fig. 212 | Fig. 213 | Fig. 214 | Fig. 215 | Fig. 216 | Chapter 13 Post-Mortem Intrauterine Changes
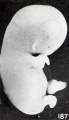
Fig. 187. Slight swelling of fetus from brief maceration. No. 2146. X2.
Fig. 188. Matted, slightly macerated villi. No. 1878. X4.
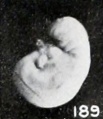
Fig. 189. A macerated, disproportional cyema 4 mm. long, showing a development of 5.5 mm. No. 786. X4.
Fig. 190. A well-preserved cyema 5.5 mm., of the same development as the preceding. No. 1380. X4.
Figs. 191. Cyemata illustrating failure of extension of the body upon maceration. No. 1299 ( X2).
Figs. 192. Cyemata illustrating failure of extension of the body upon maceration. No. 2216 ( X4).
Figs. 193. Cyemata illustrating changes in form due to maceration. No. 208 (X2).
Figs. 194. Cyemata illustrating changes in form due to maceration. No. 1296 (X2.67).
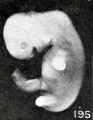
Figs. 195. Cyemata illustrating changes in form due to maceration. No. 1697 (X2).
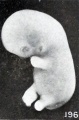
Figs. 196. Cyemata illustrating changes in form due to maceration. No. 1477 (X2).
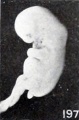
Fig. 197. Illustrating beginning changes in form due to maceration. No. 705. X1.35.
Figs. 198. Similar specimens, showing more pronounced changes. No. 2244 (X2.67).
Figs. 199. Similar specimens, showing more pronounced changes. No. 1891 (X2).
Figs. 200. Similar specimens, showing more pronounced changes. No. 1655 (X2.67).
Figs. 201. Similar specimens, showing more pronounced changes. No. 1260 (X2.67).
Figs. 202. Similar specimens, showing more pronounced changes. No. 1333 (X2.67).
Figs. 203. Similar specimens, showing more pronounced changes. No. 1379 (X2.67).
Figs. 204. Similar specimens, showing more pronounced changes. 2244 (X2.67), 1891 (X2), 1655 (X2.67), 1260 (X2.67), 1333 (X2.67), 1379 (X2.67).
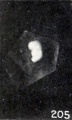
Figs. 205. Doubtful normally developed cyemata. No. 1226 (X2.67).

Figs. 206. Doubtful normally developed cyemata. No. 2361 (X4).
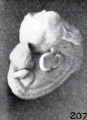
Fig. 207. A cyema illustrating post-partum changes. No. 589. X2.67.
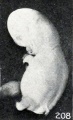
Fig. 208. Cyema showing maceration sulci and ridges, and drooping of the limbs. No. 52 If. X1.66.
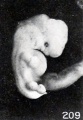
Fig. 209. Cyema showing maceration sulci and ridges, and swelling of the cord. X2.
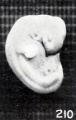
Fig. 210. Normal, well-preserved cat fetus.

Fig. 211. Normal, poorly preserved cat fetus of approximately the same length.
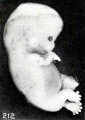
Fig. 212. Appearance of fetus before fixation. No. 1358. X2.
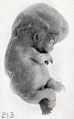
Fig. 213. The same specimen, showing wrinkling due to fixation and staining. X2.
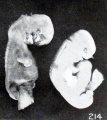
Fig. 214. Macerated, distorted single-ovum twins. No. 2258. X2.67.
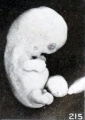
Fig. 215. Minor changes in relief, due to maceration. Swelling and constriction of cord. No. 2014. X2.
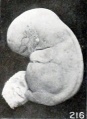
Fig. 216. An intermediate maceration form. No. 1495d. X2.67.















Plate 19

Fig. 217. Interior of the chorionic vesicle showing subchorial hematomata. No. 1495d. X0.77.
Fig. 218. A fetus and cord showing marked maceration changes. No. 797. X1.35.
Fig. 219. An older fetus, showing bleb-formation and curvature in extremities, due to maceration and retention. No. 1860. XI- 35.
Fig. 220. A fetus showing marked clubbing of the extremities and obliteration of the features. No. 1958. X0.87.
Figs. 221-222. Extremities of same specimen. X2.67.
Fig. 223. External appearance of fetus in situ. No. 1751. X0.57.
Fig. 224. An older, macerated fetus, with extremities extended instead of folded. No. 1462. X0.77. FIG. 225. A similar specimen. No. 1309. X0.23.
Fig. 226. A long-retained, soft, rolled-up form, photographed in extension. No. 1775. X0.47.
Fig. 227. A similar specimen with markedly flexed head. No. 1525. X0.66.
Fig. 228. A similar specimen. No. 1976. X0.47.
Fig. 229. A similar specimen with mummified chorionic vesicle. No. 1850. X6. FIG. 230. A decidedly mummified fetus. No. 1295a. X0.77.
Fig. 231. Unilateral development of villi in a vesicle classed as normal (?). No. 2092. (See Chapter XV.) X1.35.
Fig. 232. A somewhat macerated young vesicle with quite uniformly distributed villi which are slightly abnormal in form and structure. No. 1878. (See Chapter XV.) X4.
Fig. 233. A portion of same specimen, showing the somewhat more than normally bulbous and rather matted villi. X2.67.
Fig. 234. A very softened, macerated fetus. X0.5.
- Plate 19: Figs. 217 | Figs. 218 | Fig. 219 | Fig. 220 | Fig. 221 | Fig. 222 | Fig. 223 | Fig. 224 | Fig. 225 | Fig. 226 | Fig. 227 | Fig. 228 | Fig. 229 | Fig. 230 | Fig. 231 | Fig. 232 | Fig. 233 | Fig. 234 | Chapter 13 Post-Mortem Intrauterine Changes
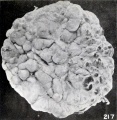
Fig. 217. Interior of the chorionic vesicle showing subchorial hematomata. No. 1495d. X0.77.
Fig. 218. A fetus and cord showing marked maceration changes. No. 797. X1.35.
Fig. 219. An older fetus, showing bleb-formation and curvature in extremities, due to maceration and retention. No. 1860. XI- 35.
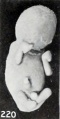
Fig. 220. A fetus showing marked clubbing of the extremities and obliteration of the features. No. 1958. X0.87.
Figs. 221-222. Extremities of same specimen. X2.67.
Figs. 221-222. Extremities of same specimen. X2.67.
Fig. 223. External appearance of fetus in situ. No. 1751. X0.57.
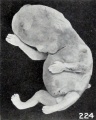
Fig. 224. An older, macerated fetus, with extremities extended instead of folded. No. 1462. X0.77.
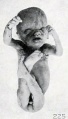
Fig. 225. A similar specimen. No. 1309. X0.23.
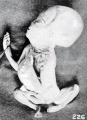
Fig. 226. A long-retained, soft, rolled-up form, photographed in extension. No. 1775. X0.47.
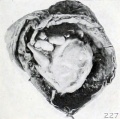
Fig. 227. A similar specimen with markedly flexed head. No. 1525. X0.66.
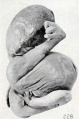
Fig. 228. A similar specimen. No. 1976. X0.47.
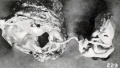
Fig. 229. A similar specimen with mummified chorionic vesicle. No. 1850. X6.
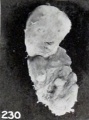
Fig. 230. A decidedly mummified fetus. No. 1295a. X0.77.
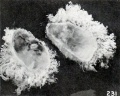
Fig. 231. Unilateral development of villi in a vesicle classed as normal (?). No. 2092. (See Chapter XV.) X1.35.
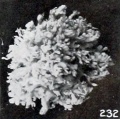
Fig. 232. A somewhat macerated young vesicle with quite uniformly distributed villi which are slightly abnormal in form and structure. No. 1878. (See Chapter XV.) X4.
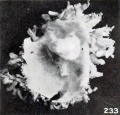
Fig. 233. A portion of same specimen, showing the somewhat more than normally bulbous and rather matted villi. X2.67.

Fig. 234. A very softened, macerated fetus. X0.5.













{kind=link}
| Embryology - 26 Feb 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Mall FP. and Meyer AW. Studies on abortuses: a survey of pathologic ova in the Carnegie Embryological Collection. (1921) Contrib. Embryol., Carnegie Inst. Wash. Publ. 275, 12: 1-364.
- In this historic 1921 pathology paper, figures and plates of abnormal embryos are not suitable for young students.
1921 Carnegie Collection - Abnormal: Preface | 1 Collection origin | 2 Care and utilization | 3 Classification | 4 Pathologic analysis | 5 Size | 6 Sex incidence | 7 Localized anomalies | 8 Hydatiform uterine | 9 Hydatiform tubal | Chapter 10 Alleged superfetation | 11 Ovarian Pregnancy | 12 Lysis and resorption | 13 Postmortem intrauterine | 14 Hofbauer cells | 15 Villi | 16 Villous nodules | 17 Syphilitic changes | 18 Aspects | Bibliography | Figures | Contribution No.56 | Contributions Series | Embryology History
| Historic Disclaimer - information about historic embryology pages |
|---|
|