Paper - The Falkiner ovum
| Embryology - 7 Aug 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Martin CP. and Falkiner N. Mcl. The Falkiner ovum. (1938) Amer. J Anat., 63: 251-271.
| Online Editor |
|---|

See also about this embryo: Falkiner NM. A description of a human ovum fifteen days old with special reference to the vascular arrangements and to the morphology of the trophoblast. (1932) J. Obst. Gynaecol. 39(3): 471-506. |
| Historic Disclaimer - information about historic embryology pages |
|---|
|
The Falkiner Ovum
C. P. Martin
Department of Anatomy, Mc:Gill Universlty, Montreal, Canada
And
Ninian Mci. Falkiner
Two Plates (Eight Figures)
Introduction
The Falkiner embryo has already been described by one of us (Falkiner, ’32) but as the previous communication was almost entirely concerned with the endometrium and the vascular relationships in the region surrounding the implantation site and did not deal at any length with the embryo proper, we feel justified in describing the latter in greater detail. Further, since the date of the previous communication one of us has had the privilege of inspecting the magnificent collection of embryos housed in the Carnegie Institution of Embryology at Baltimore, in which are several human specimens at approximately the same stage of development as our embryo and in addition a large number of macaque embryos of known age. Some of the latter are developmentally comparable with the Falkiner embryo and provide a provisional time scale for estimating its age. In the light of this examination of other specimens and also of further and critical study of the Falkiner embryo itself, we find that some of the views advanced in the previous communication require amendment, and this fact constitutes a further justification for the present communication. We would like here to emphasize our indebtedness to Dr. George Streeter of the Carnegie Institution for the very fine photographs of the specimen.
The history and clinical data of the specimen may be briefly recapitulated. The mother was aged 20 years, and had been married 3 years. She had one living child, then aged 2 years. Since her confinement she had had two abortions, the last in February, 1931. Since then she had menstruated once, beginning on March 15th and continuing for 5 days. She stated that her period had always been ‘.38 days. She was definite that coitus had occurred on March 23rd and on April 1st. She came to hospital in Marel’1, 1931, and was found to be suffering from anaemia and a sub—involuted, retroverted and retroflexed uterus. Her anaemia was treated for 1 month, and then on April 16, 1931, the uterus was curettved and the patient was operated on to rectify the displacement. Among’ the curettings was a large piece, with a small raised area on it, from the posterior wall of the uterus. This was recognized as a possible indication of an early pregnancy and the curettings were placed in normal saline. After about 2 hours they were handed over to Professor Gratenby of the Department of Zoology, Trinity College, Dublin, who fixed, sectioned and stained the specimen. The fixation was in Bouin’s fluid without acetic acid for about 18 hours. The specimen was then embedded in paraflin wax in the usual way and cut in sections 10 H thick. All the sections showing the embryo proper were stained with Ehrlich’s haematoxylin and orange Gr, but the sections at each extreme end—those at one end showing part of the body stalk and those at the other end the prolongation of the yolk sac— were stained with iron alum liaematoxylin and Biebrich scarlet.
figure 1 shows section 31 D, magnified 38 diameters, and gives a fairly good impression of the details of the specimen. The decidua is very well preserved and fixed, but unfortunately the embryo itself is not so perfect, as apparently the fixative did not sufficiently penetrate the chorionic cavity. The chorionic vesicle is a roughly spherical space rather flattened on the free surface toward the uterine cavity, which probably due to the post-operative handli11g and fixation of the specimen. The surface of the chorion abutting on the uterine wall is covered with well-developed villi, which are beginning to branch. These villi are markedly club-shaped, being constricted at their necks where they join the chorion wall and expanded at their peripheral ends. The branches arising from the villi also show this clubbing, so that a villus and its branches form a rather cactus-like figure, the main stem and each branch consisting of a narrow stalk and an expanded body. Careful examination of individual villi in successive sections demonstrated that this clubbing was an actual physical fact and the appearance was not due to the plane in which the villi were cut. The shape of these villi suggests that the mesoderm of which they consist is proliferating more rapidly at their distal than at their proximal end. This mesoderm apparently arises from the inner or cellular layer of the trophoblast and it is probable that the trophoblastic ectoderm covering the tip of a villus is more active in budding off the mesoderm than that covering the neck. Hence the villus becomes club-shaped. The mesoderm forming a villus therefore appears to originate in the trophoblast covering the villus itself and does not migrate from the chorionic mesoderm. In older specimens, when the villi have ceased to grow, or to grow so rapidly, they become more filiform and lose this club-shaped outline.
On the side of the chorion next to the cavity of the uterus the villi are very much shorter and cone—shaped with wide necks. They look as if, owing to the growth and stretching of the chorion, they were being taken into the chorionic cavity, as suggested by Fraser (’31).
Outside the villi is the cellular layer of the trophoblast and outside this is the plasmoditrophoblast which forms a sponge-like layer, thick on the side of the uterine wall, thin on that of the uterine cavity and enclosing many spaces, some of which are filled with extravasated maternal blood. This region of the ovum has been very fully described in the previous communication, and we have nothing to add to that description.
We estimate the size of the chorion to be as follows:
Maximum diameters between extreme outer limits of plasmoditrophoblast 2.05 X 1.80 X ? mm. External diameters, excluding villi and plasmoditrophoblast: 1.5 X 1.4 X ? mm.
The inside of the chorio11 is lined with a well-defined layer of extra-embryonic mesoderm, containing few spindle-shaped cells. Toward the center the cells become more condensed and form a definite layer bounding the extra-embryonic coelom and in the coelom itself are many loose strands of coagulum. The embryo proper lies near the uterine wall side of the chorion, to which it is rather eccentrically attached by the body stalk. The lower pole of the yolk sac also appears to have been attached by loose strands of mesoderm, now broken, to the opposite side of the chorion, i.e., the side next the utterine cavity. These connecting mesodermal strands are clearly visible in sections 33 A, B, C and D after the embryo itself has been passed.
The plane of sectioning is rather oblique. It makes a small angle of about 8° or 10° with the sagittal plane of the embryo and a rather larger angle with the horizontal plane of the embryonic plate. This last angle is difficult to estimate, for the embryonic plate appears to have been curved rather steeply in the transverse plane though only slightly curved in the sagittal plane. At first therefore the embryonic plate is cut very obliquely at about an angle of 45°, the cells of which it is composed have a rather polygonal outline, slightly elongated in the vertical direction, and the line of demarcation between the embryonic plate and the underlying tissue is very indefinite. Later the sections cut the embryonic plate at right angles and the cells form a well-marked columnar epithelium clearly demarcated from the underlying tissue.
The sections on slide 27 show only the body stalk. It is composed of mesoderm cells rather more condensed than in the lining layer of the chorion, and the stalk itself has a rather ill-defined boundary, especially on its ventral aspect, with some outlying strands of tissue and many indentations from the coelom entering into it. In section 28 A, a small plate of cells appears cephalad to the body stalk and on its ventral aspect. It is not connected to the bodystalk by any tissue but is adherent to it by a mass of coagulum. This plate is the Wall of the yolk sac. In section 28 B, the plate of cells is rather larger and appears to be connected with the body stalk by a thin strand of tissue composed of a double layer of cells. But in section 28 C, this connecting bridge of tissue has disappeared and the yolk sac is again completely detached from the body stalk, and in this section another rather longer and more ill-defined mass of cells appears dorsal to the yolk sac and separate from it, but directly continuous with the body stalk. This mass is the Wall of the amnion here cut through tangentially and hence displaying ill-defined outlines. In the next section, 28 D, the yolk sac Wall is still completely detached from the rest of the embryo. A large amniotic cavity has appeared but the Walls of the amnion are still rather ill-defined owing to the tangential sectioning, and on the ventral aspect of the body stalk at some distance from the yolk sac the end of the allantois has new appeared. It consists of columnar, darkly stained and closely packed cells surrounding a central cavity. In section 28 E, the yolk sac cavity appears for the first time, the amniotic cavity is now of considerable size and the dorsal part of the cavity is prolonged into the body stalk for a considerable distance. This prolongation probably represents an amniotic duct. The allantois is still visible but is not as distinct as in the previous section. It is Worth noting that the wall of the yolk sac is visible in four sections before the yolk sac cavity appears, but the amniotic Wall is only visible in one section before the cavity is visible. Obviously the yolk sac has a much thicker wall than the amnion.
The yolk sac is joined to the rest of the embryo by a broad band of loose tissue With a deep groove on both sides, that on the caudal side being the deeper. The dorsal prolongation of the amnion into the body stalk is still present, the allantois is visible but not very distinct, and in the floor of the amnion the extreme edge of the embryonic plate now appears. It is to be noted that the yolk sac is at a considerable distance from the embryonic plate and the two are connected by a broad band of tissue. In 29 A, the same features are observable but are more distinct and the embryonic plate much larger. This section is shown in figure 2. In 29 B, the yolk sac cavity is quite large; two deep indentations still constrict the bridge of tissue that connects it to the rest of the embryo, but the caudal and deeper of these is now bridged over at its mouth by an ill-defined band of tissue. The dorsal prolongation of the amnion is shorter and the allantois is very large and distinct. It makes a bend in this section and has been cut through at the bend. All these features are still present in 29 C, but the allantois is now cut twice and the indentation on the caudal side of the bridge of tissue connecting the yolk sac and the rest of the embryo has now disappeared; the indentation on the cephalic side is still present but is now very small. Section 29 D is shown in figure 3. The dorsal prolongation of the amnion into the body stalk has now disappeared. The embryonic plate is large but is somewhat damaged and torn; as already stated, in these sections it is cut rather obliquely and therefore has rather ill-defined edges, and the cells of which it is composed are polygonal in outline but somewhat elongated in a vertical direction. The cavity of the yolk sac that was visible in previous sections is now quite large and connected to the rest of the embryo by a mass of tissue in which the caudal and cephalic indentations previously noticed have completely disappeared. But now in this connecting mass of tissue a second cavity has appeared, cephalic and dorsal to the previously visible yolk sac cavity and close under the embryonic plate. This cavity, as will appear later, is also a part of the yolk sac and the two parts of the yolk sac cavity are separated by a broad band of loose tissue lined on either side by a definite and intact endothelium. It is noteworthy that on the outer surface of the yolk sac no groove corresponding to the position of this dividing septum is visible. The allantois has again been cut in two places: first, it is out almost longitudinally and can be seen as a clearly defined duct running almost directly dorsally from the caudal end of the ventral and caudal compartment of the yolk sac; and, secondly, it is cut transversely a short distance caudal to the dorsal end of the longitudinal portion. From this dorsal end of the longitudinal part a peculiar tongue-like process projects out toward the amnion, on the significance of which we can offer no suggestion. It may be a mere coincidence arising from the peculiar shape of a group of cells. The line of demarcation between the embryonic plate and the underlying tissue, though somewhat indistinct, can be made out and the plate is separated from the yolk sac cavity by a layer of loose and spongy tissue.
The next two sections, 29 E and F, present much the same picture. 29 F is shown in figure 4. In 29 E, the longitudinal portion of the allantois has been out twice, thus indicating the tortuous course it follows, but in 29 F it is only cut once. In the latter section the two compartments of the yolk sac have almost fused. The demarcation between the embryonic plate and the underlying tissue is quite visible. In both sections on the outer surface of the yolk sac a groove is present just ventral to the cephalic end of the embryonic plate, and i11 29 F, a similar groove is present just ventral to the caudal end of the plate ;the mouth of this latter groove is blocked by a small detached group of cells. The tissues lining the yolk sac cavity dorsal and ventral to these two grooves are quite different in character. Ventral to the grooves the yolk sac is lined by a thin, flattened and smooth endothelium; dorsal to the grooves, and therefore underlying the embryonic plate, it is lined by a thick, more cubical and much folded layer of tissue. These grooves, as suggested by Streeter, probably represent the earliest sign of the folding—off of the embryo and separate the gut endoderm dorsally from the yolk sac vesicle endoderm ventrally. The differentiation of the two types of endoderm is much more evident and distinct in macaque embryos.
In the next section, 30 A, these two grooves are very evident. The two compartments of the yolk sac cavity have now fused but are partially ‘separated by a broad tongue of tissue projecting inward from the cephalic side of the yolk sac. Ventral to the caudal half of the embryonic plate the line of demarcation between the plate and the underlying tissue has almost disappeared and a mass of darkly stained cells projects down to the roof of the yolk sac. This indicates that we are approaching the region of the primitive streak. The body stalk has now disappeared.
Section 30 B is shown in figure 5. The primitive streak is now evident, the caudal half of the embryonic plate passing down without a break to the roof of the yolk sac. The grooves on the outer surface of the yolk sac are present but the cephalic one is shallow. The tongue of tissue partially dividing the yolk sac cavity into two compartments is smaller, and opposite it on the outer surface of the yolk sac a peculiar circular vesicle has appeared. This vesicle persists into the next section and then disappears. Whatsits significance may be we cannot say. In section 30 C, though the embryonic plate is much damaged, the primitive streak is still evident, and the tongue of tissue partially dividing the yolk sac has almost completely disappeared. The grooves in the outside of the yolk sac are present but the one at the cephalic end is very small and shallow. Section 30 D was unfortunately destroyed.
Section 30 E is shown in figure 6. Although the embryonic plate is much damaged, it is obvious that it is now sectioned at approximately a right angle; it is composed of columnar epithelium clearly marked off from the underlying tissue, which here consists of a plate of very uniform thickness, composed of at least a double layer of rather closely packed cells. This plate roofs in the yolk sac and thins out rather abruptly at the cephalic end of the embryo. This condensed plate of tissue directly roofing the yolk sac is apparently the head process or notochordal plate, and the thinner stratum into which it passes at the cephalic end is the prechordal plate.
The grooves on the outer surface of the yolk sac have disappeared, and there is a considerable mass of tissue ventral to the caudal end of the plate. Section 30 F shows much the same picture, but here we have apparently cut through the very edge of the notochordal plate. A small detached group of cells has now appeared in the middle of the yolk sac. It persists and attains a considerable size in the next four sections and then disappears as abruptly as it appeared. Its significance is unknown.
In the next three sections, 31 A, B and C, the outlines of the embryonic" plate become more and more distinct as it is cut through more at right angles. Section 31 D is shown in figure 7. The amnion has rapidly diminished in size as we are now cutting through its edge. The amnion and embryonic plate together have become detached from the yolk sac and somewhat elevated above it. Beneath the embryonic plateis a layer of loose tissue, consisting of rather large scattered cells and connected with the lining of the yolk sac by some strands of tissue. This layer apparently is mesoderm and the notochordal plate has disappeared. On the outside of the yolk sac there are still suggestions of grooves ventral to the caudal and cephalic ends of the embryonic plate, but the difference in character of the endodermal lining of the yolk sac dorsal and ventral to these points is still very evident. At the lower or ventral pole of the yolk sac, however, the endoderm is more condensed and cubical, and resembles that under the embryonic plate more than that on the side walls of the yolk sac.
In the next three sections, 31 D, E and F, the same picture is obtained except that the embryonic plate and amnion rapidly diminish in size. In the next section, 32 A, the embryonic plate appears for the last time, and in the next one again, 32 B, the amnion makes its last appearance. In four more sections, 32 C, D, E and F, the yolk sac is intact and diminishes slightly in size, in section 33 A, it is broken on one side, and in further sections only scattered cells appear. They are especially well marked in sections 33 A, B, O and D, and apparently represent the mesodermal strands which connected the ventral pole of the yolk sac with the opposite wall of the chorion.
Having thus briefly described the whole embryo,we can now discuss the characteristics and peculiar features of each of the different parts of the specimen.
The Age and Dimensions of the Embryo
In the previous communication the age of the embryo was estimated at 15 days. We are now inclined to modify this opinion, and think it is probably a little older. Comparing it with macaque material of known age, We find that, as regards the state of development of the embryonic plate and primitive streak, it agrees very closely with macaque embryos 17 days old, but in the latter the villi are rather better developed and more branched. In the Falkiner embryo the yolk sac is larger than the amnion, in macaque embryos the amnion is always larger than the yolk sac, so this feature is of no use for comparison. But in the degree to which the roof of the amnion has become detached from the chorion, the macaque embryos of 17 days are less advanced than our specimen. Using macaque material as a time scale, therefore, our embryo appears to be about 17 days old. But of course such a conclusion is based on the assumption that the early stages of human and macaque embryos are strictly comparable. Taking all the features together, the 17-day macaque agrees more closely with our specimen than does the 15-day macaque.
On comparing the Falkiner embryo with other human specimens, we find that it is slightly more advanced than Florian’s Bi.1, though this is the specimen with which it agrees most closely, the only point of divergence being that in our specimen a head process is present, while in Florian’s specimen it is not definitely identifiable. Florian estimates the age of his embryo as 15 days. On the other hand, our specimen is definitely less advanced than the Heuzer embryo, estimated as 17 days old, and the Mateer embryo, estimated as 17 to 18 days old.
Summing up all the evidence we find:
The clinical facts suggest an age of about 15 days.
Comparison with macaque material suggests an age of 17 days.
Comparison with other human embryos suggests an age of a little more than 15 and less than 17 days, that is, of course, assuming that the age of these Florian Bi.1, Mateer and Heuzer embryos has been correctly estimated.
The actual dimensions of the Falkiner embryo are estimated as follows:
Length Breadth Height
Embryonic plate 0.15 x 0.29
Amniotic cavity 0.16 x 0.35 x 0.1
Yolk sac 0.29 x ‘'3 x 0.26
The amniotic cavity and yolk sac height was calculated from the midpoint of the embryonic plate and their length and breadth calculated in a plane parallel to the embryonic plate. As both cavities were torn these dimensions are only approximate.
The Amnion
The amnion is composed of a layer of very thin flattened epithelium, continuous with the margin of the embryonic plate and covered on the outside with a thin layer of mesoderm, which in some places has disappeared (fig. 3). The mesodermal covering appears to be directly in contact with amniotic ectoderm and no clear space intervenes between them. The roof of the amnion is completely separated from the wall of the chorion, the extra-embryonic coelom extending between the two structures, but the amnion covers the upper surface of the body stalk, and from its dorsal part in this region a broad tube-like prolongation extends for some distance into the body stalk. This we interpret as the representative of the amniotic duct so clearly seen in macaque embryos. Inside the amnion are a few scattered granules of coagulum. The amniotic cavity slightly overrides the edges of the embryonic plate.
The Embryonic Plate
This is composed of a thick plate of densely packed pseudostratified columnar epithelium, which thins out rather abruptly toward its margins where it passes into the amniotic ectoderm. In the sagittal plane it is slightly curved with a dorsal convexity, but just at its margin the dorsal convexity is replaced by a slight dorsal concavity. In the transverse plane it is more steeply curved, with a marked dorsal convexity. It is separated from the underlying tissue by a clear space except in the region of the primitive streak.
The Yolk Sac
The yolk sac is considerably larger than the amnion and its walls consist of a layer of endoderm covered on the outside with a layer of mesoderm. The mesodermal covering is thicker and much more definite than the corresponding covering of the amnion; it is also separated from the endoderm by a comparatively wide and clear space. In many places this mesodermal covering becomes thickened into small islands of tissue, probably the beginning of blood islands, though no identifiable vascular spaces were seen. As already stated, the lower pole of the yolk sac appears to have been attached to the opposite wall of the chorion by strands of this mesoderm.
The caudal end of the yolk sac is partially divided into two compartments by an oblique septum, which commences under the caudal end of the embryo and runs ventrally and cephalically round the left side of the yolk sac, thus dividing it into a dorsal or cephalic and a ventral or caudal compartment. It is quite evident that the dividing septum is a physical reality and is not in any way due to artificial deformation of the structure. The allantois comes off the ventral compartment of the yolk sac.
In some sections grooves on the outside of the yolk sac occur just ventral to the caudal and cephalic ends of the embryonic plate, and the endoderm in the dorsal wall of the yolk sac between these grooves and underlying the embryonic plate differs in character from the remainder of the endoderm. In other sections, though the grooves are not visible, the difference in the endoderm is still distinct. The endoderm lining the remainder of the yolk sac is a thin, fiat and smooth epithelium, but in this dorsal zone of the yolk sac it becomes more cubical in type and is much folded. This probably indicates the early differentiation of the endoderm into gut endoderm, which will be actually incorporated into the body of the embryo when the embryonic plate folds off and will form the lining layer of the alimentary tract, and yolk vesicle endoderm, which will remain in the umbilical cord and form the yolk stalk and vesicle. At the extreme lower or ventral pole of the yolk sac the endoderm also assumes a cubical character.
The Extra-Embryonic Mesoderm and Coelom
The extra-embryonic mesoderm forms a fairly thick layer lining the chorion. This layer contains few small spindle-shaped cells but on its inner aspect the cells are more condensed and form a sort of epithelium lining the coelom. In the body stalk the cells become more numerous and condensed but the surface of the stalk has an irregular and vesicular outline. A very thin layer of mesoderm covers the amnion and lies directly in contact with the ectoderm, no space intervening. The yolk sac is covered by a more definite layer of mesoderm with many larger cells and little islands of tissue, and this layer is separated from the endoderm by a relatively wide clear space. The impression obtained on examination of the sections is that the mesoderm covering the yolk sac is actively proliferating and growing, while that covering the amnion is atrophying. Strands of mesoderm connect the ventral pole of the yolk sac with the chorion. Ventral to the caudal and cephalic ends of the embryonic plate, in the angles between the amnion and the yolk sac, there is a considerable thickening of the mesoderm, especially at the caudal end. In this region the extra-embryonic mesoderm appears to pass into the embryonic mesoderm, or at least no clear line of division between them can be made out. The mesoderm also passes into the septum which partially divides the yolk sac.
The Primitive Streak, Notochordal Plate and Embryonic Mesoderm
The primitive streak consists of a great thickening of the dense and darkly stained ectoderm of the embryonic plate, which extends downward into the roof of the yolk sac. It occupies about one-third of the length of the plate, and as it is out rather obliquely it appears to be rather thicker than is actually the case. It occurs in three sections only, so its width must have been considerably less than its length. In the region of the primitive streak therefore the yolk sac is roofed by a mass of undifferentiated tissue which passes without a break into the ectoderm of the embryonic plate. In the caudal direction the primitive streak appears to pass rather suddenly into the thickening of the extra-embryonic mesoderm situated in the caudal angle between the amnion and yolk sac; in the cephalic direction it passes into the three germ layers and notochordal plate.
The notochordal plate is a very uniform rod of cells, extending cephalically from the primitive streak and lying directly in the roof of the yolk sac. The rod consists of at least two layers of closely packed cells, but no definite lumen can be seen. At the cephalic end of the embryo it passes directly into a thinner and more irregular layer of cells lying also directly in the roof of the yolk sac. This probably is the prechordal plate.
The embryonic mesoderm consists of a loose undulating strand of cells extending forward from the primitive streak on either side of the notochordal plate. In the early sections, where the embryonic plate is cut very obliquely, this tissue is difficult to see, but in the later ones where the plate is cut at right angles it is very clearly displayed. It is separated by a clear space from the embryonic plate, but is connected to the endodermal roof of the yolk sac by several chains of cells. In the cephalic direction it thins out but there does not appear to be a definite break between it and the extra«embryonic mesoderm.
The Allantois
The allantois is very long and tortuous though how much, if any, of this tortuosity may be due to post-operative handling is difficult to determine. It arises from the caudal and ventral compartment of the yolk sac, runs dorsally in the caudal wall of the yolk sac, and enters the body stalk. In this part of its course it undulates slightly. It then turns abruptly caudally at about a right a.ngle and runs backward in the body stalk in a curve with its convexity to the left of the embryo. Where it makes the sharp right-angled turn, a peculiar coneshaped process grows out from it toward the amnion (fig. 3). What this cone-shaped process may be is difficult to say. In the sections where the allantois is shown, the embryonic plate and amniotic ectoderm are cut very obliquely and relationships are difficult to determine, but at no time does the allantois appear to be near the primitive streak and a cloacal membrane, if present, cannot be definitely identified.
Two wax-plate reconstructions of the embryo were made, one in 1932, and the second during the last few months. A. medial section of the embryo, based on the later of these reconstructions, is shown in figure 8.
Discussion of the Special Features of the Specimen
This embryo displays two special peculiarities: first, the partial division of the yolk sac into two compartments; and, secondly, the origin of the allantois low down in the caudal side of the yolk sac and its long and tortuous course. As regards the partial division of the yolk sac, there is no doubt that this feature is not due to any deformation or damage that may have occurred. The septum which causes the division is a structure of considerable bulk and both it and the adjoining wall of the yolk sac are intact except for very slight damage to some cells on their surfaces. This partial division of the yolk sac has not apparently been previously observed in human embryos but it is interesting to record that it sometimes occurs in macaque specimens. In the Carnegie Institution collection there is an embryo (no. G. 571, macaque, aged 15 days), which shows an exactly similar feature, the yolk sac being partly divided by a definite and intact septum, two cells thick.[1]
The yolk sac apparently originates as a cavity arising in the lower pole of the inner cell mass. It is probable that such a cavity at its beginning is very irregular in outline, or even may arise as two or more separate cavities which subsequently run together. The evidence furnished by the Falkiner embryo and macaque embryo no. G. 571 suggests that toward the body stalk the yolk sac enlarges by the formation of separate bays or compartments, and that subsequently these outgrowths are taken up into the main cavity. The caudal end of the yolk sac consequently has an irregular and vesicular outline and is sometimes divided by septa into separate bays. The allantois itself may be regarded as a special instance of the tendency for this part of the yolk sac to form evaginations burrowing into the substance of the body stalk, and possibly much of the original allantois may later on be taken up into the cavity of the yolk sac. If the process of taking up the evaginations of the yolk sac and the proximal end of the allantois into the yolk sac cavity was delayed, and there is probably considerable individual variation in this process, it seems to us that we would have a state of affairs such as is presented by our specimen. Had the taking up process gone a little further, the septum partly dividing this region of the yolk sac would l1ave disappeared and the part of the allantois situated in the caudal wall of the yolk sac would have been incorporated in the yolk sac cavity. We would then have a single yolk sac with the allantois arising from its dorsal and caudal cor11er, such as has been found in so many other specimens. It is interesting to note that in the Ramsey Yale embryo in the Carnegie Institution, though the yolk sac is a single undivided cavity, the allantois arises rather low down in the caudal Wall of the yolk sac, as in our specimen (Ramsey, ’38). The very tortuous course followed by the allantois in the Falkiner embryo is probably also due to a delay in the taking up of the proximal end of this structure. The rapid growth and expansion involved in the taking up of the proximal end of the allantois into the yolk sac would also tend to straighten out the distal end of the allantois. In our View therefore the peculiar features of the Falkiner embryo can all be accounted for on the assumption that the yolk sac enlarges toward the body stalk by the formation of bays or vesicles, of which the allantois itself is only a very special instance, and that these bays are subsequently taken up into the yolk sac itself, but that in our specimen there has been for some unknown reason a slight retardation in the taking-up process.
Footnotes
- ↑ In slide 3 of embryo no. 0.571 (the number on the slide being placed above and to the left on the microscope stage, the rows of sections numbered from above downward, and the sections in each row lettered from left to right) section A of row 7 shows a single yolk sac but the extreme edge of the septum is beginning to appear; section D, of the same row, shows the septum at its best development and the yolk sac is completely divided into a small dorsal or caudal, and a large ventral or cephalic compartment; and in section J, of the same row, the small dorsal compartment ends. A structure somewhat resembling an allantois is situated to one side of the smaller compartment, but the allantois arises late in the macaque and the appearance seen in this slide is probably misleading in this respect.
Literature Cited
Falkiner NM. A description of a human ovum fifteen days old with special reference to the vascular arrangements and to the morphology of the trophoblast. (1932) J. Obst. Gynaecol. 39(3): 471-506.
FALKINER, N. MCl. 1932 A description of a human ovum fifteen days old with special reference to the vascular arrangements and to the morphology of the trophoblast. J. Obst. and Gyn. Brit. Empire, vol. 39, pp. 471-506.
FRASER, J. E. 1931 A Manual of Embryology. Bailliere, Tindall & Cox, London.
Ramsey EM. The Yale embryo. (1938) Contrib. Embryol., Carnegie Inst. Wash. Publ. 496, 27: 67-84.
Explanation of Figures
Plate 1

1 Section 31 D. X 38.
2 Section 29 A. X 150. The large amniotic cavity lies dorsal and it is prolonged into the body stalk as the amniotic duct. The small cavity of the yolk sac is separated from the amnion by a considerable interval filled with loose tissue with deep grooves on either side. In the floor of the amniotic cavity the extreme edge of the embryonic p-late has been sectioned very obliquely. The allantois, though indistinct, can be seen in the body stalk.
3 Section 29 D. X 150. Two parts of the yolk sac cavity separated by a broad band of tissue are visible in this section. One small part lies fairly close under the embryonic plate; the other and larger lies more ventrally. This last is the compartment that was visible in previous sections. The allantois is cut in two places: first it is seen as a long duct running dorsally from the caudal end of the ventral compartment of the yolk sac, and secondly it is again c.ut slightly caudal to the dorsal end of the duct-like part. From the dorsal end of the longitudinal part a wedge-shaped mass of cells projects toward the amniotic cavity.
4 Section 29 F. X 150. The two compartments of the yolk sac are fusing in this section. The allantois is still visible in the body stalk. The amnion has been torn. Two grooves constrict the outside of the yolk sac, and the endoderm in the roof of the sac between these grooves is irregular and much folded, while that below the grooves forms an even regular layer.
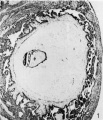
Fig. 1
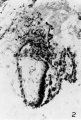
Fig. 2
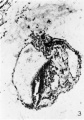
Fig. 3
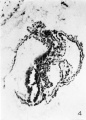
Fig. 4




Plate 2

5 Section 30 B. X150. This section passes through the primitive streak, which is seen at the caudal half of the embryonic plate. The amnion and embryonic plate have been torn. A t0ng'ue of tissue still partially divides the yolk sac.
6 Section 30 E. X 150. The amnion and embryonic plate are much damaged but the latter is now sectioned at approximately 90° and is seen to be composed of a densely packed columnar epithelium. The roof of the yolk sac is formed by an even rod of tissue, two or more cells in thickness but thinning out at the cephalic end. This is interpreted as the head process or notochordal plate.
7 Section 3.1. D. X 150. The embryonic plate is now cut through at 90°. The amnion has rapidly diminislled in size. A loose strand of mesoderm intervenes between the embryonic plate and the underlying endoderrn. The significance of the detached clump of cells in the middle of the yolk see is unknown.
8 Reconstruction of medial plane of embryo.
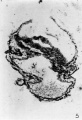
Fig. 5
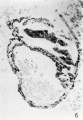
Fig. 6

Fig. 7
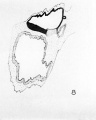
Fig. 8




Cite this page: Hill, M.A. (2026, August 7) Embryology Paper - The Falkiner ovum. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_The_Falkiner_ovum
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G