Book - Human Embryology and Morphology 20
| Embryology - 17 Apr 2026 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Keith A. Human Embryology and Morphology. (1902) London: Edward Arnold.
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
- Links: Limb Development
Chapter XX. The Limbs
The Limbs. — The limbs begin to appear at the end of the 3rd week. A slight elevation or ridge is then seen to run along the dorsal border of the somatopleure, at some distance from the row of primitive segments formed in the paraxial mesoblast (Fig. 233). The limb buds spring from this ridge as fiat processes with an upper, dorsal, or extensor surface, and a lower, ventral or flexor surface. The two borders are anterior or cephalic and posterior or caudal (Fig. 233). It is generally held that the lateral ridge, of which the limb buds are specialized parts, represents a continuous row of lateral fins. If this view is right, then the fore and hind limbs represent highly specialized fin-rays.

Fig. 233. Lateral view of a human embryo at the 28th day, showing the Limb Buds, Lateral Edges, and Primitive Segments.
A section shows each bud to be composed of undifferentiated mesoblast, with a covering of epiblast (Fig. 111 A and Fig. 237). It represents in structure a process of the undifferentiated somatopleure or body- wall. Processes grow into the limb bud from the muscle plate of each segment which is situated opposite the origin of the bud. Each corresponding segment of the spinal cord also sends to the limb bud a nerve process. At least seven body segments contribute to the formation of each limb.
Changes in External Conformation
In the 4th week (Fig. 234) the limb buds are unsegmented ; in the 5th a constriction marks the hand off; the position of the elbow being indicated in the same week. In the 7th week the fingers appear as thickenings in the webbed hand, the middle digit being indicated first. They become free at the end of the second month ; occasionally they remain attached, the child being born with its fingers in a syndactylous condition. The shoulder remains buried in the body-wall ; the skeletal structures of the shoulder are the first to appear. The corresponding parts of the lower limb appear in the same order as in the arm.

Fig. 234. Development of the Upper Limb. (After Wilhelm His (1831-1904))

Fig. 235. Development of the Lower Limb.
The Internal Differentiation of Tissues begins at the basal part of the limb and spreads towards the fingers, the terminal phalanges being the last of the skeletal parts to become differentiated. The mesoblast becomes condensed in the axis of the bud and forms the fibrous basis of the limb bones. The skeletal basis of mesoblast is continuous, but where joints are to be formed there occur opener formations in the arrangement of the cells. In the 5th week the fibrous basis of the bones begins to chondrify, and this is soon followed by the appearance of centres of ossification in the shafts of the long bones (7th week). The mesoblast between the chondrified bases of the bones opens out into a cavity and forms the synovial membranes of the joints. The joints are all formed before the 3rd month is well begun. During the 6th to the 8th week the mesoblast differentiates into muscles, bones, vessels and the sheaths of nerves ; the tissue left over, not included in these structures, forms their sheaths, and the fasciae and connective tissue of the limb. The processes of the nerve cells to form the nerves, and of the muscle plates to form the muscles, grow in very early (see Fig. 237).

Fig. 236. The Corresponding Points {A, B, C, and D) in the Ilium and Scapula.
Torsion and Rotation of the Limbs
As the limbs are developed, the extensor surfaces of the knee and elbow are directed upwards. If the body of an adult were placed in the prone position, it would be necessary, in order to restore the limbs to their embryonic position, (1) to draw them out at right angles to the axis of the body, (2) to rotate the leg outwards so that the extensor surface of the knee is directed upwards, with the great toe in front and the little toe behind. (3) The arm, on the other hand, would require to be rotated inwards to bring the elbow (extensor surface) into the dorsal position. The rotation which brings the embryonic limbs into the adult position occurs at the junction of the limb girdle with the trunk.
Rotation at the Limb Girdle
To understand the extent of this rotation it is best to compare the scapula and ilium and pick out their corresponding points. The extensors of the thigh and arm may be taken as guides. The long head of the triceps and rectus femoris of the quadriceps manifestly correspond ; their points of origin — the anterior border of the ilium and axillary border of the scapula — may be regarded as homologous points. The other corresponding points are shown in Fig. 236. The sacral articular surface of the ilium corresponds to part of the supra-spinous fossa. To restore the limb girdles to their primitive and corresponding positions, the scapula has to be rotated so that its axillary or posterior border comes to occupy the position of its spme, while the ilium has to be placed at right angles to the spine and its anterior border rotated outwards until it occupies a position corresponding to the axillary border of the scapula.

Fig. 237. Section of the Arm Bud of a human embryo at the end of the 4th week. (Alex. Low.)
There is a manifestly spiral twist in the humerus, but it is doubtful if this be in any way due to the torsion which the limb undergoes.
Nerve Supply of the Limbs
The Arm
It is important to note that the limb buds arise from the ventro-lateral aspect of the trunk (Fig. 237) at the junction of the somatopleure with the paraxial mesoblast. Therefore the nerves of the limbs are the nerves of the ventro-lateral zone— the lateral cutaneous branches of the segmental nerves (Fig. 238). The muscles are derived from the ventro-lateral sheet, which gives rise to all the muscles of the body- wall (Fig. 238). As soon as the limb buds appear, bundles of fibres from the anterior and posterior nerve roots of the corresponding body segments enter them and keep time with their growth. The limb nerves are at first so large in comparison with the size of the limb bud that they are crushed together and form a plexus. As they enter the bud, the nerves encounter the condensed skeletal inesoblast at its base and divide into a dorsal or extensor set and a ventral or flexor set (Figs. 237 and 238).

Fig. 238. Schematic Section showing the Origin and Arrangement of the Muscles and Nerves of the Limbs. (After Kollmann.)
The nerve supply assists to indicate the body segments from which the arm is developed (Fig. 239). The 4th cervical is the most anterior, the 3rd dorsal, sometimes it is the 2nd, is the most posterior segment. Hence the arm is produced from seven, or more commonly eight segments in all. Each segment contributes from its nerve, its muscle plate and probably also its artery. The typical distribution of a segmental nerve to the limb bud is shown diagrammatically in Fig. 238. Each segmental nerve, as is the case with the typical lateral cutaneous nerves, divides into a dorsal division for the extensor muscles (musculo-spiral, posterior circumflex, supra-scapular, etc.) and ventral for the flexor muscles (median, ulnar, musculocutaneous) (Fig. 238).

Fig. 239. The Distribution of the Posterior Roots of the Spinal Nerves on the Plexor Aspect of the Arm.

Fig. 240. Diagram to show the typical Manner in which the Posterior Nerve Roots are distributed in the Lower Limb (based on Sherrington's researches into the sensory distribution of the limb nerves of apes).
Clinical and experimental research has shown that each of the seven or eight segments contributes to the cutaneous supply of the limb. The classical investigations of Sherrington in the segmental distribution of the sensory nerves in the limbs of apes, showed that they are arranged in a definite and orderly manner (Fig. 240). The sensory distribution of the spinal nerves in the human arm is shown diagrammatically in Fig. 239. The distribution of the motor nerves of each segment is the following :
- 5th Cervical — To extensor muscles which raise the shoulder, extend the arm, fore-arm and proximal phalanges.
- 6th Cervical — To adductors and extensors of the arm, flexors of the thumb.
- 7th Cervical — Rotators outwards of the shoulder and flexors of the middle phalanges.
- 8th Cervical — Flexors of the fingers and extensors of the carpus.
- 1st Dorsal — Flexors of fingers, the interossei, adductors of thumb.
It will be thus seen that in the motor segments, not muscles but actions are represented ; hence the formation of the plexuses that each nerve may contain functional elements derived from several segments. Each muscle, it must be remembered, is supplied from two, three or more segments of the cord.
Only three anomalous points in the arrangement of nerves in the upper limb require attention : (1) The segments which supply nerves for the arm are nearly constant. The extent to which the 4th cervical and 3rd dorsal contribute varies ; the degree of variation is markedly less than in the lower limb. (2) A part of the musculocutaneous nerve frequently joins the median below the insertion of the coraco-brachialis ; this communication is frequently seen in lower primates ; its meaning is not known. (3) A communication between median and ulnar in the forearm is also common and is seen constantly in some primates. The communicating branch passes with the deep branch of the ulnar nerve to the palm.

Fig. 241. Plexor Aspect of the Lower Limb showing the Sensory Distribution of the Segmental or Spinal nerves.
Nerve Supply of the Lower Limb
Usually ten segments contribute to the nerve supply of the lower limb — the 12 th dorsal to the 4th sacral (Fig. 241). The sensory nerves are derived from these segments ; the motor nerves begin at the 1st lumbar segment and end at the 3rd sacral. There is a considerable variation in the number of body segments or vertebrae to which the lower limb is attached; usually it is the 25th vertebra which becomes the 1st sacral, but it may be the 26 th or 24th (page 144). Of these three forms the first is the normal type (25th); the second the post-fixed type (26th), the third the pre-fixed type (24th). There is even a greater variation in the segments which contribute nerves to the limb ; the normal motor segments are the 1st lumbar to the 3rd sacral ; in the post-fixed type (more common than the next) the motor segments commence at the 2nd lumbar and cease at the 4th sacral ; in the pre-fixed type the motor segments commence at the 12th dorsal and end at the 2nd sacral. The spinal nerve which bifurcates and joins both lumbar and sacral plexuses is known as the nervus furcalis. In the normal type it is the 4th lumbar; in the pre-fixed type it is the 3rd lumbar ; in the post-fixed type the 5th lumbar.
The nervus bigeminus, normally the 4th sacral, may also vary in a corresponding manner.
The nerves to the extensor surface of the lower limb, the anterior crural, external popliteal, etc., represent the dorsal divisions of lateral cutaneous nerves (Fig. 238). The nerves to the adductor and flexor aspects, the obturator and internal popliteal, represent the ventral divisions. In a considerable number of individuals, the dorsal division (external popliteal) and ventral (internal popliteal) of the great sciatic separate in the pelvis, the external popliteal perforating the pyriformis.
The muscles of the lower extremity are supplied in the normal type from the following segments :
Motor Segments. —
- 1st Lumbar. Psoas and Iliacus.
- 2nd Lumbar. Psoas and iliacus, sartorius, pectineus, adductor brevis.
- 3rd Lumbar. Adductors, quadriceps, obturator externus.
- 4th Lumbar. Adductors, quadriceps, obturator externus, also external rotators of hip, gluteus maximus, hamstrings.
- 5th Lumbar. External rotators of hip, gluteus maximus, gluteus medius and minimus. Muscles of leg and foot.
- 1st Sacral. External rotators of hip, gluteus maximus, gluteus medius and minimus. Muscles of leg and foot.
- 2nd Sacral. External rotators of hip, gluteus medius and minimus, flexor hallucis, extensors of foot and toes.
- 3rd Sacral. Pyriformis, muscles of calf and sole.
- 4th Sacral. Muscles of perineum.
It will be remembered that the perineal region is developed behind the limb buds of the lower extremities ; hence its nerve supply from the most posterior nerve segments (3rd and 4th sacral).
Sherrington found that the posterior roots of the limb nerves were distributed a regular and simple manner in apes. His results are applied to the lower limb of a human foetus in Fig. 240. The actual distribution in man, which has been partially worked out by clinicians, varies considerably from what might be expected from Sherrington's results (compare Figs. 240 and 241).
In the human leg and foot there is a tendency for the nerve fibres destined for the outer digits to proceed in the external saphenous nerve instead of by the musculo-cutaneous. The external saphenous nerve may supply the 4th and 5th digits, in a manner similar to the ulnar nerve in the hand ; more frequently it is confined to the outer side of the 5th digit.
Pelvic and Shoulder Girdles
In the basal part of each limb bud a cartilaginous arch is developed. It consists of a dorsal and ventral part, the joint cavity for the articulation of the limb being situated at the junction of the two parts. Fishes retain this simple primitive form of girdle.
The Pelvic Girdle has undergone less modification from the primitive type (Fig. 242) than the shoulder girdle. The primitive type of pelvic girdle, such as is seen in the crocodile or lizard, and of which the mammalian type is a derivative, is shown diagrammatically in Fig. 242. For comparison the human girdle in a 5th week foetus is shown in Fig. 243.

Fig. 242. Diagram of the Pelvic Girdle of a Lizard.
The dorsal element consists of the ilium ; it is attached by ligaments to the costal process of one or more sacral vertebrae. In the ventral portion of the cartilaginous arch are developed two elements, the pubes and ischium, both of which take part in the formation of the acetabulum (Fig. 242). Both reach the ventral median line in which cartilage is developed.

Fig. 243. The Pelvic Girdle of a Human Foetus at the 5th week. (After Kollmann )
In man the following changes may be noted: (1) The costal processes of the sacral vertebrae (2 \ usually) have fused together to form the lateral sacral mass, with these the ilium articulates ; (2) the vertebral border (crest) has become enormously elongated and gives attachment to abdominal muscles, cutting off the fibres of insertion of the external oblique which form the chief part of Poupart's ligament ; (3) the ischium does not reach the ventral line. In most birds, neither ischium nor pubes reach the ventral line. The pubes fail to meet in cases of ectopia vesicae. The symphysis pubis is formed in the ventral line. The cotyloid bone — os acetabuli — is formed in the Y-shaped cartilage between the three elements. It ossifies in the 13th year. Professor Howes has pointed out that it is this ossification which forms the pubic part of the acetabulum, and that it is really part of the pubes.
Shoulder Girdle
The duckbill (ornithorynchus, platypus) shows the most generalized type of mammalian shoulder girdle ; it resembles closely the primitive reptilian type ; from such a form the various types of mammalian shoulder girdle were probably evolved.
The dorsal part of the arch consists of (1) scapula, (2) suprascapula (Fig. 244). The supra-scapula is represented in man by the cartilage along the vertebral border ; it ossifies in the early years of manhood. The supra-spinous part of the scapula appears first in higher mammals ; it is produced late in the development of the scapula (in the 3rd month of foetal life) by the upgrowth of the supra-spinous blade of the scapula ; it is not represented in the pelvic girdle. The spine of the scapula is represented in the pelvic girdle by the ilio-pectineal line of the ilium ; but there is no pelvic representative of the acromion process. The dorsal segment of the pelvic girdle becomes fixed to the costal processes the corresponding part of the scapula remains free.

Fig. 244. The Shoulder Girdle of Ornithorynchus (platypus).
In the typical reptilian shoulder girdle, as in the pelvic (Fig. 242), two elements are formed in the ventral part of the arch — a posterior part — the coracoid, corresponding to the ischium, and an anterior — the pre-coracoid, corresponding to the pubes. Both elements reach the ventral median line in which the presternum is developed. In Ornithorynehus the coracoid element is represented by two bones — the coracoid and epi-coracoid. The coracoid helps to form the glenoid cavity at its dorsal extremity ; its ventral articulates with the presternum. In man and all higher mammals, in which mobility of the fore limb is of advantage for speed and free movement, the coracoid element is much reduced. It forms merely a process on the scapula, which it joins in man about the 15th year. It still enters into the formation of the glenoid cavity, the articular part (supra-glenoid) having a separate centre of ossification which appears in the 12 th year. It is possible that the costo-coracoid ligament may be derived from the ventral part of the coracoid element — the part which articulates with the sternum in the duckbill. The pre-coracoid in the shoulder girdle of the lizard corresponds to the pubic element in the pelvis. The pre-coracoid, which, like all the primitive elements of the pelvic and shoulder girdles is formed in cartilage, has been partly or entirely replaced in all mammals by the development over it of the clavicle, a dermal or membrane-formed bone, the first of all the bones to ossify. There is thus no true representative of the clavicle in the pelvis. The inter-clavicle so strongly developed in the ornithorynchus and in the "merry-thought" of the fowl is also a dermal bone. It is represented in man by the interclavicular ligament. It is highly probable that the clavicle does not completely replace the pre-coracoid.
The epi-coracoid of ornithorynchus probably represents part of this element (Fig. 244). It is also represented in the shoulder girdle of man by three structures all of which appear in cartilage (see Fig. 245).
- By the cartilaginous inner end of the clavicle
- by the inter-articular cartilage
- by the supra-sternal bones.

Fig. 245. The Parts in the Shoulder Girdle of a human foetus which correspond with those of Ornithorynehus.
In order to give greater mobility and speed to some four-footed mammals, the clavicle has been reduced to a ligamentous band, except at its extremities (rabbit, dog, etc.). In climbing animals, and those in which the power of grasping or embracing is highly developed, the clavicles are fully developed.
The acromion process is ossified from several centres which appear in the years of adolescence ; the epiphysis so formed may be united to the spine by fibrous tissue only. This occurs in over 8% of subjects (Symington), and may be mistaken for a fracture of the process. The coraco-clavicular ligaments may be derived from the pre-coracoid element.
The Hand and Foot
The hand and foot of man, as is the case in all primates, retain the primitive arrangement of elements much more closely than do most other mammalian orders. The primitive type of hand or foot, out of which the various forms found in mammals have been modified, are seen in such reptiles as the lizard or tortoise (Fig. 246). In the hand of man the same bones are to be seen as in the tortoise (Fig. 246), and the same arrangement with some exceptions. The elements in the foot of a typical lizard resemble closely the arrangement seen in its hand ; the same elements are present even in the highly modified human foot. The hand and foot bones have undergone great specialization in most mammals. In the evolution of the horse, for instance, one lateral digit after another has become vestigial, leaving the central digit enormously enlarged and specialized to form the lower part of the extremities. In ruminants the 3rd and 4th digits have become specialized ; the rest of the digits have become reduced until only traces of them are left ; in rodents the hallux is vestigial. The hallux and pollux are the mammalian digits most liable to undergo retrogression. In man, on the other hand, the hallux and pollux find their greatest development.
Comparison of the Tarsus and Carpus
Both are the derivatives of such a typical form as is shown in Fig. 246.

Fig. 246. The Carpal Bones of a Tortoise.

Fig. 247. The Os Trigenum and Bones of the Tarsus.
In the typical tarsus or carpus there occur the following bones :
- Radiale or Tibiale forms the scaphoid in the hand and astragalus in the foot.
- Intermedium forms the semilunar in the hand ; in the foot it is much reduced and usually unites with the astragalus to form the external tubercle of that bone. It may remain separate and form the os trigonum (Fig. 247).
- Ulnare becomes the cuneiform in the hand ; the os calcis in the foot. During the fibrous and early cartilaginous stages in the development of the human tarsus, the os calcis is in contact with the fibula. In the hand the ulnare and intermedium are bound by fibrous bands to the ulna (Fig. 246); these bands assist to form the triangular fibro-cartilage.
- Carpale or Tarsale I. becomes the trapezium in the hand, the internal cuneiform in the foot. In the prehensile foot of apes, the hallucial articular surface is directed inwards for the moveable big toe. This is also the case during the foetal development of the human foot (Leboucq). At no period of development is the hallux of man directed inwards and separated from the other toes. In man the great toe resumes a primitive position and its metatarsal lies parallel with those of the metatarsal series.
- Carpale or Tarsale II. forms the trapezoid in the hand, the middle cuneiform in the foot.
- Carpale or Tarsale III. forms the os magnum in the hand, the external cuneiform in the foot.
- Carpale or Tarsale IV. and V. have united in both hand and foot to form the unciform and cuboid. This union is seen in mammals generally. In the cat and carnivores the scaphoid and semilunar unite together.
The Os Centrale is situated between the first and second rows of the carpal or tarsal bones (Fig. 246). In the foot it forms the scaphoid — a bone which plays an important part in the formation of the plantar arch — but is yet remarkably late in beginning to ossify, viz., about the 3rd year. It appears in the membranous stage of the human carpus, but at the end of the 2nd month it has joined the dorsal and distal aspect of the scaphoid of the hand. It may be occasionally detected as a tubercle on the dorsal aspect of the scaphoid or even as a separate bone. It is a separate bone in the carpus of all primates except the gorilla, chimpanzee, and man. There are two centralia in lower vertebrate forms.
The Pisiform (ulnare laterale of Forsyth Major) is of doubtful nature. It is possible that in a very early stage of the evolution of mammals there were more than five digits — one behind the little finger — post minimi digiti; and another on the radial side of the hand — a pre-hallux. Supernumerary digits, when they appear, are commonly situated on the radial side of the thumb or ulnar side of the little finger, but they may represent merely a fission of the normal pollex or little finger. The pisiform has been regarded as the vestige of a post-minimal digit ; the sesamoid on the trapezium, in which a slip of the extensor ossis metacarpi pollicis ends, as a remnant of a pre-hallux. It is possible also to regard the pisiform as a sesamoid developed in the tendon of the flexor carpi ulnaris — for that muscle is originally a flexor of the metacarpus and ends on the 5 th metacarpal — the pisi-metacarpal ligament representing the terminal part of the tendon. The pisiform, however, is developed with the rest of the carpal bones and before the tendon of the flexor carpi ulnaris. In mammals generally, but not in man, the pisiform articulates with the ulna as well as the cuneiform, and its synovial facet opens into the wrist joint. It may be represented in the foot by the heel epiphysis of the os calcis. The gastrocnemius, which represents the flexor carpi ulnaris in the leg, is also primitively a flexor of the metatarsus ; the long plantar ligament, from which it is separated by the growth of the heel, represents the continuation of its tendon.
The Eversion of the Foot and Development of the Arch

The human foot has been highly modified for upright progression. The chief modifications are :
(1) Gradual eversion of the foot, so that the sole can be applied to the ground. Even at birth — and for some time after — and always up to and before the 7 th month of foetal life, the soles of the feet are inverted, so that when the foetal limbs are in their natural position they are directed towards the belly of the child. In club foot the natural process of eversion does not take place. The ape's foot is kept normally in the inverted position, an adaptation for prehension.
The following factors assist in producing eversion :
(a) The neck of the astragalus (Fig. 248), which in the foetal foot is long and directed downwards and inwards at an angle to the axis of its body, becomes relatively shorter and directed more in line with the axis of the articular surface of its body (Fig. 248). Further, the outer border of the tibial articular surface of the astragalus is prominent in the foetus ; the inner border is much the lower ; a growth upwards of the inner border causes the astragalus and foot to rotate outwards (Lazarus).
(b) The bones on the inner side of the foot, particularly the scaphoid and internal cuneiform, grow more rapidly than those on the outer side of the foot — especially after birth. This tends to evert the foot and also to produce the longitudinal arch.
(c) A special evertor of the foot is produced — the Peroneus tertius — a muscle peculiar to man. It is developed from the outer and lower fibular fibres of the extensor longus digitorum and represents part of the tendon of that muscle to the 5th toe. The peroneus brevis and longus may also assist, especially the latter, which in apes is a grasping muscle, acting as a flexor of the metatarsal bone of the hallux.
(2) The tarsal bones of the human foot — especially the astragalus and os calcis — are of great size when compared with the tarsal bones of other primates ; while the digital or phalangeal elements, except in the case of the great toe, which is relatively of great size, have undergone retrogression. This is especially the case in the human little toe ; some of its muscles are not infrequently fibrous, and the terminal phalanx may not be separated from the middle phalanx. The terminal phalanx is the last to be differentiated in development of the fingers and toes.
(3) The plantar arches, both longitudinal and transverse, are produced. The arch of the foot is a human character. At birth the child is flat-footed ; the head of the astragalus touches the ground. The arch is produced as the child learns to walk. The chief factor in its production is the growth of the tarsal bones — especially of the scaphoid and internal cuneiform — and 1st metatarsal. Hence in rickets, where the normal tarsal growth is disturbed, the occurrence of flat foot. Amongst the structures which help to maintain the arch are :
- (a) The growth of the os calcis into the heel separates the tendon of the plantaris from its prolongation in the sole — the middle part of the plantar fascia, which assists in maintaining the arch. In lower primates the two parts are continuous, the tendon of the plantaris plying across the os calcis in a cartilage-lined groove.
- (b) The internal lateral ligament of the ankle (anterior part) and the inferior calcaneo-scaphoid ligaments undergo great development in man.
- (c) The flexor brevis digitornm which in lower primates arises principally from the long flexor tendons in the sole, has its origin completely transferred to the os calcis in man. It can thus act more powerfully in maintaining the arch. The flexor accessorius, a detached part of the flexor longus hallucis, is specially well developed and helps to maintain the arch of the foot.
- (d) The tibialis posticus, originally a flexor of the metatarsus, corresponding to the flexor carpi radialis in the hand, obtains a secondary attachment to the scaphoid. The tibialis anticus, which answers to the extensor ossis metacarpi pollicis, becomes permanently inserted to the internal cuneiform and metatarsal. Both of these muscles, thus modified, help to maintain the arch of the foot. So does the tarsal part of the tendon of the tibialis posticus.
- (e) The long plantar ligament, originally a part of the tendon of insertion of the gastrocnemius — also assists to maintain the arch. (4) The development of the great toe and the peculiar arrangement of its muscles must also be regarded as adaptations in the foot to upright posture and progression.
Comparison of the Muscles of the Upper and Lower Extremities
As a mental exercise it may be worth the student's while to briefly review the corresponding muscles in the two limbs. He has already seen that the arm and leg buds are similar in origin and made up of similar elements. Each is subsequently modified for its special function. The key to the homology of the muscles lies in their relationship to the digits, and their position on the limb.
Pollex and Hallux. — The extensor ossis metacarpi pollicis corresponds to the tibialis anticus. The thumb muscle has commonly a carpal insertion as well as metacarpal. The extensor brevis or primi internodii pollicis is constant in man only; it is a segment of the extensor ossis metacarpi pollicis.
The extensor brevis hallucis is not represented in the thumb.
Second Digit. — In the lower primates each finger has two extensors — a deep and superficial. The deep in the second digit becomes the extensor indicis ; in the little finger it forms the extensor minimi digiti. The deep extensor muscles have disappeared in man from the 3rd and 4th digits, but occasionally reappear. In the leg the deep extensors have migrated to the foot, and form the extensor brevis digitorum. That for the little toe, however, has not descended ; it is always vestigial, if present, and is commonly absent. It runs beneath or with the peroneus brevis, and is known as the peroneus quartus or peroneus quinti digiti.
Flexors and Extensors of the Metacarpus. — These have retained their primitive insertions in the hand; their modifications in the foot have been already mentioned. Both at the knee and elbow joint the origins of these muscles have undergone much shifting and migration.
The comparison already made between the scapula and ilium (p. 291), will help the student to understand the correspondence between the muscles of the thigh and arm.
Vestigial and Abnormal Muscles in the Limbs and Trunk


- The muscles of the human ear and scalp may be described as vestigial when compared to the development in other mammals. Although their action on the ear and scalp is feeble, yet they serve as most important bases into which certain psychological states are reflected.
- The levator claviculae (omo-trachelian) is a muscle which passes from the upper transverse cervical processes to the outer end of the clavicle or acromion process. It is well developed in climbing primates. It is not a common muscle in man. It can be recognised during life in the posterior triangle of the neck.
- The latissimo-condyloideus (dorso-epitrochlearis), a climbing muscle, is always represented in man, commonly by a fibrous bundle between the tendon of the latissimus dorsi and the long head of the triceps (Fig. 249). It may be occasionally muscular. In apes it passes from the latissimus dorsi at the axilla to the inner aspect of the elbow and arm, which it retracts in climbing. It belongs to the same sheet as the coraco-brachialis. The ligament of Struthers — a strip of fibrous tissue over the internal intermuscular septum, above the internal condyle — represents part of the tendon of this muscle. The muscular slips occasionally found crossing the brachial or axillary artery from the latissimus dorsi to the coraco-brachialis or biceps are derivates of this muscle. Other slips found crossing the floor of the axilla, between the adjacent borders of the pectoralis major and latissimus dorsi, are parts of the muscular sheet out of which these two muscles are developed.
- The pectoralis externus arises from the 4-5-6 ribs and costal cartilages, beneath the axillary border of the peetoralis major. This is its normal condition in most mammals, but in man it is commonly fused with, and forms part of, the peetoralis major.
- The sternalis is a new muscle. The peetoralis major is formed from the same ventral longitudinal sheet as the rectus abdominis and sterno-mastoid. The fibres of the sternalis, which lie along the sides of the sternum, superficial to the origin of the peetoralis major, represent a persistent part of the primitive longitudinal sheet. The sternalis is a derivative of the sphincter colli, part platysma-sheet (Parsons).
- In the sterno-mastoid, four elements are recognised : sternomastoid, sterno-occipital, cleido-mastoid, cleido-occipital. The cleido-occipital fibres, which form part of the same sheet as the trapezius, are often absent. On the other hand the cleidooccipital fibres may be continuous with the trapezius.
- The peetoralis minor is sometimes inserted to the capsule of the shoulder and great tuberosity of the humerus. This is the primitive insertion of the muscle. The coracoid insertion is a secondary attachment seen only in man and some of the higher primates. "When the peetoralis minor is inserted to the coracoid, the former fibres of insertion become fused with, and forms part of, the coraco-humeral ligament. The coraco-humeral ligament represents the ischio-capsular of the hip joint.
- In some apes (such as the Gibbons) the biceps has four heads — the two usual, the long and short, and two others, one from the inner border of the humerus and one from the bicipital groove. These two extra heads appear frequently in man.
- The epitrochieo-anconeus is frequently present. It crosses the ulnar nerve from the internal condyle to the olecranon.
- The palmaris longus and its homologue in the leg, the plantaris, are vestigial, aberrant in form, and often absent. The plantar and palmar fasciae represent their divorced tendons. The plantaris and palmaris undergo retrograde changes in the Primates with the transformation of claws to nails.
- Each digit (fingers and toes) in lower Primates, such as monkeys, is provided with three short muscles which arose from the carpus or tarsus. The three muscles are (Pig. 250): (1) a short flexor on the radial side of the digit ; (2) a short flexor on the ulnar side ; (3) a contrahens or adductor muscle (always absent in the middle digit). The ten short flexor muscles form a deeper sheet than the four contrahentes. Of this form the arrangement of the short muscles in the human hand is a derivative. The remnants in the human hand and foot of the contrahentes are: (1) The adductors of the 1st digit (pollex or hallux) ; (2) fibrous remnants of the others occur over the deep plantar or carpal arch (Fig. 250). The short flexors in man have become (1) the seven interossei ; (2) the fiexores breves (ulnar and radial) and opponens of the first digit ; (3) the flexor brevis and opponens of the fifth digit (see Fig. 250). The ulnar flexors of the thumb and great toe are absent or fibrous.
- The Pyramidalis is often absent in man or vestigial. It is the tensor of the linea alba.
- Eemnants of the extensors and flexors of the tail may occur between the sacrum and coccyx (page 136).
- The Coccygeus is vestigial; its superficial part forms the small sacro-sciatic ligament.
- Fibres of the biceps of the thigh may be followed into the great sacro-sciatic ligament. This ligament, which is almost peculiar to man — in other primates it is quite thin and slender — may contain fibres derived from the caudo-femoral group of muscles, such as the tenuissimus, a long strap-like muscle which passes from the coccyx to the femur and leg in lower mammals. The sacrosciatic ligament is mainly derived from the great median sheet, out of which the middle layer of the lumbar fascia is also formed. Parsons regards the short head of the biceps as a derivative of the tenuissimus, while others regard it as part of the muscular sheet which forms the peroneal muscles. Amongst Primates, it is found only in man, the anthropoids, and some South American apes. The short head of the biceps corresponds to the brachialis anticus in the arm, and is supplied by the external popliteal nerve.
- The psoas parvus is also vestigial. It acts primarily as a flexor of the pelvis on the spine. It begins to disappear with the assumption of the erect posture.
- The scansorius is a separated segment of the gluteus medius and minimus. It rises from the anterior border of the ilium and passes to the great trochanter. It corresponds to the teres minor. It is not constant in any animal.
- The flexor brevis digitorum to the little toe and the adductor transversus of the great toe are often fibrous.
Vessels of the Limbs
The Vas Aberrans
This vessel gives rise to a number of anomalous arrangements of the brachial artery. If a newly-born child be well injected, a number of branches derived from the axillary, brachial and main arteries of the fore-arm will be found to form an anastomatic chain along the superficial aspect of the median nerve. The upper end of the chain is formed by a branch from the axillary ; the lower end is completed by a branch of the ulnar or radial at the elbow. In South American apes this anastomosis always opens up to form a supplemental brachial artery — the vas aberrans — ending usually as the radial. From the manner of its origin and disposition in front of the median nerve, it will be seen that the vas may arise from the axillary or upper part of the brachial, and terminate in the radial, ulnar or brachial, according to the branches which participate. It may be so large as to simulate a large branch of the axillary or a division of the brachial. Such a condition is spoken of as high division of the brachial. The vas may supplant the brachial artery altogether, which is then represented by a trunk which ends high up in the arm by giving off the superior and inferior profunda arteries. The brachial vessel formed from the vas is recognised by its superficial position to the median nerve. In lower apes, the brachial artery commonly divides a little above the elbow. This position of division is rare in man. In cases of high division the radial, or even the ulnar, may be superficially placed in the forearm.
Internal Saphenous Artery
In most mammals — in all primates except man — the anastomotica magna of the femoral artery is as large as the popliteal and passes over the inner side of the tibia with the internal saphenous vein to reach the dorsum of the foot, where it forms the dorsalis pedis artery. This vessel is known in them as the internal saphenous artery, and corresponds to the radial artery in the forearm and wrist. The superficial branch of the anastomotica magna rarely assumes such a development in man ; but the course taken by the internal saphenous artery explains the position of its accompanying vein, the internal saphenous, in front of the internal malleolus. In man only is the internal saphenous vein continued up the thigh to a saphenous opening in the groin. Its primitive termination is in the femoral vein at the lower end of Hunter's canal.
The Superficial Plantar Arch, formed from the internal plantar artery, is seldom complete in man. The pressure to which it is subjected in the standing posture has led to its partial obliteration. It corresponds to the superficial palmar arch.
The Supra-condylar Process is well developed in lemurs, the lowest primates, and in mammals of many orders. Its function is unknown. It occasionally appears in man. It is developed from the humerus about two inches above the internal condyle as a hook-like process of bone. It lies in front of the internal inter-muscular septum, and when well developed the brachial artery and median nerve may pass beneath it, as they do in such animals as the squirrel and cat.
Third Trochanter. — A tubercle may appear in the gluteal ridge which receives this name. It is cartilaginous until the 20th year, when a separate centre of ossification appears in it (Dixon). It is well developed in the horse.
Ligaments and Joints of the Limbs
The Inter-articular Cartilages are of doubtful origin. In their first appearance, by the condensation of the mesoblast in the axis of the limb bud, the bones are continuous. The joints are formed between the bones, by the mesoblastic tissue remaining fibrous, while that which forms the bases of the bones undergoes chondrification. The mesoblastic tissue thus left between the cartilages opens out and forms a synovial cavity and a fibrous capsule. Hence the periosteum of the bones is continuous with and derived from the same primitive layer as the joint capsules. The inter-articular cartilages of the knee, for instance, may be simply masses of mesoblast left between the bones, originally part of the condensed mesoblastic bar out of which the femur, tibia and fibula were developed. The inter-articular cartilage of the temporo-maxillary joint may be a derivative of Meckel's cartilage ; that of the sterno-clavicular of the pre-coracoid. The triangular fibrocartilage of the wrist joint is derived from the ulno-carpal ligaments, and interosseous membrane (Parsons). The cartilage may also contain a post-minimal element (Corner). The cartilages of the knee are of obscure derivation ; they may be parts of muscles formerly inserted on the upper end of the tibia (Sutton). Parsons has shown that in all primates, except man, the external semilunar cartilage is circular in form and is attached to the posterior crucial ligament.

Fig. 251. Showing the Origin of the Ligamentum Teres and Reflected Bundle of the Capsular Ligament.
The Ligamentum Teres is to be regarded as a part of the capsule of the hip joint which has been cut off by the outgrowth of the articular surface of the head of the femur (Fig. 251). The crucial ligaments are semi-isolated fasiculi from the capsule of the knee joint, separated in a similar manner to the ligamentum teres by the outgrowth of the condyles of the femur (Fig 252). At first the ligamentum mucosum, crucial ligaments and posterior part of the capsule are continuous and form parts of the capsule of the joint ; they come to occupy a position within the joint as the femoral condyles grow backwards.

Fig. 252. Showing the Origin of the Crucial Ligaments of the Knee.
In reptiles and in lower mammals the fibula enters into the formation of the knee joint as the ulna does in that of the elbow. This is at no time the condition in the human foetus (G-runbaum). The external malleolus in primates is generally shorter than the internal ; this is the case in man until the 3rd month of foetal life ; after the 3rd month the external is the longer (Wilgress).
Chapter Figures
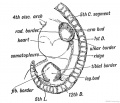
Fig. 233. Lateral view of a human embryo at the 28th day, showing the Limb Buds, Lateral Edges, and Primitive Segments.
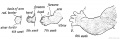
Fig. 234. Development of the Upper Limb.
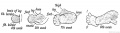
Fig. 235. Development of the Lower Limb.
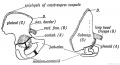
Fig. 236. The Corresponding Points {A, B, C, and D) in the Ilium and Scapula.
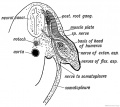
Fig. 237. Section of the Arm Bud of a human embryo at the end of the 4th week.
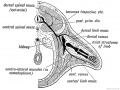
Fig. 238. Schematic Section showing the Origin and Arrangement of the Muscles and Nerves of the Limbs.
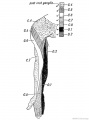
Fig. 239. The Distribution of the Posterior Roots of the Spinal Nerves on the Flexor Aspect of the Arm.
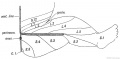
Fig. 240. Diagram to show the typical Manner in which the Posterior Nerve Roots are distributed in the Lower Limb.
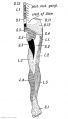
Fig. 241. Flexor Aspect of the Lower Limb showing the Sensory Distribution of the Segmental or Spinal nerves.
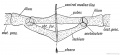
Fig. 242. Diagram of the Pelvic Girdle of a Lizard.
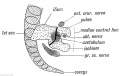
Fig. 243. The Pelvic Girdle of a Human Foetus at the 5th week.
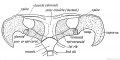
Fig. 244. The Shoulder Girdle of Ornithorynchus.
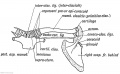
Fig. 245. The Parts in the Shoulder Girdle of a human foetus which correspond with those of Ornithorynehus.
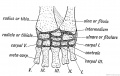
Fig. 246. The Carpal Bones of a Tortoise.
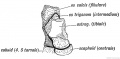
Fig. 247. The Os Trigenum and Bones of the Tarsus
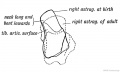
Fig. 248. The Foetal and Adult (in dotted outline) Forma of the Astralagus contrasted.
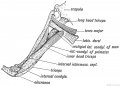
Fig. 249. Latissimo-condyloideus Muscle.

Fig. 250. The Morphology of the Short Muscles of the Digits.
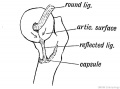
Fig. 251. Showing the Origin of the Ligamentum Teres and Reflected Bundle of the Capsular Ligament.
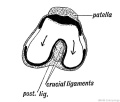
Fig. 252. Showing the Origin of the Crucial Ligaments of the Knee.
- Links: Limb Development
| Historic Disclaimer - information about historic embryology pages |
|---|
|
Human Embryology and Morphology (1902): Development or the Face | The Nasal Cavities and Olfactory Structures | Development of the Pharynx and Neck | Development of the Organ of Hearing | Development and Morphology of the Teeth | The Skin and its Appendages | The Development of the Ovum of the Foetus from the Ovum of the Mother | The Manner in which a Connection is Established between the Foetus and Uterus | The Uro-genital System | Formation of the Pubo-femoral Region, Pelvic Floor and Fascia | The Spinal Column and Back | The Segmentation of the Body | The Cranium | Development of the Structures concerned in the Sense of Sight | The Brain and Spinal Cord | Development of the Circulatory System | The Respiratory System | The Organs of Digestion | The Body Wall, Ribs, and Sternum | The Limbs | Figures | Embryology History
Reference
Keith A. Human Embryology and Morphology. (1902) London: Edward Arnold.
Cite this page: Hill, M.A. (2026, April 17) Embryology Book - Human Embryology and Morphology 20. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Book_-_Human_Embryology_and_Morphology_20
- © Dr Mark Hill 2026, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G