Paper - The Sexual Cycle in the Human Female as revealed by Vaginal Smears (1933)
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Papanicolaou GN. The sexual cycle in the human female as revealed by vaginal smears. (1933) Amer. J Anat. 52: 519–637.
| Online Editor |
|---|
 The original black and white photomicrographs were hand-coloured by the author to match his microscopic observations. |
| Historic Disclaimer - information about historic embryology pages |
|---|
|
The Sexual Cycle In The Human Female As Revealed By Vaginal Smears
George N. Papanicolaou
Department of Anatomy, Cornell University Medical College, and Woman’s Hospital, New York City
- This work has been aided by the Committee for Research on Sex Problems of the National Research Council, and by the National Committee on Maternal Health.
Three Figures and Ten Plates (Eighty-One Figures)
- drawings of cells in human vaginal smears
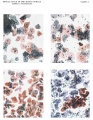
Plate 8
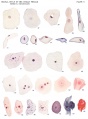
Plate 9
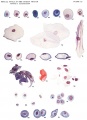
Plate 10
I. Introduction
The study of the female sexual functions in mammals has been greatly stimulated and advanced in recent years by the application of the vaginal smear method. This method, as originally applied to the guinea pig by Stockard and Papanicolaou in 1917, consists in the microscopic examination of smears prepared at frequent intervals from the fluid content of the vagina. The vaginal fluid usually has a mucous consistency and contains a variety of desquamated cells, as well as leucocytes, lymphocytes, often erythrocytes, and a large number of bacteria. As the relative number and the distribution of these elements change periodically, smears prepared from such fluid show modifications in their composition and structure. The successive alternation of periods of sexual activity and inactivity, which characterizes the mammals, imparts to the vaginal fluid a rhythmical sequence of typical cellular stages which can be easily recognized.
These cyclic changes affect the entire genital tract, and, consequently, every change in the vaginal fluid is strictly correlated With corresponding changes in the other organs of
the female genital system, particularly the uterus and the ovaries. The time of ovulation may be accurately detected by this method in living mammals, while, before its application, no such information could be obtained without an operation or the sacrifice of an animal.
In the guinea pig, which is polyoestrous, the sexual cycles
return periodically throughout the year every 15 to 16 days
(15.73 in average). The longest part of each sexual cycle,
i.e., about 12 days, is occupied by a period of relative inactivity or rest, which is called ‘clioestrus.’ During this time
the vaginal smear, as described by Stockard and Papanicolaou ( ’l7), consists chiefly of leucocytes and a varying num-
ber of atypical squamous cells. The period of increased
sexual activity lasts 3 to 4 days and is characterized by a
succession of stages, which have been designated as stages
I, II, III, IV. During stage I the leucocytes almost disappear
from the smear, the secretion of mucus becomes more abundant, and the cells which dominate the smear are of a squamous type with very small pyknotic nuclei which are at times
fragmented. At the end of stage I there is an intermediate
period, characterized by the prevalence of ‘elongate, cornified
cells Without nuclei.’ Stage II also shows a scarcity of leucocytes and a prevalence of cells which are derived from the
deeper layers of the vaginal epithelial wall, being thus less de-
generated and having a larger nucleus and a more compact
form. Stage III is characterized by the reappearance of
myriads of leucocytes and by cells mainly of the II and III
type. The III cells are as a rule modified type II cells, the
bodies of which had been penetrated by the invading leucocytes. Stage IV is practically the same as stage III, with the
difference that erythrocytes are also present in the smear as
the result of slight bleedings.
These four smear stages are of short duration, succeeding one another rapidly and are strictly coordinated with corresponding stages of the uterine and ovarian cycles. During
stage I the ovary contains large ripening follicles and regressing corpora lutea. The uterus is congested and hypertrophied and its epithelial lining consists of high cuboidal or columnar cells. During the intermediate or cornified stage the follicles as well as the uterus reach their highest development. The onset of the catabolic processes, i.e., the bursting
of the ripe follicles and the denudation of the uterine mucosa,
is associated with the appearance of vaginal stages II and III.
Ovulation in the guinea pig is thus characterized by definite
vaginal changes and occurs at about the time of the sloughing
off of the uterine mucosa. Comparable conditions have been
found to exist in other rodents, such as the rat and the mouse.
Long and Evans (’22), in their monograph on the oestrous
cycle in the rat, described definite relations between the vaginal cycle, as revealed by vaginal smears, and the uterine and
ovarian cycles. The rat has normally a very short sexual
periodicity of about 5 days’ duration. Long and Evans recognized five smear stages: Stage 1: Leucocytes disappear;
great numbers of small round nucleated epithelial cells of
strikingly uniform appearance and size; duration 12 hours.
Stage 2, or ‘cornified cell stage’: leucocytes still absent;
appearance of cornified cells. Stage 3, or ‘late cornified cell
stage’: leucocytes absent; rich in cornified cells, forming
large cheesy masses; duration of stages 2 and 3 about 30
hours. Stage 4, or ‘leucocytic—cornified cell stage’: reappearance of leucocytes; gradual disappearance of the cornified
cells and appearance of epithelial cells; duration 6 hours.
Stage 5 corresponds to the dioestrus, which is of an approximate duration of 3 days and is characterized by the presence
of leucocytes and atypical epithelial cells in the vaginal smear.
The cornification of the vagina is much more pronounced
in the rat than in the guinea pig and this accounts for some
of the smear differences between these two animals. In the
rat the cornification extends over two well-defined stages, the
second and third of Long and Evans, whereas in the guinea
pig it is usually overlapped by the I and II to the III stages;
on this account, it had been originally described by Stockard
and Papanicolaou as an intermediate period. In the rat the
uterine growth is completed during the first and second
stages. The catabolic processes in the uterus begin during
the cornified cell stages, and the ovulation occurs during the last hours of this same period. The uterine epithelium is
vacuolized and degenerated, but an actual denudation, as in
the guinea pig, does not occur. The various phases of the
sexual cycle in the rat proceed in rapid succession, but their
coordination is maintained.
The corresponding vaginal smear stages in the guinea pig and the rat could be illustrated as follows:
| Vaginal Smear Comparison Table | |
|---|---|
| Guinea pig | Rat (Long and Evans) |
| I. Superficial squamous cells with pyknotic nuclei; progressive leucopenia. | 1. Small round nucleated cells; disappearance of leucocytes. |
Intermediate (cornification) period
|
Cornification period
|
| II, III, IV. Appearance of deep layer cells; reappearance and great exodus of leucocytes; gradual disappearance of cornifiecl cells; sometimes erythrocytes present. | 4. Leucocytic-cornified cell stage; reappearance of leucocytes; gradual disappearance of the cornified cells. |
| V. Dioestrus: Leucocytes and atypical atypical vaginal cells. | 5. Dioestrus: Leucocytes and vaginal cclls. |
| |
This comparison indicates that the succession of stages is similar in both animals. Ovulation occurs at practically the same moment in relation to the above stages, i.e., at the end of the intermediate period in the guinea pig and during the last hours of the cornification period (second and third) in the rat.
In the mouse, Allen (’22) has also recognized five stages, the entire sexual cycle averaging about 4% days.
Selle (’22) has reclassified the vaginal smears of the guinea pig after a detailed study of the changes in the vaginal epithelium. He has recognized a separate ‘cornified cell stage’ similar to the same stage in the rat. Such a stage may be actually present in a large number ‘of cases, but, as a rule, it is indistinct and overlaps both the preceding and the following stages. This cornified cell stage of Selle corresponds to the ‘intermediate period’ of Stockard and Papanicolaou, and not to the stage 2 as given in Selle’s table. His stage 3 corresponds to the intermediate +II, whereas his stage 4 is equivalent to the stages III and IV of Stockard and Papanicolaou.
Murphey (’22) and Frei and Metzger (’26) have recognized four stages in the oestrous cycle in the cow with the vaginal smear method. Hartman (’23), in his study of the oestrous cycle in the opossum, also recognized four stages corresponding to the ones described in the rodents.
In 1924, McKenzie and Zupp, and in 1926, Wilson studied vaginal smears in swine. They found periodic variations in the cellular and leucocytic make-up of the smears, indicative of a cyclic rhythm.
A further step was made by Corner’s (’23) application of
this method to the Primates. His observations on Macacus
rhesus, though not entirely in line with the smear findings
in the rodents, yet revealed a rhythm in the vaginal reactions.
The average length of cycle in the Macacus, when the regular
cycles are considered, is about 27 days, which is almost the
same as in the human. During menstruation, which lasted
4 to 6 days, the vaginal smear contained erythrocytes, epithelial cells, and leucocytes. In the first half of the intermenstrual interval there were relatively few epithelial cells and
many leucocytes. About the middle of the interval a sharp
drop in leucocytes occurred or even total disappearance.
Leucocytes sometimes reappeared a few days later or were
absent until onset of next menstruation or a few days before.
During the second half of the intermenstrual interval there
was an increased desquamation of epithelial cells. The pre-menstrual smear seemed to be thick and caseous, whereas the
postmenstrual smear was rather thin and scanty.
These observations, though not establishing a definite succession of clear-cut stages, reveal the existence of a rhythm expressed mainly in the periodical increase and decrease in the number of the leucocytes and the epithelial cells. As Corner concludes:
- There is to some degree a cycle of the vaginal secretion in this species. There is hardly enough evidence to warrant a correlation with the much sharper cycle of the rodents; moreover there was seen in the monkeys no massive desquamation of completely cornified epithelial cells and no swarming of leucocytes into the epithelial debris. However, . . . . it seems very likely that the disappearance, or diminution in number of vaginal leucocytes, which usually happened about the tenth to the fifteenth day before the onset of menstruation (in regular cycles of 25 to 30 days) is to be compared with the disappearance of leucocytes from the vagina of rodents at oestrus or shortly before the moment of ovulation.
In a later paper, 1927, in which the presence of menstruation without ovulation in Macacus rhesus was reported, Corner stated that daily vaginal smears, taken from animals with ovulative and non-ovulative cycles, were practically alike. The presence or absence of ovulation could not be ascertained by the examination of vaginal smears.
Allen, in 1927, also studied vaginal smears of the monkey, Macacus rhesus, and found varying numbers of epithelial cells in different stages of cornification. Some of these cells were quite normal, others were flattened and their nuclei were pyknotic. The epithelial elements were present in greatest numbers during the latter half of the second and the whole third week of the cycle. Completely cornified, non-nucleated cells frequently also appeared at these times. Leucocytes were present in greatest numbers before, during, and after menstruation and in least numbers or absent between the tenth and the twentieth to twenty-fourth day of the shorter cycle. During menstruation varying numbers of erythrocytes were present.
Allen’s findings are more or less in line with Corner’s observations, especially in regard to leucocytes. Corner found many leucocytes during and after menstruation and Allen before, during, and after. At about the tenth day both noticed a progressive diminution in the number of the leucocytes. This diminution lasted for a few days or almost up to the onset of next menstruation. In regard to ovulation, they agree that, whenever present, it occurs in the mid-period, between the tenth and the fifteenth days.
The preparation of vaginal smears from the human has been in usein pathology for a long time for the study of various conditions, especially of bacterial infections. However, the application of smears to morphological and physiological studies has been extremely limited up to the last decade.
As early as 1847, Pouchet, in his book on “Ovulation and other related phenomena,” gave a description of human vaginal smears, which is chiefly interesting from a historic standpoint. Though unaided by modern technical methods, he
was able to recognize the existence of a rhythmical reaction
in the vaginal secretion. His work, however, was largely lost
sight of and in no way stimulated attention upon the value
of the vaginal content as an indicator in analyzing the phases
of the sexual cycle. The recent interest and activity in these studies can in no sense be connected with or attributed to this early pioneer effort by Pouchet.
He describes the vaginal mucus as becoming less dense
shortly before menstruation and as acquiring a peculiar odor
(une odeur ‘sui generis’), to which he attributed an exciting
effect upon sex desire. Microscopically, the vaginal fluid
showed fragments of epithelium and lacerated pieces, some
consisting only of the ‘tubercule central’ (he evidently meant
the ‘nucleus’), also large numbers of ‘globules muqueux’ (meaning probably the leucocytes) and some erythrocytes.
During the menstrual phase (‘période d’état’) he records the enormous quantity of erythrocytes, of mucous globules, and of small and transparent epithelial fragments. He believed that ovulation occurs toward the end of the menstrual phase, when the sex desire is most imperative. The menstrual phase is followed by a ‘period of desquamation,’ lasting approximately 10 days. This period is characterized by the detachment of a considerable quantity. of epithelial plates (‘Plaques d ’épithélium ’) .
During the sixth and seventh days after the end of menstruation (eleventh to twelfth day after onset) the vaginal mucus begins to lose its transparency and becomes heavier. Large numbers of epithelial plates are present and the mucous globules become more abundant. At about this time, or on the eighth day after menstruation (thirteenth day since the onset), some women experienced a feeling of heaviness or sharp pains lasting 1 to 2 days. Pouchet attributed these to contractions of the fallopian tubes, and not to the ovulative process, which he thought to occur much earlier.
Between the tenth and fiftenth days after cessation (fifteenth to twentieth day after onset of menstruation) pieces
of uterine decidua (‘flocon membraneux’) were expelled in
a number of cases. Pouchet interpreted this as an abortive
process and believed that a woman could only conceive between the time of menstruation and the spontaneous fall of
this decidua.
In 1921, Lehmann made a study of the diagnostic value of
the human vaginal smear. Interested chiefly from a pathological and diagnostic point of view, he did not attempt to
establish definite morphological and physiological relations
between the changes in the vaginal fluid and the ovarian and
uterine cycles. He recognized, however, the dependence of
certain vaginal conditions, such as secretion of glycogen,
acidity, or bacterial growth on ovarian and uterine functions.
In 1925, I announced in a preliminary report some of the early results of my human vaginal smear studies. I held that there are definite morphological changes in the vaginal fluid by which a diagnosis of certain physiological and pathological conditions is made possible.
Pregnancy, cystic or other degenerative changes of the
ovaries, inflammatory processes, growth, etc., affect the entire
genital tract, including the vagina, in a way which produces
definite and typical changes in the consistency and make-up
of the vaginal smear. The presence or absence of different
types of desquamated cells, as well as the varying form and
number of leucocytes, lymphocytes, erythrocytes and bacteria,
offer a variety of criteria upon which a diagnosis of certain.
conditions may be based.
These observations were mainly on the morphological changes of the various constituents of the smear, and not merely on quantitative estimates of the relative number of
leucocytes or cells.
In the same year, Allen (’25) reported tests on human vaginal smears from gynecological patients in collaboration
with Dr. Q. Newell. He stated that—
- Although they found a decided variation in the number of leucocytes and epithelial cells at different times in the cycle, results were not nearly so clear-cut as in rats, because normally no cornification occurs in the vaginal epithelium of women. Furthermore, the smear test is of greatest value in animals, in which sexual changes are not as clearly marked externally. In the primates menstruation furnishes such a prominent milestone that vaginal smears seem of secondary importance for diagnosis.
A year later, King ( ’26) published her studies on human vaginal smears. She found that “in general the secretion is more scanty during the first few days following menstruation and the cells show less degeneration during the first part of the cycle than in the late intermenstrual and premenstrual phases. The leucocytes are of the polymorph type, although an occasional mononuclear can be seen.” She also found a definite decrease in the number of leucocytes and a relatively high content of epithelial cells in one cycle of case D, on the thirteenth day after one onset and 16 days before the next. A similar condition was seen in the third interval of case F, beginning in the middle of a 22-day cycle.” She considers this fall in leucocytes as being probably comparable to the intermenstrual decrease noticed in some of Corner’s monkeys which may bear a relation to the time of ovulation. She states, however, that there were great irregularities in this respect in other cases.
Two types of secretion described by Lehmann for normal adults may be found, according to her, in the same individual and, in addition, there is a type in which the content in both
leucocytes and epithelial cells is high. She concludes that—
The cellular content of the vaginal secretion of the normal human female is exceedingly variable and is a doubtful index of changes transpiring in the ovary and uterus. The indications of periodic variation are even less evident than those found by Corner for the monkey. Considering the higher degree of specialization, this is the condition which might reasonably be expected.
A Mexican gynecologist, Ramirez[1] published, in 1928, a study of the human vaginal smear cycle. He recognized five provisional types of cells: I) Cells with a large and round nucleus clearly outlined, finely reticulated, and taking a violet-reddish color with Leischmann’s stain; cytoplasm reticulated, pale-blue, with or without inclusions. II) Cells with a large round or oval nucleus having a thick reticulum, granular or semigranular; cytoplasm spongy, bluish, with inclusions and vacuoles. III) Cells with a much smaller nucleus having a thick reticulum or forming granules, irregularly outlined, taking a blue-violet color; cytoplasm condensed, bluish, vacuolated with inclusions. IV) Cells with a small, pyknotic granular or simply structureless nucleus of bluish or wine shade; cytoplasm spongio-granular with abundant inclusions and with or without large vacuoles. V) Anucleate cells and cells with only a trace of a nucleus. The nucleus appears as a mass of granules which seem to be dispersed in the granular cytoplasm. All the granulations, nuclear or cytoplasmic, display the same bluish color.
Ramirez found that during menstruation the prevailing cellular type is: I (49 per cent), II (27 per cent), III (20 percent), IV (4 per cent), and V (0); leucocytes, scarce at first, increase toward the end. During the post menstrual period (2 to 3 days after menstruation) the cell percentages were as follows: I (1 per cent), II (9 per cent), III (24 per cent), IV (60 per cent), V (6 per cent) ; many polynuclear neutrophilic leucocytes. During the early interval (4 to 8 days after menstruation): I (0), II (15 per cent), III (25 per cent), IV (52 per cent), and V (8 per cent). During the interval (8 days after menstruation) : I (0), II (1 per cent), III (42 per cent), IV (45 per cent), and V (12 per cent) ; leucocytes exceptional.
Type V was prevalent in pregnancy, where the cellular percentages were as follows: I (0), II (0), III (18 per cent), IV (12 per cent), and V (70 per cent). The aspect of the smear was entirely different during pregnancy and the cells showed a tendency to form large groups or laminae. Ramirez arrived at the same conclusion that I had in 1925, that pregnancy may be diagnosed through the cytological characteristics of vaginal smears. His cellular classification is, however, different from mine and his diagnosis is based rather on quantitative estimates. The increase of leucocytes during and immediately after menstruation and their decrease during the interval, as observed by Ramirez, are in accordance with the previous observations of Corner and Allen in monkeys and of King in the human being.
- ↑ A preliminary report on the same subject was published by this author in 1922, but unfortunately I have been unable to obtain the article.
Hartman, in his paper on gestation in Macacus rhesus (’28), presented some data on vaginal smear changes under normal conditions and during pregnancy. He found that the only successful mating in his Macacus took place at the ninth to twelfth day. During the fertile mating leucocytes were still present in the Vagina (though on the decline). Soon after copulation there was a temporary vaginal leucocytosis. He recognized two different types of leucocytes: those with unstained nuclei and those whose nuclei stained intensively with methylene blue. He stated that—
The total number of leucocytes increases during the second
half of intermenstruum, reaching a maximum just before (or
during) menstruation, whereas they go down to nearly zero in
the mid-interval; the curve of greatest desquamation from
the vaginal wall rises to a maximum in the latter part of the
interval, to fall, usually very low, about the time of menstruation.
This is in accord with Corner’s and Allen’s views. He also made observations on the cellular changes during gestation. He noticed the presence of the cells of Papanicolaou (navicular). He also found erythrocytes in the pregnancy smear for about 23 days, beginning at the twenty-sixth day since the onset of last menstruation (or 14; to 17 days after fertile copulation). He compares this with the bleeding observed by Long and Evans at the fourteenth day of gestation in the rat. In later stages of pregnancy Hartman found a leucocytic decrease and an increase of the resorption of the placental hormone as revealed by the exacerbation of the sex color. From these observations, he believes that “the disappearance of leucocytes from the vaginal smear is due to the follicular hormone.”
In the same year a detailed cytological study of the human
vaginal secretion was published by Moser (’28). She studied
ten cases of virgins and one sexually abstaining woman. She
examined vaginal smears daily or every second day during
one or two menstrual periods. The fluid was taken with the
aid of a ‘platinum loop’ and the smears were dried and then
fixed with methyl alcohol and stained with Delafield’s haematoxylin and eosin. The results were classified according to the relative number of epithelial cells, leucocytes, cornified cells (Schollen), and detritus.
Moser was unable to detect any regularity in the consistency of the vaginal fluid during the successive phases of the menstrual cycle. Epithelial cells were always present. Their cytoplasm and nucleus showed various degrees of degeneration. The whole structure was often covered by abundant bacteria. The nuclei of the cells were degenerated, pyknotic, or fragmented.
Isolated eosinophilic cells (Schollen) were found. Only in one case were they very numerous (about 7 per cent of the total). Moser also described nucleated cells which were stained darker with eosin and which could be interpreted as
transitional forms. She found no evidence that the human
vaginal epithelium undergoes a periodical cornification.
There was great fluctuation in the number of leucocytes, but
no typical leucopenia. In one case there was a premenstrual
increase in the number of leucocytes, but this was not constant. Most of the leucocytes were polymorphonuclear, some enclosed within the cells. Mononuclears were also occasionally observed. Most of the leucocytes were in process of degeneration. Her conclusion was that the morphological changes of the vaginal secretion are not typical enough to supply information regarding the functional condition of the ovaries.
A comparative review of all the investigations of this character conducted in various classes of mammals shows that
the results obtained in the rodents are by far the most satisfactory. The vaginal smear changes in these animals are
clear-cut and characteristic of the more fundamental changes
occurring in the ovary and the uterus. The time of ovulation can be accurately diagnosed in the rodents by definite
smear syndromes. The moments in their sexual rhythm may
be easily" established by the microscopical examination of
a sample of vaginal fluid. This has greatly advanced our
knowledge of the sexual and reproductive phenomena in mammals, and endowed us with indicators which may be profitably applied for obtaining valuable experimental information.
Various problems related to the ovarian hormones in particular as well as other hormonal reactions can now be attacked with greater accuracy.
In the higher mammals and in the Primates the results obtained so far have not been of equal clearness. In monkeys
and in the human female, in which the sexual rhythm is revealed by the menstrual bleeding, Very little information has
been gained by the application of the vaginal smear method.
We have learned that Primates, as Well as other mammals,
possess a definite vaginal rhythm. It has also been observed
that certain physiological and pathological conditions of the
female genital tract are revealed by vagi.n.al discrepancies.
These facts fully justify continued researches in this field,
and it is believed that the present contribution furnishes much
additional material toward a clearer understanding of the
human cyclic changes as evidenced by an analysis of the
vaginal smears of women.
2. Application of the Vaginal Smear Method in the Human
Material and technique
The correlation of the vaginal smear stages, as described in the rodents and in various other mammals, with analogous stages in women offers great difficulties. The extreme prolongation of the human female sex cycle has had a modifying effect upon the manifestation and the sequence of the vaginal fluid changes. The various stages are less typically expressed than in some of the lower mammals and are subject to a greater variability.
During the first 4 years of my studies (1920 to 1924), I was unable to correlate my observations on the human with those obtained in other mammals and more particularly in the rodents. In the spring of 1924, however, I first appreciated a stage interpreted as a transition between the copulative and the ovulative stage. And yet both these stages, and especially the second one, looked quite different from the corresponding stages in the rodents.
An explanation for these differences was offered one year later, when I first examined a human post-partum smear. This type of smear showed a surprising similarity to the missing
stage III, which, in the guinea pig, is Very closely and typically
related to ovulation. It thus became apparent that the guinea
pig stage III was not primarily an ovulative one. Its almost
infallible relation to ovulation was a mere coincidence. It
should be interpreted as resulting from utero-vaginal catabolic processes, corresponding to a pseudo-postpartum condition. It also became evident, from a morphological standpoint, that the prooestrous hyperplasia should be interpreted
as a pseudopregnant condition, and the onset of the cyclic desquamatory and catabolic phenomena of the utero-vaginal
tract as the equivalent of a pseudoparturient or pseudoabortive process.
If this explanation were correct, it would necessarily lead to the old assumption that human menstruation and its associated catabolic phenomena are of a pseudo parturient or pseudoabortive character (Beard, ’97 ). lonsequently, a
pseudo-postpartum stage, corresponding to the guinea pig
stage III, ought to appear in the human toward the end of
the menstrual process. This has been fully corroborated by
later findings. Toward the end of menstruation a pseudo—III
or pseudo—postpartum stage has been identified marking the
end of the destructive period, just as the guinea pig stage III marked the end of the utero-vaginal destructive phenomena. A positive explanation has thus been offered for the significance of certain smear stages, and their classification may be thus based on a fundamental interpretation of the female sex-cycle processes.
Most of the smears which I examined up to February, 1925, were obtained from a single human case (special case). This was an ideal case to work with, because of perfect regularity
in the menstrual periodicity and of a complete lack of any serious disturbance or of any bacterial contamination. It has been by far the most normal and typical case that I have had an opporunity to study. Several smears obtained during this first period from the clinic of Cornell Medical College in New York City, through the courtesy of Dr. John McGrath, were taken mostly from pathological cases. These have been little utilized for the study of normal standards.
In February, 1925, a valuable association was established with the Woman’s Hospital of New York City. This was rendered possible through the cooperation of Dr. George Gray Ward, director of the Woman’s Hospital, Dr. Charles R. Stockard, administering a grant from the Committee on Problems of Sex of the National Research Fund, and Dr. Robert L. Dickinson, representing the Maternal Health Committee. A special assistant was assigned to this work, whose technical skill in the preparation of smears has been extremely valuable.
Our first effort was to obtain a. sufficient number of control smears from normal cases. We selected twelve healthy young women, the majority from the personnel of the hospital, and arranged to obtain daily vaginal smears through several menstrual periods without interruption. Later, the material was extended to include smears from pregnant women who came to the clinic of the hospital for examination, and also from various pathological cases treated at the hospital.
From the start of this Work at the Woman’s Hospital up to the end of 1929, more than 900 cases were examined. Almost every important pathological condition has been more or less thoroughly considered. There have been smears from cases of pregnancy, abortions, threatened abortions, ectopic pregnancies, various infections or inflammatory processes of the genital tract, cystic ovaries, pus tubes, and various benign and malignant tumors of the ovaries, uterus, cervix, vagina, and mammary glands. There were also several cases of sterility due to various causes.
This study of pathological smears has helped in an understanding of normal processes and opened a new possibility for diagnosing various gynecological conditions. In a preliminary note (1925), I referred to the application of the vaginal smear method as a guide in the diagnosis of pregnancy and of pathological conditions.
In the last few years attention has been given to the characteristic smear changes Which seem to accompany cases of malignant tumors of the genital tract. A preliminary report on these observations was given at the “Third Race Betterment Conference” (1928).[1] Several types of abnormal cells with enlarged, deformed, or hyperchromatic nuclei are present in such smears.‘ Leucocytes are numerous and display a high phagocytic activity. Mononuclears become especially prominent. Erythrocytes are commonly present. A more detailed discussion of this phase of the study will be given in a subsequent publication. In this paper We intend to describe only the normal smear findings and to discuss their possible application in a detailed analysis of the human female sex cycle.
- ↑ Unfortunately, many stenographic errors have been included in this article, some altering the meaning of the text.
The technique used in these studies has been rather simple. In dealing with human material one benefits greatly by simplifying methods and avoiding unnecessary technical difficulties. In order to obtain the samples of vaginal fluid a glass pipette 8 inches long and 3/8 of an inch in diameter was used. The pipette was slightly curved at the end. The opening was small and a strong rubber bulb at the opposite end was used to produce suction in collecting the fluid (fig. 1). The sample of vaginal fluid may be taken with the aid of one of these pipettes by a technician or a nurse. Some of these patients were able to take their own smears. The sample is obtained from a region5 or ‘6 cm. within the vagina and is spread on a glass slide as a heavy film, much heavier than an ordinary blood smear. Soon after the spreading and before the smear has dried the slide is placed in a jar containing equal parts of 95 per cent alcohol and ether for quick fixation. When ether was not available, plain alcohol was used.
Fig. 1 Pipette used for taking samples of human vaginal fluid.
Proper fixation of the vaginal smears before drying is important, although this has been somewhat underestimated by most of the workers in this field. A smear which has been allowed to dry either before or after fixation may serve for the simple recognition of a typical stage, as has been done on rodents, but is not suitable for a detailed cytological study, especially in the human. The cells, as well as all other elements, are affected by the drying and lose their true appearance. Several investigators have probably failed to recognize certain morphological characteristics because of the use of dry smears.
The slides were kept in the alcohol-ether fluid for from half an hour to one hour, but smears may be kept in this fluid for several hours or even days without much harm. After fixing in this way the slides are carried through 80 per cent, 70 percent, and 50 per cent alcohol into distilled water as preliminary to staining.
The staining of vaginal smears with haematoxylin-eosin
gives good general results and a fair differentiation of the
cornified cells, and such a stain may be considered as very
suitable for the recognition and study of the fundamental
morphological changes in the smear of rodents. This stain,
however, in the study of human smears, did not give satisfactory results.
After using various staining fluids, I succeeded in developing a simple stain, consisting of a combination of haematoxylin, eosin, and waterblue, which seems to offer certain
advantages for the general staining of vaginal smears. This
secures a sharp outline of the various cell types with a variety
of shades from an intense blue to an eosin—red. The cornified
cells, which are almost constantly present in the vagina, take
an intense eosin color and are sharply differentiated. On
the other hand, cells derived from the deeper vaginal layers,
which are free from cornification, are stained strongly blue.
Partly cornified cells show various shades of purple. Superficial secretory cells, containing mucus, display a typically
characteristic bluish—purple tone. Leucocytes, as well as bacteria, are also sharply outlined. Eosinophiles show distinctly
red granulation. Mucus takes a bluish or a purplish shade. The contrasts are much sharper than with ordinary haematoxylin-eosin and the cornified cells are particularly differentiated. This is important for the Vagina, in which the cornification is Very pronounced, especially during certain periods of the cycle.
The exact method is as follows: Slides are first stained with Ehrlich’s haematoxylin for about 5 minutes, then carried into running water for at least a quarter of an hour. After this they are stained with a 0.5 per cent eosin for about 3 to 4 minutes, rinsed well in water, and finally stained for 1 minute
in a 0.5 per cent solution of waterblue. Following this they
are rinsed well in water and then carried through 50 per cent,
70 per cent, 80 per cent, 95 per cent, and absolute alcohol
into xylol and mounted in Canada balsam. The stain is permanent and does not fade easily.
Several other methods have been used, but none was as
satisfactory as the above. Many of the slides have been
stained with specific bacterial stains, especially with Gram’s,
for the study of the bacterial flora.
The twelve selected cases from which daily smears were
obtained were all normal and in good health. Racially, eight
were Americans, two Irish, one British, and one Italian. Ages
ranged from 19 to 28 years, one, however, being 35 years old.
Five were unmarried and seven, though previously married,
were not living with their husbands. They were of average
height and their weights ranged from 100 to 152 pounds.
Smears were taken daily, except on holidays, for 2 to 3
months, beginning February 9, 1925.
In addition to these, many smears have been examined
from other cases. The largest number has been taken from
one individual (our special case), covering about fifteen
periods and studied at different times since 1920. A number
of smears was also obtained from sixteen operated cases at
the Woman’s Hospital. These were patients with slight
pathological disturbances, in whom laparotomy was necessary. Smears were taken before and after operation, and
the ovaries were examined during the operation in order to
ascertain the relation of the various smear stages to the condition of the ovaries and especially to ovulation. This has
been rendered possible by the helpful cooperation of the director of the Woman’s Hospital, Dr. George Gray Ward.
The total number of smears studied is 1022, arranged as follows:
First day of menstruation: 39 smears (13 from the special case and 26 from the other cases); second day: 39 (13 + 26); third day: -39 (13+26); fourth day: 39 (12+27); fifth day: 42 (13 + 29); sixth day: 43 (12 + 31); seventh day: 39 (13 + 26); eighth day: 41 (14+ 27); ninth day: 40 (13 + 27); tenth day: 42 (14+ 28); eleventh day: 41 (14+27); twelfth day: 43 (14—1— 29); thirteenth day: 46 (13 + 33); fourteenth day: 42 (13 + 29); fifteenth day: 42 (13 +29); sixteenth day: 39 (12 +27); seventeenth day: 41 (12 + 29) ; eighteenth day: 41 (13 + 28) ; nineteenth day: 40 (13 + 27) ; twentieth day: 38 (12 -1- 26) ; twenty—first day: 33 (13 + 20) ; twenty—second day: 35 (12 + 23) ; twenty-third day: 30 (13 + 17); twenty-fourth day: 31 (13 + 18); twenty-fifth day: 29 (12 -1- 17); twenty—siXth day: 20 (4 + 16); twenty-seventh day: 13 (0+ 13); twenty-eighth day: 7 (0+7); twenty-ninth day: 4 (0 +4); thirtieth day: 1 (0 + 1); thirty-first day: 1 (0 + 1); thirty—second day: 0; thirty-third day: 1 (O + 1); thirty—fourth day: 1 (0 -1- 1). Total, 1022 (326 + 696).
3. The Morphology of the Vaginal Epithelium as the Basis of Smears
Most of the cells present in the vaginal fluid are derived from the vaginal epithelium, and especially from its superficial layers. Cells lining the vagina are subject to continuous desquamation and may be constantly found in its lumen. Cells of the deeper layers are less frequently thrown offthese only appear en masse after an active denudation following a period of excessive growth. Such a phenomenon occurs in the human commonly after parturition, whereas in other mammals, as the guinea pig, it may recur periodically during oestrus.
The recognition and classification of the various forms of cells in the vaginal fluid offer difficulties owing to extreme variability within the types. The free cells of the vagina
cannot be directly compared with the cellular make-up of the vaginal epithelium. The latter consists of normal living epithelial cells of relatively uniform types, whereas the smear
is characterized by a large number of dead and highly modified cells displaying great variety in form and structure. Still further, cells belonging to the same type may show great differences in the various groups of mammals. The so-called type I cells in the guinea pig show entirely different appearances in closely related species, such as mice or rats. Therefore, a strict morphological classification of smear cells is hardly attainable, especially from the comparative point of view, without a proper consideration of general topographical relations.
The general structure and topography of the vaginal epithelium have certain peculiarities which seem common to all
mammals. There are at least two distinct types or rather extremes of structural architecture to be encountered in the
same animal, one associated with low, and the other with high
sexual activity. The first is characterized by an epithelium
consisting of only two zones, an inner one, basal or germinative, and an outer one, superficial. The cells of the superficial zone arise from the germinative layers and usually differentiate into secretory cells, producing a mucoid substance.
The two-zone condition, as illustrated in the guinea pig,
occurs before puberty, during dioestrus or anoestrus, and
generally at times of sexual inactivity.
Whenever the vagina is stimulated to growth by an increase
in sexual activity, the basal or germinative zone undergoes
considerable proliferation and gives rise to a third zone, the
outer basal or pavement cell zone, interpolated between the
germinative and the superficial. The cells of the third zone
are large, having a round or oval nucleus and are connected
by a net of intercellular bridges, whereas the germinative cells
are smaller, with a thin cytoplasmic coat around the nucleus.
The three zones may be found normally in prooestrus.
During oestrus, when sexual activity is more accentuated, the outer part of the pavement cell zone undergoes a progressive cornification, the extent of which differs greatly
among the various mammalian species. In the rodents this cornification is extensive and a very distinct fourth zone appears, the intermediate or cornified zone, which is placed
between the outer basal and the superficial. This fourth, or cornified zone, is derived chiefly from the third zone, Whereas the second (superficial) and the third (pavement cell) arise directly from the germinative zone.
All cells of the second, third, and fourth zones are so far differentiated as to be unable to give rise to other types of cells. The old idea of an actual metaplasia occurring in these
various types of differentiated cells is untenable. The cornification of the outer basal zone cells is a process of degeneration such as occurs in the skin. Only the undifferentiated
germinative cells retain the ability to undergo differentiation either into a secretory form (superficial mucous type) or into a supporting form (pavement cell type). The most differentiated superficial cells cannot change into any other type, whereas the outer basal cells may, under certain conditions, undergo cornification or a marked vacuolization. The process of cornification may be considered as a protective adaptation brought about by the formation of a peripheral sheath, consisting of dead cornified elements. The vacuolization of the outer basal cells may be due to a secretory function (formation of glycogen) or to a degenerative process.
Cells belonging to the three differentiated zones are partly
or totally eliminated when the destructive phenomena set in
at the end of the sexual or the gestation period. The extent
of vaginal denudation differs greatly in different species. In
the guinea pig the elimination of the three zones is practically
complete after each sexual period.
In the human, the study of the vaginal epithelium and especially of its cyclic changes had not been actively taken up until very recently. In 1925, Stieve published a study on the vaginal changes during pregnancy. He found an extensive proliferation of the vaginal epithelium from its germinative zone chiefly during the first months of gestation and a gradual increase in the number of cells and layers of the epithelial Wall. There was also a noticeable increase in the size of the cells and of their nuclei. With the onset of parturition degenerative changes appeared and all the excess growth material was eliminated. He also studied a few normal women and found a considerable increase in the height of the epithelium during premenstruum, but not so extensive as during pregnancy.
Dierks, in 1927, gave a detailed account of conditions prevailing in the normal vaginal epithelium of women. He described three distinct zones: the basalis, the cornified zone, and the functionalis. During menstruation the two upper zones (functionalis and cornified) are cast off and only a thin basalis remains. Gradually the basalis grows thicker and, at about the seventh day from the onset of menstruation, a new cornified zone and a new thin functionalis appear. This new functionalis, which originally consists of only 2 or 3 cell layers grows actively up to the premenstruum, reaching a considerable thickness of about 155 p (30 to 34 cell layers). Accompanying this there is increasing hyperaemia, reaching its peak at menstruation.
The cells of the cornified zone were flattened and had
shrunken and pyknotic nuclei. The cells of the functionalis
were rather large with round nuclei. During the last days
of premenstruum the functionalis cells displayed extensive
karyopyknotic and chromatolytic degeneration extending
deep toward the cornified zone. Dierks concludes that there
is a distinct cycle and a periodic cornification in the human
Vagina.
In the same year Puccioni (’27), after a thorough study of several specimens of human Vaginae, arrived at similar conclusions. He found that during the intermenstrual period
the regeneration was completed. The vagina was covered by
a basal zone of cylindrical cells and, outside this, by one or two layers of cuboidal cells and by Various layers of pavement cells, the most superficial being almost entirely cornified. During the early part of the premenstrual period the cellular proliferation of the basal layers continued, resulting in a uniformly elevated epithelial lining, consisting of well-preserved cells up to the cornified zone. During the latter part of pre-menstruum the proliferation ceased and the desquamation of the stratum corneum as well as the degeneration of the superficial strata began. During menstruation there was a progressive desquamation of the epithelium.
Adler, in 1928, also described the development of the functionalis and the cornified zone in the human vaginal epithelium. He found a considerable increase of the functionalis during the second part of the intermenstrual period. During the first week after the onset of menstruation there was no functionalis present." Its growth, as well as the differentiation of the cornified zone, began with the second week.
In pregnancy the functionalis reached a Very high development, which might be used as a sign in diagnosing the condition. After parturition it gradually degenerated and disappeared. In girls and p0st—climacteric women no functionalis was present.
Like most investigators, Adler emphasizes the great Variability in the extent of the growth and degeneration processes.
This variation prevails not only among different individuals,
but also in any given person. Specimens taken from two
neighboring parts of the vaginal epithelium may often show
structural differences. The differentiation of the cornified
zone is particularly unstable. In one and the same section,
within a very small area, one may find spots with a distinct
cornified zone, and other places where there is not the slightest indication of it. The development of the functionalis is
especially variable during the first part of the inter menstrual period, its growth being more constant in premenstruum. The exceptional variability prevailing in the structure of the vaginal epithelium has been largely responsible for certain divergences of opinion among investigators.
Lindeman, in 1928, found irregularity in the appearance of the various zones of the vagina. A naked basalis (low) was observed at various times, not associated with menstruation (1, 3, 5, 7, 10, 11, 12, 15, 16, 17, 19, 25, and 27th day). A two-zone epithelium (basalis-functionalis) was also found at different times, independent of the menstrual cycle (1, 2, 4, 12, 17, 20, 21, 24, and 28th day). The three-zone epithelium (addition of an intermediate cornified zone) appeared also without a definite relation to the cycle (1, 4, 8, 12, 13, 16, 18, 23, 26, and 27th day). The cornified zone was found intra-epithelially (no more than five cell layers from the surface) and superficially, when the funetionalis was sloughed off. Lindeman concludes that the various constructive and destructive processes which alternate in the vaginal epithelium are evidently independent of the menstrual cycle.
N iirnberger, in 1928, found no cyclic periodicity. He contends that, contrary to Dierk’s (’27) description, the intra-epithelial cornified zone may be present during menstruation, whereas it may be entirely absent in premenstruum.
Pankow, in 1928, adheres to Dierk’s views of a cyclic
rhythm, but does not consider it as regular and typical. In
one case he saw a distinct three—zone arrangement at the
seventh day; the outer zone (functionalis) showed a partial
desquamation. At the twelfth day he found the cornified zone
more distinct and a much more intense desquamation of
the functionalis, which was altogether absent from certain
places. On the twenty-second day the middle zone was well
developed and the functionalis showed a more extensive desquamation. During menstruation the denudation process is
more pronounced, extending partly into the cornified zone.
He places the early development of a new functionalis a
little later than Dierks: after the end of the menstrual flow.
In one case, even at the thirteenth day, the basalis was practically naked, covered only partially by a newly developed
functionalis. Pankow also believes that the elimination of
the functionalis in individual cases may occur during the
interval as well as during menstruation. The examination
of the vaginal smears gave additional support to this view.
Samples taken during the interval had shown much desquamation. In one case only, he observed a cornified stage
(Schollenstadium). He concluded that the human vagina
does not undergo the same changes as the vagina of the rodents. He also found that in a large number of cases there
was an increase in the extent of bacterial contamination (Verschlechterung des Reinheitsgrades) shortly before or during menstruation.
In discussing Pankow’s report, Dyroff stated that he examined vaginal smears of normal women and found an increased epithelial desquamation before, during, and after
menstruation, whereas during the interval there were fewer epithelial cells. His observations are in support of the View that there is a cyclic return of proliferative and desquamative processes in the human Vagina, though less pronounced and less typical than in the uterus.
Stemshorn, in 1928, found the three-zone arrangement especially in the interval and, in a few sections, shortly after the onset of menstruation. On the other hand, he was unable
to observe clear differentiation of the three zones immediately
before the period, as Dierks described. The intermediate
zone was not considered by him to be definitely cornified and
was interpreted rather as a thickening of the epithelium (Verdichtungszone). His conclusions were contrary to the conception of a typical vaginal rhythm.
Smith, in 1929, described certain specimens of human vaginae which showed a typical three-zone arrangement. He also noticed and emphasized, like others, the extreme variability in the appearance of the cornified zone.
Dierks, in the same year (1929), tried injections of follicular hormone on an ovariectomized woman and found a proliferation and hyperplasia in the Vaginal epithelium with the
appearance of all three zones. He concludes that the vaginal
cycle depends upon the ovarian secretions. He further observed that the growth of the vaginal epithelium was not
entirely uniform, and this accounts for its great developmental variability.
A general review of the human studies offers considerable
evidence in favor of the existence of a vaginal rhythm. The
periodic return of a premenstrual growth and of a menstrual
destructive process, as well as the appearance of an intermediate cornified zone can scarcely be disputed. On the other
hand, it is unlikely that a complete denudation of the cornified
zone or even of the functionalis occurs at each menstruation,
since most investigators find both these zones present at practically every stage of the cycle. The existence of a vaginal epithelial rhythm, as claimed by Dierk, seems to be fundamentally true, but his assumption that the functionalis and the cornified zone undergo a total elimination and a regeneration during each menstrual period is not supported by the
findings of others.
The three zones described in the human as basalis, cornified zone, and functionalis correspond topographically to the three zones described in the guinea pig as basal (inner and outer), cornified or intermediate, and superficial or mucous. The separation of the basalis into two zones, an inner basal or germinative and an outer basal or pavement cell zone, may
not seem justifiable to those who have studied the morphology of the human vagina in sections. Yet, in human vaginal smears, one may find, under various conditions, cells corresponding to those from a well—differentiated outer basal zone. Such cells appear very regularly and in abundance during post-partum (figs. 27, 28, 30). It seems evident that a suitable description of the vaginal smear changes in the human as well as in the lower mammals should be based on a four-zone classification and a subdivision of the basalis into an inner (undifferentiated) and an outer (differentiated) zone.
4. The Vaginal Smear Changes During the Various Phases of the Human Cycle
Based on the following description and from comparative considerations, I have adopted a general division of the human female sex cycle into four fundamental phases or periods: First, the menstrual or congestive phase, extending from the first to seventh day after onset of menstruation. This is characterized by various symptoms of congestion and a bloody discharge. Second, the ripe follicular or copulative phase, from the eighth to the twelfth day, during which the pre-ovulative ripening of the large follicles and the copulative morphological reactions occur. This is closely followed by the ovulative movement, which in normal typical cases usually appears at the twelfth to thirteenth day, and is characterized by the follicular rupture. Third, a post.-ovulative or proliferative phase, extending from the thirteenth to the seventeenth day. During this period the proliferative phenomena are more uniformly and more typically expressed. Fourth, the growth and secretory or pre-menstrual phase, from the eighteenth day to the onset of the next menstruation. This is especially characterized by higher functional and growth phenomena.
The more detailed analysis and interpretation of this classification will be given in a following section.
In considering the cellular make—up of the vaginal fluid,
one should keep in mind that, with the exception of some of
the leucocytes, all other cells contained in the lumen of the
vagina are desquamated, dead, and partly deformed elements.
Their description becomes difficult on account of extreme
variety in form. An effort to describe and classify as many
of the Vaginal smear cell varieties as possible resulted in the
recognition of many different cell forms. A. detailed descrip-
tion of these would be of little theoretical or practical interest
and far beyond the scope and plan of this work. Only such
types or groups of cells are described in this paper as could
be considered rather regular and typical in their appearance
and form.
The topographical origin of the various cells encountered
in the vaginal smears is not always clear. It is practically
impossible to link every cell with the zone from which it arose.
One may, however, draw certain conclusions in regard to the
original topographical relations of the various cells encountered in the smear by their general conformation and their
staining reactions.
The cells of the intermediate zone are characterized by
their cornification and the pyknotic condition of their nuclei
(figs. 47, 49 to 54; also figs. 32 to 34). They are, as a rule,
large squamous, flat, or partly folded cells. Sometimes they
have an elongate or elliptic form (fig. 54). When slightly
cornified, they may have a purple or pale bluish color (figs.
47, 48). The nucleus is pyknotic and ordinarily stained dark
with haematoxylin. It often shows fragmentation with a number of irregularly shaped granules dispersed into the cytoplasm (fig. 47). With the progress of cornification the nucleus may gradually disappear (figs. 52, 53). Totally corniffied cells become irregularly folded and are stained intensely red with eosin (figs. 60, 78). They are very similar to the typical eosinophilic cells found in abundance in the vagina of rodents during oestrus.
The cells of the outer basal or pavement cell zone are
smaller and more compact, usually rounded or oval, with a
relatively large and slightly oval nucleus and a rather dense
cytoplasm, which stains deep purple or blue (figs. 61 to 67).
Some of these cells are invaded by leucocytes (fig. 68) similar
to the type III cells in the rodents.
During post—partum, typical outer basal cells are found.
They are round or oval, sometimes elongate or navicular, and
have a large round or oval nucleus (figs. 82, 83, 84). A large
peri— or paranuclear vacuole is one of their characteristic
features. They also show vacuolization within the cytoplasm,
and are stained intensely blue. Numerous outer basal cells
are also present in the post—menopause smears.
Cells belonging to the inner basal zone cannot be easily identified in vaginal smears, being insufficiently differentiated. They have a relatively large, normal nucleus and a very small amount of cytoplasm. Such cells may appear in smears, but their definite identification. is very difficult.
The cells of the superficial zone are more differentiated,
more variable in size and form, and more deformed, depending upon the extent of differentiation, growth, or degeneration of the superficial zone. Most of the superficial cells
are of the ordinary squamous type wi.th flattened or partly
folded form and irregular outline (figs. 36 to 40). Their
nucleus varies greatly in size and is often folded or irregularly shaped. Sometimes it stains faintly, as if it were in a
process of gradual resorption (fig. 37); at other times it is
surrounded or adjoined by a vacuole (fig. 38). The usual
color of the cells varies between purple and blue. Under
certain conditions, some of the cells are greatly deformed and display very irregular and atypical forms and partial fragmentation (fig. 59). Some of the cells are invaded by bacteria (fig. 58). Others contain coarse cytoplasmic granules, some in contact with the nucleus (figs. 56, 57).
Many cells have higher differentiated forms and their topographical relations to the various zones are somewhat uncertain. They may be elongate, elliptic, or navicular (figs. 41,
44, 45, 46). Their nucleus is shrunken, slightly elongate, and
lies eccentrically, often close under the cellular surface. The
interior of the cell is very scantily supplied with cytoplasm,
which is often vacuolated. Its periphery is covered with a
heavy membrane. Some cells of this type have a round or oval form (figs. 42, 43).
A very typical and characteristic differentiation of navicular cells occurs during advanced stages of pregnancy (figs.
72 to 76). The nucleus of these cells is large in relation to
normal cells and their contour is heavier (compare with figs.
42 to 46). Their coloration is dark purple or blue.
The human vagina is continuously exposed to infection by
bacteria and always contains a rich and varied bacterial flora.
Due to the great variety of bacterial forms found in the vagina, it is very difiicult to give a detailed account of them.
Only certain typical forms, such as the bacillus vaginalis,
may be often identified. Aside from this bacillus, there are
many other types of bacilli and Various forms of cocci, either
gram-positive or negative. Certain bacterial forms are most
frequently encountered in pathological cases, especially in
pathogenic infections. Some thin and long and some comma-
like bacilli, or prominent chains of cocci, evidently belong to
this class. Spores of parasites have been also observed, usually in pathological conditions.
A. Vaginal fluid changes olnmng the menstrual phase
This phase is prominently characterized by the appearance of the menstrual bleeding. Though the definition is somewhat uncertain, we speak of the first day of menstruation as the day when the bloody discharge becomes externally visible.
Of course, erythrocytes, indicating the presence of blood, may
be found in the vagina sometimes before the actual onset of
the menstrual flow. Theoretically, it might seem entirely cor-
rect to consider this as evidence of beginning flow, and yet
any attempt to modify the prevailing conceptions in this mat-
ter would lead only to confusion. Menstruation has been
primarily considered as an external sign and should be main-
tained as such.
Samples of vaginal fluid taken from the vagina during the
menstrual phase have a bloody or yellowish appearance. In
most cases the visible bleeding stopped at about the fourth
day. Erythrocytes, on the other hand, were generally found
in abundance up to the seventh day, after which there was
a rather abrupt decrease (fig. 2a).
Fig.2a Showing the percentage of cases in which erythrocytes have been found in the vaginal smears during the consecutive days of the'sex cycle (solid line) and the percentage of cases in which the erythrocytes were abundant (broken line). The absolute figures are given under each day. For total number of cases studied on each day see pages 537 and 538.
The largest number of erythrocytes is generally found at the second and third days. Many erythrocytes, especially toward the end of the bleeding, are deformed, vacuolized, or
fragmented. Others entirely lose their hemoglobin and become pale and colorless. It requires experience and the use of a high magnification in order to recognize some of these
modified forms. The longer they remain in the Vagina the
more likely are they to be deformed. Since the menstrual
blood is normally extravasated in the uterus, and descends
into the vagina mixed with mucus and other detriment, one
would expect to find a high percentage of abnormal erythrocytic forms during the advanced stages of menstruation. At
times the smear seems fibrinated (fig. 4). This usually occurs
at the beginning of the period, before the full onset of the
menstrual flow.
Erythrocytes in the vaginal smears are either isolated or in piles and groups (figs. 5, 6, 7). In rare instances they are seen within the body of cells. An actual phagocytosis of erythrocytes by leucocytes is a very uncommon occurrence during the menstrual phase, though such a phagocytosis may occur at other times.
The general consistency of the vaginal secretion is semifluid mucous. The amount increases toward the second and
third days as it becomes more fluid and bloody in character.
Toward the end of this phase it decreases in quantity and
becomes more and more mucous and somewhat denser. The
secretion of mucus undergoes great fluctuations, but is
usually fairly abundant, especially during the first 4 to 5 days.
The mucus, in stained smears, is rather diffuse and spread in
the form of fibers, taking a blue-purple coloration (figs. 5, 6, 7 ). Its distribution through the smear is irregular, consisting of thinner and heavier spots.
The number of leucocytes, though changeable. and seemingly atypical in all human smears, shows on the average
certain periodic variations during the various phases of the
cycle, indicating the existence of a rhythm. The polymorphonuclears are generally numerous during the menstrual phase. An actual leucopenia is rather rare (fig. 2b). Most of
the leucocytes have a normal appearance and well-defined
cytoplasm. Their activity is evidenced by the phagocytosis
of large numbers of bacteria. They often appear in groups
or aggregations, more commonly around the third or fourth
day. Their color is usually pale or slightly bluish. A certain number of leucocytes show signs of degeneration and disintegration.
Fig. 2b Showing the total number of cases (solid line) and the percentage of cases (broken line), in which typical leucopenia has been observed in the vaginal smears during the consecutive days of the sex cycle.
In some typical cases a marked increase in the number of
leucocytes occurs after the third and up to the seventh day
(fig. 9). This is highly suggestive of a true leucocytic exodus,
as evidenced in other mammals in early metoestrus. The appearance of the leucocytes and their compact form are rather indicative of recent migration into the lumen of the vagina. Many of them show a slightly dark bluish taint. It is a common occurrenee to see some enclosed within the body of cells (fig. 9). The erythrocytes are on the decrease during this particular stage and are, usually, outnumbered by the leucocytes.
In many cases the expression of this condition, like every
other one in the human, is extremely atypical. The smears
appear rather dry and diffuse, with poorly defined outlines
of cells and leucocytes, all embedded in rich masses of bacteria (fig. 10). The leucocytes are then poor and highly
degenerated.
The mononuclears are, as a rule, prominent through the entire menstrual phase——more so than at any other period. In addition to their increased number, they often show a certain enlargement in the size of their cytoplasm and nucleus, which frequently becomes kidney—shaped. Some show vacuolization of the cytoplasm and others display a certain phagocytic activity by assimilating detritus, dead polymorphonuclears, or bacteria. The highest activity, especially of the larger mononuclears, seems to be during the first 4. days.
Degenerating mononuclears are also present. A rather
specific form of degeneration consists in the granular fragmentation of the nucleus and its progressive resorption. Such
mononuclears acquire a characteristic form and may often
appear in large numbers and in groups or aggregations. They
are more frequently found during menstruation than during
the other phases of the cycle.
Lymphocytes are numerous during the menstrual bleeding,
reaching their maximum at about the third day, after which
they gradually decrease in number. Plasma cells may be
seen, especially in the early stages of menstruation. Eosinophiles have been repeatedly observed. They were relatively
numerous in only six cases, at the second, fifth, and sixth days.
The relative number of epithelial cells in the vaginal fluid is not altogether uniform during the entire menstrual phase. The number of epithelial cells is larger during the first day, then decreases with the increase in bleeding up to the third or fourth day, after which time such cells tend to increase again. Most of the cells present during this period may be considered either as superficial or as intermediate cells (figs. 5, 6, 7). They are squamous cells with a medium-sized, shrunken, or with a pyknotic nucleus. As menstruation progresses the intermediate cells tend to become more prominent. These are cells with a pyknotic nucleus and their cytoplasm often shows early signs of cornification. One such cell is illustrated in figure 48.
Another cell type quite characteristic of this period shows
a faintly stained nucleus, as though the nucleus were in a
process of resorption (fig. 37). Two of these cells are illustrated in figure 6 along with an intermediate cell. The resorption of the nucleus is a degenerative process, which does
not seem to have any particular significance. Such cells are
often found in large numbers in certain pathological cases,
after operations, and more frequently after an intra-vaginal
treatment with radium. The nuclear resorption is probably
a sign of progressive cellular necrosis.
The majority of the ‘intermediate cells do not reveal definite cornification. There are present, however, a number of
more or less typically cornified cells, varying in individual
cases. Enucleated eosinophilic cells may be found either isolated or in small groups.
Cells from deeper layers, evidently belonging to the outer
basal or pavement cell group, are often present during
thisphase (as in figs. 61 to 66), but n.ot in all cases. They
are smaller and more compact than the ordinary cells and
have a. denser cytoplasm and a more normal nucleus. Their
form is usually round or oval. ' A
The relative scarcity of such cells indicates that, as a rule, the destructive processes during menstruation do not reach the outer basal zone, being mostly limited to the superficial
and to the outer layers of the cornified zone.
Another cell form characteristic of the ‘menstrual phase often appears in large, irregular groups (fig. 8). This is a small cell without sharp outlines, with a very small amount
of cytoplasm and a relatively large nucleus. Cell aggregations of this type have been observed 32 times in all, most
frequently and typically at the second and third day, when
the bleeding and the destructive processes reach their maximum. They also have been observed less frequently during
the first, fourth, fifth, sixth, and seventh days. At other
times than the menstrual phase they only appeared in four
instances in a rather atypical way, once at the eighth, once
at the ninth, once at the twelfth, and once at the twenty-first
day. In pathological cases I found them oftener i11 metrorrhagias and menorrhagias, where they may be considered of
diagnostic value.
The general conformation of these cell groups shows a
resemblance to fragments of uterine compacta, undergoing
desquamation. When the bleeding is intense, such fragments
may be washed down into the vagina by the menstrual blood.
The fact that they often appear in metrorrhagias is in favor
of this interpretation. One cannot, however, exclude the pos-
sibility that they may be fragments of the inner basal (germinative) zone of the vagina, which are expelled u11der severe
desquamative conditions.
Denudated uterine tissues usually decay before reaching
the vagina and dissolve into a cellular detritus, which is
phagocytized. Such bluish detritus of dirty appearance may
be ordinarily seen in the va.ginal smears of this period.
Toward the end of menstrual bleeding, usually after the
third day, when the peak of the destructive activity has been
reached, and the leucocytic exodus made its appearance, several cells are shown with their bodies invaded by leucocytes
(fig. 9). This occurrence suggests the so-called stage III in
the guinea pigs, during which a large proportion of epithelial
cells containing leucocytes and a high leucocytic exodus predominate.
Such cells are evidently derived from the subcornified layers either of the intermediate or of the outer basal zone and their presence in large numbers may be considered as an indication that the cornified zone has been, to a certain extent, removed.
In the guinea pigs and other mammals, in which the cornification process is extensive during oestrus, and the under- lying outer basal zone well differentiated, the type III cells are almost exclusively of the outer basal type. ‘ In the human, on the other hand, both zones—cornified and ‘outer basal— are incompletely differentiated during menstruation and their cells less typically formed. The cells invaded by the leuco- cytes are usually atypical. Only occasionally one finds such cells having the outer basal type (fig. 68), and closely resembling the typical III cells.
The penetration of these cells by leucocytes is somewhat
puzzling. Are the leucocytes penetrating the epithelial cells
actively in order to phagocytize them, or do they become passively included within them”! The acceptance of phagocytic
activity on the part of the epithelial cells can hardly be considered, since it is evident that, in general, the cells and their
nuclei suffer from this parabiosis and not the leucocytes. The
nuclei of such cells often show distinct signs of phagocytosis
and dissolution caused by the leucocytic invaders.
The explanation which I am inclined to give of this occurrence is based on two facts: when a solid and intact cornification zone is present in the vagina, the leucocytes are
hindered in their migration toward the lumen, and often
accumulate in large numbers just underneath the cornified
sheath (Stockard and Papanicolaou, ’17). Moreover, the cells
of the sub-cornified outer basal zone are usually equipped
with intercellular bridges. It is therefore possible that, when
the degeneration phenomena begin and the cells lose their
intercellular connections and retract their impaired bridges,
the crowding leucocytes lying close to them penetrate some
of the cells.
Many atypical cell forms may be found in the smears during the menstrual phase. Some show‘ vacuolization of their
cytoplasm and some are plasmolyzed. In others the nucleus
is surrounded or adjoined by a large vacuole (as in fig. 38). Certain cells have a somewhat naviculated, form (as in figs.
44, 45, 46). Whenever these cells are present in abundance,
they seem to indicate the presence of a somewhat hypertrophic condition. Several cases were characterized by the persistence of such cells not only during the menstrual phase, but
throughout the whole cycle. Other cells show enlargement,
elongation, or fragmentation of their nuclei. Cells with various types of granules dispersed within the cytoplasm may
also be observed (as in fig. 56). In some of these the granules seem to merge from the nucleus as in a budding process.
Various degenerated cells, such as the ones illustrated in
figure 59, are found, especially in dry smears with poorly
preserved cells. Others are invaded by and some practically
loaded with bacteria. (as in fig. 58).
The various morphological changes occurring during the
menstrual phase may be grouped into three distinct vaginal
smear stages, succeeding one another more or less typically:
first, the ‘early menstrual,’ characterized by the predominance of epithelial cells over the erythrocytes and leucocytes;
second, the ‘mid-menstrual,’ showing excessive bleeding and
a large increase of erythrocytes; and third, the ‘late menstrual’ stage, characterized in typical cases by a leucocytic
exodus and its related phenomena. This stage, which may
be considered as a pseudo-III stage, marks the end of the
menstrual desquamative processes.
The number of bacteria usually decreases with the onset
of the abundant blood flow. During the second and third
day, when the bleeding reaches its maximum, bacteria are not
very conspicuous. Later, toward the end of this phase, their
number increases, sometimes to a very high degree. This is
especially noticeable in atypical dry smears, in which the
bacterial flora appears to be extremely rich (fig. 10). It is
probable that the constant flow of blood through the vagina
during the first few days aids considerably in the removal of
bacteria and checks their growth. One finds that in dry
smears there is always a high bacterial content. Very often,
and especially during the bleeding, the bacteria form small
groups or conglomerations.
The typical form of bacillus vaginalis has been found most
frequently toward the end of the menstrual phase. Also other
types of long bacilli have been seen at this time. During the
first days of bleeding there is a relative prevalence of cocci.
A detailed description of the morphological characteristics
of vaginal smears in man and monkeys during menstruation
has been given by earlier investigators. Pouchet (1847) long
ago found in women an enormous number of erythrocytes at
about the third day, many leucocytes (globules muqueux), and
small epithelial fragments. The erythrocytes were often arranged in piles; some were deformed. Mucus was usually
noticed with the blood. During the later part of the menstrual phase (his period of cessation) he noticed a decrease
in the menstrual flow and in the number of erythrocytes, which
became scarce. A fairly large number of leucocytes and
pieces of epithelial tissue were present.
Corner (’23) described the menstrual smear of Macacus as consisting of erythrocytes, epithelial cells, and leucocytes. In the first half of the inter-menstrual interval, he found relatively few epithelial cells and many leucocytes. The post-menstrual smear was thin and scanty.
Ramirez (’28) found the leucocytes scarce in the beginning
and increasing in numbers toward the end of menstruation.
Shortly after menstruation, there were many polymorphonuclear neutrophilic leucocytes. With the end of menstruation, epithelial cells showing small pyknotic nuclei exceeded
gradually in number those with larger nuclei. Cells encumbered with leucocytes, as observed by us especially toward
the end of the menstrual phase, have been occasionally seen
by Moser ( ’28), though not in relation to any definite stage.
B. The vaginal fluid during the copalative phase
In most mammals the time of copulation is well fixed within what is called the period of heat. The examination of the vaginal fluid during this particular period reveals typical and characteristic changes. As a rule, there is an increase in the
(insert page 40 onwards here)
Plates
Plate 1

4 Second day of menstruation. Show extreme fibrination. 0c. 3, Obj. 8, B. 19.
5 Third day of menstruation. Shows typical condition during early menstrual phase. Most of the small dark cells are erythrocytes. Some leucocytes are also present. 0c. 6, Obj. 16, B. 23.
6 Enlarged from figure 5. 0c. 6, Obj. 8, B. 19.
7 Enlarged from figures 5 and 6. 0c. 6, Obj. 8, B. 30.
Plate 2

8 Third day of menstruation. Shows aggregations of small compact epithelial cells. 0c. 3, Obj. 8, B. 19.
9 Seventh day. Shows typical Ieucocytic exodus and cells invaded by 1eucocytes. 0c. 6, Obj. 16, B. 23.
10 Ninth day. Shows extreme bacterial growth. 0c. 3, Obj. 8, B. 19.
11 Tenth day. Early copulative phase with extreme mucous Secretion. 0c. 6, Obj. 16, B. 23.
Plate 3

12 Tenth (lay. Early copulative phase with mucus and some leucocytes. Oc. 6, Obj. 16, B. 23.
13 Twelfth day. Typical copulative smear with leucopenia. Compare with figure 32. 0c. 6, Obj. 16, B. 23.
14 Twelfth day. Enlarged cells of copulative type. 0c. 6, Obj. 8, B. 19.
15 Thirteenth day. Typical 0v11la.tive smear. Numerous leucocytes are present. Compare with figure 33. 0c. 3, Obj. 8, B. 19.
Plate 4

16 Fourteenth day. Post-ovulative type of smear. 0c. 3, Obj. 8, B. 19.
17 Fifteenth clay. Post-ovulative smear with continued high cornification and Ieucopenia. Compare with figure 34. 0c. 6, Obj. 16, B. 23.
18 Twenty-third day. Typical premenstrual smear. 0c. 6, Obj. 16, B. 23.
19 Twenty-eventh day. ‘Late premenstrual type of smear. 0c. 3, Obj. 8, B. 19.
Plate 5

20 Twenty-seventh day. Late premenstrual smear. (Same as i11 figure 19.) 0c. 6, Obj. 8, B. 30.
21 Early pregnancy. Thirty-fifth day after last menstrual date. 0c. 3, Obj. 8, B. 19.
22 Typical smear of pregnancy. Sixty-fifth day after last menstrual date. 0c. 6', Obj. 16, B. 23. -
23 Same as in figure 22. 0c. 3, Obj. 8, B. 19.
Plate 6

24 Late pregnancy. Smear taken 5 days before normal delivery. Shows unusual occurrelme of extreme leucopenia. O0. 3, Obj. 8, B. 19.
25 Typical smear of pregnancy. Sixty-fifth day after last menstrual date. (Same as figures 22 and 23). Shows typical navicular cells highly magnified.
26 Pregnancy: one hundred and fifty-second day. Shows cells of ‘oyster shell’ type.
27 Sixth day after normal delivery. Shows typical cells of the postpartum phase. 0c. 6, Obj. 8, B. 30.
Plate 7

28 Twelve hours after normal delivery. The small cells are erythrocytes and leucocytes. 0c. 3, Obj. 8, B. 19.
29 Stage III of guinea pig. The small cells are all leucocytes, Oc.3, Obj. 8, B. 19.
30 Same as in figure 28. Shows one typical cell of type III. The smaller cells are leucoeytes and erythrocytes. O0. 6, Obj. 8, B. 30.
31 Five days after delivery. Shows one group of large mouonuclears. The smaller cells are polyniorphoiiuclears and erythrocytes. 0c. 6, Obj. 8, B. 43.
Plate 8
Photomicrographs of human vaginal smears colored by hand as in stained smears, in order to show the relative number of cornified cells (stained red).

32 Twelfth day. Typical copulative smear with leucopenia. Same as figure 13.
33 Ovulative type of smear. Fourteenth day. Oc. 3,0bj. 8, B. 19.
34 Fifteenth day. Post-ovulative smear with continued high cornification and Ieueopenia. Same as figure 17.
35 Twenty-third day. Typical premenstrual smear. Same specimen as in figure 18. 0c. 6, Obj. 16, B. 23.
Plate 9
Drawings of various types of cells found in normal human vaginal smears.

- 36 to 60 Cells from human vaginal smears at different stages of the normal menstrual cycle.
Plate 10
Drawings of various types of cells found in normal human vaginal smears.

- 61 to 68 Cells from human vaginal smears at different stagesrof the normal menstrual cycle.
- 69 to 79 Characteristic types of cells found in human vaginal smears during pregnancy.
- 80 Normal mononuclears during menstruation.
- 81 Large mononuclears found in post—partum.
- 82 to 84 Cliaraeteristic types of cells found in postpartum.
Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - The Sexual Cycle in the Human Female as revealed by Vaginal Smears (1933). Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_The_Sexual_Cycle_in_the_Human_Female_as_revealed_by_Vaginal_Smears_(1933)
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G