Paper - An early stage of human implantation
| Embryology - 27 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Latta JS. and Tollman JP. An early stage of human implantation. (1937) Anat. Rec. 69(4): 443-463.
| Online Editor |
|---|

|
| Historic Disclaimer - information about historic embryology pages |
|---|
|
An Early Stage of Human Implantation
John Stephens Latta and J. Perry Tollman
Departments of Anatomy and Pathology, University of Nebraska College of Medicine, Omaha
One Text Figure And Three Plates (Fourteen Figures) 1937
Introduction
Some time ago a specimen of a uterus surgically removed from a sexually active woman was sent to the pathology laboratory for examination. It was found later that the endometrium contained an ovum in an early stage of development. Careful examination of the implanted tissues revealed no traces of embryonic disc, amnion or yolk sac. Nevertheless histological study indicated that the tissues were very well preserved and several interesting features of implantation were well shown. In View of the fact that only relatively few ova in situ in such an early stage of development have been described, we have thought that an account of the features presented by our specimen would prove interesting, even though necessarily incomplete.
Clinical History
A married white woman, aged 40, entered the University Hospital complaining of back ache, frequency, urgency and irregular periods. Her past history showed three spontaneous miscarriages and marked menstrual irregularities over a period of 8 years. The periods had been generally at 28day intervals, one to four periods occasionally being missed, until 11 months before the present specimen was obtained. Since that time they had occurred at 20-day intervals. A curettage 5 months before the present admission showed normal endometrium. The uterus measured 6 X 5 X 5 cm. The endometrium averaged 4 mm. in thickness and exhibited no gross changes.
Microscopic Preparation
After gross examination of the endometrial surface and before any fixation of the tissue, a thin section of it extending to and including some of the muscularis was cut out with a safety razor blade, fixed in formalin, dehydrated in alcohol, cleared in chloroform, infiltrated and embedded in paraffin. Inspection of the cut edge of this block of tissue had shown a small cyst about 5 mm. in diameter just beneath the endometrial surface. A sample section was cut from this block and stained routinely at the hospital laboratory with hematoxylin and eosin, Examination of this section showed that this cystic structure was a chorionic vesicle in a very early stage of implantation and development. The remainder of the block of tissue was immediately presented to the department of anatomy together with blocks of tissue removed from the endometrium on either side of the region from which the original block was removed, both of which were found to contain portions of the chorionic vesicle. The original cuts in the fresh endometrial surface had, therefore, cut directly through the embryonic membrane. Serial sections were made of each of these blocks of tissue, some of the sections being 10 [J and others 15 u in thickness. Most of the series was stained with Mayer’s hematoxylin and eosin, a few slides being stained with Delafield’s hematoxylin and azur II-eosin.
The sections cut from these three different blocks of mucosa were arranged as accurately as possible to make one nearly complete series through the implanted ovum. Vvhere the series from one block passed over into the adjacent one some tissue was lost, and there were undoubtedly slight changes in the plane of sectioning of the three pieces. At the beginning and end of each block incomplete sections were cut, all of which were mounted as part of the series. These factors made the determination of measurements rather inexact, although we believe that the measurements given are Very nearly correct.
General Description
Our first examination of the implanted ovum showed that it was completely covered by a layer of decidua 0.14 mm. thick after fixation. Unfortunately it was found that an artificial break had occurred, probably during manipulation of the fresh tissue, which extended through the covering decidua and the underlying chorion into the chorionic cavity. In addition and possibly as a result of this rupture of the vesicle there was found to be considerable distortion of the chorionic mesoderm, consisting principally of withdrawal of the mesodermal cores of the shorter villi and occasional breaks without displacement in the chorionic epithelium. In View of the necessary method of preparation and these evidences of distortion, we were not surprised, though greatly disappointed, that we have been unable to discover any traces of embryonic disc, amnion or yolk sac.
Regardless of the above-mentioned evidences of distortion, the chorionic vesicle appears to have been relatively undisturbed, having retained a practically spherical shape, and the chorionic-mucosal relationships have remained intact. The developmental state of the chorion and the size of its cavity, together with its position in and relations to the uterine mucosa, give ample evidence that this material presents a very early stage of implantation.
Measurements and Relation to Other Described Early Implantations
In order to more accurately determine the size of the implantation cavity and the limits of the embryonic trophoblast, several photographs were taken of sections through the center of the vesicle. Measurements were made from the photograph in which the vesicle was of greatest diameter. In this section there Was some displacement of the edges of ruptured chorion previously mentioned and overlying decidua capsularis (fig. 1). In determining the diameter of the vesicle in a plane perpendicular to the surface, measurements were taken from both of these broken edges. The average of these two measurements give a vertical diameter of 4.25 mm. The greatest diameter in a plane parallel to the endometrial surface as measured from the same photograph was 4.54 mm. Estimation of the diameter of the Vesicle in a line perpendicular to the plane of section was inexact because of the incomplete sections at adjacent edges of the three blocks of tissue. Our computation of this measurement from the sections gave a diameter of 5.5 mm. The external dimensions of the chorionic vesicle, including the trophoblast, and likewise the dimensions of the implantation cavity were approximately 4.25 X 4.54 X 5.5 mm. These values are quite similar (slightly less) than those computed by Stieve (’26) for ‘Ei Hugo’ and less than those given for the Mateer (Streeter, ’20) and the Von Herff (Von Spec, 1896) embryos, somewhat larger than for the embryo described by Kindred (’33) and very nearly the same as those given for the Grreenhill specimen ( ’27).

Fig.1 A section near the center of the implanted ovum. The enlargement of the uterine glands and the numerous papillary folds are clearly shown. Note the comparative absence of glands in the more compact tissue immediately surrounding the implantation cavity and also compactness of the glandular tissue in the region immediately underneath the ovum. X 18.
True villi were present, extending from all sides of the chorionic surface. These were considerably longer and slightly more complex in the basal and marginal zones than in the region of the covering decidua capsularis. In no instance, however, was there any great degree of complexity, there being few indications of branching and the largest villus measured being slightly less than a millimeter in length. Compared with other ova of similar stages of development, the villi of this specimen do not appear to be as long and complex as those of the Mateer, Carnegie no. 763 (Streeter, ’20), or the Goodwin embryo of Kindred (’33); but somewhat more complex than represented for the Greenhill specimen. In respect to chorionic development our case apparently most closely resembles the 1342--day-old embryo described by Stieve (’26), although his illustrations of this feature are inadequate for exact comparison.
Measurement of the diameter of the chorionic cavity was made only from a photograph of the largest section in planes parallel and perpendicular to the mucosal surface, the dimensions not including the mesoderm being 2.6 X 2.6 X 3.7 mm.
From the incomplete data available in the specimen here presented, we have concluded that it should be considered as a younger example of ova of group II, stage 4 of Streeter’s classification (’20) and class C1 of Grrosser’s classification (’24). Therefore the embryo associated with these adnexa, which was so unfortunately lost must have been in a presomite stage, probably exhibiting a primitive streak in the embryonic disc.
Uterine Mucosa
The mucous membrane in general was somewhat thickened and presented a11 appearance quite similar to the mucosa in the premenstrual phase, as observed by several others coincidental to early stages of implantation (e.g., Linzenmeier, ’14; von Miillendorff, ’21; Stieve, ’26; Streeter, ’26). The connective tissue stroma was very edematous and markedly infiltrated with polymorphonuclear leucocytes. The glands were very large and tortuous, with numerous papillary folds extending into their lumina (fig. 1). The glandular epithelium was columnar and had evidently been actively secreting, as shown by secretory globules extending into the lumen from many cells and the presence of secretory products within the glandular lumina (as described by Linzenmeier, ’14; Falkiner, ’32). The stroma appeared as columns between the vertically placed glands, and in the columns or septa were found many sections of small, thick-walled arteries. The stroma was less edematous and the glands more closely packed, with increased tortuosity and folding immediately underneath the basal zone of implantation (fig. 1).
Approaching the tissues immediately surrounding the chorionic vesicle the glands were found to have changed their direction so as to go around the area of implantation (Teacher, ’25; Greenhill, ’27). The ovum with its covering of decidua. was shown by the sectioned material to have extended into the uterine lumen behind the general surface. This covering decidua capsularis, as stated earlier, was complete and of a thickness of 0.14 mm. There was no fibrin plug, discontinuity of the covering epithelium (as in the ovum described by Tennant and Ramsay, ’34) (except where artificially ruptured in handling), or operculum of trophoblast cells or other evidence of the locus of the portal of entry as were found by Falkiner (’32), Stieve (’26), Grreenhill (’27) and Kindred (’33) in implantations of about the same age and described in detail by Teacher (’25) in younger stages. We did note in this case, as observed by von Mollendorff (’25) that the surface epithelium, as well as the underlying connective tissue cells over the capsularis, was much flattened.
In the zone immediately outside the trophoblast (the ‘Umlargerung’ zone of some German investigators) the connective tissue seemed more compact than elsewhere. Very few glands were encountered, and in those found there was little tortuosity and no papillary folds (fig. 1). A lack of uniformity in the height of the epithelial cells was encountered, although generally it was lower than in glands encountered elsewhere. In the glands nearest to the trophoblast, the epithelial lining was often incomplete and the cells somewhat piled up. From the shrunken appearance of the nuclei and the rather indefinite cytoplasmic staining, it is thought that this heaping up of cells was a part of a degenerative process. Occasionally cell strands extend through such breaks into the glandular lumen, the strands being traceable directly to the embryonic trophoblastic syncytium (fig. 3).
Detailed examination of the connective tissue in this surrounding zone revealed that the stroma here was quite edematous. There was possibly some increase in size of the connective tissue cells (Linzenmeier, ’14), but we could find no evidences of definitive decidual cells and stages leading to them such as von Miillendorff (’25) described in embryo ‘W0 ’ only in the marginal zone. The infiltration with polymorphonuclear leucocytes previously noted was more marked here than elsewhere, but we also found numerous pycnotic nuclei and other signs of degeneration of connective tissue cells (fig. 2). There was, however, no necrotic zone as found in earlier stages by Bryce and Teacher (Teacher, ’25), von Mollendorff (’21) and others. Strands of syncytial tissue extended for some distance into the maternal tissue from the general trophoblast boundary. Many of these consisted of large multinucleated masses with deeply staining cytoplasm characteristic of the embryonic syncytium bordering the intervillous space. Many of the nuclei of these trophoblastic elements were very pycnotic, indicating degeneration. These invasive elements gave little demonstrable evidence of ability to digest the mucosa proper (fig. 2). Most investigators of early stages of human implantation have described a marked vascularity of the surrounding maternal tissue. The present example is no exception in that numerous large endothelial lined sinuses were encountered in this zone in very close relationship to the outer limits of the trophoblast. There were numerous evidences in the sectioned material of the disruption of the endothelial lining of some of the spaces by extensions from the syncytial trophoblast, in a few cases the syncytium forming one wall of the sinus for a short distance. In such regions there was usually a direct communication between the vascular lumina and the intervillous space (fig. 4).
Chorion and Trophoblast
The chorionic vesicle which was enclosed by this specially differentiated zone of the endometrium had been ruptured in preparation, as stated earlier, and the lumen contained nothing except a few maternal blood corpuscles and two fragments of villi. The chorionic wall was made up of a thin layer of mesoderm covered by a double layer of ectodermal cells (trophoblast). The chorionic villi projecting from the wall were variable in length and complexity, the greatest development being in the basal and marginal zones. In no instance was there found any great degree of complexity in branching. The mesoderm had been withdrawn either partially or completely from the villous cores, causing the mesoderm lining the chorionic vesicle to appear thicker than it actually was and those villi themselves to appear as hollow ectodermal tubes, continuations of the basal trophoblast. From the distal ends of these true villi extended massive columns of trophoblastic cells, which, as they approached the surrounding mucosa, spread out and fused with distal ends of adjoining cell columns to form an incomplete layer of trophoblast cells (‘trophoblast shell’) (fig. 7) in juxtaposition to the surrounding maternal tissues. Usually the peripheral trophoblast lay directly upon the decidual tissue and was easily distinguishable from it by differences in staining reaction. The two opposing tissues were intermingled in certain regions where long streamers from the syncytial trophoblast extended into the decidua, including maternal blood vessels and glands (as described earlier). There were frequent interruptions in this outer layer of trophoblast, most or all of which formed communicating channels between maternal blood sinuses and the intervillous spaces.
Upon detailed examination of the two-layered trophoblast which clothed the basal chorion and the sides of the mesodermal villi, it was seen that the epithelial nuclei lying next to the embryonic mesoderm were comparatively small and regularly arranged, with chromatin knots evenly dispersed. In the basal chorion the cytoplasm of these layers was somewhat granular, but the cell membranes were usually heavy and distinct (fig. 5). Along the sides of the villi the cytoplasm of the cells coInp’osing this layer, the cytotrophoblast, was often entirely clear, with cell membranes correspondingly more prominent (fig. 6) and numerous mitotic figures were found in this layer, particularly along the sides of the villi.
Covering this layer and bordering the intervillous space, there was a syncytial layer of granular, deeply stained cytoplasm in which no cell membranes were demonstrable and nuclei were irregularly interspersed. These nuclei were usually very large and vesicular with prominent nucleoli and heavy nuclear membranes (fig. 5). Along the villi the nuclei were‘ sometimes quite regularly arranged, smaller and not so vesicular, resembling quite closely those of the internal cytotrophoblast (fig. 6). Occasional cells with very clear cytoplasm, evidently originating from the cytotrophoblast, were found in the midst of the syncytial layer (plasmatrophoblast).
From the terminal ends of the true villi thick columns of cells extended toward the end of the implantation cavity. Near the villus these cells were closely packed with no intercellular space. They were in direct continuity with the cytotrophoblast and presented the same appearances of clear cytoplasm, prominent cell membranes and nuclei with evenly distributed chromatin characteristic of the inner epithelial layer of the villus (fig. 8). Mitotic figures were occasionally encountered in this region. Proceeding further from the ends of the villi, the cells comprising these columns became more spread out; so that spaces were apparent between some of them. Other cells in this part of the column became separated from the general mass so as to lie in spaces; such cells having very indefinite, degenerate cytoplasm and either very faded or shrunken and pycnotic degenerate nuclei, giving the impression that. the spaces arise in the cell columns through degeneration of some of the cells. The cytoplasm of the remaining cells was quite stretched out and more deeply stained, cell membranes were rarely demonstrable and the nuclei much larger and more vesicular than in the cells nearer the villi. In many of these larger nuclei prominent nucleoli were observed. It was apparent that the cellular tissue here was intermediate between typical cytotrophoblast and plasma trophoblast. Some of the space fourid between the cells here had established connection with the intervillous spaces as shown by the presence of maternal blood cells in the former (fig. 9).
Covering most of the cell cords and separating them from the definitive intervillous space, there was a layer of typical syncytial trophoblast, a continuation of that covering the cytotrophoblast of the true villi. This layer was somewhat incomplete so that at some places the typical or transforming cytotrophoblast was directly exposed to maternal blood in the spaces in the trophoblast or the definitive intervillous spaces (fig. 7). From the covering syncytial layeriof the villi and cell cords there were frequent extensions of syncytial masses into the intervillous space. Approaching the surrounding decidua the amount of syncytial trophoblast increased markedly; so that we found no place in which the cytotrophoblast was in contact with the decidual tissue, an incomplete layer of syncytium intervening. These large masses of syncytium had sometimes apparently lost contact with the layer covering the villi and cell cords and lay as isolated masses in the implantation cavity. The nuclei in such masses presented great variety in size and distribution. Some were extremely large, vesicular and widely separated from each other, others quite small and very closely clumped together (fig. 10). In some of these syncytial masses near the basal side, spaces or vacuoles of great variety as to size and number were found. Nuclei of such syncytial masses were usually quite shrunken and pycnotic. Irregular projections from the surface of some masses were found in which the typical deeply stained cytoplasm had faded, groups of deeply stained granules being present (fig. 11). Some such degenerating masses containing no nuclei were apparently lying free in the intervillous space. There were also found large syncytial masses near or adjacent to the maternal tissue in which both cytoplasm and nuclei had almost completely faded out. The de generative forms of syncytium surely indicate a regressive condition and should be designated as ‘symplasmata’ as according to Linzenmeier (’14), Grosser (’12) et al. We are undoubtedly in this case dealing with the degeneration of excessive syncytium of the second generation, the resorption syncytium (Grosser, ’22; Fetzer and Florian, ’30). From our study in this specimen, in which the majority of the trophoblast exposed to maternal blood was syncytial in character, that transformation of cytotrophoblast into syncytium was dependent upon contact with maternal blood or decidual tissue, as predicated by Peters in 1899 (see Grosser, ’12). This transformation might further be considered as of the nature of a degenerative process Which, in the exuberant masses of syncytium, is carried on to complete disintegration.
It is, of course, the layer of syncytium immediately covering the mucosa that gives origin to the long radiating syncytial strands extending for considerable distances into the maternal connective tissue (fig. 2), disrupting glandular epithelium (fig. 3), and invading and partially lining maternal blood vessels (fig. 4) as described previously.
The mutual antagonism mentioned by Von Mollendorff (’21) in his study of the ‘Sch’ embryo is further shown by the evidences of degeneration in the syncytial strands penetrating the maternal connective tissue and by the pycnotic nuclei so frequently encountered in the cells of this connective tissue, as well as the disrupted glands and blood vessels. We found almost no fibrin in the surrounding zone, such as described and illustrated in embryos described by Greenhill (’27), Stieve (’26), Falkiner (’32) and many other investigators in younger embryos.
The absence of a necrotic zone in the uterine stroma, its penetration by syncytial tissues, together with the absence of any space around the penetrating syncytial elements indicate that ‘the ovum is entering upon a phase during which it will become anchored to the surrounding decidua.
Chorionic Mesoderm
Adequate study of this tissue was impossible because of the great distortions encountered which had occurred in preparation. Next to the chorionic cavity was a thin layer composed of spindle-shaped cells, arranged parallel to the chorionic surface and lying in a practically homogeneous matrix. The few evidences of fibers seen were considered to be finely drawn out cell processes. This layer was definitely set off from the chorionic cavity which contained no magma, but no complete surface layer of mesodermal cells could be demonstrated. The spindle-shaped cells possessed elliptical nuclei with finely divided and evenly distributed chromatin. Some small groups of three or four partially rounded up cells were encountered, containing spheroidal, vesicular nuclei with prominent nucleoli. Individual spherical cells were found lying free in the meshwork, containing large nuclei with chromatin collected around the nuclear membrane and definite central nucleoli, but otherwise clear. The cytoplasm of such cells was stained rather deeply with basic stain (fig. 12). Cells with quite long processes were seen containing similar nuclei. These were most numerous in the mesoderm adjacent to the cytotrophoblast.
The mesodermal cores had been to a great extent either partially or completely withdrawn (figs. 1, 7 and 13). Masses of mesoderm lying next to the epithelium in which the fusiform cells lay perpendicular to the surface undoubtedly represented mesoderm originally within the villi. When the mesoderm was only partially withdrawn from at villus it could be seen to have separated cleanly from the cytotrophoblast and in villi in which the mesoderm was intact there was a distinct line of separation between it and the overlying epithelium (fig. 13). We failed to find any evidence in this case to indicate that either angioblastic or mesodermal cells were arising from the cytotrophoblast as shown by Hertig (’35) in embryos of a similar stage and in macaque embryos. Cells comprising the villous mesoderm were of the same nature as those encountered in the basal mesoderm. Cells in various stages of rounding up to become free, possessed vesicular nuclei and nucleoli were quite numerous (fig. 14). Groups of three or four free cells with basic staining cytoplasm and large vesicular nuclei were sometimes found here also (fig. 15). Throughout the mesoderm, particularly in the villi, there were some strands of larger fusiform or stellate shaped cells with large vesicular nuclei with nucleoli and rather deeply staining cytoplasm which might be interpreted as the endothelium of beginning Vasculogenesis (figs. 13 and 15) (see Stieve, ’26; Hertig, ’35). (Considering the distortions encountered in the prepared tissues, we do not feel justified in drawing conclusions as to the nature of the single or grouped free spherical cells, although they have many features characteristic of primitive blood cells. Intermediate cell stages in the chorionic mesoderm indicate, surely, a mesodermal origin for both these free spherical cells and the endothelial ('1?) strands.
Literature Cited
FALKINER, N. M. 1932 A description of a human ovum, fifteen days old with special reference to the vascular arrangements and to the morphology of the trophoblast. J. Obst. and Grynaec. Brit. Emp., vol. 39, p. 471.
FETZER, M., AND J. FLORIAN 1930 Der Embryo ‘Fetzer’ mit beginniender Axialmesodernibildung und bereits augelegter Kloakenmembran. Ztsehr. f. mikr. anat. Forsch., Bd. 21, S. 351.
GREENHILL, J. P. 1927 A young human ovum in situ. Am. J. Anat., vol. 40, p. 315.
GROSSER, O. 1912 Development of the egg membranes and the placenta; menstruation. Keibel and Mall, Manual of Human Embryology, vol. 1, p. 91. J. B. Lippincott 00., Philadelphia and London.
1922 Zur Kenntnis der Trophoblastsehale bei jungen menschlichen Eiern. Ztschr. f. Anat. u. Entwicklngsgesch., Bd. 66, S. 179.
1924 Junge mensliehe Embryonen der dritten und vierten Woche. Ergebn. ll. Anat. u. Entwicklngsgesch., Bd. 25, S. 391.
HERTIG, A. T. 1935 Angiogenesis in the early human ehorion and in the primary placenta of the macaque monkey. Contrib. to Embryol., no. 146, vol. 25, p. 37.
KINDRED, J. E. 1933 A human embryo of the presomite period from the uterine tube. Am. J. Anat., vol. 53, p. 221.
LINZENMEIEK, G. 1914 Ein junges menschliches Ei in Situ. Arch. f. GrynZei.k., Bd. 102, S. 1.
MGLLENDORFF, W. VON 1921 Uber (lie jiingste bisher erkannto mensehliche Abortivei (Ei SCH). Ztschr. f. Anat. u. Entwieklngsgeseh, Ed. 62, S. 352. 9
1925 Das mensliehe Ei WO(lfring). Implantation. Verschlusz (ler Implantationsiiffnung und Keimesentwicklung beim Mensohen vor Bildung des Primitivstreifens. Ztschr. f. Anat. 11. Entwieklngsgesek, Bd. 76, S. 16.
STIEVE, H. 1926 Ein 131} Tage altes, in (ler Gebiirmutter erhaltenes und (lurch Eingriff gewonncnes menschliches Ei. Ztschr. f. mikr. anat. Forsch., Bd. 7, S. 295.
STREETER, G. L. 1920 A human embryo (Mateer) of the presomite period. Contrib. to Embryol., no. 43, vol. 9, p. 389.
1926 The ‘Miller’ ovum—the youngest human embryo thus far known. Contrib. to Embryol., no. 92, vol. 18, p. 31.
TEACHER, J. H. 1925 ()n the implantation of the human ovum and the early development of the trophoblast. Ztschr. f. Anat. u. Entwicklungsgeseh., Bd. 76, S. 360.
TENNANT, R., AND E. M. RAMSEY 1934. An early human embryo in situ. Surg., Gynec. and Obst., vol. 58, p. 968.
Plates
Plate 1

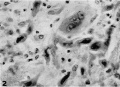
2 A portion of the maternal mucosa lying immediately outside the implantation cavity, showing invasion by deeply staining strands of syncytial trophoblast, the nuclei of which are mostly quite pycnotic. One large trophoblastie mass is shown which contains several typical nuclei. There are no indications of any histolytic activity on the part of this trophoblast. Note evidences of degeneration of connective tissue cells and polymorphonuelear infiltration. X 600.
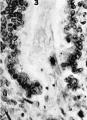
3 The lumen of a gland being invaded by a strand of syncytial trophoblast. The disorganization and piling up of the epithelial nuclei is quite apparent. X 600.
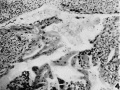
4 This shows the invation and partial lining of a maternal blood sinus in the marginal decidua by syncytial trophoblast. Decidua above, trophoblast and implantation cavity below. X 78.
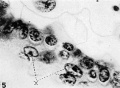
5 Cellular detail of the cytotrophoblast and the syncytial trophoblast (x) in the basal chorionic epithelium. Note the distinct cell membranes of the cytotrophoblast. Compare the nuclear appearances in the two layers. X 1340.
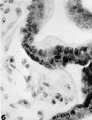
6 Junction of a villus with the basal chorion. In the covering of the villus many of the nuclei in the syncytial layer closely resemble those of the cytotrophoblast. The mesodermal core of the villus has been partially retracted. X 600.

1
2
3
4
5
6





Plate 2

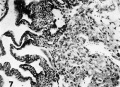
7 Cell columns continuing from the terminal ends of chorionic villi and uniting together peripherally. The mesodermal cores of the villi have been retracted in preparation. X 78.
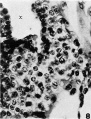
8 Tip of a, chorionic villus and the proximal end of a cell column in continuity with it. The cells of the cytotrophoblast clothing the villus are identical with those comprising this portion of the cell column. Villus core (X). X 600.
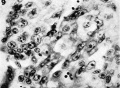
9 The character of the cells making up the column somewhat peripheral to region illustrated in figure 8. The cytological characteristics shown here are indicative of transition between cytotrophoblast and syncytium. The appearance of spaces due to the degeneration of ecrtain cells is clearly indicated. x 600.
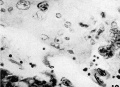
10 Syncytial trophoblast from the periphery of the implantation cavity. Note the great variation in the concentration of nuclei in the protoplasmic mass. X 600.
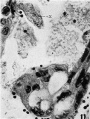
11 Syncytial trophoblastic masses projecting into the intervillous space. In the lower mass the nuclei are shrunken and pycnotic and large vacuoles are present. Projecting from the mass at the upper right is a non-nucleated. fragment (x) containing numerous deeply staining granules. Much of the debris in the intervillous space appears as disintegrating syncytial tisue. X 600.
7
8
9
10
11





Plate 3

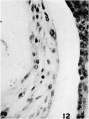
12 Mesoderm of the basal ehorion. Most of tl1e cells are fusiform with elongate nuclei containing scattered, finely divided chromatin. Occasional free spherical cells are to he found. Chorionic epithelium to the right with shrinkage space between it and the rnesoderm. X 600.
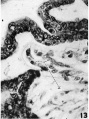
13 Linearly arranged vells (X) with large nuclei which form protoplasmic strands which might be interpreted as evidence of vasculogencsis. X 600.
14 Cellular detail from the core of a villus illustrating stages (1, 2, 3, 4) in the transformation of typical mesodermal cells into free spherical cells with basic rrytoplzxsm and very vesicular nuclei with prominent nucleoli characteristic of hemoblasts. X 1340.
15 An accumulation of free round cells at the tip of a villus suggestive of the appearance of blood islands. Near the lower border of the picture is a column of mesodermal cells suggestive of angiogenosis. X 600.
12
13
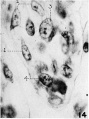
14
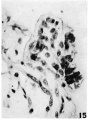
15




Cite this page: Hill, M.A. (2024, April 27) Embryology Paper - An early stage of human implantation. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_An_early_stage_of_human_implantation
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G