|
|
| (21 intermediate revisions by the same user not shown) |
| Line 1: |
Line 1: |
| {{Header}} | | {{Header}} |
| {{Ref-Conklin1905}} | | {{Ref-Conklin1905}} |
| | {{Conklin1905 TOC}} |
| | {| class="wikitable mw-collapsible mw-collapsed" |
| | ! Online Editor |
| | |- |
| | | [[file:Mark_Hill.jpg|90px|left]] Ascidians ([[Sea Squirt Development|sea squirts]]) are the evolutionary link between invertebrates and vertebrates. Conklin's 1905 detailed developmental cell lineage study of the ascidian embryo, that inspired similar studies in other species. This also set the stage for identifying embryonic patterning. |
| | <br> |
| | Edwin Grant Conklin (1863 – 1952) was an American biologist and zoologist at the Department of Biology at Princeton University. He was Chair of Biology for twenty-five years and studied the embryonic development of marine animals. |
| | <br> |
| | <br> |
| | |
| | See also - {{Ref-Conklin1905b}} |
| | <br> |
| | <br> |
| | |
| | [[Sea Squirt Development]] | [[Media:Conklin EG-The Organization and Cell-Lineage of the Ascidian Egg-1905.pdf|Book PDF]] |
| | |
| | [[Historic Embryology Papers]] |
| | |} |
| {{Historic Disclaimer}} | | {{Historic Disclaimer}} |
|
| |
| =The Organization and Cell-Lineage of the Ascidian Egg= | | =The Organization and Cell-Lineage of the Ascidian Egg= |
| | [[File:Edwin Conklin.jpg|thumb|alt=Edwin Conklin|Edwin Conklin (1863 – 1952)]] |
| | [[File:Conklin 1905 plate01.jpg|thumb|Living Egg of Oynthia partita - Maturation and Fertilization]] |
|
| |
|
| By Edwin G. Conklin. | | By Edwin G. Conklin. |
| | |
|
| |
|
| Professor of Zoology, University of Pennsylvania, | | Professor of Zoology, University of Pennsylvania, |
|
| |
|
| WITH PLATES I-XII.
| |
|
| |
|
| | With Plates I-XII. |
|
| |
|
| Philadelphia : 1905
| |
|
| |
|
| | Philadelphia - 1905 |
|
| |
|
| ==Contents== | | ==Contents== |
|
| |
|
| Introduction. | | [[#Introduction|Introduction]] |
| | | * A. Organization of the Egg |
| A. Organization of the Egg | | * B. Ascidian Embryology |
| | | * C. Methods and Material |
| B. Ascidian Embryology | |
| | |
| C. Methods and Material | |
| | |
| | |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 1|I. The Ovarian Egg]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 1|I. The Ovarian Egg]] |
| | | # Development of the Ovocyte |
| 1. Development of the Ovocyte
| | # Test Cells and Chorion . |
| | | # Structure of Fully Formed Ovocyte |
| 2. Test Cells and Chorion .
| |
| | |
| 3. Structure of Fullv Formed Ovocyte
| |
| | |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 2|II. Maturation and Fertilization]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 2|II. Maturation and Fertilization]] |
| | * A. Maturation |
| | # Disappearance of Nuclear Membrane |
| | # Chromosomes |
| | # Nucleolus |
| | # Spindle Formation |
| | # Movements of Spindle and Nuclear Plasm ; Formation of |
|
| |
|
| A. Maturation
| | * B. Fertilization |
| | | # Entrance of Spermatozoon |
| 1. Disappearance of Nuclear Membrane
| | # Movements of Ooplasm |
| | | ## Localization of Yellow Protoplasm |
| 2. Chromosomes
| | ## Localization of Clear Protoplasm and Yolk |
| | | # Development of Sperm Nucleus and Aster |
| 3. Nucleolus
| | # Path of Spermatozoon within the Egg . |
| | | # The Egg Nucleus and its Movements |
| 4. Spindle Formation
| | # Sperm Amphiaster and First Cleavage Spindle |
| | | # Dispermy |
| 5. Movements of Spindle and Nuclear Plasm ; Formation of
| | * Polar Bodies |
| | |
| B. Fertilization | |
| | |
| 1. Entrance of Spermatozoon
| |
| | |
| 2. Movements of Ooplasm
| |
| a. Localization of Yellow Protoplasm
| |
| b. Localization of Clear Protoplasm and Yolk
| |
| | |
| 3. Development of Sperm Nucleus and Aster
| |
| | |
| 4. Path of Spermatozoon within the Egg .
| |
| 5. The Egg Nucleus and its Movements
| |
| | |
| 6. Sperm Amphiaster and First Cleavage Spindle
| |
| | |
| 7. Dispermy
| |
| | |
| Polar Bodies | |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 3|III. Orientation of Egg and Embryo]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 3|III. Orientation of Egg and Embryo]] |
| | | # Van Beneden and Julin's System of Orientation |
| 1. Van Beneden and Julin's System of Orientation
| | # Seeliger's System |
| | | # Samassa's System |
| 2. Seeliger's System
| | # Castle's System |
| | | # Evidences in Favor of Van Beneden and Julin's System |
| 3. Samassa's System
| |
| | |
| 4. Castle's System
| |
| | |
| 5. Evidences in Favor of Van Beneden and Julin's System
| |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 4|IV. Cell-Lineage]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 4|IV. Cell-Lineage]] |
| Line 86: |
Line 79: |
|
| |
|
| B. Cleavage of the Egg ; First to Seventh Generation of Cells | | B. Cleavage of the Egg ; First to Seventh Generation of Cells |
| | | # First Cleavage ; 1-2 Cells |
| 1. First Cleavage ; 1-2 Cells
| | # Second Cleavage ; 2-4 Cells . |
| | | # Third Cleavage; 4-8 Cells . |
| 2. Second Cleavage ; 2-4 Cells .
| | # Fourth Cleavage; 8-16 Cells |
| | | # Fifth Cleavage ; 16-32 Cells |
| 3. Third Cleavage; 4-8 Cells .
| | # Sixth Cleavage ; 32-64 Cells |
| | |
| 4. Fourth Cleavage; 8-16 Cells
| |
| | |
| 5. Fifth Cleavage ; 16-32 Cells
| |
| | |
| 6. Sixth Cleavage ; 32-64 Cells
| |
|
| |
|
| C. Gastrulation ; Seventh to Ninth Generation of Cells | | C. Gastrulation ; Seventh to Ninth Generation of Cells |
| Line 104: |
Line 91: |
|
| |
|
| 8. Eighth Cleavage ; 112-132 Cells, 132-218 Cells | | 8. Eighth Cleavage ; 112-132 Cells, 132-218 Cells |
|
| |
|
| |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 5|V. Later Development]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 5|V. Later Development]] |
| | | # Closure of Blastopore |
| 1. Closure of Blastopore
| | # Development of Larva |
| | |
| 2. Development of Larva
| |
| | |
| | |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 6|VI. Comparisons with A.mphioxus and Amphibia]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 6|VI. Comparisons with A.mphioxus and Amphibia]] |
| | | # Axial Relations of Egg and Embryo |
| 1. Axial Relations of Egg and Embryo
| | # Entrance of Spermatozoon |
| | | # Cleavage |
| 2. Entrance of Spermatozoon
| | # Blastula and Gastrula |
| | | # Closure of Blastopore |
| 3. Cleavage
| | # Neural Plate |
| | | # Chorda |
| 4. Blastula and Gastrula
| | # Origin of Mesoderm |
| | |
| 5. Closure of Blastopore
| |
| | |
| 6. Neural Plate
| |
| | |
| 7. Chorda
| |
| | |
| 8. Origin of Mesoderm
| |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 7|VII. The Organization of the Egg]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 7|VII. The Organization of the Egg]] |
| | | * A. Polarity |
| A. Polarity | | * B. Symmetry |
| | | * C. Cytoplasmic Localization |
| B. Symmetry | | # Localization in the Cleavage Stages Cytoplasmic Organization and the Nuclear |
| | | # Localization before Cleavage |
| C. Cytoplasmic Localization | | * D. Genesis of the Organization of the Egg |
| | | # Role of the Nucleus in Differentiation - Inheritance Theory |
| 1. Localization in the Cleavage Stages
| | # Factors of Localization |
| | | ## Cytoplasmic Movements |
| | | ## Cell Division as a Factor of Localization |
| Cytoplasmic Organization and the Nuclear | | * Types of Germinal Localization ; Evolution of Types |
| | | # Annelid-Mollusk Type |
| 2. Localization before Cleavage
| | # Ctenophore Type |
| | | # Echinoderm Type |
| D. Genesis of the Organization of the Egg | | # Ascidian Type |
| | |
| 1. Role of the Nucleus in Differentiation - Inheritance Theory
| |
| | |
| 2. Factors of Localization
| |
| | |
| a. Cytoplasmic Movements
| |
| | |
| b. Cell Division as a Factor of Localization
| |
| | |
| E. Types of Germinal Localization ; Evolution of Types
| |
| | |
| 1. Annelid-Mollusk Type
| |
| | |
| 2. Ctenophore Type
| |
| | |
| 3. Echinoderm Type
| |
| | |
| 4. Ascidian Type
| |
|
| |
|
| [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 8|Summary]] | | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 8|Summary]] |
| | # Ascidian Embryology |
| | # Cytological Results |
| | # Organization of the Egg |
|
| |
|
| I. Ascidian Embryology
| | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 9|Literature Cited]] |
|
| |
|
| II. Cytological Results
| | [[Paper - The Organization and Cell-Lineage of the Ascidian Egg 10|Explanation of Figures]] |
|
| |
|
| III. Organization of the Egg | | <gallery> |
| | File:Conklin 1905 plate01.jpg|Plate I |
| | File:Conklin 1905 plate02.jpg|Plate II |
| | File:Conklin 1905 plate03.jpg|Plate III |
| | File:Conklin 1905 plate04.jpg|Plate IV |
| | File:Conklin 1905 plate05.jpg|Plate V |
| | File:Conklin 1905 plate06.jpg|Plate VI |
| | File:Conklin 1905 plate07.jpg|Plate VII |
| | File:Conklin 1905 plate08.jpg|Plate VIII |
| | File:Conklin 1905 plate09.jpg|Plate IX |
| | File:Conklin 1905 plate10.jpg|Plate X |
| | File:Conklin 1905 plate11.jpg|Plate XI |
| | File:Conklin 1905 plate12.jpg|Plate XII |
| | </gallery> |
|
| |
|
| Literature Cited
| | <gallery> |
| | | File:Conklin 1905 fig01-02.jpg|Fig 1-2 |
| Explanation of Figures
| | File:Conklin 1905 fig03-06.jpg|Fig 3-6 |
| | File:Conklin 1905 fig07-08.jpg|Fig 7-8 |
| | File:Conklin 1905 fig09-12.jpg|Fig 9-12 |
| | File:Conklin 1905 fig13-16.jpg|Fig 13-16 |
| | File:Conklin 1905 fig17-20.jpg|Fig 17-20 |
| | File:Conklin 1905 fig21-24.jpg|Fig 21-24 |
| | File:Conklin 1905 fig25-26.jpg|Fig 25-26 |
| | File:Conklin 1905 fig27-33.jpg|Fig 27-33 |
| | File:Conklin 1905 fig34-35.jpg|Fig 34-35 |
| | </gallery> |
| | ==Introduction== |
|
| |
|
| | ===A. Organization of the Egg=== |
| | Recent years have witnessed a revival of the ancient controversy as to the nature and contents of the germ cells. On the one hand are those who with Weismann maintain that the egg must contain the elements or determinants of very many structures which will appear in the course of development; on the other hand are ranged the modern epigenesists who find in the egg cell only complex chemical substances which have the capacity under certain outer conditions of undergoing regular transformations into other substances which incidentally have peculiar forms, just as crystals have. |
|
| |
|
|
| |
| ==Introduction==
| |
|
| |
| A. Organization of the Egg. Recent years have witnessed a revival of
| |
| the ancient controversy as to the nature and contents of the germ cells. On
| |
| the one hand are those who with Weismann maintain that the egg must contain
| |
| the elements or determinants of very many structures which will appear in the
| |
| course of development; on the other hand are ranged the modern epigenesists
| |
| who find in the egg cell only complex chemical substances which have the capacity
| |
| under certain outer conditions of undergoing regular transformations into other
| |
| substances which incidentally have peculiar forms, just as crystals have.
| |
|
| |
|
| But while this modern controversy recalls the ancient one between the | | But while this modern controversy recalls the ancient one between the |
| Line 202: |
Line 174: |
| somewhere between these two extremes ; the real problem is how much or how | | somewhere between these two extremes ; the real problem is how much or how |
| little of organization is present, and not whether the germ is organized at all. | | little of organization is present, and not whether the germ is organized at all. |
| | |
|
| |
|
| Though the controversy as to evolution and epigenesis has thus been narrowed within relatively small limits, and has therein- lost much of its startling and | | Though the controversy as to evolution and epigenesis has thus been narrowed within relatively small limits, and has therein- lost much of its startling and |
| Line 211: |
Line 184: |
| structures in earlier and earlier stages of development and so finally in the | | structures in earlier and earlier stages of development and so finally in the |
| unsegmented egg itself. | | unsegmented egg itself. |
| | |
|
| |
|
| It is not many years since all embryological studies were dominated by the | | It is not many years since all embryological studies were dominated by the |
| Line 224: |
Line 198: |
| the basis of any thorough study of development, inheritance and evolution. | | the basis of any thorough study of development, inheritance and evolution. |
|
| |
|
| B. Ascidian Embryology. Anyone who has observed the ascidian egg will | | ===B. Ascidian Embryology=== |
| understand why it has been such a favorite object of study. The cleavage of the | | Anyone who has observed the ascidian egg will understand why it has been such a favorite object of study. The cleavage of the |
| egg is so beautifully regular and can be observed so readily in life that it is not | | egg is so beautifully regular and can be observed so readily in life that it is not |
| surprising that ascidians were among the first animals to which the "cell-lineage" | | surprising that ascidians were among the first animals to which the "cell-lineage" |
| Line 263: |
Line 237: |
| me to say that I have spared no pains or labor to make them accurate. | | me to say that I have spared no pains or labor to make them accurate. |
|
| |
|
| ('. Material and Methods. Early in July. 1903, while working at the Marine
| | ===C. Material and Methods=== |
| Biological Laboratory, Woods Holl, Mass.. I began the study of the maturation | | |
| | Early in July. 1903, while working at the Marine Biological Laboratory, Woods Holl, Mass.. I began the study of the maturation |
| and fertilization of the egg of Ciona intestinalis (L.) Flemming, with the aim mentioned in the preceding paragraph. Only a small number of these animals was to | | and fertilization of the egg of Ciona intestinalis (L.) Flemming, with the aim mentioned in the preceding paragraph. Only a small number of these animals was to |
| be found at that time at Woods Holl. though they occurred more abundantly later | | be found at that time at Woods Holl. though they occurred more abundantly later |
| Line 281: |
Line 256: |
| I took up also the living eggs of Ciona and Molgula, and finally I fixed and prepared for microscopical examination, both as whole objects and as serial sections, | | I took up also the living eggs of Ciona and Molgula, and finally I fixed and prepared for microscopical examination, both as whole objects and as serial sections, |
| the eggs and embryos of all three of these genera. | | the eggs and embryos of all three of these genera. |
| | |
|
| |
|
| Castle (1896) has described in considerable detail the time and manner of egg | | Castle (1896) has described in considerable detail the time and manner of egg |
| Line 298: |
Line 274: |
| another individual, hut if fertilized with sperm from the same animal the eggs | | another individual, hut if fertilized with sperm from the same animal the eggs |
| rarely if ever develop, as Castle has shown. This is due to the fact that such | | rarely if ever develop, as Castle has shown. This is due to the fact that such |
| spermatozoa never enter the egg, though they may be quite active. Morgan (1904) | | spermatozoa never enter the egg, though they may be quite active. Morgan (1904) has recently discussed this interesting fact in a suggestive manner. |
| lias recently discussed this interesting fact in a suggestive manner.
| | |
|
| |
|
| The method which I employed in studying the living eggs of these ascidians | | The method which I employed in studying the living eggs of these ascidians |
| Line 315: |
Line 291: |
| of several eggs through consecutive portions of it, I chose the latter and easier | | of several eggs through consecutive portions of it, I chose the latter and easier |
| method. | | method. |
| | |
|
| |
|
| All my studies of the living eggs of these ascidians were made with a dry lens, | | All my studies of the living eggs of these ascidians were made with a dry lens, |
| Line 325: |
Line 302: |
| difficult" to explain. I can only account for it by supposing that he obtained the | | difficult" to explain. I can only account for it by supposing that he obtained the |
| eggs in the evening and studied them by yellow artificial light. | | eggs in the evening and studied them by yellow artificial light. |
| | |
|
| |
|
| Preserved material was fixed in various fluids, Perenyi's,. Kleinenberg's, PicroAcetic, Sublimate and Sublimate-Acetic. For the study of entire eggs and embryos | | Preserved material was fixed in various fluids, Perenyi's,. Kleinenberg's, PicroAcetic, Sublimate and Sublimate-Acetic. For the study of entire eggs and embryos |
| Line 336: |
Line 314: |
| satisfactory results. Such material was stained on the slide in Delafield's Hematoxylin and Eosin or in Iron Hematoxylin and Bordeau red. | | satisfactory results. Such material was stained on the slide in Delafield's Hematoxylin and Eosin or in Iron Hematoxylin and Bordeau red. |
|
| |
|
| Castle states that he found it necessary to remove the egg envelopes by | | |
| drawing the egg into a pipette through an opening so small that the egg alone could puss in, after the manner recommended l>\ Chabry. In most cases I have | | Castle states that he found it necessary to remove the egg envelopes by drawing the egg into a pipette through an opening so small that the egg alone could puss in, after the manner recommended by Chabry. In most cases I have |
| found that the presence of the egg envelope's (iocs not seriously interfere with clear | | found that the presence of the egg envelope's (iocs not seriously interfere with clear |
| seeing, possibly owing to the fact that in the study of preparations 1 have used an | | seeing, possibly owing to the fact that in the study of preparations I have used an immersion lens in which the depth of locus is relatively slight. In late stages, |
| | | however, the test cells are sometimes confusing, and in the case of Ciona I found |
| immersion lens in which the depth of locus is relatively slight. In late stages, | | that these, together with the other envelopes, could he removed by simply rolling the eggs under the cover glass. In Cynthia the envelopes may sometimes he removed in the same way, though not so easily as in Ciona. |
| however, the test cells are sometimes confusing, and in the case of Ciona 1 found | |
| that these, together with the other envelopes, could he removed by simply rolling | |
| the eggs under the cover glass. In Cynthia the envelopes may sometimes he | |
| removed in the same way, though not so easily as in Ciona. | |
| | |
| | |
| SUMMARY.
| |
| | |
| | |
| I. Ascidian Embryology.
| |
| | |
| 1. The orientation of the ascidian egg and embryo adopted by Van Beneden
| |
| and Julin is correct, that of Seeliger, Samassa and Castle is wrong (pp. 20-37).
| |
| | |
| 2. The cell-lineage given by Castle is correct for the early stages; from the
| |
| 48-cell stage on it is wrong (pp. 5 65 U ) .
| |
| | |
| 3. The egg axis corresponds very nearly with the gastrular axis; during the
| |
| closure of the blastopore this axis is shifted so that it is no longer dorso-ventral as
| |
| in the early stages, but is antero-ventral and postero-dorsal in direction in the larva
| |
| (pp. 73, 75-77).
| |
| | |
| 4. The relation of the germinal layers to the cleavage planes is very different
| |
| from the account given by Van Beneden and Julin and by Castle, and is more
| |
| nearly in accord with that of Seeliger, Davidoff and Samassa. All cells above the
| |
| equator (3d cleavage plane) are ectodermal and neural plate cells; all below are
| |
| endodermal, mesodermal and neural plate (pp. 47-48).
| |
| | |
| ''5. The factors of gastrulation are (a) change of shape of the cells of the
| |
| animal and vegetal hemispheres, (b) overgrowth of the marginal cells (pp. 64-65).
| |
| Peculiarities of the gastrula are foreshadowed in the egg at a very early stage
| |
| (pp. 45. 50. 59).
| |
| | |
| / 6. The muscle and mesenchyme cells arise from a common base, the mesodermal crescent, which surrounds the posterior side of the egg just below the
| |
| equator (pp. 19-21); ultimately these cells surround the posterior margin of the blastopore (pp. 51-55); the most laterally situated of these cells become the large
| |
| muscle cells of the tail of the tadpole, the portion of the crescent lying nearest the
| |
| dorsal mid-line becomes mesenchyme (pp. 61-07). In the overgrowth of the dorsal
| |
| lip of the blastopore the muscle cells become separated from the mesenchyme (pp.
| |
| 69, 84-87).
| |
| | |
| 7. The chorda arises at the anterior border of the blastopore from yolk-laden
| |
| cells which resemble endoderm (pp. 53, 61, 62, 70-72, 83, 84).
| |
| | |
| 8. The neural plate arises on the anterior side of the egg from cells of both
| |
| hemispheres ; it extends from the margin of the anterior lip of the blastopore to a
| |
| point about one-third of the way from the equator to the animal pole (pp. 52-54,
| |
| 61-63, 66-68, 70, 72, 73. 82, 83).
| |
| | |
| 9. The nervous and muscular systems do not arise from a common base as
| |
| claimed by Castle; there is no nerve ring around the blastopore (pp. 61, 72, 73).
| |
| | |
| ' 10. A comparison of the early development of ascidians with that of Amphioxus and amphibians shows that there is fundamental agreement among them in
| |
| axial relations of egg and larva, in bilaterality of cleavage, in the method of closure
| |
| of the blastopore and probably in the origin and position in the embryo of the
| |
| neural plate, the chorda and the mesoderm (pp. 73-87).
| |
| | |
| II. Cytologic al Results.
| |
| | |
| 11. The maturation spindles are peculiar; they have no centrosomes nor
| |
| asters ; they are formed entirely within the nuclear area from nuclear linin and
| |
| chromosomes ; their fibres at first radiate in all directions, and finally they form a
| |
| barrel-shaped spindle. The chromosomes separate without any possible influence
| |
| from centrosomes or traction on the part of spindle fibres (pp. 15, 16).
| |
| | |
| 12. In the first and second cleavages a small nuclear spindle like those present during maturation, lies between the two large asters, and in Ciojia it is quite
| |
| distinct, from them. The separation of the daughter chromosomes takes place here
| |
| as in the maturation divisions (pp. 40, 41).
| |
| | |
| 13. The spermatozoon enters near the lower pole and rotates after entering
| |
| the egg so that its centrosome is directed forward ; the centrosome is derived from
| |
| the middle piece of the spermatozoon and can be followed without interruption until
| |
| it divides, at right angles to the copulation path, and gives rise to the sperm amphiaster and finally to the cleavage centrosomes (pp. 22-24). A netrum is formed in
| |
| the division of all centrosomes (p. 40).
| |
| | |
| III. Organization of the Egg.
| |
| | |
| 14. In the ovocyte of Cynthia parti/a there is a peripheral layer of yellow
| |
| protoplasm, a central mass of gray yolk, and a large clear germinal vesicle, which is
| |
| eccentric toward the animal pole. These same parts are jiresent in the eggs of
| |
| other ascidians, but are differently colored (pp. 11, 12).
| |
| | |
| 15. When the wall of the germinal vesicle dissolves at the beginning of the
| |
| maturation divisions a large amount of clear protoplasm, containing dissolved oxyebromatin, is liberated into the cell body. This clear protoplasm is eccentric
| |
| toward the animal pole and is distinct from the yolk and peripheral layer (pp.
| |
| 13, 17).
| |
| | |
| lii. Immediately after the entrance of the spermatozoon the yellow and clear
| |
| protoplasm How rapidly to the lower pole, where the yellow protoplasm collects
| |
| around the point of entrance; the clear protoplasm lies at a deeper level. The
| |
| yellow protoplasm then spreads out until it covers the surface of the lower hemisphere. This flowing of protoplasm to the point of entrance of the sperm is comparable with what takes place in many animals, though here much more extensive
| |
| and rapid than elsewhere (pp. 1921, 77).
| |
| | |
| 17. The withdrawal of protoplasm from the upper pole leaves the maturation
| |
| spindles closely surrounded by yolk. The polar bodies are thus formed at the
| |
| middle of a yolk-rich hemisphere, which is, however, the animal pole and not
| |
| the vegetal pole as was claimed by Castle (pp. 19-21, 29, 30, 36, 37, 87-90).
| |
| | |
| 18. The sperm nucleus moves from the point of entrance toward the equator
| |
| in a path which is apparently predetermined. Tins path lies in the plane of the
| |
| first cleavage and the point, just below the equator, at which the sperm nucleus
| |
| stops in its upward movement, becomes the posterior pole of the embryo. The
| |
| median plane and the posterior pole are probably not determined by the path of the
| |
| spermatozoon, but by the structure of the egg. All the axes of the future animal
| |
| are now clearly established, antero-posterior, right-left, dorso-ventral (pp. 22, 26,
| |
| 90-93).
| |
| | |
| 19. As the sperm nucleus moves to the posterior pole the clear and the yellow
| |
| protoplasm move with it ; the latter collects into a yellow crescent with its middle
| |
| at the posterior pole and its horns extending about halfway around the egg just
| |
| below the equator. This position it retains throughout the whole development,
| |
| giving rise to the muscle and mesenchyme cells mentioned in 6 (pp. 19-21, 97, 98).
| |
| | |
| 20. After the sperm and egg nuclei have met at the posterior pole they move
| |
| in toward the center of the egg and the clear protoplasm goes with them ; the outplace where the latter remains in contact with the surface is along the upper border
| |
| of the crescent. At the close of the first cleavage the nuclei and clear protoplasm
| |
| move into the upper hemisphere, and thereafter, throughout development, this
| |
| hemisphere contains most of the clear protoplasm and gives rise to the ectoderm
| |
| (pp. 20, 21, 42, 102).
| |
| | |
| 21. The yolk which before maturation was central in position is shifted
| |
| toward the animal pole when the protoplasm flows down to meet the spermatozoon ;
| |
| when the sperm nucleus and surrounding protoplasm move to the posterior pole
| |
| the yolk is moved down around the anterior side of the egg to the lower pole, and
| |
| when the clear protoplasm moves into the upper hemisphere the yolk is largely
| |
| collected in the lower hemisphere. This yolk rich area gives rise to the endoderm
| |
| (pp. 20', 33-35, 12, 102).
| |
| | |
| 22. At the close of the first cleavage the principal germ regions of the embryo
| |
| are visible in their definite positions and proportions, viz. : the muscle-mesenchyme crescent and the ectodermal and endodermal areas. The chorda and neural plate
| |
| areas are also visibly different from surrounding areas at this stage (pp. 42, 50, 95,
| |
| 97, 98, 108).
| |
| | |
| 23. In many eases the cleavage planes do not follow the lines of differentiation but cut across them. Although cleavage is, under normal conditions, constant
| |
| in form, it is less constant and fundamental than the type of localization, and the
| |
| two are relatively independent (pp. 103, 104).
| |
| | |
| 24. The chief factor of localization is protoplasmic flowing; cell division is a
| |
| factor of subordinate value (pp. 102-104).
| |
| | |
| 25. Experiments which demonstrate the totipotence of blastomeres or regions
| |
| of the egg prove nothing with regard to the presence or absence of differentiation
| |
| in those parts. Some eggs with a high degreee of differentiation have at the same
| |
| time great capacity for regulation, e.g., those of ascidians ; 1 others with no greater
| |
| differentiation have little regulative capacity, e. g., ctenophores and mollusks.
| |
| Therefore the potency of any part of an egg or embryo is no satisfactory measure of
| |
| the degree of its differentiation (pp. 93-95).
| |
| | |
| 26. The organization of the ovocyte is not the initial organization. The yellow protoplasm (mesoplasm) of the Cynthia egg is probably derived, at least in part,
| |
| from sphere material (archoplasm) which arose from the nucleus at the last ovogonic division. / The yolk (endoplasm) is formed by the activity of the "yolk
| |
| matrix" (Crampton) which also is probably sphere material. The clear protoplasm
| |
| (ectoplasm) is derived from the germinal vesicle at the first maturation division. Thus
| |
| many important regions of the egg come, at least in part, from the nucleus, and a
| |
| method is therein suggested .of harmonizing the facts of cytoplasmic localization
| |
| with the nuclear inheritance theory (pp. 99-101)./
| |
| | |
| 27. There are several distinct types of germinal localization. /The annelidmollusk type does not approach that of chordates or echinoderms in the earliest
| |
| stages of localization more closely than in the cleavage or gastrular stages. There is
| |
| no convergence toward a common type in the earliest stages (p. 104-109).
| |
| | |
| 28. Embryonic repetitions (recapitulations), as well as many other homologies,
| |
| probably result from similarities of egg organization common to each type (p. 109).
| |
| | |
| 29. "Precocious segregation" is not a satisfactory explanation of the origin
| |
| of germinal organization (pp. 109, 110).
| |
| | |
| / 30. The evolution of animals must be accompanied by an evolution of the
| |
| type of germinal organization ; modifications of this organization are probably the
| |
| immediate causes of evolution. Transformations which would be impossible in
| |
| adults are readily brought about by modifications in the organization of the egg
| |
| {e.g., inverse symmetry). Perhaps profound mutations or even the origin of distinct types may be so explained (pp. 110, 111).
| |
| 1 See foot-note p. 95.
| |
| | |
| | |
| | |
| ==Literature Cited==
| |
| | |
| | |
| BANCROFT, F. \V. (1899).
| |
| | |
| Ovogenesis in Distaplia occidentalis, Ritter. Bull. Mus. Camp. Zoo/, xxxv.
| |
| BENEDEX, E. van kt CH. JULIN, (1884).
| |
| | |
| La segmentation chez les ascidens dans ses rapportes avec l'organization de la larve. Arch, de Biol., v.
| |
| BENEDEN, E. van et CH. JULIN, (1886).
| |
| | |
| Recherches snr la Morphologie des Tuniciers. Arch, de Biol., vi.
| |
| BENEDEN, E. van et A. NEYT, ( L887).
| |
| | |
| Nouvelles recherches sur la fecondation et la division mitosique chez 1'Ascaride Megalocephale. Bull.
| |
| /'Acad. Row Belgique, 3eser., XI V.
| |
| BIGELOVV, M. A (1902 .
| |
| | |
| The Early Development of Lepas. A study of Cell-Lineage and Germ Layers. Bull. Mus. Comp. Zoo/., xl.
| |
| BLOCHMANN, FR. (1882). ,
| |
| | |
| Beitriige zur Kenntniss der Gasteropoden. Zeit. wiss. Zoo/., xxxviii.
| |
| BOYERI, TH. (18S7, 1SSS, 1890).
| |
| | |
| Zellen Studien, I, II, III. Jena.
| |
| BOVERI, TH. (1901).
| |
| | |
| Ueber die Polaritat des Seeigeleies. Verh. Phys. Med. Ges. Wurzburg, xxxiv.
| |
| BOVERI. TH. (1901).
| |
| | |
| Die Polaritat von Ovocyte, Ei und Larve des Strongylocentrotus lividus. Zoo/. Jahrb., .xiv.
| |
| BROOKS, W. K. (1S93).
| |
| | |
| The Genus Salpa. Baltimore.
| |
| CARAZZI, D. (1904).
| |
| | |
| Ricerche embriologiche e citologiche sull 'uovo di Myzostoma glabrum Leuckart. Monitoro. Zool.
| |
| Italiano, xv.
| |
| CASTLE, W. E. (1894).
| |
| | |
| On the Cell-Lineage of the Ascidian Egg. A preliminary notice. Proc. Amer. Acad. Arts and Sci., xxx.
| |
| CASTLE, W. E. (1896).
| |
| | |
| The Early Embryology of Ciona intestinalis, Flemming (L. ). Bull. Mus. Comp. Zoo/., xxvii.
| |
| CHABRY, L, (1887
| |
| | |
| Contribution a l'embryologie normal et teratologique des Ascidies simples. Jour. Ana/, et Physiol, xxiii.
| |
| CHILD, C. M. (1900).
| |
| | |
| The Early Development of Arenicola and Sternaspis. Arch. Entw. Meek., ix.
| |
| C( INKLIN, E. G. (1897)
| |
| | |
| The Embryology of Crepidula. Jour. Morph., xiii.
| |
| CONKLIN, E. G. (1899).
| |
| | |
| Protoplasmic Movement as a Factor of Differentiation. Woods Ho// Bio/. Lec/ures, 1S98.
| |
| CONKLIN, E. G. (1902).
| |
| | |
| Karyokinesis and Cytokinesis in the Maturation, Fertilization and Cleavage of Crepidula and other
| |
| Gasteropoda. Jour. Acad. Nat. Sci. Philadelphia, xii.
| |
| CONKLIN, E. G. (1903).
| |
| | |
| The Cause of Inverse Symmetry. Anal. Anz., xxiii.
| |
| CONKLIN, E. G. (1903).
| |
| | |
| The Earliest Differentiations of the Egg. Abstract of paper before Amer. Philos. Soc, Science, xvii.
| |
| CRAMPTON, H. E. (1894).
| |
| | |
| Reversal of Cleavage in a Sinistral Gasteropod. Ann. New York Acad. Sci., viii.
| |
| CRAMPTON, H. E. (1896).
| |
| | |
| Experimental Studies on Gasteropod Development. Arch. Entiv. Mech., iii.
| |
| CRAMPTON, H. E. (1S97).
| |
| | |
| The Ascidian Half- Embryo. Ann. Nezv York Acad. Sci., x.
| |
| CRAMPTON, H. E. (1899).
| |
| | |
| Studies upon the Early History of the Ascidian Egg. Jour. Morph., xv. Supplement.
| |
| DAYIDOFF, M.v. (1889-1891).
| |
| | |
| Untersuchungen zur Entwicklungsgeschichte der Distaplia magnalarva, Delia Yalle. Mitth. Zool.
| |
| Stat. Neapel, ix.
| |
| | |
| DRIESCH, H. (1895).
| |
| | |
| Von der Entwicklung einzelner Ascidienblastomeren. Arch. Entw. Mech., i.
| |
| DRIESCH, H. (1896).
| |
| | |
| Betrachtungen iiber die Organization des Eies und ihre Genese. Arch. Eniw. Mech., iv.
| |
| DRIESCH, H. (1903).
| |
| | |
| Ueber Aenderung der Regulationsfahigkeiten im Verlauf der Entwicklung bei Ascidien. Arch. Entw.
| |
| Mech., xvii.
| |
| FISCHEL, A. (1897, 1898).
| |
| | |
| Experimentelle Untersuchungen am Ctenophorenei. Arch. Entw. Mech., vi, vii.
| |
| FISCHEL, A. (1903).
| |
| | |
| Entwicklung und Organ-Differenzirung. Arch. Entw. Mech., xv.
| |
| FLODERUS, M. (1896).
| |
| | |
| Ueber die Bildung der Follikelhiillen bei den Ascidien. Zeit. wiss. Zool., lxi.
| |
| GARBOWSKI, T. (1S98).
| |
| | |
| Amphioxus als Grundlage der Mesodermtheorie. Anat. Anz., xiv.
| |
| GOLSKI, St. (1899)'.
| |
| | |
| La Maturation et la fecondation de 1'oeuf de Ciona intestinalis F. Bull. Internat. Acad. Sci. Cracovie.
| |
| HACKER, V. (1899).
| |
| | |
| Praxis und Theorie der Zellen und Befruchtungslehre. Jena.
| |
| HATSCHEK, B. (1881).
| |
| | |
| Studien iiber die Entwicklung der Amphioxus. Arb. Zool. /fist, ll'ien, iv.
| |
| HERTWIG, O. (1892).
| |
| | |
| Urmund und Spina Bifida. Arch. mik. Anat., xxxix.
| |
| HERTWIG, R. (1904).
| |
| | |
| Ueber physiologiscbe Degeneration bei Actinosphaerium Eichorni. Fcstschr. zum siebzigsten Geburtstage v. E. Haeckel.
| |
| HILL, M. D. (1895).
| |
| | |
| Notes on the Fecundation of the Egg of Sphaerechinus granulans and on the Maturation and Fertilization of the Egg of Phallusia mammalata. Quart. Journ. Mic. Sci., xxxviii.
| |
| JORDAN, E. O. (1893).
| |
| | |
| The Habits and Development of the Newt. Jour. Morph., viii.
| |
| JULIN, CH. (1893).
| |
| | |
| Ovog^nfese, spermatogn<;se et fecondation chez Styelopsis. Bull. Sci. France et Belg., xxv.
| |
| KLAATSCH, H. (1896).
| |
| | |
| Bemerkungen iiber die Gastrula des Amphioxus. Morph. Ja/ub., xxv.
| |
| KOFOID, C. A. (1S94).
| |
| | |
| On Some Laws of Cleavage in Limax. Proc. Amer. Acad. Arts and Sci., xxix.
| |
| KOPSCH, Fr. (1900).
| |
| | |
| Ueber das Verhaltniss der embryonalen Achsen zu den drei ersten Furchungsebenen beim Frosch.
| |
| Internat. Mona/schr. f. Anat. u. Physiol, xvii.
| |
| KORSCHELT, u. HEIDER (1903).
| |
| | |
| Lehrbuch der vergleich. Entwicklungsgeschichte der wirbellosen Thiere. Allgemeiner Theil, 2te Lieferung. Jena.
| |
| KOVVALEVSKY, A. (1866)
| |
| Entwicklungsgeschichte der einfachen Ascidien. Mem. V Acad. St. Petersbourg, Ser. 7, x.
| |
| KOVVALEVSKY^ A. (1867).
| |
| | |
| Entwicklungsgeschichte der Amphioxus lanceolatus. Idem, xi.
| |
| KOWALEVSKY, A. (1871).
| |
| | |
| Weitere Studien iiber die Entwicklung der einfachen Ascidien. Arch. mik. Anat, vii.
| |
| KUPFFER, C. v. (1870).
| |
| | |
| Die Stammverwandschaft zwischen Ascidien und VYirbelthieren. Arch. mik. Aval., vi.
| |
| LILLIE, F. R. (1895).
| |
| | |
| The Embryology of the Unionidte. Jour. Morph., x.
| |
| LILLIE, F. R. (1899).
| |
| | |
| Adaptations in Cleavage. Woods Holt Biol. Led., 1898.
| |
| LILLIE, F. R. (1901).
| |
| | |
| The Organization of the Egg of Unio, etc. Jour. Morph., xvii.
| |
| LILLIE, F. R. (1902).
| |
| | |
| Differentiation without Cleavage in the Egg of the Annelid Chaetopterus pergamentaceus. Arch.
| |
| Entw. Mech., xiv.
| |
| | |
| | |
| LWOFF, B. (1894).
| |
| | |
| Die Bildung der primaren Keimblatter und die Entstehung des Chorda und des Mesoderms bei den
| |
| W'irbelthieren. Bull. Soc. Imp. Nat. d. Moscou, N. S., viii.
| |
| LYON, E. P. (1903).
| |
| | |
| Experiments on Artificial Parthenogenesis. Amer. Jour. Physiol., ix.
| |
| MAC BRIDE, E. \V. (1898).
| |
| | |
| The Early Development of Amphioxus. Quart. Jour. Mic. Sei., xl.
| |
| MORGAN, T. H. (1S90).
| |
| | |
| The Origin of the Test Cells of Ascidians. Journ. Morph., iv.
| |
| MORGAN, T. H. (1901).
| |
| | |
| Regeneration. New York.
| |
| MORGAN, T. H. (1903).
| |
| | |
| Evolution and Adaptation. New York.
| |
| MORGAN, T. H. (1904).
| |
| | |
| Self Fertilization Induced by Artificial Means. Jour. Exp. Zool., i.
| |
| MORGAN and HAZEN (1900).
| |
| | |
| The Gastrulation of Amphioxus. Jour. Morph., xvi.
| |
| MOSKOWSKI, M. (1902).
| |
| | |
| Ueber den Einfluss der Schwerkraft auf die Entstehung und Erhaltung der bilateralen Symmetric des
| |
| Froscheies. Arch. mik. Anat. Ix.
| |
| MOTTIER, D. M. (1897).
| |
| | |
| Beitriige zur Kenntniss der Kerntheilung in den Pollenmutterzellen. Jahrb. wiss. Bol., xxx.
| |
| NEMEC, B. (.1S99).
| |
| | |
| Ueber die karyokinetische Kerntheilung in den Wiirzelspitzen von Allium. Idem, xxxiii.
| |
| OSTERHOUT. \V. J. V. (1897).
| |
| | |
| Ueber Entstehung der karyokinetischen Spindel bei Equisetum. Idem, xxx.
| |
| PFLUGER, E. (18S3).
| |
| | |
| Ueber den Einfluss der Schwerkraft auf die Theilung der Zellen. Pfluger's Archiv., xxxi, xxxii.
| |
| RABL, C.'(1892, 1S96).
| |
| | |
| Theorie des Mesoderms. Morph. Jahrb., xv, xix, xxiv.
| |
| ROUX, W. (1883).
| |
| | |
| Ueber die Zeit der Bestimmung der Hauptrichtungen des Froschembryo. Gesammelte Werke, ii.
| |
| ROUX, W. (1885).
| |
| | |
| Ueber die Bestimmung der Hauptrichtungen des Froschembryo im Ei und iiber die erste Theilung des
| |
| Froscheies, Idem.
| |
| ROUX, W. (1887).
| |
| | |
| Die Bestimmung der Medianebene des Froschembryo d.urch die Copulationsrichtung des Eikerns und
| |
| des Spermakerns. Idem.
| |
| ROUX, \Y. (1S92).
| |
| | |
| Ueber das Entwicklungsmechanische Vermogen jeder der beiden ersten Furchungszellen des Eies.
| |
| Verh. d. Anal. Ges. zu Wien, 1892.
| |
| ROUX, W. (1895).
| |
| | |
| Einleitung. Arch. Entw. Mech., i.
| |
| ROUX, W. (1903).
| |
| | |
| Ueber die Ursachen der Bestimmung der Hauptrichtungen des Embryo im Froschei. Anat. Am., xxiii.
| |
| SAMASSA, P. (1894).
| |
| | |
| Zur Kenntniss der Furchung bei den Ascidien. Arch. mik. Anat. xliv.
| |
| SAMASSA, P. (1898).
| |
| | |
| Studien iiber den Einfluss des Dotters auf die Gastrulation und die Bildung der primaren Keimblatter
| |
| der W'irbelthiere. IY Amphioxus. Arch. Entw. Mech., vii.
| |
| SEELIGER, O. (1882
| |
| | |
| Zur Entuicklungsgeschichte der Ascidien. Sitzb. Akad. Wiss. Mien., Ixxxv.
| |
| SEELIGER, O. (1885
| |
| | |
| Die Entwicklungsgeschichte der Socialen Ascidien. Jena. Zeitschr., xviii.
| |
| SOBOTTA, J. (1S97).
| |
| | |
| Die Reifung und Befruchtung des Eies von Amphioxus. Arch. mik. Ana/., 1.
| |
| SOBOTTA. J. (1897 .
| |
| | |
| Beobachtungen iiber den Gastrulationsvorgang beim Amphioxus. Verh. Phys. Med. Ges. II Urzburg, xxxi.
| |
| VEJDOVSKY, Fr. (1888-1892).
| |
| | |
| Entwicklungsgeschichtliche Untersuchungen. Pi
| |
| | |
| | |
| WHEELER, W. M. (1897).
| |
| | |
| The Maturation, Fecundation and Early Cleavage in Myzostoma. Arch, dc Biol., xv.
| |
| WHITMAN, C. O. (1893).
| |
| | |
| The Inadequacy of the Cell Theory of Development. Woods Holl Biol. Lectures, 1893.
| |
| WILSON, E. B. (1892).
| |
| | |
| The Cell-Lineage of Nereis. Jour. Morph., vi.
| |
| WILSON, E. B. (1893).
| |
| | |
| Amphioxus and the Mosaic Theory of Development. Jour. Morph., viii.
| |
| WILSON, E. B. (1903).
| |
| | |
| Experiments on Cleavage and Localization in the Nemertine Egg. Arch. Entw. Meek., xvi.
| |
| WILSON, E. B. (1904).
| |
| | |
| Experimental Studies on Germinal Localization. Jour. Exp. Zoo/., i.
| |
| WILSON and MATHEWS (1895).
| |
| | |
| Maturation, Fertilzation and Polarity in the Echinoderm Egg. Jour. Morph., x.
| |
| WILSON, H. Y. (1900).
| |
| | |
| Formation of the Blastopore in the Frog's Egg. Anat. A?i~., xviii.
| |
| YATSU, N. (1904).
| |
| | |
| Experiments on the Development of Egg Fragments in Cerebratulus. Biol. Bull., vi.
| |
| ZIEGLER, H. E. (1902).
| |
| | |
| Lehrbuch der Vergleichenden Entwicklung^geschichte der niedereu Wirbelthiere. Jena.
| |
| ZUR STRASSEN, O. (1896).
| |
| | |
| Embryonalentwicklung der Ascaris megalocephala. Arch. Entw. Mcch , iii
| |
| | |
| | |
| | |
| ==Explanation Of Figures==
| |
| | |
| All figures represented in the plates were drawn with the aid of the camera lucida. Plates I-V
| |
| represent the living eggs of Oynthia (Styela) partita, and the colors used are in no sense diagrammatic
| |
| but are as nearly as possible those which appear in life; plate VI-X show fixed and stained' eggs of the
| |
| same species; plates XI anil XII, lixed and stained eggs of Citmn hitextintdk. The figures of the
| |
| living eggs of Cynthia i plates I-V) were drawn at the stage level under Zeiss Apochromatic Obj. 4
| |
| mm., Oc. 4. and are therefore magnified 250 diameters. With the exception of figs. 46-49 all the
| |
| details shown in these drawings were observed in living eggs without reference to fixed and stained
| |
| material; in the figures specified certain cell outlines were added to the drawings of the living eggs
| |
| after a study of prepared material. The figures of plates VI-XII, with the exception of figs. 61-75,
| |
| were drawn at the stage level under Zeiss Apochromatic Obj. 3 mm., Oc. 4; in the process of reproduction they have been reduced about one-fifth so that as they now appear they represent a magnification of about 266 diameters. Figs. 61-75, plate VI, were drawn under Zeiss Apochromatic Obj. 1.5
| |
| nun., Oc. 4, and therefore represent a magnification, after reduction, of about 535 diameters. The
| |
| chorion and test cells, though present in most of the eggs drawn, have been omitted from all the
| |
| figures, except those of plates I, VI and VII; the chorion, without the test cells, is shown in plates
| |
| II-V.
| |
| | |
| | |
| | |
| A.
| |
| a.p.bp,
| |
| ch.cn.c. p.Cr.
| |
| Cr. s,
| |
| | |
| D.
| |
| end.
| |
| f. c
| |
| g. v.kp.
| |
| mb.
| |
| m'ch.
| |
| ms.
| |
| n.
| |
| n. p.
| |
| | |
| | |
| Reference
| |
| | |
| -Anterior.
| |
| | |
| -Animal pole.
| |
| | |
| -Blastopore.
| |
| | |
| -Chorda
| |
| | |
| -Chorion.
| |
| | |
| -Clear protoplasm.
| |
| | |
| -Crescent of mesodermal substance (yellow
| |
| | |
| in Cynthia).
| |
| -Substance of crescent.
| |
| -Dorsal.
| |
| -Endoderm.
| |
| | |
| -Follicle cells (outside of chorion).
| |
| -Germinal vesicle.
| |
| -Karyoplasm (achromatic substance from
| |
| | |
| nucleus).
| |
| -Thick membrane which becomes chorion.
| |
| -Mesenchyme.
| |
| -Muscle cells.
| |
| -Nucleus.
| |
| -Neural plate.
| |
| | |
| | |
| | |
| Letters.
| |
| | |
| nl. Nucleolus.
| |
| n. t. Neural tube.
| |
| | |
| P. Posterior,
| |
| p. b. Polar body.
| |
| | |
| 1 p. b. First bolar body.
| |
| | |
| 2 p. b. Second polar body.
| |
| | |
| p. 1. Peripheral layer of protoplasm.
| |
| | |
| 1 P. S. First polar spindle.
| |
| | |
| 2 P. S. Second polar spindle.
| |
| | |
| Sn . Sperm atozoon .
| |
| t. c Test cells.
| |
| V. Ventral,
| |
| v. end. Ventral (caudal) endoderm.
| |
| v. p. Vegetal pole. y. h. Yellow hemisphere of egg.
| |
| | |
| ' yk. Yolk.
| |
| | |
| z Zwischenkorper (mid-body).
| |
| | |
| n. Sperm nucleus.
| |
| | |
| | |
| | |
| Nomenclature of Cells.
| |
| | |
| A Anterior half of egg; B Posterior half ; AB Right half; AB Left half ; A and A Right
| |
| and left anterior quadrants; B and B Right and left posterior quadrants. From the 8-cell stage
| |
| onward the cells of the eudodermal hemisphere are designated by capital letters, those of the ectodermal hemisphere by lower case. The first exponent indicates the cell generation, counting the unsegmented egg as the first ; the second exponent the position of the cell relative to the animal and vegetal
| |
| poles, cells which lie nearest the vegetal pole having the lowest exponent, those farthest away (nearest
| |
| the animal pole) having the highest. See p. 37.
| |
| | |
| | |
| | |
| Plate I.
| |
| Figures of the Living Egg* of Oynthia partita; Maturation and Fertilization.
| |
| | |
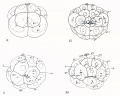
| Fig. 1. Unfertilized egg before the fading of the germinal vehicle, showing central mass of gray yolk,
| |
| peripheral layer of yellow protoplasm, test cells and chorion.
| |
| | |
| Fig. 2. .Similar egg after the disappearance of the nuclear membrane, showing the spreading of the
| |
| clear protoplasm of the germinal vesicle at the animal pole.
| |
| | |
| Fig. 3. Another egg about five minutes after fertilization, showing the streaming of the peripheral
| |
| protoplasm to the lower pole where the spermatozoon enters, thus exposing the gray yolk
| |
| of the upper hemisphere; the test cells are also carried by this streaming to the lower
| |
| hemisphere.
| |
| | |
| Figs. 4 and 5. Other eggs -bowing successive stages in the collection of the yellow and clear protoplasm at the vegetal pole; clear protoplasm lies beneath and extends a short distance
| |
| beyond the edge of the yellow cap.
| |
| | |
| Figs. 6-10. Successive stages of the same egg drawn at intervals of about five minutes; viewed from
| |
| the vegetal pole. In fig. 6 the area of yellow protoplasm is smallest, and the sperm
| |
| nucleus is a small clear area near its center. Figs. 7.-10 show stages in the spreading of
| |
| this yellow protoplasm until it covers nearly the whole of the lower hemisphere; at the
| |
| same time the sperm nucleus and aster move toward one side of the yellow cap and the
| |
| yellow protoplasm begins to collect into a crescent at this side.
| |
| | |
| Fig. 11. Side view of an egg of about the same stage as fig. 10, showing the eccentric position of the
| |
| sperm nucleus and a small area of clear protoplasm at the upper pole where the polar
| |
| bodies are being formed.
| |
| | |
| Fit;-. 12. Polyspermic (?) egg, viewed from the vegetal pole, showing four collections of yellow protoplasm around as many sperm (?) nuclei (see p. 24).
| |
| | |
| | |
| | |
| | |
| PLATE 1.
| |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| | |
| Plate II.
| |
| | |
| Living Eggs of Cynthia partita ; First Cleavage.
| |
| | |
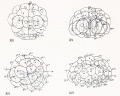
| Figs. 13 and 14. Side views of egg, showing the formation of the crescent (cr.) from the yellow hemisphere ; in all the figures the animal pole is above, the vegetal pole below. Above the
| |
| yellow crescent is an area of clear protoplasm (c. p.).
| |
| | |
| Fig. 15. Similar stage of another egg showing the aggregation of test cells over the crescent and the
| |
| protrusion of the chorion at this place, an unusual phenomenon.
| |
| | |
| Fig. 16. Egg showing crescent, viewed nearly from the vegetal pule
| |
| | |
| Fig. 17. Another egg showing crescent and clear protoplasm, viewed from posterior pole; the animal
| |
| pole is above, the vegetal below.
| |
| | |
| Figs. 18, 19, 20. Successive stages of the same egg drawn at intervals of about two minutes, showing
| |
| the approaching division of the egg.
| |
| | |
| Figs. 21, 22, 23, 24. Succesive stages in the first cleavage of an egg, drawn at intervals of about two
| |
| or three minutes. The egg is viewed from the posterior pole and shows the complicated
| |
| forms taken by the yellow crescent during the division, also the enlargement of the area
| |
| of clear protoplasm and its extension toward the animal pole.
| |
| | |
| | |
| | |
| | |
| Plate III.
| |
| | |
| Living Eggs of Oynthia partita; First to Fourth Cleavage.
| |
| | |
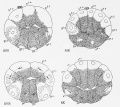
| Figs. 25 and 26- Same egg a* the one shown in figs. 21-24; final stages in the first cleavage.
| |
| | |
| Fig. 27. Another egg at the close of the first cleavage ; seen from the posterior pole.
| |
| | |
| Fig. 28. End view of egg of same stage as preceding, showing the lateral limits of the yellow crescent, the clear protoplasm in the upper hemisphere and the yolk in the lower. The
| |
| anterior portion of the lower hemisphere is composed of light gray material ; this is the
| |
| gray crescent and gives rise to chorda and neural plate.
| |
| | |
| Fig. 2'.). Four-cell stage, viewed from the animal pole.
| |
| | |
| Fig. 30. Similar egg seen from the vegetal pole; the crescent covers about half of the posterior
| |
| blastomeres.
| |
| | |
| Fig. 31. Fight-cell stage: the crescent is limited entirely to the two posterior blastomeres at the vegetal pole : while under observation the furrow between B 4 ' and b 4-2 shifted from the position indicated by the faint line to that shown by the heavy line, thus giving rise to the
| |
| "cross furrow" shown in the next figure.
| |
| | |
| Fig. 32. Fight-cell stage, viewed from the right side, showing a small amount of yellow protoplasm
| |
| around all the nuclei.
| |
| | |
| Fig. 33. Same stage viewed from the vegetal pole, showing 1% yolk laden endoderm cells and the
| |
| crescent.
| |
| | |
| Fig. 34. Same stage viewed from the posterior-animal pdreTshowing the clear ectodermal cells and
| |
| the crescent.
| |
| | |
| Fig. 35. Same stage seen from the anterior-vegetal pole; yellow protoplasm around all the nuclei.
| |
| | |
| Fig. 36. -Fourth cleavage of the egg seen from the vegetal pole.
| |
| | |
| | |
| | |
| | |
| Plate IV.
| |
| | |
| Living Eggs of Cynthia partita ; Fourth Cleavage to Gastrula.
| |
| | |
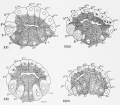
| Fig. 37. Sixteeu-cell stage viewed from vegetal pole.
| |
| | |
| Fig. 38. Sixteen-cell stage, from the animal pole, yellow protoplasm around the nuclei.
| |
| | |
| Fig. 39. Twenty-two cell stage, from the vegetal pole; four mesoderm cells (yellow), ten endoderm,
| |
| | |
| chorda and neural plate cells (gray) and eight ectoderm cells (clear).
| |
| Fig. 40. Same stage viewed from the posterior pole.
| |
| | |
| Fig. 41. Egg passing into the 32-cell stage; postero-dorsal (vegetal pole) view.
| |
| Fig. 42. Thirty-two-cell stage, postero-dorsal view.
| |
| Fig. 43. Forty-four- cell stage; posterior view, showing separation of mesenchyme (m'ch) from muscle
| |
| | |
| cells (ms.).
| |
| Fig. 44. Same stage, dorsal view, showing subdivision of endoderm cells.
| |
| Fig. 45. Similar stage, posterior view, showing separation of another mesenchyme cell from a muscle
| |
| | |
| cell.
| |
| Fig. 4(j. Seventy-four cell stage, dorsal view, showing division of 4 chorda and 4 neural piatc cells;
| |
| | |
| there are 10 mesenchyme and 6 muscle cells, besides 10 endoderm cells.
| |
| Fig. 47. One hundred and sixteen cell-stage, showing the beginning of gastrulation, also the neural
| |
| | |
| plate, chorda, muscle and mesenchyme cells.
| |
| Fig. 48. Slightly older stage showing advancing gastrulation with inrolling of cells at edge of blastopore.
| |
| | |
| | |
| | |
| | |
| Plate V.
| |
| | |
| Living Embryo* of Oynthia partita; Gastrula to Tadpole.
| |
| | |
| Fig. 49. Gastrula showing neural plate, chorda and muscle cells; about 176 cells, 96 ectoderm, 32
| |
| | |
| mesoderm, 20 endoderm, 8 chorda, 8 dorsal neural plate and 12 ventral neural plate cells.
| |
| Figs. 50 and 51. Two stages of one embryo, the second drawn about ten minutes after the first; the
| |
| | |
| yellow cells in the mid-line (m'ch.) are mesenchyme cells, the others (ms. ) muscle cells.
| |
| Fig. 52. Posterior view of elongated gastrula, the blastopore reduced to a narrow slit.
| |
| Fig. 53. Dorsal view of similar stage, the blastopore a small opening at the posterior end of a groove
| |
| Fig. 54. An older embryo, the blastopore covered by the forward growth of the posterior lips.
| |
| Fig. 55. Embryo with inrolling neural plate and with muscle cells arranged in three rows.
| |
| Fig. 56. Embryo of about the same stage as that shown in figs. 52 and 53; seen from the left side
| |
| | |
| showing neural groove, three rows of muscle cells which run transversely to the long
| |
| | |
| axis, and a row of mesenchyme cells in the long axis.
| |
| Fig. 57. Embryo viewed from posterior end, showing blastopore-raphe with open groove above and
| |
| | |
| with three rows of muscle cells on each side, also a row of mesenchyme cells.
| |
| Fig. 58. Young tadpole seen from dorsal side, neural groove open in front and closed behind, smallcelled mesenchyme in front of large muscle cells.
| |
| Fig. 59. Same stage as preceding seen from the right side, showing neural groove, mesenchyme and
| |
| | |
| three rows of muscle, cells.
| |
| Fig. 60. Tadpole of slightly older stage viewed from the posterior (caudal) end showing on each
| |
| | |
| side three large muscle cells each of which belongs to a row of such cells (<;/'. tig. 59).
| |
| | |
| These muscle cells are connected across the mid-line at the posterior end by a few
| |
| | |
| small mesenchyme cells.
| |
| | |
| | |
| | |
| PLATE VI.
| |
| | |
| SeHitiits of Eggs of Cynthia partita; Maturation and Fertilization.
| |
| Figs. 617S5 magnified 5S5 diameters; Figs. 70-87 magnified :.>t!<! diameters.
| |
| | |
| Fig. 61. Ovarian egg fully formed, showing germinal vesicle surrounded by yolk, and peripheral
| |
| | |
| layer of protoplasm containing test cells and yellow granules (small spheres in figure).
| |
| Fig. 62. Free egg shortly after the dissolution of the nuclear membrane, showing in the middle of the
| |
| | |
| clear karyoplasm fragments of nucleolus, chromosomes and a granular mass from which
| |
| | |
| spindle fibres arise; the peripheral protoplasm contains yellow granules.
| |
| Fig. 63. Egg similar to the preceding, but with the spindle fibres more fully formed.
| |
| Fig. 64. Similar to preceding, spindle fibres radiate in all directions.
| |
| | |
| Fig. 65. The first polar spindle lies near the surface of the egg and its fibres are approximately paratangential ; the peripheral layer of protoplasm has streamed away from the animal pole
| |
| | |
| and the karyoplasm from the germinal vesicle has spread out here in a broad disk.
| |
| Fig. 66. Metaphase of first polar spindle which is nearly parallel with surface; no centrosomes present.
| |
| Fig. 67. Anaphase of first polar spindle which is turning into a radial position.
| |
| Fig. OS. Separation of first polar body.
| |
| | |
| Fig. 69. Metaphase of second polar spindle, which is paratangential in position.
| |
| Fig. 70. Anaphase of second polar spindle.
| |
| Fig. 71. Second polar spindle approaching a. radial position.
| |
| Fig. 72. Separation of second polar body.
| |
| | |
| Fig. 73. Fusion of chromosomal vesicles in egg to form egg nucleus.
| |
| Fig. 74. Vegetal pole of egg of the stage shown in figs. 65 and 79, showing the entrance of the sperm
| |
| | |
| into the egg and the collection of yellow granules around the sperm head.
| |
| Fig. 75. Later stage in the entrance of the sperm ; formation of sperm aster from the middle- piece.
| |
| Fig. 76. Free egg before the solution of the nuclear membrane but after the extrusion of the tesl
| |
| | |
| cells; the chromosomes at the periphery of the germinal vesicle.
| |
| Fig. 77. Egg after being laid but before fertilization ; chromosomes and granular substance which
| |
| | |
| forms spindle fibres in the center of the karyoplasm. The egg remains in this condition
| |
| | |
| until fertilized.
| |
| Fig. 78. Same as preceding, save that spindle fibres are forming and karyoplasm has moved nearer to
| |
| | |
| the animal pole.
| |
| Fig. 79. Egg showing the entrance of the spermatozoon near the vegetal pole and the spreading of
| |
| | |
| the karyoplasm into a thin cap at the animal pole.
| |
| Fig. 80. Slightly more advanced stage showing development of sperm aster and collection of yellow
| |
| | |
| granules at vegetal pole, spermatozoa have entered some of the test cells.
| |
| Fig. 81. First polar spindle assuming a radial position ; increase of cytoplasmic area surrounding the
| |
| | |
| sperm nucleus and aster, the latter are moving across the egg axis and hence in the
| |
| | |
| longest path toward the equator.
| |
| Fig. 82. Stage slightly more advanced than the preceding; sperm nucleus, aster, clear and yellow
| |
| | |
| protoplasm becoming eccentric toward the posterior side.
| |
| Fig. 83. First polar body formed ; prophase of second polar spindle.
| |
| Fig. 84. Metaphase of second polar spindle; yellow protoplasm collecting into crescent.
| |
| Fig. 85. Anaphase of second polar spindle, spermatozoa in some of the test cells.
| |
| Fig. 86. Telophase of second polar spindle.
| |
| Fig. 87. Movement of sperm nucleus and aster and of surrounding protoplasm to the posterior side
| |
| | |
| of the egg; approach of the germ nuclei.
| |
| | |
| | |
| | |
| PLATE VII.
| |
| | |
| Sections of Eggs of Cynthia partita; Fertilization and Early Cleavages.
| |
| | |
| Fig. 88. Division of sperm aster ; crescent substance at periphery on posterior side.
| |
| | |
| Fig. 89. Similar to preceding but showing egg nucleus.
| |
| | |
| Figs. 90 and 91. Sections at right angles to first cleavage plane but oblique to egg axis, showing the
| |
| union of pronuclei; clear and yellow protoplasm and sperm amphiaster in posterior half
| |
| of egg.
| |
| | |
| Fig. 92. Section in the plane of the first cleavage (future median plane) showing the union of the
| |
| germ nuclei, the clear protoplasm and crescent on the posterior side of the egg, and the
| |
| polar bodies above.
| |
| | |
| Fig. 93. Early prophase of the first cleavage, in the plane of the spindle axis.
| |
| | |
| Fig. 94 and 95. Two dispermic eggs; the first showing two sperm nuclei on the posterior side of the
| |
| egg with a single crescent ; the second, two sperm nuclei and one egg nucleus, with the
| |
| clear protoplasm and crescent about equally divided, by a tongue of yolk, between the
| |
| two sperm nuclei.
| |
| | |
| Fig. 96. Section in the plane of the first cleavage and transverse to the spindle, showing a cross section of the equatorial plate near the middle of the egg, surrounded by clear protoplasm,
| |
| also the crescent at the posterior border with some of the yellow spherules all around
| |
| the spindle.
| |
| | |
| Fig. 97. Metaphase of the first cleavage; equatorial section at right angles to the preceding.
| |
| | |
| Figs. 98 and 99. Early and late anaphases of the first cleavage; sections in equatorial plane.
| |
| | |
| Fig. 99. Constriction of cell body; chromosomal vesicles at the ends of the nuclear spindle. In both
| |
| | |
| figures the daughter centrosi is are elongating in the equatorial plane and at right
| |
| | |
| angles to the first spindle axis.
| |
| | |
| Fig. 100. Telophase of the first cleavage, showing the rotation of nuclei, eentrosomes and cytoplasm
| |
| toward the animal pole of the egg.
| |
| | |
| Fig. 101. Equatorial section, prophase of second cleavage; crescent substance at the posterior pole.
| |
| | |
| Fig. 102. Section at right angles to the preceding and through the axis of one of the spindles; nucleus
| |
| and cytoplasm lie above the equator, yolk and crescent below.
| |
| | |
| Fig. 103. Equatorial section showing precocious division of the eentrosomes in one of the blastomeres.
| |
| | |
| Fig. 104. Anaphase of second cleavage; equatorial section ; chromosomal vesicles at ends of nuclear
| |
| spindles.
| |
| | |
| Fig. 105. Telophase of second cleavage ; equatorial section, showing the bending of the middle of the
| |
| spindle toward the center of the egg, and a large amount of clear cytoplasm aud of
| |
| crescent substance in the posterior quadrants.
| |
| | |
| Figs. 106 and 107. Two sections through one and the same egg in the eight-cell stage ; the first
| |
| through the anterior blastomeres ; the second through the posterior. The cytoplasm is
| |
| most abundant in the cells at the animal pole, the crescent substance in the two posteriorvegetal cells, though it is also found around all the nuclei. The polar bodies, which are
| |
| shown in stippled outlines, are not in the plane of either of these sections, but in the
| |
| region between them.
| |
| | |
| | |
| | |
| PLATE VIII.
| |
| | |
| Surface Views of Entire Eggs of Cynthia partita; Eight to Forty-four Cells.
| |
| | |
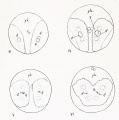
| Fig. 108. Eight-cell stage ; left side of egg ; showing spindles of third cleavage.
| |
| | |
| Fig. 109. Anterior view of 8-cell stage, showing cytoplasm most abundant in the animal pole cells,
| |
| | |
| and the yolk largely collected in the anterior cell of the vegetal hemisphere.
| |
| Figs. 110, 111,112. Stages in the fourth cleavage; figs. 110 and 112 viewed from the animal pole, fig.
| |
| | |
| Ill from the. vegetal pole.
| |
| Fig. 113. Telophase of fourth cleavage, vegetal poje view ; caps of deeply staining protoplasm lie at
| |
| | |
| the hinder borders of the small posterior cells (B 5,2 ).
| |
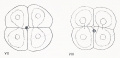
| Figs. 114 and 115. Anterior and posterior views of the 16-cell -stage; fig. 115 showing caps of
| |
| | |
| deeply staining protoplasm at the posterior pole, which later go into the posterior mesenchyme cells (B' 6 , figs. 130, 131).
| |
| Figs, lit! and 117. Ventral and dorsal views of a 20-cell stage, showing the cells at the vegetal pole
| |
| | |
| dividing before those at the animal pole.
| |
| Fig. 118. Slightly older stage with some of the animal pole cells dividing.
| |
| Figs. 119-123. Five views of one and the same egg; fig. 119, ventral; 120, dorsal; 121, anterior;
| |
| | |
| 122, posterior; 123, right side; the latter shows in dotted outlines the great elongation
| |
| | |
| of the cells at the animal pole and the flattened shape of the cells at the vegetal pole ;
| |
| | |
| all the designations of cells in tig. 123 should be underscored ; 44 cells, 16 ectoderm, 10
| |
| | |
| endoderm, 10 mesoderm, 4 chorda and 4 neural plate cells.
| |
| Figs. 124-129. Six different views of one and the same egg in the 44-cell stage showing the divisions
| |
| | |
| of the ectodermal cells and the second cells of the crescent (B 6 ' 4 ) ; when these divisions
| |
| | |
| are completed there will be 62 cells. Fig. 124, ventral; 125, dorsal; 126, anterior; 127,
| |
| | |
| postero-dorsal.
| |
| | |
| | |
| | |
| | |
| | |
| Plate IX.
| |
| | |
| Surfact Views oj Entire Eggs of Cynthia partita; Forty-four to Tiro Hundred and Eighteen Cells,
| |
| | |
| Figs. 128, 129. Same egg as shown in figs. 124-127. Fig. 128 from the left side; the equator of the
| |
| egg (plane of the third cleavage) is the heavy line running between A ami 1' and
| |
| separating cells designated by lower case from those designated by capital letters. Fig.
| |
| 129. View from the posterior pole.
| |
| | |
| Figs. 130 and 131. Ventral and dorsal views respectively of one and the same egg; 64 cells, 32 in
| |
| each hemisphere, distributed as follows: Ventral hemisphere, 26 ectoderm, 6 neural plate
| |
| cells (a 7 -', a 7 -', a 7- ' 3 ) ; Dorsal hemisphere, 10 endoderra, 4 chorda (A 73 , A 77 ), 4 neural plate
| |
| (A'-, A 7 - 8 ), 10 mesenchyme (B 7 - 6 , B 7 5 , B", B 73 , A 7 ' 6 ) and 4 muscle (B 78 , B 7 ->).
| |
| | |
| Fig. 132. Postero-dorsal view of an egg in the same stage as the preceding showing direction of
| |
| division of mesenchyme cell (B 73 ).
| |
| | |
| Figs. 133 and 134. Ventral and dorsal views respectively of one and the same egg; 110 cells; Ventral hemisphere 64 cells, 52 ectoderm, 12 neural plate (a 8 ' 9 , a 8= , a 8 ' 7 , a 8 ' 8 , a 8,25 , a 8=6 ) ;
| |
| Dorsal hemisphere 46 cells, 10 endoderm, 8 chorda (A 85 , A 86 , A 8 - 13 , A 8 ' 4 ), 8 neural plate
| |
| (A 8 - 7 , A 8 - 8 , A 8 -' 3 , A 8 ' 6 ), 12 mesenchyme (B 7 - 6 , B", B 7 7 , B 8 ', B 86 , A 7 - 6 ), 8 muscle (B 8 - 7 , B 88 ,
| |
| B 8 ' 5 , B 8 " 6 ). Gastrulation has begun.
| |
| | |
| Fig. 135. Dorsal view of a slightly more advanced stage showing increasing gastrulation ; 118 cells,
| |
| ventral hemisphere 64 cells, dorsal hemisphere 54, viz., 10 endoderm, 16 mesenchyme, 12
| |
| muscle, 8 chorda, 8 neural plate; when the divisions indicated by spindles are completed
| |
| there will 4 additional endoderm and 2 additional mesenchyme cells.
| |
| | |
| Figs. 136-139. Four views of one and the same egg; fig. 136 dorsal, 137 posterior, 138 anterior, 139
| |
| ventral; gastrulation well advanced. 124 cells; Ventral hemisphere 64 cells, 52 ectoderm, 12 neural plate (a 8 -'", a 8 -*", a 8 " 7 , a 8 ' 8 , a 8 - 23 , a 6 - 56 ); Dorsal hemisphere 60 cells, 14
| |
| endoderm, 8 chorda, 8 neural plate, 18 mesenchyme, 12 muscle. Spindles are already
| |
| present for divisions, which, when completed, will lead to 178 cells, viz., 96 ectoderm, 12
| |
| neural plate of ventral hemisphere, 12 neural plate of dorsal hemisphere, 8 chorda. 211
| |
| mesenchyme, 12 muscle, 18 endoderm.
| |
| | |
| Figs. 140-143. Four views of one and the same egg; 140 dorsal, 141 posterior, 142 anterior, 143
| |
| ventral. Gastrulation is here far advanced. 180 cells; Ventral hemisphere 108 cells,
| |
| 96 ectoderm, 12 neural plate; Dorsal hemisphere 72 cells, 12 neural plate of dorsal hemisphere, 8 chorda, 20 mesenchyme. 12 muscle, 20 endoderm; when the divisions indicated
| |
| by spindles are completed there will be 4 additional neural plate cells.
| |
| | |
| Figs. 144 and 145. Two views of the same egg; 144 dorsal, 145 median optical section in sagittal
| |
| plane. In fig. 145 the polar body is not visible, but its supposed position is indicated by
| |
| the dotted outline at the animal pole; the dotted outlines at the lower pole indicate the
| |
| mesoderm cells which lie in the lateral lip of the blastopore and out of the plane of the
| |
| section ; the rolling in of the muscle cells in the lateral lips is well shown.
| |
| | |
| Fig. 146. Anterior view of an egg of about the same stage as the preceding, showing the division of
| |
| the 12 neural plate cells of the ventral hemisphere.
| |
| | |
| Fig. 147. Ventral view of a similar stage with the ectoderm omitted in order to show the endoderm
| |
| and mesoderm from the ventral side. At this stag.' all the ectoderm cells have passed
| |
| into the 9th generation, all the endoderm into the 8th or 9th, all the mesoderm except
| |
| IV" into the 8th or 9th, all the chorda and neural plate cells into the 9th. There
| |
| are 218 cells; Ventral hemisphere 128 cells, 104 ectoderm, 24 neural plate of ventral
| |
| hemisphere; Dorsal hemisphere 90 cells, viz., 16 mural plate, 16 chorda, 20 mesenchyme,
| |
| 12 muscle, 26 endoderm.
| |
| | |
| | |
| Plate X.
| |
| Entire Embryos of Cynthia partita; Gastrula to Tadpole.
| |
| | |
| Muscle cells shaded by vertical lines, mesenchyme by transverse line*, nerve plate and tube by fine
| |
| | |
| stipples, chorda by coarse stipples.
| |
| | |
| Figs. 148-151. Four view? of the same embryo; fig. 14.8 dorsal, 149 ventral -posterior, 150 same view
| |
| but deeper level, showing mesoderm and endoderm, 151 right side. Many of the ectoderm cells are passing into the 10th generation.
| |
| | |
| Figs. 152-154. Three views of : embryo ; fig. 152 dorsal, showing closure of the blastopore from in
| |
| | |
| front, 153 same view but deeper level, showing cells beneath the ectoderm, 154 ventral
| |
| view of mesoderm and endoderm below ectoderm.
| |
| | |
| Figs. 155 and 156. Two ventral views of an advanced gastrula, the first showing the superficial ectoderm, the latter the endoderm and mesoderm lying beneath the ectoderm.
| |
| | |
| Fig. 157. Left side of embryo, showing, in optical section, muscle cells, mesenchyme, chorda and nerve
| |
| plate.
| |
| | |
| Figs. 158 and 159. Two optical sections of the same embryo, the former (158) through the open blastopore at the posterior end of the embryo, the latter (159) through the region anterior to
| |
| the blastopore.
| |
| | |
| Figs. 160-162. Three views of the same embryo from the ventral side but drawn at different levels;
| |
| 160 surface view showing ectoderm ; 161 same view deeper level, showing mesoderm and
| |
| endoderm : 162 same view still deeper level, showing chorda, mesoderm and endoderm.
| |
| | |
| Fig. 163. Dorsal view of embryo of same stage as preceding showing nerve plate and tube, mesoderm
| |
| and chorda.
| |
| | |
| Figs. 164 and 165. Two views of same embryo from right side ; fig. 164 median optical section showing chorda, nerve tube and endoderm ; 165 more superficial view showing muscle cells
| |
| and mesenchyme.
| |
| | |
| Fig. 166. Older embryo from right side, showing in optical section nerve tube, chorda, mesoderm and
| |
| endoderm.
| |
| | |
| Fig. 167. Young tadpole from left side showing in optical section nerve tube, chorda and mesoderm.
| |
| | |
| | |
| | |
| PLATE XL
| |
| | |
| Eggs of Ciona intestinalis ; Ovocyte to Fourth Cleavage.
| |
| | |
| Figs. 168-179 actual section*; figs. 180-187 whole eggs. Crescent substance and clear protoplasm
| |
| stippled. Polar bodies are actually present where drawn.
| |
| | |
| Fig. 168. Half grown ovarian egg, showing test cells within the egg cytoplasm, which is composed of
| |
| | |
| a layer of yolk surrounding the nucleus and a peripheral layer of clear protoplasm ;
| |
| | |
| around the whole egg is a layer of small follicle cells which later develop into the very
| |
| | |
| large conical cells which surround the chorion.
| |
| Fig. 169. Older ovarian egg, showing the area of the yolk increased and the test cells limited to the
| |
| | |
| peripheral layer of protoplasm, which is sharply differentiated from the yolk.
| |
| Fig. 170. Still older ovarian egg, showing the test cells arranged in groups or "nests" at the periphery
| |
| | |
| of the egg.
| |
| Fig. 171. Ripe ovarian egg ready to escape from the ovary, showing the extrusion of the test cells and
| |
| | |
| the formation of a thick homogeneous membrane around the egg, which ultimately
| |
| | |
| becomes the chorion.
| |
| Fig. 172. Free hut unfertilized egg which lias shrunken away from the chorion, showing the first
| |
| | |
| maturation spindle, the peripheral layer of protoplasm collected over the lower pole of
| |
| | |
| the egg and the achromatic substance of the germinal vesicle (kp) spread in a broad
| |
| | |
| layer over the upper pole and around the central yolk.
| |
| Fig. 173. Extrusion of first polar body and entrance of spermatozoon ; the sperm nucleus lies in the
| |
| | |
| peripheral layer surrounded by clear protoplasm in which astral rays are developing.
| |
| Fig. 174. Nearly equatorial section of the egg showing sperm nucleus and amphiaster at the posterior
| |
| | |
| side of the egg; at the surface is the granular protoplasm of the peripheral layer.
| |
| Fig. 175. Section of egg in plane of first cleavage (median plane), showing the approach of the germ
| |
| | |
| nuclei and the movement of the peripheral layer of protoplasm from the lower pole to
| |
| | |
| the posterior side to form the crescent ; in the darkly staining substance of the crescent
| |
| | |
| is a clear triangular area which corresponds to the clear area surrounding the sperm
| |
| | |
| nucleus in fig. 173.
| |
| Fig. 176. Section of slightly older stage in plane of first cleavage, showing the union of the germ
| |
| | |
| nuclei ; polar body out of the plane of section.
| |
| Fig. 177. Anaphase of first cleavage, showing the complete separateness of the nuclear and astral
| |
| | |
| portions of the mitotic figure; the crescent substance shows at the ends of the spindle.
| |
| Fig, 178. Telophase of first cleavage, showing " zwischenkorper " (z), also the bending of spindle axis
| |
| | |
| and shifting of cytoplasm and nuclei toward the animal pole.
| |
| Fig. 179. Anaphase of second cleavage, showing separateness of nuclear and astral portions of mitotic
| |
| | |
| figure, also position of crescent on postero- dorsal side ; polar body out of the plane of
| |
| | |
| section.
| |
| Fig. 180. Entire egg of Ciona, showing formation of second polar spindle, peripheral layer of protoplasm, sperm nucleus and aster.
| |
| Fig. 181. Anaphase of first cleavage viewed from posterior pole.
| |
| Fig. 182. Two-cell stage seen from posterior pole, cytoplasm and nuclei lie near animal pole, crescent
| |
| | |
| near vegetal,
| |
| fig. 183. Four-cell -stage seen from vegetal pole ; the crescent covers about one half the surface of the
| |
| | |
| two posterior blastomeres.
| |
| Fig. 1S4. Eight cell-stage, left side, showing cap of deeply staining protoplasm at posterior pole of
| |
| | |
| cells (B 4-1 ), which later goes into the posterior mesenchyme cells (B 7 ' J , fig. 200.)
| |
| Fig. 185. Eight cells, posterior view; spindles of the fourth cleavage present.
| |
| Fig. 186. Fourth cleavage, vegetal pole.
| |
| Fig. 187. Fourth cleavage, animal pole; 16 cell.-.
| |
| | |
| | |
| | |
| | |
| | |
| PLATE XII.
| |
| | |
| Whole Eggs of Ciona intestinalis ; Sixteen Cells to Gastrula.
| |
| | |
| Crescent substance stippled in nil the figures; clear protoplasm also stippled in figs. 188-191.
| |
| Pobty bodies are actually present where drawn.
| |
| | |
| Fig. 188. Sixteen cell stage, anterior pole.
| |
| | |
| Fig. 189. Sixteen cells seen from vegetal pole; all but two of the cells are passing from the 5th to the
| |
| 6th generation.
| |
| | |
| Fig. 190. Similar stage viewed from animal pole.
| |
| | |
| Fig. 191. Animal pole view, 5th to the 6th generation; 20 cells; when the divisions are completed,
| |
| which are here indicated by spindles, there will be 32 cells.
| |
| | |
| Figs. 192-195. One and the same egg in the 32-cell-stage. Fig. 192. From animal pole. Fig. 193.
| |
| From vegetal pole, spindles present in many of the cells, which are about to pass into the
| |
| 7th generation. Fig. 194. From posterior pole; all nuclei at the animal pole in a
| |
| resting condition. Fig. 195. Antero-vegetal view.
| |
| | |
| Figs. 196 and 197. One and the same egg seen from the animal (ventral) and from the vegetal (dorsal) poles; most of the cells at the vegetal pole have just passed into the 7th generation ;
| |
| spindles are present in all the cells at the animal pole preparatory to their division into
| |
| the 7th generation ; 44 cells, 20 of the 6th and 24 of the 7th generation.
| |
| | |
| Figs. 198-204. With the exception of fig. 199, all these figures represent views of one and the same
| |
| egg. Fig. 198. View from animal pole showing all the ectoderm cells in process of
| |
| division. Fig. 199. Another egg of the same stage as the preceding, but seen from a
| |
| more anterior point of view. Fig. 200. Vegetal pole showing all the cells at this pole
| |
| in a resting condition. Fig. 201. Same egg seen from a more anterior point of view.
| |
| Fig. 202. Same egg viewed from the anterior pole. Fig. 203. Viewed from the right
| |
| side showing in dotted outline the great elongation of the cells of the vegetal pole and a
| |
| corresponding flattening of the cells at the animal pole. Fig. 204. Viewed from the
| |
| posterior pole. 76 cells, viz., 32 ectoderm, !< endoderm, 18 mesoderm, 8 chorda, 8 neural
| |
| plate; when the divisions indicated in the ectoderm cells are completed there will be 64
| |
| ectoderm cells and 108 cells in all.
| |
| | |
| Fig. 205. Gastrula corresponding in stage to fig. 140 of the Cynthia egg; 178 cells, vis., 108 ectoderm
| |
| and neural plate (ventral), 20 endoderm, 18 mesenchyme, 12 muscle, 8 chorda, 12 neural
| |
| plate (dorsal).
| |
| | |
| Fig. 206 and 207. Older gastrula with blastopore much restricted by the growth of the anterior lip
| |
| Fig. 206. View of superficial ectoderm and mesoderm. Fig. 2(17. Deeper view of
| |
| same embryo showing endoderm and mesoderm.
| |
| | |
| | |
|
| |
|
|
| |
|