Paper - The Development of the Pars Membranacea Septi in the Human Heart
| Embryology - 24 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Odgers PN. The development of the pars membranacea septi in the human heart. (1938) J Anat. 72(2): 247-59. https://www.ncbi.nlm.nih.gov/pubmed/17104688 PMID 17104688]
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
The Development of the Pars Membranacea Septi in the Human Heart
By P. N. B. Odgers
From the Department of Human Anatomy, University of Oxford (1938)
THE accounts which have been published of the development of this region of the heart, the method in which the interventricular foramen is occluded to form the membranous part of the septum, or whether indeed this occurs at all, have been so vague or so contradictory that a re-examination of the question seemed worth while.
For this investigation I was able to study human embryos of 11.2, 11.4-, 12.5, 14, 14.5, 15.5, 17, and 17.5 mm. in crown-rump length. Of these the 14 mm. embryo was the least valuable for my present purpose, as the heart is over-distended with blood in this specimen. I have therefore not included it among the detailed descriptions which follow. Reconstruction models were made of this region in the 11.2, 11.4, 14.5, 15.5, and 17 mm. specimens.
11.2 mm Embryo
At this stage (Text-fig. 1) the two bulbar ridges (Frazer) or the proximal bulbar swellings (Tandler) are seen united above to form the bulbar septum, but soon separating below into right and left moieties. The right one (R.B.R.) can be traced down on the right ventricular wall to become continuous with the anlage of the right or inferior cusp of the tricuspid valve, while the left one (L.B.R.) joins the right side of the interventricular septum and on that soon disappears. Where they are fused and for a few sections after their separation the two ridges are seen to be of equal size, but at a. lower level the right one soon appears to be not only the more extensive but the more substantial of the two.
The interventricular foramen (I. V.F.) is seen in thirty sections, i.e. its cranio-caudal diameter is 300 p.. It is bordered ventrally by the inter ventricular septum (I. V.S.) and dorsally by the fused auriculoventricular cushions (hereafter abbreviated into the A.-V. cushions). Above, the dorsal (cranial) border of the interventricular septum joins the fused A.-V. cushions about their centre, while below it joins their right margin.
Below the caudal border of the foramen the septum is fused to the inferior A.-V. cushion in fourteen sections (i.e. for 140 p.). This must presumably represent the amount of closure of the interventricular foramen which has already occurred since about the 7 mm. stage. It may be noted here, however, that in our 7 mm. embryo, where the interventricular septum is represented by a
low ridge which does not yet make contact with the A.-V. cushions, the cranio-caudal -measurement of the interventricular foramen is only 220 μ in contrast to 300 μ in the specimen under consideration. This illustrates how difficult it is to compare accurately the condition of any single region in different embryos. There cannot be in the nature of things any constant initial size of foramen to be obliterated. Secondly, the foramen may not be diminishing steadily with the age of the embryo. Rather is it reasonable to suppose that it shares in the general growth and that its gradual closure is a compromise between two competitive processes.

Text-fig. 1. Drawing of a model of this region of the heart in an 11-2 mm. embryo ( x 100) after removal of the ventral wall of the bulbus and right ventricle and of ahnost the whole of the interventricular septum, viewed from the right side. R.B.R. right bulbar ridge; L.B.R. left bulbar ridge; R.S.T. right superior tubercle of the A.-V. cushions; R.I.T. right inferior tubercle of the A.-V. cushions; I. V.S. interventricular septum; I . V.F. interventricular foramen; R.A.V. right auriculoventricular orifice; A.0. aortic channel; P.A. pulmonary artery.
It is interesting to observe the method of closure here. In the three sections immediately below the foramen this is seen to be taking place by a narrow tongue—shaped proliferation of the right border of the A.-v. cushion making contact with and extending Ventralwards on the right side of the septum (Pl. I, fig. 1). From the left margin of this proliferation contact with the septum gradually broadens, but there are ten sections below the foramen before the dorsal border of the interventricular septum and the A.-V. cushion are joined.
The right and left edges of the fused A.-V. cushions are seen to project as prominent swellingsto form the medial boundaries of the right and left auriculoventricular orifices respectively. Further, in both these swellings superior and inferior tubercles can be distinguished corresponding presumably to the original dorsal and ventral components of the fused cushions. Over the upper border of the right superior tubercle (R.S.T.), which is well cranial to the upper limit of the right auricular opening, the aortic channel passes to gain the left ventricle through the upper and dorsal portion of the interventricular foramen above the level of the lowest attachment of the left bulbar ridge to the interventricular septum.
11.4 mm Embryo
I suspect that this embryo is older than its crown-rump length suggests. There is no record of its length before fixation, and in estimating this from an enumeration of the sections I expect an insuflicient allowance was made for shrinkage. The model (Text-fig. 2) begins below the level of the bulbar septum and the two bulbar ridges appear quite separated. The left one (L.B.R.) is seen, as before, to join the interventricular septum well to the right side and ventral to its dorsal border. On this it soon disappears. The right ridge (R.B.R.) is the more prominent of the two and, as it is traced caudalwards, is found to become continuous with the projection of the commencing anterior cusp of the tricuspid valve. Between the two bulbar ridges are seen the marked elevations of the right tubercles of the fused A.-V. cushions, and now the separation between the superior (R.S.T.) and inferior (R.I.T.) is much more pronounced than in the earlier embryo.

Text-fig. 2. Drawing of a model of this region of the heart in an 11-4 mm. embryo ( x 100) after removal of the ventral wall of the right ventricle and the greater part of the inter ventricular septum, viewed from the right side. R.B.R. right bulbar ridge; L.B.R. left bulbar ridge; .R.S.T. right superior tubercle of the A.-V. cushions; R.I.T. right inferior tubercle of the A.-V. cushions; I . V.S. interventricular septum; I. 17.1‘. interventricular foramen; R.A. V. right auriculoventricular orifice.
The relation of the right bulbar ridge to these tubercles must be described in some detail. While superiorly the two structures are quite separate, farther down the left margin of the right bulbar ridge gradually joins the right side of the lower portion of the superior and the upper portion of the inferior tubercle. In this way a part of the bulbar cavity is cut off and is found to be continuous below with that of the right auricle. This must mean that the right bulbar ridge has grown across the upper end of the right auriculoventricular orifice (R.A. V.) to meet the right A.-V. tubercles. In conformity with this, while the latter orifice measures 190 p. in its vertical extent in the 11.2 mm. embryo, it is now only patent for 40 μ. The interventricular foramen (I. V.F.) is very much smaller than in the earlier specimen. It is now only seen in eight sections, i.e. its craniocaudal diameter is only 80p.. Caudal to its lower margin the ventricular septum and the inferior A.-V. cushion are fused for 320 μ in contrast to 140 μ in the first embryo. Further, a comparison with the 11.2 mm. specimen shows that the foramen is not only less extensive but is more obviously divided now into two parts. That portion of it cranial to and dorsal to the lower end of the left bulbar ridge is certainly forming the aortic channel, while the caudal and ventral part of it is being gradually obliterated. The closure of the foramen can be observed to be taking place here by the proliferation of the inferior tubercle in a cranial direction and it is this proliferation which makes the sulcus, separating the two tubercles, so much more pronounced (Pl. I, fig. 2). The microphotograph is that of the fourth section cranial to the foramina closure and shows the rapidly extending cushion tissue bare of endocardium in contrast to the interventricular septum, which is still covered by it.
14.5 mm Embryo
(Text-fig. 3).
The bulbar septum is now very much more extensive and forms the cranial border of the still patent interventricular foramen. The junction between its two component ridges can be distinguished as a more deeply staining seam, and by tracing this down it can be seen that it is the left bulbar ridge (L.B.R.) which joins the interventricular septum, as it does in the earlier embryos, but now it does so practically at its dorsal border and nothing like so much to its right side compared with the 11-4 mm. specimen. It soon fades out on the septum and does not appear to contribute any cushion tissue to it or to take any part in closing the foramen. The right bulbar ridge (R.B.R.), as it is traced caudalwards, is much the more extensive of the two. Just above the level of the interventricular foramen its left border is seen to be forming the greater part of the right wall of the aortic channel, while its right border bulges out prominently to form the anterior cusp of the tricuspid valve. Projecting into the aortic channel (A.0.) from its dorsal wall is the superior portion of the right tubercle of the A.-V. cushions (R.S'.T.) ; it is impossible now to define accurately the division between superior and inferior tubercles. The interventricular foramen (I.V.F.) is patent in four sections (20 μ in thickness). It is bounded now cranially, as has been said, by the bulbar septum, caudally by A.-V. cushion tissue, on its dorsal border by the right tubercle of the A.-V. cushions (R.S.T.) and on its ventral border by the inter ventricular septum (I. V.S.). The microphotographs show how the foramen is being closed. In P1. I, fig. 6, cushion tissue is seen proliferating from the A.-V. cushion, while a small disk-shaped mass of similar tissue caps the interventricular septum to the right of its dorsal border. In the next sections below this one this proliferation becomes more extensive until it reaches and fuses with the tissue capping the septum (Pl. I, fig. 7), so that the succeeding sections show a single fused mass joining the A.-V. cushion to the septum.

Text-fig. 3. Drawing of a model of this region of the heart in a 14.5 mm. embryo ( x 100) viewed obliquely from above and from the left side. R.B.R. right bulbar ridge; L.B.R. left bulbar ridge; R.S.T. right superior tubercle of A.-V. cushions; I. V.S. interventricular septum; I . V.F interventricular foramen; A.0. aortic vestibule; RA .0. bulbo-auricular channel.
The only interpretation which can be put on these appearances is that proliferating A.-V. cushion tissue is growing up from below on the ventral (septal) boundary of the interventricular foramen. These facts are shown in Text-fig. 4, which is a diagrammatic enlargement in silhouette of this region.
Further, P1. I, fig. 7, shows that just at the caudal boundary of the foramen the left border of the right bulbar ridge is joining the tissue proliferating from the A.-V. cushion rather than the cushion itself as it was seen to do in the 11.4. mm. embryo. In either case, however, this junction is cutting off a bulbo-auricular channel which is far larger in thepresent specimen than in any of my others and which still communicates inferiorly with the right auricle.
12.5 mm Embryo
In this specimen, while the interventricular foramen is more extensive, 180 μ in its vertical diameter, in some particulars the development of this region is more advanced. The right bulbar ridge and the superior tubercle of the A.-V. cushions, which seems to be poorly developed here, have blended above. It is not until the level of the first section which shows the closure of the interventricular foramen that a narrow cleft is seen to be separating the two (Pl. I, fig. 3). When this narrow cleft is traced caudalwards through the succeeding four sections (40 μ) it is found to communicate with the right auricle and this is all that is seen of the much larger bulbo-auricular channel of the 14-5 mm. embryo.
A microphotograph of the next section below the first one (Pl. I, fig. 4-) shows that the right bulbar ridge is in close coaptation at once with the right tubercle of the A.-V. cushions, with cushion tissue capping the interventricular septum, and with cushion tissue connecting them.

Text-fig. 4. A.V.C. a.-v. cushions from which tissue is proliferating to fill up from below the inter-ventricular foramen, which is black, and to climb up on the dorsal border of the interventricular septum, I . V.S. R.B.R. right bulbar ridge joined with the left bulbar ridge, L.B.R., to form the bulbar septum and the cranial boundary of the foramen. R.B.R.’ is the lower margin of the right bulbar ridge at its junction with the proliferating A.-V. cushion tissue.
In the next three sections the connexion between the right tubercle and the cushion tissue capping the septum gradually seems to be more and more pronounced. They are still demarcated, however, from each other by a distinct notch on either side (Pl. I, fig. 5). This notch becomes indistinct in the course of the next few sections and then the A.-V. cushions appear as a uniform mass in contact with the interventricular septum. The origin of the horn-shaped cap of cushion tissue on the interventricular septum (Pl. I, fig. 3), which can be recognised in the three sections cranial to this level, might be thought to be doubtful from the appearances presented by the microphotographs (P1. I, figs. 3-5). In this series it is seen to be in contact with the right bulbar ridge 10 μ rostral to the level at which it is connected with the right A.-V. cushion, but two facts make its origin from the latter certain. As it is traced caudal-wards it obviously increases in size, which would surely be impossible if it were a derivative of the bulbar ridge. Further it is found to actually fuse with the right A.-V. cushion, while it is simply in coaptation with the right bulbar
ridge, which does not appear to contribute anything to its formation.
I think therefore it can be truly regarded as having proliferated from the A.-V. cushions and the appearances presented here thus fall into line with those exhibited in the previous, 14-5 mm., specimen.
15.5 mm Embryo
As in the 14-5 mm. embryo, the bulbar septum is seen here to form the cranial boundary of the interventricular foramen. The seam between its two constituents is distinct here, and the left bulbar ridge fuses with the dorsal border of the interventricular septum (Pl. II, fig. 8) and is contiguous with the cushion tissue which caps it (Pl. II, fig. 9). The foramen is now 30 ft in its craniocaudal diameter.
Above the level of the interventricular foramen the right bulbar ridge and
the right superior tubercle of the A.-V. cushions are completely fused, and it is
only at the caudal border of the foramen that a cleft appears which separates
them (Pl. II, fig. 10). While this cleft separates the right bulbar ridge from the
A.-V. cushion, it still permits the former to meet the cushion tissue which has
proliferated to fill the foramen. As it is traced caudalwards the cleft gradually
becomes wider and ends in the posterior portion of the right auriculoventri-
cular orifice. The condition found here is almost exactly similar to that seen
in the 12-5 mm. specimen and represents the remains of the bulbo-auricular
channel of the earlier embryos, but owing apparently to a gradual extension
cranialwards of the tricuspid orifice, the cleft is more correctly described now
as communicating with that orifice rather than with the auricle.
In the left wall of the cleft immediately posterior to the junction of the
right bulbar ridge and the cushion tissue capping the interventricular septum,
a notch may be seen. This notch can be seen in seven sections (70 p.) caudal to
the interventricular foramen and is in continuity above with its lower margin.
It represents without doubt, I consider, the partial separation of the A.-V.
cushion from the cushion tissue proliferated from it and is in the same situation
as the notches referred to above in my description of the 12-5 mm. specimen.
17 mm Embryo
At this stage it is becoming less easy to distinguish the different masses of cushion tissue, the bulbar septum and the A.-V. cushions.
The interventricular foramen is just completely closed, although in one
section this is only effected by two strands of single cells, while in the section
next below this the foramen is closed on either side with its central part still
unobliterated. In both these sections the closing foramen appears as a cleft
between two masses of cushion tissue. Above this level, as in the 15-5 mm.
embryo, the right bulbar ridge and the superior tubercle of the A.-V. cushions
are completely fused. At the level of the closing foramen, While the)‘ are still
connected, they can be distinguished from each other, but in the next section
below this a deep cleft divides them, as the right bulbar ridge separates off to
enter into the formation of the right cusp of the tricuspid valve. For six sections (120 μ) caudal to the closing foramen and in continuity with it a notch
is seen on the left wall of the right ventricle, which partially separates a wedge-
shaped mass of cushion tissue capping the interventricular septum from that of
the A.-V. cushion (Pl. II, fig. 11). This is the same notch as that seen in the
preceding specimen and, like it, represents the unobliterated portion of the
groove between the A.-V. cushion and the cushion tissue which, springing from
it, has grown up round the dorsal -border of the interventricular septum. But,
while in the earlier embryo it appears as a notch in the left wall of a discrete
cavity, i.e. the caudal portion of the bulbo-auricular channel, it is now seen in
the left wall of the posterior part of the right ventricle just above the upper limit
of the tricuspid orifice. I think this difference can be explained in the following
way. Owing to the enlarging ventricular cavity the connection between the
right bulbar ridge and the right margin of the proliferation from the A.-V.
cushions has become, as noted above, so much less extensive inferiorly that
immediately below the level of the final closure of the interventricular foramen
the previously discrete channel is thrown into that of the right ventricle by the
disappearance of its anterior wall.
17.5 mm Embryo
In this embryo the development of this region of the heart is now complete. The closure of the interventricular foramen is consolidated, and all the evidence one can see now for the processes I have described is a small indentation on the left wall of the posterior part of the right ventricle (Pl. II, fig. 12) which can be traced through nine sections (90 p.) and which is undoubtedly the remains of the more distinct notch observed in the earlier specimens.
The mass of cushion tissue capping the interventricular septum appears
to be more obviously wedge—shaped than it does in the preceding photograph
(Pl. II, fig. 11). Its configuration is due, of course, to the fact, to which attention has already been drawn, that the contact which the proliferating cushion
tissue makes with the septum is asymmetrical. It joins it first to the right of
and slightly ventral to its dorsal border and spreads subsequently as far as,
but no farther than the latter point.
Discussion
Collating my own observations I would say at once that while the upper portion of the interventricular foramen as seen in my 11-2 mm. specimen remains as the aortic channel, its lower portion is closed in the following way. At first the caudal portion of the right extremity of the inferior A.-V. cushion and the interventricular septum are approximated and fuse with each other to the right of the dorsal border of the septum. In both my 11-2 and my 11-4 mm. embryos this fusion appears to be effected by active proliferation of the cushion tissue (Pl. I, figs. 1, 2). In the later stages, as shown in my 12-5, 14-5 and 15-5 mm. specimens, there is, as I interpret it, some actual upgrowth of cushion tissue over the dorsal border of the interventricular septum, so that the final closure is effected not, as in the earlier period, by apposition of the interven- tricular septum and the A. -V. cushion, but by the coalescence of the latter with proliferating cushion tissue derived from itself. Such a process as this is suggested by the microphotographs of the closing foramen (Pl. I, figs. 3-7) and also by the situation of the notch seen after the closure is complete in the 15-5, 17 and 17-5 mm. embryos (Pl. II, figs. 10-12).
I cannot say whether both the right A.-V. tubercles are involved in this
obliteration of the foramen, because after the 11-4 mm. stage it is difficult to
differentiate one from the other. The whole of the inferior tubercle certainly
takes part in it and the upper portion of the superior one certainly does not,
but one cannot be more precise than this.
With regard to the bulbar ridges, the left one is seen in the 11-4 mm.
embryo to be attached at its lower end to the right side of the interventricular
septum and thus to be demarcating the upper (dorsal) portion of the original
interventricular foramen as the aortic channel from the lower (ventral) portion
which is closed in the way I have just described. The level of the attachment
of the left bulbar ridge to the septum seems to alter little. It is obviously
difficult to obtain a fixed point in a rapidly developing organ like this, but, if
the distance from the upper border of the right superior tubercle of the A.-V.
cushions to the lower limit of the attachment of the left bulbar ridge to the
septum is measured, this is found to be 50 μ in the 11-2 mm., 70 μ in the 11-4 mm.
and 60 μ in the 14-5 mm. embryo. In the later 14-5 and 15-5 mm. specimens
the two ridges have joined to form the bulbar septum, but the smaller left
ridge can be still distinguished in the fused septum, and its inferior edge, as before, joins the interventricular septum. Here it forms the cranial boundary
of the still patent foramen.
The right bulbar ridge has been shown in the 11-4 mm. embryo to grow
across the upper portion of the right auriculoventricular orifice to reach the
right border of the lower end of the right superior tubercle and the upper
portion of the right inferior tubercle of the A.-V. cushions. In this way a
portion of the original bulbar cavity is cut off, which communicates with the
right auricle.
This bulbo-auricular channel is still patent in my 14-5 mm. embryo, but in
my 12-5 and 15-5 mm. stages the upper portion of it has been closed by the
approximation and fusion of the right bulbar ridge with the right superior
tubercle of the A.-V. cushions and has been thus incorporated in the wall of the
aortic vestibule. The lower end of the channel persists for a longer time. While
this maintains its connexion with the right auricle up to the 14-5 mm. stage,
thereafter, owing to the increasing extension cranialwards of the tricuspid
orifice and the consequent decrease in the extent of the attachment inferiorly
of the right bulbar ridge to the proliferation from the A.-V. cushions, it
appears to be thrown into the posterior portion of the right ventricle in my
17 mm. embryo.
As far as I can ascertain, the above facts were first clearly observed by Frazer (1916-17). He found that the track was closed at its aortic end in his 16 mm. specimen, while in his 18 mm. embryo no indication of its former position could be recognized with any certainty.
Do the bulbar ridges contribute anything to the closure of the interventricular foramen? If the right one is considered first, the lower portion of its
left border was seen in the earlier stages (11-4 mm.) to join the ventral right
margin of the tubercles of the A.-V. cushions. In the 12-5, 14-5, and 15-5 mm.
embryos this border is connected not with the tubercles themselves, but rather
with the cushion tissue proliferating from them to fill up the interventricular
foramen, so that the notch, which I have already described as separating the
A.-V. cushion from cushion tissue proliferating from it, is seen in the left wall
of the bulbo-auricular channel (Pl. I, fig. 5; Pl. II, fig. 10). While the right
bulbar ridge joins this proliferating cushion tissue, in no case does it appear to
contribute anything to it, and it therefore cannot be said to take any part in
closing the foramen. The result of this connexion, however, is that the anterior
cusp of the tricuspid valve, which I believe is entirely formed from the free
border of the right bulbar ridge, is continuous with the ventral end of the septal
cusp when the development of the heart is completed.
The left bulbar ridge, where it joins the interventricular septum, is forming
the cranial boundary of the interventricular foramen in my 14-5 and 15-5 mm.
embryos, and must therefore fuse, as closure is completed, with A.-V. cushion
tissue which has grown up to it along the dorsal border of the septum. Beyond
this I cannot see that it helps in closing the foramen.
If it be objected that in the later stages, such as are shown in my 15-5 mm.
embryo (Pl. II, figs. 8-10), the cushion tissue capping the interventricular
septum might as well be interpreted as being derived from the left bulbar ridge
growing downwards as from the A.-V. cushion growing upwards, I would
maintain that the clue to its origin must be sought in the earlier specimens,
where it has been shown clearly to be proliferating from the A.-V. cushions.
I have described the fate of the two bulbar ridges separately. Of course,
when the 14-5 mm. stage is reached, their adjacent margins have completely
joined with each other to form a perfect proximal bulbar septum, although the line of their coaptation can be distinguished as a seam even in the 17-5 mm.
embryo.
The lower end of this septum is thus fastened eventually anteriorly to the interventricular septum and posteriorly to the right side of the A.-V. cushions and between these two points to the cushion tissue derived from their proliferation, which is filling up the foramen and which may thus be said to complete the lower margin of the bulbar septum.
In the descriptions of the closure of the interventricular foramen and the
formation of the pars membranacea which have been published, different views
have been expressed as to the relative importance of the part played therein
by the interventricular septum, the proximal bulbar septum, and the A.-V.
cushions. Born (1889), in his classical account of the development of the rabbit’s heart, wrote that the ventricular septum spreads upwards and unites gradually with the whole right margin of the fused A.-V. cushions and lastly
with the right tubercle of the upper cushion. The lower margin of the bulbar
septum unites with the free edge of the interventricular septum so that the
remainder of the interventricular foramen is drawn into the aorta.
Mall (1912) described the interventricular foramen in his 11 mm. embryo as
being bounded “above by the union of the septum aortopulmonale and the
anterior (A.-V.) cushion, in front by the septum aortopulmonale, behind by the
extended portion of the posterior cushion and below by the muscle of the
ventricular septum”. By the union of these different masses the foramen is
closed and the membranous septum is formed.
Tandler (1912) gave no more detailed account than that of Mall. “The
interventricular foramen becomes closed in the following manner. The proximal septum bulbi grows downward and reaches the septum interventriculare.
The right end of the endocardial cushions, which have fused in the meantime,
undoubtedly participate in the fusion of the two septa, but the extent of their
participation cannot be exactly determined, since the entire circumference of
the foramen interventriculare is surrounded by endocardial growths which
pass into one another.”
Frazer (1916—17, 1931) gave a much more precise, but very different, description of these points. According to him, the pars membranacea marks the
position of the fused A.-V. cushions, but the interventricular foramen remains
as the opening into the aortic vestibule from the general cavity of the left
ventricle. By the growth of the two bulbar ridges, particularly of the left one,
which spreads along the edge of the interventricular foramen, and by their
subsequent fusion the aortic channel is roofed in and separated from the right
cavity continuous with the pulmonary orifice. To quote from his original paper
(1916): “There is no closure of the true interventricular foramen, for the
structures concerned in the separation between the two channels reach the
margin of the foramen and leave its lumen patent, affording passage to the
blood from the left ventricle to the aortic channel.”
I find it difficult to co-ordinate my description of the development of this
region with Frazer’s account of it. If Frazer’s “ true ” interventricular foramen
is the upper portion of the foramen above the level of the attachment of the
left bulbar ridge to the interventricular septum, then all other accounts would
agree that this does not close, but that it persists as the aortic vestibule. His
figures, however, suggest that more than this of the original foramen remains
patent, and not only does he not describe any filling up of the lower portion of
it by proliferating A.-V. cushion tissue, but insists (1931) in his description of
his 10 mm. embryo that the interventricular septum “ has not grown upwards
is evident from the fact that it retains its old level of attachment to the lower
A.-V. cushion: this level is original and persistent throughout”, Nor can I
confirm his description of the left bulbar ridge extending along the free margin
of the interventricular septum to reach its fellow. As I have mentioned already, I found that the lower limit of its attachment to the septum remains fairly con-
stant after the 11-4 mm. stage.
All other accounts of the development of this region regard the ventral portion of the pars membranacea as marking the site of the primitive inter- ventricular foramen, and Frazer appears to be alone in denying this. While there is general agreement that the fused A.-V. cushions must form its posterior or upper portion, i.e. the part which separates the aortic vestibule from the right atrium (Hochstetter, 1898), Frazer is singular in making them responsible for the whole membranous part of the septum. My own observations would rather confirm, with the addition of some details, the classical description given by Born, who described the development of the rabbit’s heart nearly fifty years ago.
Conclusions
The attachment of the left bulbar ridge to the interventricular septum may be said to divide the original interventricular foramen into two parts. That part of it, which is above and dorsal to this attachment, remains patent as the aortic channel; that part, which is below and ventral to it, is filled up by the active proliferation of the right tubercles of the fused auriculoventricular cushions to form the lower, ventral, half of the pars membranacea septi. The upper, dorsal, portion of the membranous septum is derived from the fused A.-V. cushions themselves. '
The proximal bulbar septum is finally attached by its inferior border
anteriorly to the interventricular septum, posteriorly to the right margin of
the A.-V. cushions and between these two points to the cushion tissue which
has proliferated from the latter to close the interventricular foramen, this
proliferating tissue connecting the two bulbar ridges and so completing the
bulbar septum inferiorly. While the bulbar septum is certainly responsible for
the separation of the aortic from the pulmonary channel, it does not contribute
anything to the occlusion of the interventricular foramen or to the formation of the pars membranacea septi.
I gratefully acknowledge my indebtedness to Prof. E. Fawcett and to
Prof. D. M. Blair for their kindness in lending me embryos from their collections to amplify our own series here. I must also thank Mr W. Chesterman of
this department for his assistance in the making of my models and for the preparation of the microphotographs.
References
Born, G. (1889). Arch. mikroekop. Anat. Bd. xxxm, S. 284.
Frazer, J. E. (1916-17), Frazer JE. The Formation of the Pars Membranacea Septi J. Anat., Lond., voL LI, p. 19.
Frazer, J. E. (1931). A Manual of Embryology, p. 316.
Hochstetter, F. (1898). Wien. klin. Wachr. Vortrag gehalten in d. Wiss. Aerztegesellschaft zu Innsbruck.
Mall FP. On the Development of the Human Heart Am J Anat. 1912;13: p240.
Tandler, J. (1912). Manual of Human Embryology, vol. II, p. 561. Keibel and Mall.
Explanation of Plates
Abbreviations
- R.B.R. right bulbar ridge.
- L.B.R. left bulbar ridge.
- R.S.T. right superior tubercle of the A.-V. cushions.
- R.I.T. right inferior tubercle of the A.-V. cushions.
- I.V.S. interventricular septum.
- A.V.C. auriculoventricular cushions.
- B.A.C. bulbo-auricular channel.
Plate I

Fig. 1. A section immediately caudal to the interventricular foramen in an 11-2 mm. embryo ( x 80) to show that a tongue-shaped proliferation from the A.-V. cushion, R.I.T., is making contact with the right side of the interventricular septum, I. V.S., ventral to its dorsal border.
Fig. 2. A section through the interventricular foramen in an 11-4 mm. embryo ( x 80). It shows the right bulbar ridge, R.B.R., in contact with the right superior tubercle of the A.—V. cushions, R.S.T., to form the bulbo-auricular channel, B.A.0. R.I.TL marks the inferior A.-V. tubercle proliferating cranialwards.
Figs. 3-5. Sections through this region of the heart in a 12-5 mm. embryo ( x 80). Fig. 3 shows the interventricular septum, I. V.S., capped by cushion tissue (X), which is in contact with the right bulbar ridge, R.B.R. In Fig. 4, which is the next section below Fig. 3, this tissue X is connected with both the right bulbar ridge, R.B.R., and with the A.-V. cushions, A.V.0. Fig. 5 is a section three below the last and shows the proliferating tissue joining the A.-V. cushions, A.V.0. The junction between the two is marked on either side by a notch. The bulbo-auricular channel, B.A.C'., appears in Fig. 3 as a. narrow cleft between the right bulbar ridge, R.B.R., and the A.-V. cushions, A. V.0.: in Fig. 4 this channel is better marked, while in Fig. 5 it is seen to be continuous with the right auricle.
Figs. 6-7. These are sections through the heart of a 14-5 mm. embryo ( x 80). In Fig. 6, which is through the centre of the interventricular foramen, the A.-V. cushion, A. V.0'., is seen to be proliferating, while similar cushion tissue, X, caps the interventricular septum, I. 17.8. In Fig. 7 the proliferating cushion tissue is making contact with that on the septum, X, while its right border is joined by the right bulbar ridge, R.B.R., as it encloses the bulbo-auricular channel, BA .0.
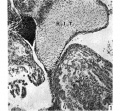
Fig 1.

Fig 2.

Fig 3.
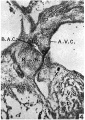
Fig 4.

Fig 5.

Fig 6.

Fig 7.







Plate II

Figs. 8-10. These are microphotographs of sections from a 155 mm. embryo (x 80). Fig. 8 is immediately cranial to the interventricular foramen and shows that in the proximal bulbar septum the right, R.B.R., and left, L.B.R., bulbar ridges can still be distinguished. The right one is fused with the right superior tubercle of the A.-V. cushions, A. 17.0., while the lefi one joins the interventricular septum, I . V.S. Fig. 9 is a section through the centre of the foramen and shows the septum, I. 17.8., capped with cushion tissue, X. The latter, as is seen in Fig. 10, which is the fourth section caudal to the level of the foraminal closure, joins the A.-V. cushion. A. V.C’., on its left side but is still separated from it on the right by a notch, which appears as a diverticulum from the anterior end of the inferior portion of the bulbo-auricular channel, B.A.C'.
Fig. 11. This is a section through the heart of a 17 mm. embryo ( x 80) five sections below the closing foramen. It shows the A.-V. cushion, A.V.0., separated, as in the last figure, by a notch on the right side from the cushion tissue, X, which has proliferated from it to join the interventricular septum, I. 17.18’.
Fig. 12. This is a section from a 17-5 mm. embryo ( x 80) and shows a notch in the same situation as that seen above, but now it is much less well marked.
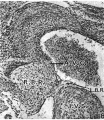
Fig 8.
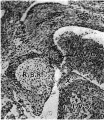
Fig 9.
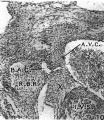
Fig 10.

Fig 11.

Fig 12.





Cite this page: Hill, M.A. (2024, April 24) Embryology Paper - The Development of the Pars Membranacea Septi in the Human Heart. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_The_Development_of_the_Pars_Membranacea_Septi_in_the_Human_Heart
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G