Paper - Origin of the sex cells in man
| Embryology - 20 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Simkins CS. Origin of the sex cells in man. (1928) Amer. J Anat. 41: 248-272.
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
Origin of the Sex Cells in Man
Cleveland Sylvester Simkins
Division of Anatomy, University of Tennessee Medical School, Memphis, Tennessee
Ten Plates (Forty-Six Figures) 1928
Introduction
Investigations dealing solely with the origin of the sex cells in man appear very infrequently, due, no doubt, to the scarcity of suitable material. The few contributions that have been oifered toward the solution of this problem seem to the author to be quite limited in scope.
Fuss (’11), upon the basis of Rubaschkin’s (’10) proposed method of differential coloration of the germ cells, located what he identified as ‘Extraregionare Geschlechtzellen’ in the intestinal epithelium of a four~weeks human embryo. The supposed sex cells were disposed in such a way as to suggest an active migration from the entoderm of the gut into the visceral peritoneum. He extended his investigations into several genera of mammals including man; these were published in 1913. His later conclusions confurmed, but added little to, his previous observations.
Felix (’12), influenced probably by the work of Lams et Doorme (’07), Rubaschkin (’10), et al., distinguished two categories of cells in the course of gametogenesis, which he designated as primary and secondary genital cells. The primary genital cells, he contended, had a special origin from the segmentation cells, and hence were extraregional in origin; i.e., they arose directly from the blastomeres and remained distinctly reproductive cells destined to migrate into the gonads where they were found to degenerate and to contribute nothing to the definitive procreation elements. The secondary genital cells he found to be derived from the epithelium of the gonads, and these were metamorphosed directly into the ova or sperms; the two categories, then, were found to have no physiological or genetic connection.
Kohno (’25), working upon the ‘Keimbahn’ of man, recognized what he considered to be germ cells in the lateral plates of mesoderm of a 2.3—mm. embryo, and as these plates were folded under the gut in 2.8-mm. embryos, the germ cells moved up into the epithelium of the gut and mesentery, from which place they migrated farther into the developing gonads of 5- and 7-mm. embryos. The four embryos, 2.3, 2.8, 5, and 7 mm., made up the entire series upon which he based his conclusions. He did not mention when the migration ceased. There is the possibility that his ‘Urgenitalzellen’ are not germ cells at all, but somatic cells in different phases of metabolic activity——a point that I shall discuss more in detail.
The problem at present is in rather a confused state. Two prevailing and opposed opinions may be gathered, viz., 1) that the eggs and sperms have extraregional forebears derived from the early segmentation cells and these may or may not degenerate, and 2) that the definitive reproductive cells are products of the germinative epithelium.
Materials and Methods
Only human embryos have been used in this study and, with certain breaks, they constitute a series from embryos of 2.6 mm. to infants six months postpartum. The material was fixed in 10 per cent formalin, sectioned from 7 to 10 p by the paraffin method and stained in haematoxylin and eosin. A few were stained in carmine followed by azure II. Embryos up to 20 mm. C.R. length were sectioned in their entirety, but in the more advanced ones the gonads were dissected out and sectioned alone. The gonads of stillborn and premature infants were removed at autopsy and fixed at once in formalin. No measurements were made upon these infants. They were entered in the series as so many months postpartum or as full term.
Inasmuch as my primary object was to secure a series in an ascending order, one method of measurement was chosen and adhered to at all times ; this was the crown-rump length, hereinafter referred to as C.R.; all figures in millimeters, then, represent the C.R. height. Merely for the convenience of treatment, the embryos were arranged into three groups, shown in table 1. The first group includes thelembryos whose gonads, if present at all, are in the indifferent stage, of which the table shows ten; the second group includes the females, and the third, the males.
Table 1
| Table 1 - Tabular view of the embryos used in the present study. All measurements are in millimeters and represent the crown-rump length | ||
|---|---|---|
| Indifferent | Female | Male |
| 2.6 mm. | 20.0 mm. | 25.0 mm. |
| 3.2 mm. | 25.0 mm. | 43.0 mm. |
| 5.0 mm. | 30.0 mm. | 54.0 mm. |
| 7.4 mm. | 38.0 mm. | 70.0 mm. |
| 7.6 mm. | 50.0 mm. | 154.0 mm. |
| 10.1 mm. | 90.0 mm. | 190.0 mm. |
| 15.0 mm. | 144.0 mm. | 316.0 mm. |
| 18.0 mm. | 154.0 mm. | 350.0 mm. |
| 20.0 mm. ? | 167.0 mm. | 3 at full term. |
| 195.0 mm. | 1 at 6 months postpartum | |
| 345.0 mm. | ||
| 25 at full term. | ||
| 1 at 5 months postpartum | ||
Observations
In embryos of the first month (2.6 and 3.2 mm.), the peritoneal epithelium is composed of a single layer of cells whose nuclei are spherical and present such a slight variation in size that none of them stand out conspicuously. No large cells of a startling appearance are to be found in the body, neither in the gut epithelium nor in the mesoderm of the somites, and, furthermore, none of them react in any specific manner to the various stains employed. Now and then, as one searches through the sections of these early embryos, certain clear circular areas are met with whose center has a pycnosed knot of chromatin suggesting a nucleus, and it bears a very close resemblance to the ‘Urgenitalzellen’ figured by Kohno in his excellent microphotographs; a ‘diagnosis’ with which I cannot agree, since these so—called cells are not true cells at all, but liquefied areas surrounding a mitotic or degenerating nucleus.
In embryos of 4.2 and 5.0 mm. the mesonephros begins to form and causes a slight bulge into the dorsal compartment of the body cavity (fig. 44, c0r.ms’nph.). The epithelium along the ventromedian border of the mesonephros is slightly thickened, but shows no cells that differ from the somatic cells in other epithelial layers of the body. Some of the cells are a little larger than others, but to no startling degree. I have been unable to discover any cells of peculiar size or staining reaction that bear an identical or precise appearance to the unquestionable germ cells of the early ovary anywhere in the body of embryos as young as the first month. Even in 7-mm. embryos, after the mesonephros has caused an extensive bulge into the peritoneum (fig. 45), no cells of startling size could be located either in the mesentery or in the germinal epithelium which is beginning to thicken on the ventromedian surface of the mesonephric body. There are no anatomical landmarks yet formed to demarcate the extent of the gonadogenic area of the mesonephros, hence the epithelium covering it is often incorrectly called the germinal epithelium, since it is yet but a peritoneal epithelium. Such a condition does not lon.g obtain, for there soon takes place a thickening of the epithelium 011 the ventromedian surface of the mesonephros, extending from the anterior end of that body rearward to its posterior end. (At first the epithelium does not undergo extensive proliferation, the thickening results from the cells’ assuming a cylindrical form, and later dividing and moving inward. The cells of the epithelium vary only slightly insize (fig. 45), none of them having acquired as yet the darkly staining cytoplasm so characteristic and constant of the primordial oocytes that develop later in the indifferent gonads.
Immediately following the 7 mm stages, the gonads enter upon a period of growth which restricts the extent of the sexual anlage and at the same time thickens the gonogenic mass by a rapid proliferation of cells from the peritoneal epithelium. By the time the embryo has attained a length of 10 mm., the genital anlage is confined to the ventromedian surface of the mesonephros and is marked off from the non-sexual epithelium by slight furrows fore and aft. The wolflian body constitutes an extensive mass that bulges into the caudal partof the visceral cavity and extends from the second or third thoracic segment cranially to the urogenital sinus caudally. The proliferation of cells from the coelomic epithelium which marks the beginning of the formation of the sexual gland does not proceed at the same rate over the entire median surface of the mesonephric body, but diminishes in activity rearward beyond the posterior one-fourth of the Wolflian body, hence the primordium covers the anterior three-fourths of the ventromedian surface of the mesonephros. In this circumscribed area the cells from the epithelium only slightly invade the stroma of the wollfian body, and none of them are large enough to attract more than passing attention; in actual measurement the diameters of the nuclei vary from 7 to 9 u.
The 10 to 11 mm stages are critical ones in the development of the human reproductive gland, for during that time the activity of the cells of peritoneal epithelium initiate those changes that eventually terminate in the formation of the gonads by transforming somatic cells into the primary propagative elements. The actual change is soon seen in the tendency of certain cells derived from the epithelium (now a true germinative epithelium) to arrange themselves into short cord—like processes extending from the superficial epithelium down into the stroma. These are the early sex cords. The relative size of mesonephros becomes less, due to the accentuation of the‘ gonads by the deeper penetration of the cord—like processes into the region of the future sexual gland and the actual increase in size of gonad, rendering much more pronounced the furrows that bound the anlage. The sex cords of the cranial part of the sexual anlage penetrate deeper than anywhere else, reaching, in that part of the gland, almost to the capsules of the mesonephric glomeruli. In both the stroma and epithelium of the in-difierent gonad of tlie 11—mm. stage there are to be found large and small nuclei; difference in size is all that can be attributed to them, for they have not yet acquired a different reaction to the stain, both types of which must be looked upon as derivatives of somatic cells with peculiar propagative possibilities.
In embryos of 15 mm the most cranial of the mesonephric tubules show retrogressive changes, which retrogression extends to the anterior end of the gonad so that the cranial ends of both gonads and the mesonephric bodies lie at the same level. Anteriorly, the sex gland is distinctly raised and separated from the wolffian body by deep sulci; these play out caudally, so that the gonads are only slightly elevated from the caudal surface of the mesonephros. Differentiation of the genital gland proceeds from before backward, so that the anterior end of the anlage is slightly in advance of the caudal, hence the sulci demarcating the lateral limits of the genital ridge are pronounced cranially, but shallow caudally. The cells of the sex gland which showed an incipient cord-like arrangement in the 10-mm. stage are slightly more pronounced and extend into the wolffian body, sometimes branched, sometimes straight, until they reach the region of the mesonephric glomeruli. The cells composing the sex cords are larger than the cells lying between them (fig. 4). The larger cells of the sex cords vary only slightly in size from each other (fig. 34) and are large and spherical, lying in an intercellular substance that stains darkly. Between the sex cords there lie smaller connective-tissue cells and indifferent cells, whose nuclei stain darkly and the cytoplasm lightly. The contrast between the staining reaction of the sex cords and the cells between them is most striking and remains constant throughout all the stages of the indifferent gonad. A distinction should be made here between the large cells of the sex cords and the genitaloid cells, because of the conspicuousness and the late diflerentiation of the latter. The genitaloid cells may either lie Within or without the cords. They, like the large cells of the sex cords, have large spherical nuclei that stain lightly and a definite mantle of cytoplasm about each of them that stains darkly; differing from the large cells of the cords only in that the cytoplasm is limited and bounded instead of lying in a syncytial mass. The number of genitaloid cells is small compared to the number of large cells in the sex cords~—a condition that can be explained by assuming that the genitaloid cells are specially differentiated recruits from the germinative epithelium and the cords and at such an early time (15-mm. stage) very few have formed. The cords (fig. 4) are the most characteristic structures of the 15-min. embryo and extend throughout the entire gonad from the rete cords inwardly (re., fig. 4) to the germinative epithelium outwardly. ‘
The germinative epithelium is thick (e’th.g., fig. 4) and composed of more than one layer of cells; from its inner surface hillocks of cells project toward the stroma, but in no place connect morphologically with the peripheral ends of the sex cords. Some of the cells of the epithelium are large, but none so large as the genitaloid cells or the cells of the cords. The larger ones lie on the inner surface of the investing layer where they seem to be detaching themselves from it and invading the spaces between the cords, probably to form new ones and eventually to anastomose with those already formed; some of them, too, may contribute to the ranks of the genitaloid cells. i
The mesonephros degenerates from the cranial toward the caudal end, and by the 18—mm. stage this degeneration has attacked the tubules as far back as the anterior end of the sex gland. The sex gland itself (fig. 6), is composed chiefly of undifferentiated cells, the genitaloid cells being met with about as often as in the 15—mm. stage. The sex cords have more deeply invaded the stroma until they seem to unite with the rete cords which are differentiated from the inner ends of the sex cords and serve as connections between the tubuli recti of the seminiferous tubules and the genital tubules of the mesonephros in the males, but the rete cords disappear in the females. The sex cords are more complex in their branching~—a condition brought about by the formation of additional cords and the additional anastomoses.
The large cells of the sex cords, the gcnitaloid cells, and the small fusiform cells are the most important cells of the gonad, and for the most part show no great advance in specialization over the same types in the 15 mm embryos.
The differentiation of the indifferent sexual gland has to do with the formation of an ovary and a testis in which the genital cells lead on to the formation of spermatogonia and oogonia; but in order for these cells to arise the gland must take on the structure of an ovary or a testis. The most characteristic structures of the male gland are the testis cords and the broad tunica albuginea, while the abortive or incomplete formation of these two is characteristic of the ovary. The diverging lines of development followed by the gonads after the 18—mm. stage lead to a definite differentiation of the sexes. The rapid differentiation of the sexual apparatus is concerned mostly with the sex cords, and this process will be dealt with in the following pages.
The embryonic gonad differentiates along the lines of its predetermined sex after the 18-mm. stage (Felix (’11) places the stage at 13 mm.), and by the time the embryo has increased in length to 20 mm. the sex of the individual can be accurately determined. In 20-mm. embryos the mesonephros has degenerated considerably and the vestigial remains of that organ hang suspended from the dorsal body wall by a narrow mesentery (fig. 9). Traces of the glomeruli are still present and the mesonephric duct is still patent. The sex gland swings from the degenerated mesonephros by a mesentery in which the well-defined rete are seen. The sex cords show a distinctive tendency to differentiate along diverging lines, one of which leads to the characteristic structure of a testis and the other to the ovary. In the ovary (fig. 9) the cords begin to break up into masses separated from each other by connective-tissue cells. In these masses there are large cells (fig. 43) which are probably the forerunners of primordial ova. These are the genitaloid cells which are more numerous in females than in males.
In the 25 mm embryo the genitaloid cells (fig. 16) lie thickly scattered in the ovary, but in the male are found within and without the cords, separated from each other by strands of densely staining small spherical cells. The sex cords are bounded by discontinuous fusiform nuclei which probably are the embryonic connective-tissue walls. The sex cords began to assume a definite arrangement during the 20—mm. stage, the walls of small fusiform cells surrounding the cords and containing the larger cells, which are never so large as the genitaloid cells of the ovary. The formation of incipient connective-tissue walls about the sex cords tends to confine them in connective-tissue tubes, which is a definite step toward the formation of the male gland. At the same time the differentiation in the opposite sex tends to break up the sex cords into irregular masses and indefinite cords without the regular connective~tissue walls to surround them, hence the early ovary from embryos of 20 mm. difiers from the male in the looseness of the cords and the irregular manner in which the connective tissue ramifies through the gland. Sexual diiferentiation begins in the early stages, but it is not until the 18- to 20—mm. stage that the differences between the two sexes is great enough to enable a definite and certain determination of the sex; nevertheless, the tentative criterion is reliable that the sex cords and rete cords, forerunners of the seminiferous and rete tubules, respectively, progressively differentiate in the male, but cease to do so in the female. From this time on the history of the early germ cells follows two diverging lines: one leads to the forma.tion of the oocytes and the other to the spermatocytes.
History of the Male Sex Cells
The complete differentiation of the testes concerns chiefly the union of the sex cords to the rete cords and the formation of connective—tissue sheaths around this system of primordial tubules. Very early (15 mm.), the cells of the gonad destined to build the connective tissues of the gland elongate, assume an elipsoid outline, and finally transform into long slender fusiform cells, with conspicuous and densely staining nuclei. Blood cells present in the gonad are large and spherical and may easily be mistaken for genitaloid cells. The difference is that the true genitaloid cells have nuclei that stain lightly and cytoplasm that stains darkly, while the blood cells have nuclei and cytoplasm both staining lightly. There are large and small cells present, which belong neither to the category of connective-tissue cells nor to the blood cells; these shall command immediate attention, because their relation to the developing sex cords and to the spermatogonia has a peculiar significance.
The large cells, even in the indifferent gonad (figs. 23, 24), have large spherical nuclei and are surrounded by a cytoplasmic mantle that takes the stain intensely (fig. 40). The diameter of the large cells is often twice that of the smaller cells. The smaller nuclei stain intensely, the chromatin being compact and granular, the cytoplasm differing in its reaction to the stain in different places, sometimes staining faintly and at others densely (compare figs. 34 and 35). The large cells, however, are characteristic of the sex cords in embryos of 20 mm., only an occasional genitaloid cell being observed in the spaces between the early tubules. The genitaloid cells stain as they did in the indifferent gonad, i.e., the nucleus stains lightly and the cytoplasm takes the stain very darkly. When the genitaloid cells lie within the confines of the tubules, their conspicuousness is lost by lack of contrast, for the large cells of the tubules almost equal them insize. The constant difference between the large cells and the genitaloid cells of the sex cords lies in the appearance of the cytoplasm, most important being that the cytoplasm of the large cells is not definitely arrayed around a single nucleus, while in the genitaloid cells it is; large cells lie in a syncytial cytoplasm, but the genitaloid cells lie in circumscribed masses. The distinction here is finely drawn in anticipation of the fate these cells are to suffer.
In male embryos of 25 mm. the sex cords anastomose peripherally in loops and join in twos and threes centrally before uniting with the rete. The sex cords consist for the most part of indifferent epithelial cells, some of which are large, with an occasional genitaloid cell interposed. The testis as a whole is larger than the wolfi‘ian body and has assumed an ovoid outline with its long axis directed somewhat transversely.
So far the differentiation of the testis has been concerned with the cords rather than with the cells that compose them. Their inner ends reach and unite with the rete testes, which in turn show a close relation to the genital tubules of the mesonephros, the relationship indicating what will eventually take place, namely, the union of rete and genital tubules. The peripheral ends of the cords are club-shaped, beyond which there intervenes a layer of connectiVe—tissue cells, out of which the tunica albuginea develops. Grenitaloid cells are to be found lying between the sex cords as far inward as the rete; in each case they are large, spherical, and have the characteristic light nuclei and dark cytoplasm.
The testis of a 54—mm. embryo is made up of a series of darkly staining cords (fig. 21) each one of which is surrounded by a delicate connectiVe—tissue she-ath; outside of this sheath lie the large genitaloid cells, some of them, however, are entrapped in the sex cord by the developing sheaths, where they sutier a change from light staining nuclei with dark cytoplasms to dark nuclei and pale cytoplasms, while their fellows not so entrapped (fig. 33, cl..90.) retain their characteristic reaction, viz., dense cytoplasm and light homogeneous nucleus. These cells are distinctive and persist for some time in the sex cords; eventually, however, they undergo dissolution, while those lying in the interstitial spaces develop and specialize along certain well-defined lines and are the forerunners of the cells of Leydig (fig. 41).
The cells making up the sex cords undergo a series of changes which forecast the formation of the seminiferous tubules (fig. 27). The change is most noticeable in the arrangement of the cells into concentric rows around the periphery of the tubules, the small cells taking their positions nearest the incipient basement membrane and the large ones lying nearer the future lumen where degenerative changes begin. The extratubular genitaloid cells (figs. 23 and 24) are so similar in shape, in size, and in staining reaction to the genitaloid cells of the ovary that one must consider them identical and to have a common origin, yet their fate is by no means similar. A
The differentiation of the testis shows its continuous advancement in the distinct delimitation of the seminiferous tubules, the extensive Vascularization of the organ, the formation of the testicular septa, and the thickening of the tunica albuginea. The large genitaloid cells unincorporated into the cords or tubules lie in groups or singly in the interstitial spaces of gonads of embryos ‘of 154 mm. (fig. 36). Within the confines of the tubules the small cells lie, more or less arranged in concentric rows, most of them being of spherical form, with an occasional fusiform nucleus interspersed.
The testes increase greatly in size as a result of the growth and the peripheral anastomoses of the primordial tubules, some of which, by the time the embryo has attained a length of 190 mm., begin to acquire lumens. Testicular septa (fig. 14) enclose bundleslof tubules centrally and extend toward the periphery of the gonad, where they unite with the tunica albuginea. The peripheral ends of the tubules are clubshaped and somewhat contorted, the central parts are straight and join the rete tubules. The contorted parts of the tubules (fig. 5) still contain some large cells, while in the kinks and interstitial spaces one encounters numerous genitaloid cells. Many of the large, genitaloid cells that were entrapped i.n the tubules show signs of degeneration, but those in the interstitial spaces retain their characteristic appearance (fig. 15).
The tubules of the testes at three months after birth (fig. 12) are definitely formed, most of them having well—defined lumens. The large cells have about all disappeared from them, but are still numerous in the interstitial spaces. The walls of the tubules are thick and the smaller cells lying close to them are arranged in a regular order; an occasional mitotic figure is encountered indicating that the testis, now within the scrotum, is advancing toward sexual maturity. The disappearance of the large genitaloid cells from the tubules is of great importance, for it indicates that the definitive spermatogonia arise from the smaller cells, while outside the tubules they contribute to the formation of the interstitial cells. In the ovary these large genitaloid cells have a different fate, which will be considered in the next section.
The comportment of the cells in the developing testes has to do with the large and small units. From the time that a testis can be distinguished from an ovary (about the 20-mm. stage) these two types pursue a diverging course. Both types arose from the germinal epithelium (converted ooelomic epithelium) and progressively differentiated into genitaloid cells and genital cells. Not only have the forerunners of the sexual elements come from the epithelium, but the rete cords (Wilson, ’26) and connective-tissue cells (some of the connective-tissue cells migrate inward from the hilus, according to Felix, ’12), and nurse cells as well. As soon as the genital swelling has attained a sufiicient size, the sex cords develop quite suddenly within it; in the female these cords undergo fragmentation, but in the male they give rise to the seminiferous tubules. They stain densely and for the most part are composed of large nuclei, which stain feebly, and a cytoplasmic matrix that stains darkly. Connective-tissue sheaths form around the cords entrapping many of the large genitaloid cells. Those left on the outside persist and become the interstitial cells, while those confined within the tubules perish; they do not disappear all at once, some of them are present as late as six months postpartum. The smaller cells arrange themselves around the inner wall of the tubules and enter upon those transformations preparatory to the production of the male propagation cell. With the six—month postpartum infant my study ends, hence I am unable to say at what time the last genitaloid cell disappears from the tubules.
History of the Female Sex Cells
Felix (’12) states that it is possible to identify an ovary or a testis by excluding certain characters, such as branched, anastomosing sex cords and a broad tunica albuginea as early as the 13—mm. stage; but he hastens to add that the retarded diflerentiation of a. male renders the identification of a female par exclusionem quite unreliable until the embryo has attained a length (OR) of at least 20 mm.
As has been mentioned above in the present study, I have found that the sex cords develop equally well in the indifferent gonad up to the 18-mm. stage, but at about the stage of 20 mm. the cords begin to fragment or break up into elongated and irregular masses, which means that the progressive difierentiation of the sex cords ceases in one sex, which is the ovary, hence they remain in the simpler or unspecialized condition for a longer time than in the testes (fig. 9). The ovaries are traversed by the indefinitely outlined cords that extend in a general way from the periphery to the hilus, where they end in close proximity to the rete, with which, however, they never unite (fig. 9, re.). In the 20-mm. ovary the large and small cells are present, although the contrast between them is not striking (fig. 43).
In the 25-mm. embryos the ovary (fig. 16) hangs suspended by the mesovarium and consists of an enlarged medulla of epithelial origin, sometimes called the epithelial core or nucleus, and a superficial epithelium, both well demarcated from one another. In the medullary portion there are cords of large and small cells having a tendency to alternate with each other and radiate outward toward the periphery. The large cells are loosely distributed, each has a spherical nucleus that stains lightly, and each one is surrounded by a sharply outlined body of cytoplasm that stains darkly; these are the sex genitaloid cells and are identical with the genitaloid cells described as occurring in the early testis. The superficial epithelium is made up of small nuclei that stain intensely and are held in a syncytium of intranuclear cytoplasm. It is of great significance that no large genitaloid cells are to be found in the superficial epithelium, because it indicates, I believe, that the delivery of cells to the core has about ceased, all the forerunners of the germ cells having previously moved into the epithelial nucleus or core. The distinction between the large and small cells is clear-cut, which enables one to trace their fate with accuracy.
The superficial epithelium is the source of all cells of the incipient ovary, and at the 25-mm. stage begins to take on the character of an insignificant investing layer more or less completely separated from the core of the gland. Such a complete separation and cessation of cellular proliferation is not attained suddenly, for in embryos of 30 and 38 mm. there are still signs of an actual delivery of cells from the investing layer into the stroma. In an embryo of 30 mm. (fig. 11), the superficial epithelium is a true germinative epithelium, for cells can be seen moving from its inner border into the stroma, some of which have all the characters of genitaloid cells. It is possible that the delivery of cells into the epithelial core ceases only when the tunica albuginea forms and prevents further ingress. This View gains weight from the fact that in all the early stages of the formation of the ovary, prior to the completion of the tunica albuginea, there are areas observed where no separation of the outer and inner parts exists and of course the germinative epithelium and epithelial core are in morphological contact (fig. 20), indicating that the disjunction is not everywhere completed at the same time and entailing no abrupt cessation to the movements of cells from the superficial layer into the deeper layer, and rendering thereby the likelihood of a second proliferation and the formation of a second epithelial nucleus in addition to the first quite uncertain and unnecessary.
The outer or investing‘ epithelium of the gonad of a 30-mm. embryo (e’th..g., fig. 20) is compo-sed of a single layer of cells, the nuclei of which are small and stain homogeneously dark. In the epithelial core are to be found the large genitaloid cells (cl._q., fig. 20) and the smaller cells, too. Out of the general assemblage of cells that make up the core of the gonad, three lines of specialization lead: one line tends to surround the large genitaloid cells, another fills in the spaces between them, and a third type includes the genitaloid cells themselves. The sex cords are recognized with difficulty, their remains persist for a longer time near the hilus, at which place the blood vessels begin to penetrate the gland and progress toward the periphery. The sex cords become more and more indistinct as time passes, fewer of their remains being seen in the'38-mm. stage, fewer still in the 43-min. stage; very little change other than the disruption of the cords takes place until the embryo has reached a length of 51 mm.
The outstanding features of the 51-mm. ovary (fig. 34) are the sex cords and the rete cords. The sexcords are few in number, the rete are but indefinite rows of small cells (re., fig. 34). The superficial epitheliumis very thin and the tunica albuginea is thick. In ovaries of the same age (not figured) one is able to recognize a cortical zone and a looser, pale-staining medullary zone; the differentiation of these two zones finally leads to the complete obliteration of the sex cords in the ovary. In embryos of 90 mm. (fig. 25) the cortical zone is sharply, though irregularly delimited from .the medullary zone, very few large cells remaining in the cortical zone. The medulla contains indefinite rows of small pycnotic nuclei-—-the remains of the rete cords; the large cells lying around the peripheral border of the medulla take unto themselves a concentric layer of smaller nuclei (figs. 29, 30)-—the incipient follicular cells. Divisions of connective tissue grow from the hilus toward the periphery of the gonad invading the cortical zone and spreading out just under the investing epithelium augmenting the thickness of the tunica albuginea, which is completed about the fifth month of gestation, after which time no more germ cells or any cellular recruits from the germinative epithelium reach the epithelial nucleus. Hence, from the 90-mm. stage onward the histogenesis of the ovary is concerned only with the cells already included within the core, By the fifth month the cells of the core have differentiated into the large spherical germ cells surrounded by the incipient follicular cells and the fusiform cells of the tunica albuginea and septa ovarii.
The large cells may now be safely called the genital cells; their nuclei are invariably spherical and, because of the attenuated condition of the chromatin, are quite conspicuous by their lightness, which is accentuated by the mantle of cytoplasm around them which stains homogeneously dark. The small cells destined to become the follicular cells begin to surround the genitaloid cells in stages of 80 mm.; at such a time most of them (genitaloid cells) lie freely scattered in the epithelial nucleus; but with the formation of the septa ovarii they move toward the peripheral margin of the medullary zone, invading the cortical zone and there enter upon a period of growth, first characterized by the acquisition of follicular cells around the peripheral margin of the darkly staining cytoplasm. By the time the embryo has attained an age of five months, the genital cells have all been surrounded by the smaller follicle cells and appear as incipient follicles (fig. 38). The rapid enlargement of the follicles probably exerts a peripheral pressure that tends to flatten the connective—tissue cells surrounding them (fig. 39); portions of the septa ovarii containing connective—tissue cells ramify among the follicles contributing their quota to the formation of the theca externa and interna. Some follicles begin to form in advance of others, the greater number being well advanced at the fifth month, but some start later, so that at birth some follicles are mature and others yet in the primary stage (compare figs. 17, 18, 19, 31, 28, 42, and 32; different stages in the formation of the follicles all taken from a full-term ovary).
The smaller cells which are entrapped in the thecae about the developing ova proliferate apace, augmenting their numbers as the size of the egg increases. The follicles begin to form in the medulla in the region of the rete blastema and move gradually into the cortex, Where continued enlargement brings them to the surface of the ovary. The movement of indifferent cells takes place in the same manner, fromthe medulla to the cortex; during this migration the somatic cells undergo transformations into the genitaloid cells, acquire the follicular arrangement, and begin to increase in size. Hence in the cortical zone are to be found follicles in all stages of development ranging from the immediately transformed genitaloid cells (figs. 30, 40, and 42) to mature follicles. The transformation seems to take place in a continuous and progressive manner from the cells of the epithelial nucleus, there being no indication that a new or neogenic zone develops over the old epithelial nucleus by a second delivery of cells from the germinative epithelium.
There is a progressive increase in size of certain cells in the developing ovary from the 20-mm. stage onward, which attain conspicuous and characteristic features in the latter part of the fourth month (fig. 30), and these are to be regarded as the true forerunners of the propagation cells. At the same time that these primordial oocytes that have come from the epithelial nucleus begin to acquire the characteristic structure of incipient follicles, other indifferent cells of the late medullary zone are transforming into oocytes; thus the process of transformation becomes continuous, but the recruits come from two different sources: the earliest oocytes coming from the nucleus of the gonad and the later ones from the medullary zone; both categories, however, strive toward the same end, viz., the production of definitive ova, which end the primary ones never reach.
In order to determine the fate of the large primordial genital cells and the later ones, the gross changes of the developing ovary must be clearly conceived and kept so in mind, because the changing relationships of the different sex zones constantly alter the conditions from one stage to the next. Shortly after the formation of the epithelial nucleus, which I believe to be the result of a continuous delivery of cells from the germinative epithelium, and not from an early and late delivery, two well-defined parts can be recognized in the gonad, the nucleus itself and the investing or germinative epithelium; the latter continues to deliver cells into the stroma until the tunica albuginea forms and prevents their ingress; this continues as late as the fifth or sixth month. The primordial germ cells, those delivered from the germinative epithelium during the formation of the indifierent gonad, lie deep within the stroma, almost" to the hilus, where they are found in greater abundance near the peripheral ends of the rete ovarii. Here they remairruntil the tunica albuginea is formed and the stroma takes 0117 the character of the cortical and medullary zones. The large cells lie yet within the medullary zone, where, because of their large size, thin chromatin, and disunion, they give to that zone its characteristic pale and loose structure. In the medullary zone some of them take on the appearance of follicles, lie loosely associated in nests, and disposed toward the cortex, into which zone they eventually move-along with the later transformed indifferent cells, Where with the second type of genital cells they enlarge apace, some of them degenerating, others attaining the status of mature follicles at birth. The growth of the primary genital cells into follicles is similar to the secondary genital cells, which follows this course; the large cell becomes surrounded by a few small spherical cells, outside of which there are a few small fusiform cells (fig. 17). The small spherical cells arrange themselves in an orderly manner around the peripheral margin of the cytoplasm of the oocyte (fig. 18), and proliferate to form an irregular double layer (fig. 19). As yet there is no definite cytoplasmic membrane formed around the gonocytes, but its margin can be easily determined by the intense staining reaction of the cytoplasm itself. The irregular disposal of the follicle cells soon passes into an orderly one (fig. 31), and soon thereafter the vitelline membrane can be distinguished. The follicles increase in size, so that at birth certain ones (fig. 42) have acquired the parts of a mature follicle. Many of these follicles have attained a diameter at birth of 2 mm. containing eggs that show no signs of degeneration (fig. 32), perfect in outline, with a thick zona pellucida, vitelline membrane (m.vt., fig. 32), and corona radiata. I have found such follicles in the ovaries of 60 per cent of all full-term infants examined, the eggs seem to be normal in every histological detail and in follicles that apparently were about ready to rupture. Never are the secondary genital cells so far advanced at birth.
Whether or not these follicles from the primary genital cells ever rupture is a matter of doubt. I have found no corpora lutea in the ovaries of full—term infants. Runge ( ’06) reports but one true corpus luteum in ovary of ‘a newborn child. Felix (’11) states that the usual fate of these early graafian follicles is atresia and cystic degeneration.
Pycnotic, fragmented, and crenated nuclei are to be found in" all the later stages of development of the ovary; too, excessively swollen nuclei with little "or no chromatin are met with just as often, all being indications of cellular degeneration. The frequency of the occurrence of these signs increases from the fifth to the seventh month, especially in the medullary zone, so that at birth none of the large cells remain in that zone, all of -them that have not degenerated or grown into large follicles are to be found in the cortex. Because of the increasing dissolution of the large primordial sex cells of the ‘ovary after birth, it is doubtful whether any of the large follicles ever rupture at, or shortly after the.time _of, parturition. The possibility must be recognized, however, that some of them may rupture and initiate the formation of the first corpus luteum. It may very well be that the primary sex cells give rise to follicles that rupturein precocious ovulation and pregnancy. The occurrence is so rare and this fact, coupled with the report of Runge (’O6) that he has found but one corpora lutea in full-term ovaries, is strong evidence to support the contention that the precocious follicles snx CELLS IN MAN 269
are abortive attempts to form eggs, and that the definitive or fertile eggs arise from the smaller cells, which are transformed indifferent cells, that were delivered to the epithelial nucleus from the germinative epithelium. Contributions from the germinative epithelium make up the entire gonad, certain cells of which later pass from the medullary to the cortical zone and differentiate into follicles, precisely like, but at a later date than the follicles initiated by the primordial genital cells. The primary genital cells that come early from the germinative epithelium give rise to follicles that rarely if ever rupture, although those follicles almost reach maturity in the ovaries of full-term infants, only to suffer the fate of dissolution; a second draft of cells is called for from the medullary zone; from them the fertile follicles develop.
Discussion
The most recent contribution to the history of the sexual elements in man is a paper by Kohno (’25), in which the photomicrographs are so excellent that a detailed study of the section from which they were taken can be made. In the figures published there are to be seen large structures, which Kohno has called primordial genital cells (Urgenitalzellen), scattered in various regions of the embryo; some of them are found in the Wall of the gut, in the neural tube, mesentery, and in the mesonephric body. The pictures show a uniform structure having a dark pinched-up nucleus surrounded by a circular space entirely free from cytoplasm or filled with cytoplasm that stained too lightly to be photographed. Such units cannot be primordial germ cells, destined to contribute anything to the formation of the gonads, they a.re found so often in the wall of the neural tube that they have been called the germinal cells of the spinal cord, without reference What~ ever to the reproductive gland. The same class of units was recognized in the early embryos of mice by Simkins (’23_), who concluded that they were not primitive sexual units at all, but somatic cells in mitosis or some other phase of metabolic activity. Another reason Why these cells cannot be forerunners of the definitive sexual element is their reaction to the stain; in the present study, in the gonocytes of whose identity there can be no doubt, the nucleus is large and vesicular, not pinched and pycnotic; the cytoplasm is thick and stains intensely, not thin and pale: compare figures 38, 39, and 40 and then 17, 18, and 19, in the most essential features the ‘Urgenitalzellen’ of Kohno are the exact reverse of the true gonocytes.
Proponents of the view that the human sex cells have an origin remote from, and antedating the establishment of, the gonad offer as evidence to support their contention the results of early castrations. It l1as been thought that the removal of the primordial germ cell from the early‘ embryo would render the adult completely and permanently sexless: the results have not been convincing. The germ cells have been removed along with the entire gonad and its most important part, the germinative epithelium, and after the lapse of a certain amount of time, in which some opportunity was given for the regeneration of at new gonad, no such result occurred. In View of certain experiments to be cited below, one would rather expect to find some regeneration of the gonads in the embryos of amphibians, but not to any great extent, since the germinative epithelium, from which the definitive sex cells come, is removed, too. These experiments would be indeed weighty if the germinative epithelium were left intact, and it, after the removal of all other parts of the gonad, never produced any more germ cells.
There is very excellent evidence, on the other hand, that regeneration of reproductive tissue may occur in animals as complex as the mammals. - Davenport (’25) found that 16.8 per cent of ovariotomized mice regenerated ovaries. Parks et al. (’27) found that 9.1 per cent regenerated new ovaries after all precautions had been taken to see that all the ovarian tissue was removed and that the regeneration which subsequently occurred did so from somatic tissue, and not from residual pieces of the gland. Castle and Phillips (’11) observed that 7.8 per cent of guinea-pigs and rabbits regenerated new gonads. Jordan and Kindred (’26, p. 291) state sex that such evidence “compels the abandonment of the conception of a strict independence between germ cells and soma cells, and the doctrine of a specific germ-plasm and its continuity.”
Several cases of pregnancy following complete removal of both ovaries have been reported. In each instance the authors offer the explanation that a portion of the ovary must have been left behind, although at the time of the operation they were careful to remove all the gland. Robertson (’90), Kossman (’OO), Morris (’01) and Doran (’O2) all agree that some residual part of the gland must have escaped them at the operation. Davenport (’25) offers the additional explanation that even in humans there may be some regeneration of ovarian tissue from the peritoneal epithelium.
In view of the results on other animals, Where it has been clearly demonstrated that the ovaries reform from the peritoneal epithelium, as in Trionyx, Simkins (’25) and in amphibians as reported by many authors, it is not unreasonable to think that regeneration of the ovaries is a possibility deserving consideration.
Summary
- All cells that go into the fundaments of the indifferent human gonads are derived from the germinative epithelium.
- During the development of the indiflerent gonad the sex cords are formed and large primary genitaloid cells appear, associated with smaller spherical cells.
- The genitaloid cells degenerate in the seminiferous tubules, but form the interstitial cells of the testes when not incorporated into the tubules.
- The genitaloid cells produce precocious follicles in the full-term ovary, some of which may possibly rupture; most of them, however, become atretic.
- The definitive cells of both ovary and testis come from the smaller cells of the gonad and are transformed somatic cells.
- A strict independency of somatic and germ cells is not demonstrable in the history of the human gonads. 272 CLEVELAND SYLVESTER SIMKINS
Bibliography
CASTLE AND PHILLIPS 1911 On germinal transplantation in vertebrates. Carnegie Institution of Washington, publication no. 144.
DAVENPORT, C. B. 1925 Regeneration of ovaries in mice. Jour. Exp. Zool., vol. 42, pp. 1-12.
DORAN, A. 1902 Pregnancy after removal of both ovaries for cystic tumor. J our. Obs. Gyn. British Empire, vol. 2, pp. 1-10.
Felix W. The development of the urinogenital organs. In Keibel F. and Mall FP. Manual of Human Embryology II. (1912) J. B. Lippincott Company, Philadelphia. pp 752-979.
FUSS, A. 1911 U'ber extraregionare Geschlechtzellen bei einem menschlichen Embryo von vier Woehen. Anat. Anz., Bd. 39. 1913 Uber die Geschlechtzellen des Menschen und der Siiugetiere. Arch. f. mikr. Anatomie, Bd. 81.
HUMPHREY, R. R. 1927 Extirpation of the primordial germ cells of Amb1ystoma: its effect upon the development of the gonad. Jour. Exp. Zool., vol. 49, pp. 363-400.
JORDAN AND KINDRED 1926 Textbook of embryology. New York.
KOHNO, S. 1925 Zur Kenntnis der Keimbahn des Menschen. Arch. f. Gyniik., Bd. 126, S. 310-326.
KOLMER, W. fiber der Keimbahn des Menschen. Zentralb. f. Gynak., Leipzig, Bd. 49, S. 1875.
KOSSMAN 1900 Pregnancy and normal delivery one and one-half years after removal of both ovaries. Amer. Jour. Obs., vol. 41, p. 839.
MORRIS, M. M. 1901 Pregnancy following removal of both ovaries and tubes. Boston Med. Surg. Jour., vol. 144, p. 86.
PARKES, A. S., FIELDING, AND BRAMBELL 1927 Ovarian regeneration in the mouse after complete double ovariotomy. Proceedings Royal Society, B. vol. 101, pp. 328-354.
ROBERTSON, J. A. 1890 Renewal of menstruation and subsequent pregnancy after removal of both ovaries. Brit. Med. J our., vol. 2, p. 722.
RUBASCHKIN, W. 1908 Zur Frage von der Entstehung der Keimzellen bei Siiugetierembryonen. Anat. Anz., Bd. 32, S. 222-224.
1910 Uber das erste Auftreten und Migration der Keimzellen bei Siiugetierembryonen. Anat. Hefte, Bd. 41, S. 243.
SIMKINS, C. S. 1923 On the origin and migration of the so-called primordial germ cells in the mouse and the rat. Acta Zoologica, Bd. 4, S. 241-278.
— 1925 Origin of the germ cells in Trionyx. Am. Jour. Anat., vol. 36, pp. 185-211.
WILSON, K. M. 1926 Origin and development of the rete ova:-ii and the rete testis in the human embryo. Contri. Embryo., vol. 17, pp. 69-89.
Plates
- Simkins 1928: plate 1 | plate 2 | plate 3 | plate 4 | plate 5 | plate 6 | plate 7 | plate 8 | plate 9 | plate 10
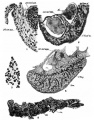
plate 1
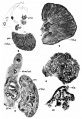
plate 2
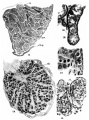
plate 3
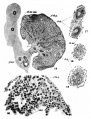
plate 4
plate 5
plate 6
plate 7
plate 8

plate 9
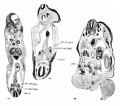
plate 10
Plate 1

All figures have been reduced approximately one-half.
1 Peritoneal epithelium of a human embryo of 5 mm. X 750. plinm-gen., urogenital fold; cor.m.9’nph., mesonephric body.
2 Cross-section of the peritoneal epithelium of a 7-mm. human embryo. X 750. gZ., glomeru1us;ms’em'.d., dorsal mesentery; e’th.p12’tn., peritoneal epithelium; cor.ms’nph., mesonephric body.
3 Types of cells in the ovary of a 43-mm. human embryo. X 750.
4 Longitudinal section of the genital thickening of a human embryo of 15 mm. X 200. gem, sex cord; e’th.g., germinative epithelium; re., rete.
5 Seminiferous tubule from an embryo of 190 mm. X 200. 11%., blood vessel; tbl., tubule, because of its contortion is out several times.
Plate 2

6 Cross~section of the gonad of a. human embryo of 18 mm. X 200. The mesentery lies to the right. gl’ml., glomerulus.
7 Cross—section of the ovary of a 43-mm. fetus. X 200.
8 Cross~section of the ovary of a 38-mm. human embryo. X 200.
9 Cross-section of a 20mm. human embryo. X 200. dt.ms’nph., mesonephrie duct. Miillerian duct lies above.
10 Genitaloid cells and small cells from a 38-mm. human ovary. X 750.
Plate 3

11 Cross-section of part of a human ovary of 30 mm. X 200.
12 Tubules from the testis of an infant three months postpartum. X 750
13 Large cells and small cells in detail from the ovary of a 30-mm. human. X 1500.
14 Cross~section of the testis of a fetus, 190 mm. X 40.
15 Detail of the tubules of a 190-mm. fetus. X 750.
Plate 4

16 Cross-section of the ovary of a 25~1nm. human embryo. X 200.
17 Primary follicles from an ovary at term. X 750.
18 Follicle forming in an ovary at term. X 750.
19 Beginning of secondary follicle in an ovary at term. X 750. (Figs. 17, 18, 19, 28, and 31, all taken from the same ovary.)
20 Detail of the cells in an ovary of 30 mm. X 750.
Plate 5

21 Cross—section of the testis of an embryo of 54 mm. X 200.
22 Detail of the large and small cells in the ovary of a 51-mm. embryo. X 1500.
Plate 6

23 Detail of the sex cords from the testis of a 70-mm. embryo. X 750.
24 The genitaloid cells (cl.g.) in the gonad of a 70-mm. human embryo. X 1500.
25 Cross-section of the ovary of a 90~mm. fetus. X 200.
26 Detail of the large and small cells in the gonad of a 25-mm. human embryo. X 750.
Plate 7

27 Detail of the large and small cells in the testis of a 54-mm. embryo. X 1500.
28 Secondary follicle from an ovary at term. X 750. A nucleus could not be found in the ovum.
29 Large and small cells in detail from the ovary of a 90-mm. fetus. X 750.
30 Grenitaloid cell in detail from the ovary of a 51-mm. embryo. )< 1500.
31 First indication of a cytoplasmic membrane (vitelline membrane) enclosing the ovum. From an ovary at term. X 350.
32 Mature ovum in the follicle of a full—term ovary: the walls of the follicle have not been shown. X 350. /m.vt., vitelline membrane.
33 Detail of tubule from a 54-mm. embryo. X 1500. cl.so., interstitial cell.
Plate 8

34 Cross-section of an ovary of a 51-mm. embryo. crd.s., sex cords. X 200.
35 Cross—section of the gonad of a 70-mm. embryo, showing the sex cords and large genitaloid cells. X 200.
36 Peripheral portion of the testis of a. 154-mm. human embryo. X 200.
37 Detail of a seminiferous tubule of a 154—mm. fetus. X 750.
Plate 9

38 Peripheral portion of an ovary from a seven-month fetus. X 750.
39 Peripheral portion of an ovary from a full-term fetus. cl.g., genitaloid cell. X 7 50.
40 Portion of an ovary from a fetus five months postpartum. X 750.
41 Cross-section of incipient tubules of a 54-mm. embryo. X 750.
42 Advanced follicle from an ovary at full term. X 200.
43 Detail of cells from the gonad of a 20-mm. human embryo. X 750. cl.g., genitaloid cell.
Plate 10

44 Cross—seetion of a 5-mm. human embryo through the mesonephros, showing the formation of the urogenital fold, plahun-gen., urogenital fold; c0r.ms’nph., mesonephric body; ao.d., dorsal aorta.
45 Cross—section through the mesonephric body of a 7.4—mm. human embryo.
Legends same as in figure 44.
46 Cross-section through the genital gland of a 15~mm. embryo. gem, genital gland.
Cite this page: Hill, M.A. (2024, April 20) Embryology Paper - Origin of the sex cells in man. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_Origin_of_the_sex_cells_in_man
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G