Paper - An Early Human Ovum (Thomson) in situ
| Embryology - 16 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Odgers PN. An early human ovum (Thomson) in situ. (1937) J Anat. 71(2): 161-168.3. PMID 17104634
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
An Early Human Ovum (Thomson) in situ
Odgers PN.
In March 1913 the late Prof. Arthur Thomson received the uterus, which contained this ovum, with its adnexa from Prof. H. M. Turnbull of the London Hospital. He briefly demonstrated it to the Anatomical Section of the XVIIth International Congress of Medicine, which was held in London in the summer of that same year, but he never published any detailed description of it.Some five years ago he suggested to me that it was well worth further examination, but it was not until after his death that the slides of this specimen with his serial microphotographs and notes about it were brought to my notice. I shall call it in this paper after his name, and I hope that, if it is ever referred to in the future, it may be known as the Thomson ovum.
Prof. Turnbull discovered it in the decidua of the posterior wall of the uterus of an unmarried woman, aged 20, who died 5 hours after severe burns on 4 March 1913. The right ovary contained a corpus luteum, which measured 24.5 x 17.5 mm. The woman's menstrual history was as follows. It first occurred at 15 years of age and had always been regular. Her last period commenced on 3 February, exactly 29 days before her death. No dates of coitus were available. The post-mortem examination took place 26 hours after death. The specimen was fixed in 10 per cent. formalin and cut into sections 10 thick.
From the menstrual history the maximal age of this ovum must be about 16 days. The inside measurements of the blastocyst are 2.1x 151 x 0.7 mm.; the embryonic disc measures 0.26 x 0.31 (?) x 0.16mm. (?).
In the following list it is placed among its contemporaries. Dr S. Zuckerman has kindly computed for me from the inside measurements of the blastocyst cavity a rough estimate of its cubic capacity in the several ova, the cavities being regarded for this purpose as perfect spheres, of which the diameters were taken as the mean of the dimensions given. These figures are added in a separate column.

As is seen in the general view (PI. I, fig. 1), the blastocyst cavity and the trophoblast are well preserved, but the embryonic disc, particularly the ectoderm, has suffered a good deal of disorganization, the amniotic cavity being partly obscured by cellular detritus. While this forbids any detailed description of the disc itself, I wish to draw attention here to:
- the formation of the extra-embryonic coelom, and
- the wall of the yolk sac.
I. The Extra-Embryonic Coelom

On either side of the amnio-embryonic vesicle a space is seen (P1. I, fig. 2), the one on the left being triangular in section, while the right one appears more quadrangular in shape. These are marked off from the rest of the blastocyst cavity by a regular layer of mesothelial cells, which vary in different sections in their number and their shape, most of them being round, while a minority are flatter and spindle-shaped. They are partially filed by coagulum similar to that of the magma. If the almost symmetrical channels thus formed are traced in one direction, they are seen to coalesce and surround this end of the embryonic disc (PI.I,fig.1), the common channel extending for thirteen sections beyond the disc: traced in the opposite direction they are observed to narrow and finally disappear, the right before the left, sixteen sections from the disc, but here they are always separated from each other by a strand of mesenchyme and never join (PI.I,fig.4).
Text-fig. 1 is a diagram drawn roughly to scale to show the disposition of these channels (ExC) around the embryonic disc (ED) viewed from the dorsalaspect. The letters A,B,C correspond to the levels of the microphotographs (PI. I, fig. 1; PI. I, fig. 2; and PI. I, fig. 4). While, as I have said, these channels are well marked out from the rest of the blastocyst cavity for the most part, there are in the case of either of them gaps in their parietes where they become continuous with the adjacent magma spaces. When traced in serial sections, such spaces are seen to be increasingly outlined by mesothelial cels so that eventually they appear as little bays in the lateral wall of the channel which is thereby correspondingly enlarged.
The only other specimen which shows anything exactly like this one is the Peters ovum, but that of Tennant & Ramsey presents a condition very comparable to it. The former of these was re-examined by Prof. 0. Grosser in 1908, and he-has been good enough to send me his microphotograph of this ovum and very kindly permits me tore produce it here for comparison (PI.I,fig.2).
The only differences in the appearance presented by these two sections are that in the Peters ovum (1) the lateral wall of the right channel has been torn away, (2) the containing wall of the intact left channel appears to be more celular, and (3) itextends on to the ventral wall of the yolk sac, while in the Thomson ovum, although they are wider, these channels are confined to the neighbourhood of the amnio-embryonic vesicle. Grosser described the channel in the Peters ovum as horseshoe-shaped; itsurrounds the cranial end of the disc as a small fissure. This is rather different from the wide communication presented by the present specimen.
Similarly, Tennant & Ramsey described "a space around the embryonic mass which is limited by a single layer of flattened mesodermal cells having the appearance of an endothelium (PI.II, fig. 3).This layer not only surrounds the space but likewise completely encloses those surfaces of the embryonic mass not directly attached to the mesenchyme." In their specimen, too, the space is horseshoe-shaped, as must be inferred from the fact that they found that "the left end of the embryonic disc stops bluntly and is entirely free of meso- thelial attachment while the right is completely encased in mesenchyme".
The three ova, Thomson, Peters and Tennant & Ramsey, seem to represent three stages in the development of these channels. In the first they border the amnio-embryonic vesicle only, in the second they have extended on to the lateral side of the yolk sac, while in the last they practically enclose the yolk sac on all sides. Here, although the authors describe a space, I think there is a fine strand in the microphotograph they reproduce between the mesothelial layer covering the ventral wall of the yolk sac and the parietes of the space to the right of the midline which may indicate its bilateral origin (PI.II, fig. 3), while in their reconstruction they show the ventral surface of the embryo as almost entirely free of mesenchyme, "with the exception of a small band of attach- ment extending about half way along this surface from the encapsulated end ".
In 1908 Keibel & Elze and Grosser described these channels as the com- mencement of the extra-embryonic coelom. Grosser (1924) suggested that they gradually grow ventrallyr ound the yolk sac, their opposed walls possibly forming, when they meet, the "distaler Nabelblasenfaden", the mesodermal strand which may connect the ventral pole of the yolk sac with the chorion. Further, he thought that these exocoelomic channels extend lateralwards by taking up the magma spaces nearest to them so that eventually the whole cavity of the blastocyst becomes the extra-embryonic coelom. He wondered if the loss of the lateral wall in the left channel in the Peters embryo might not be due to its having already broken down so that on this side the exocoelom was continuous with the intercellular spaces of the magma. I have mentioned above that in the Thomson ovum there is evidence of the commencement of the same process, while Tennant & Ramsey wrote that in the specimen they described "on the surface towards the main chorionic cavity the continuity of this membrane-like structure is less well maintained and irregularly distributed openings in it offer probable avenues of communication between the larger and the smaller spaces ".
These three ova are peculiar then in being the only ones so far described which exhibit definite symmetrical coelomic channels.[1] The Fetzer (Grosser, 1924) and Herzog (Lewis, 1912) ova and the much more advanced embryo described by Waterston (Mall, 1916) have al been considered by these
different observers to show something similar, but I would suggest that at al events the illustrations, which accompany the descriptions of these specimens, are far from convincing from this point of view. In al other specimens the whole of the blastocyst cavity seems to represent the extra-embryonic coelom (Bryce,1924-5). An accumulation of magma fluid appears according to this author's account near its centre close to the embryonic disc, which gradually leads to the stretching and absorption of the mesodermal bands traversing it and to the isolation of the embryo, so that ultimately all the magma spaces run together and the whole of the magma cavity becomes the exocoelom. Grosser (1913) and v. Mollendorif (1921) both suggested that there are thus two alternative ways in which this coelom may be formed.
It is, however, perhaps pertinent to observe that in the Peters ovum the trophoblast is already lined with "morula" mesoderm (Stieve, 1926), which also covers the ventral wall of the yolk sac beyond the limits of the exo- coelomic channel. The Thomson ovum presents the same appearance, although the mesodermal cells covering the yolk sac are few. In either of these cases, apparently, a gradual liquefaction of the magma would result in a perfect coelom lined with mesoderm without the help of the mesothelium of the channels at all.
With regard to the later fate of these exocoelomic channels there is only Grosser's surmise to guide us, and one can not help thinking that their formation as seen in the three specimens in which they occur serves no useful purpose. In any case their appearance in the Tennant-Ramsey and Thomson ova prove that the Peters ovum isno longer quite exceptional.
II. The Yolk Sac
As seen in PI. I, fig. 2, this appears as a space beneath the amnio-embryonic disc, outlined from the rest of the magma cavity by a thin membranous wall. It is flattened dorsoventrally. On the left side, its ventral wallis connected by an irregular strand of mesenchyme cels to the chorion, but this is the only evidence of a commencing "Nabelblasenfaden" so obvious in the Schlagenhaufer-Verocay specimen. While the dorsal wall of the yolk sac beneath the embryonic disc has a lining of cubical entodermal epithelium its lateral and ventral walls are formed by a sparse layer of flattened spindle- shaped cells mesothelial in character. These latter appear to be similar to many of the cels lining the extra-embryonic coelomic channels, and the coagulum within the yolk sac seems to be exactly like that filing these channels and the magma spaces. Clothing the lateral walls and extending on to the ventral angles of the sac are a few round mesodermal cells. It is interesting to compare the condition found here with that noted or illustrated in the other ova in the list given above.
Linzenmeier. There is no mention of this point in the text, but the accom- panying illustration shows flattened cells on the ventral side of the yolk sac with a cubical entoderm under the embryonic disc.
Stieve. "The yolk sac is outlined by spindle-shaped mesoderm cells, while the entoderm is only represented by a single layer beneath the ectoderm of the disc."
Peters. Here the whole sac is lined by a cubical epithelium of the same type alround (P1.I,fig.2). In Peters' original drawing the cells on the ventral
wall are more irregularly spaced than are those beneath the disc.
v. Milendorif, EiOP. "In its whole circumference the yolk sac is lined with markedly flattened cells with lens-shaped nuclei." His figure shows a condition very much like that I have described in the Thomson ovum.
Fetzer. The sac is lined except beneath the disc with a "thin low entoderm". Sporadically the cells are larger, cubical or short cylindrical in type (Fetzer & Florian).
Falkiner. He gives no description of this point, but his illustration shows the lining of the yolk sac very much like that in the Thomson ovum.
Tennant & Ramsey. "The yolk sac is lined with entodermal cels, which hare cuboidal on the dorsal surface and flattened on the ventral surface"(P1.I,fig.3).
Herzog. Lewis (1912) described the sac in this ovum thus: "Over the greater part of it the entoderm forms a very thin layer resembling endothelium."
Heine & Hofbauer.The yolk sac is lined by a single layer of flattened cells except under the disc.
Schlagenhaufer & Verocay.The yolk sac is lined throughout with entoderm.
Jung. The yolk sac shows a simple layer of flat entoderm cells.
With the Peters and Schagenhaufer ova standing out as exceptions this analysis confirms Bryce's (1924-5) statement that in the first instance the yolk sac is always lined with flattened endothelial-like cels. In older embryos, as Grosser (1924) pointed out, there is a complete change; in these (e.g. Rossen- beck, Grosser and Graf Spee ova) while the peripheral part of the sac is lined with cuboidal cells the portion of it beneath the disc has the flatter type of epithelium. Similarly Florian & Hill (1934-5) described the yolk sac wall in their specimen-" while the endoderm of its cranial and lateral walls is formed of flattened cels,that of its ventral wall is distinctly higher."
In attempting to explain these differences Stieve interpreted the appearance presented by the ovum (Werner) he described as follows. The future yolk sac is here represented by an enlarged magma space lying beneath the embryonic disc and outlined by mesoderm cels. Later the entoderm of the disc must gradually grow ventrally and surround this space on its inner aspect and so form a definite yolk sac. He followed Corning (1925), who described the entoderm in the human ovum as beginning as an entodermal plate-the lowest layer of cels of the embryonic disc-rather than as an entodermal vesicle. In favour of such a conception are, I think, the following points:
- It brings the formation of the human yolk sac in line with that which has been found to occur in most other mammals (Stieve).The only observations inc omparative embryology, as far as I am aware,which suggest the development of the yolk sac by the dehiscence or vacuolization of a primitively solid entodermal mass are those of Hubrecht (1889-90) in Erinaceus and Hill (1932) in Nasalislarvatus.
- The entodermal vesicle of the Teacher-Bryce ovum (TB. I.) lying in the magma and divorced from the amnio-embryonic vesicle might very well be described as a magma space (Stieve). Bryce (1924-5) himself described it as being lined with flattened cels precisely like those forming the wall of the sac in Linzenmeier's embryo.
- In the Tennant-Ramsey ovum the cubical entoderm appears to be surrounding the yolk sac and to have left only a portion of its ventral wall lined by flattened cells (PI.I, fig.3).
- In nearly al the early ova the yolk sac, lined with flat mesothelial-like cels,is filed with a loose coagulum which seems to be indistinguishablefrom that of the magma spaces (Triepel, 1916). On the other hand Schlagenhaufer & Verocay found in their ovum that the contents of the yolk sac stained more deeply with eosin than did that of the magma. Tennant & Ramsey described the yolk sac in their specimen as "partially filed by pink-staining finely granular material, while in other sections large yolk granules are present in the cavity". Peters figured some drops of (?) yolk-like material in his original drawing of the yolk sac in his ovum. In all these three instances, as has been noted already, the sac is lined more or less completely with a cubical entoderm. One might, therefore, suggest that there is some correlation between the character of the contents of the yolk sac and of its cellular lining.
The appearance of the yolk sac in the Thomson ovum isvery much like that shown in Stieve's illustration. On Prof. J. P. Hill's advice I ventured to send the serial microphotographs of this ovum to Prof. Grosser, and he was kind enough to give me his opinion on this point after examining them. His criticism of both the Thomson and the Stieve specimen is the same. Inboth, according to him, the entodermal lining of the yolk sac has disappeared except under the disc. I confess I think this explanation is a difficult one. Allowing that 26 hours elapsed between the woman's death and the fixation of the Thomson ovum, the entoderm could scarcely have vanished in this time without leaving a trace. If the cels had been shed, they or their debris should surely still be found in the yolk sac, unless its walls were broken. But the yolk sac appears to be intact, filed with coagulum, but containing only an occasional detached cel. If there had been a dehiscence in its wall, the fluid content would certainly have escaped before its cellular lining. Further, as I have shown above, in most of the early ova the cels lining the yolk sac are described as being of this same character, and, while in different specimens they vary somewhat both quantitatively and qualitatively, most embryologists would follow Bryce indescribing them as flattened and endothelial-like. But, if one accepts Grosser's explanation for the Stieve and Thomson ova, is one to condemn others of about the same age which show a similar condition? One is left apparently with two alternatives. There is, on the one hand, the view generally held that these mesothelial-like cels lining the early yolk sac, so similar to many of those outlining the exocoelomic channels and to those scattered elsewhere throughout the blastocyst cavity, grow eventually into the cubical or low columnar ettoderm of the older sacs. On the other hand, there is the conception of Corning and Stieve of a yolk sac originally preformed asamagma space and outlined in mesoderm,which later acquires an epithelial lining from the growing margins of an entodermal plate. This latter idea apparently has much to recommend itand I should prefer myself to interpret this specimen as rather corroborating it.
While I realize to the full the weight that rightly should be attached to Prof. Grosser's opinion, I would suggest that the appearance of the cellular lining of the yolk sac in this ovum is not so remarkable.
Summary
- The interior measurements of the blastocyst cavity in the Thomson ovum are 2-1x 1-51 x 07 mm.; itsmaximal age isabout 16 days.
- Theextra-embryonic coelom is here seen to be commencing as fairly symmetrical channels on either side of the amnio-embryonic vesicle. A comparable appearance to this has been observed previously only in the Peters and Tennant-Ramsey ova.
- The yolk sac has a lining of cubical entoderm beneath the disc; elsewhere it is merely outlined by a sparse layer of spindle-shaped mesothelial-like cells.This condition is shown not to be exceptional in the earlier ova and is held to corroborate Stieve's conception of an entodermal plate spreading ventrally to form the proper entodermal wall of the yolk sac as seen in later embryos.
I am very much indebted to Prof. J.P. Hill for his kindness in examining the slides of this specimen, and to Prof. 0. Grosser for giving me his opinion on the serial microphotographs of it, and for his kind permission to reproduce his own photograph of the Peters ovum. I must also thank the publishers of Surgery, Gynecology and Obstetrics for allowing me to reproduce the illustration, PI.I,fig.3,which appeared originallyin thatjournal. The other micro- photographs are the work of Mr W. Chesterman of this Department.
References
BRYCE, T. H. (1924-5). Tranm. roy. Soc. Edinb. VOL. LM, pt. 3, p. 533.
CORNING, H. K. (1925). Lehrbtuch d. Entwicklungsgesch. d. Menwchen, 2. Aufl. Munchen, S. 79.
FAijWER, N. M. (1932). J. Obdtet. Gynaec. vol. XXXIX, p. 471.
FETZER, M.(1910). Verh.anat.Ges.Bd.xxxvii,S.116.
FETZER, M. & FLORIAN, J. (1930). Z. miikr.-anat. Forsch. Bd. xxi, S. 351.
FLORIAN,J.& HILL, J.P. (1934-5). J.Anat.,Lond., Vol.LXIX, p.399.
GROSSER,0.(1908). Vergleich.Anat.u.Entwicklungsgesch.d. Eihduteu. d. Placenta. Wienu. Leipzig.
- (1913). Anat.Hefte,Bd.XLVII,S.649.
- (1924). Z.ges.Anat.3Abt.Bd.xxv,S.391.
- (1927). Frilentwickllung, Eihautbildung u. Placentation d. menschen u. d. Sdugetiere. Munchen.
HEINE & HOFBAUER, J. (1911). Z. Geburtsh. Gyndk. Bd. LXVIII, S. 665.
HERZOG, M. (1909). Amer. J. Anat. vol. IX, p. 361.
HILL, J.P. (1932). Philos. Trans. B, vol. CCXXI, p.108.
HUBRECHT,A.A.W. (1889-90). Quart. J. micr. Sci. vol. XXX, p.283.
JUNG,H.(1907). Mschr. Geburtsh. Gynak. Bd. XXV, S.901.
KEIBEL, F. & ELZE, K. (1908). Normentafel z. Entwicklungsgesch. d. Menschen. Jena.
LEWIS, F.T. (1912). Manual of Human Embryology, vol.I, p.298. Keibel and Mall.
LINZENMEIER, G. (1914). Arch.Gynak.Bd.CI,S.1.
MALL, F.P. (1916). Contrib. to Embryology, vol.IV.
V. MOLLENDORFF, W. (1921). Z. ges. Anat. Abt. 1, Bd. LXIT, S. 406.
PETERS, H. (1899). Uber d. Einbettung d. menschlichen Eies. Leipzig u. Wien.
SCHLAGENHAUFER & VEROCAY (1916). Arch. Gyndk. Bd. cv, S. 151.
STIEVE,H.(1926). Z.Mikr.Anat.Bd.VI,S.295.
- (1931). Verh.anat.Ges.Bd.XL,S.44.
TENNANT, R. & RAMSEY, E. M. (1934). Surg. Gynec. and Obstetr. vol. LVIII, P. 968.
THOMSON, A. (1913). XVII th Internat. Congress of Medicine, Section 1, pt.2, p.28.
TRIEPEL, H. (1916). Anat. Hefte, Bd.LIV, S.149.
Explanation Of Plates
Plate I

Fig. 1. The section photographed is through the middle of the blastocyst cavity (x44).
Fig. 2. The same section as fig. 1 (x 100). This shows the exocoelomic channels, ExC, on either side of the embryonic disc and ventral to this the yolk sac, YS. Above the disc is the amniotic cavity obscured by cellular debris which partially fills it. This section corresponds to the plane lettered B in Text-fig.1.

Fig. 1
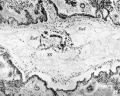
Fig. 2


Plate II
Fig. 1. A section (x100) 20, u beyond one extremity of the embryonic disc. It corresponds to the plane A in Text-fig.1 and shows that the two exocoelomic channels have joined round this end of the disc.
Fig. 2. A photograph of a section (x100) of the Peters ovum, kindly sent me by Prof.0. Grosser. It shows the exocoelomic channel on the left intact, bordering the embryonic disc and the yolk sac, while that on the right has only its medial wall left.
Fig. 3. A section (x 150 ?)ofthe Tennant-Ramsey ovum (reproduced by courtesy of the Editors of Surgery, Gynecology and Obstetrics). It shows the amniotic cavity, the embryonic disc and the yolk sac surrounded on either side and ventrally by the extra-embryonic coelom.
Fig. 4. A section (x100) 20/.z beyond the opposite extremity of the disc showing the two channels still separate from each other. This corresponds to the plane C in Text-fig.1.
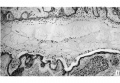
Fig. 1
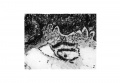
Fig. 2

Fig. 3
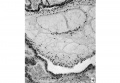
Fig. 4




Reference
Odgers PN. An early human ovum (Thomson) in situ. (1937) J Anat. 71(2): 161-168.3. PMID 17104634
Cite this page: Hill, M.A. (2024, April 16) Embryology Paper - An Early Human Ovum (Thomson) in situ. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Paper_-_An_Early_Human_Ovum_(Thomson)_in_situ
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G
- ↑ Since this was written K. Hiramatsu (Fol. Anat. Japon, 1936, Bd. XIV, p. 15) has published an account of the ovum, Ei-Ando. In this he describes two small cavities on either side of the dorsal portion of the yolk sac, which he thinks correspond to these exocoelomic channels.