Developmental Signals - TGF-beta
| Embryology - 19 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Introduction
Transforming Growth Factor-beta (TGF-beta, TGF-β) this is a superfamily signaling pathway with many different roles during development.

TGF Members: AMH
| Factor Links: AMH | hCG | BMP | sonic hedgehog | bHLH | HOX | FGF | FOX | Hippo | LIM | Nanog | NGF | Nodal | Notch | PAX | retinoic acid | SIX | Slit2/Robo1 | SOX | TBX | TGF-beta | VEGF | WNT | Category:Molecular |
Some Recent Findings
|
| More recent papers |
|---|
 This table allows an automated computer search of the external PubMed database using the listed "Search term" text link.
More? References | Discussion Page | Journal Searches | 2019 References | 2020 References Search term: TGF-β Embryology <pubmed limit=5>TGF-β Embryology</pubmed> |
Structure
The TGF precursor protein has three distinct regions:
- signal peptide - targets it to the endoplasmic reticulum and secretion
- propeptide - or the latency associated peptide
- mature peptide - cleaved from the precursor protein and is actively involved in signalling
- cleaved by Furin - a convertase
- cleaved at a dibasic arginine-X-X-arginine (RXXR) site
Function
Implantation
Expressed at the fetal-maternal interface and implicated in the promotion of implantation and post-implantation development.[5]
Signaling Pathway
Receptor
- active peptide forms a hetero- or homodimer
- binds to a specific TGF-β Type II receptor
- Type II receptor then recruits a TGF-β Type I receptor
- phosphorylates it via its serine/threonine kinase domain
- phosphorylated Type I receptors then phosphorylate receptor-associated Smad proteins (R-Smads), including Smad1/5 and Smad2/3
- Type II receptor - MlTgfRII
- Type I receptors - MlTgfRIa, MlTgfRIb, and MlTgfRIc
Intracellular Signaling
- R-Smad proteins are composed of two main functional domains
- Mad-homology domains 1 and 2 (MH1 and MH2)
- Smad1/5 - associated with BMP-like signalling.
- Smad2/3 - associated with TGF-β-like signaling.
SMAD
SMAD5
Mothers against decapentaplegic homolog 5 (SMAD5) is a transcriptional modulator activated by BMP (bone morphogenetic proteins) type 1 receptor kinase.
OMIM: SMAD5
Bone morphogenetic protein 15

BMP15 evolution among family members[7]
Growth Differentiation Factor 1
Growth Differentiation Factor 9 (GDF1) in involved in early left-right patterning.[8]
Growth Differentiation Factor 9

Growth Differentiation Factor 9 (GDF9) and bone morphogenetic protein 15 (BMP15) are both members of the transforming growth factor-β (TGF-β) superfamily. They have both been identified as growth factors required oocyte to granulosa cell signaling for ovarian follicle development (folliculogenesis).[10][11][12]
A recent study in pig has shown that oocyte-derived BMP15 but not GDF9 is required for down-regulation of connexin43 expression leading to a decrease in intercellular gap junction communication in granulosa cells.[3]
- Links: TGF-beta | Bone Morphogenetic Protein | oocyte | ovary | menstrual cycle | OMIM - GDF9
Nodal
A secretory protein (gene 10q22.1) important for gastrulation.[13] Nodal forms a heterodimer with Vg1 (GDF3) required for formation and mesendoderm induction.[14]
OMIM
About OMIM "Online Mendelian Inheritance in Man OMIM is a comprehensive, authoritative, and timely compendium of human genes and genetic phenotypes. The full-text, referenced overviews in OMIM contain information on all known mendelian disorders and over 12,000 genes. OMIM focuses on the relationship between phenotype and genotype. It is updated daily, and the entries contain copious links to other genetics resources." OMIM
References
- ↑ Pang K, Ryan JF, Baxevanis AD & Martindale MQ. (2011). Evolution of the TGF-β signaling pathway and its potential role in the ctenophore, Mnemiopsis leidyi. PLoS ONE , 6, e24152. PMID: 21931657 DOI.
- ↑ Urban DJ, Anthwal N, Luo ZX, Maier JA, Sadier A, Tucker AS & Sears KE. (2017). A new developmental mechanism for the separation of the mammalian middle ear ossicles from the jaw. Proc. Biol. Sci. , 284, . PMID: 28179517 DOI.
- ↑ 3.0 3.1 Chang HM, Cheng JC, Taylor E & Leung PC. (2014). Oocyte-derived BMP15 but not GDF9 down-regulates connexin43 expression and decreases gap junction intercellular communication activity in immortalized human granulosa cells. Mol. Hum. Reprod. , 20, 373-83. PMID: 24413384 DOI.
- ↑ Islam SS, Mokhtari RB, Kumar S, Maalouf J, Arab S, Yeger H & Farhat WA. (2013). Spatio-temporal distribution of Smads and role of Smads/TGF-β/BMP-4 in the regulation of mouse bladder organogenesis. PLoS ONE , 8, e61340. PMID: 23620745 DOI.
- ↑ Jones RL, Stoikos C, Findlay JK & Salamonsen LA. (2006). TGF-beta superfamily expression and actions in the endometrium and placenta. Reproduction , 132, 217-32. PMID: 16885531 DOI.
- ↑ Pang K, Ryan JF, Baxevanis AD, Martindale MQ (2011) Evolution of the TGF-β Signaling Pathway and Its Potential Role in the Ctenophore, Mnemiopsis leidyi. PLoS ONE 6(9): e24152 PLoS ONE
- ↑ Auclair S, Rossetti R, Meslin C, Monestier O, Di Pasquale E, Pascal G, Persani L & Fabre S. (2013). Positive selection in bone morphogenetic protein 15 targets a natural mutation associated with primary ovarian insufficiency in human. PLoS ONE , 8, e78199. PMID: 24147118 DOI.
- ↑ Rankin CT, Bunton T, Lawler AM & Lee SJ. (2000). Regulation of left-right patterning in mice by growth/differentiation factor-1. Nat. Genet. , 24, 262-5. PMID: 10700179 DOI.
- ↑ Hosoe M, Kaneyama K, Ushizawa K, Hayashi KG & Takahashi T. (2011). Quantitative analysis of bone morphogenetic protein 15 (BMP15) and growth differentiation factor 9 (GDF9) gene expression in calf and adult bovine ovaries. Reprod. Biol. Endocrinol. , 9, 33. PMID: 21401961 DOI.
- ↑ McNatty KP, Moore LG, Hudson NL, Quirke LD, Lawrence SB, Reader K, Hanrahan JP, Smith P, Groome NP, Laitinen M, Ritvos O & Juengel JL. (2004). The oocyte and its role in regulating ovulation rate: a new paradigm in reproductive biology. Reproduction , 128, 379-86. PMID: 15454632 DOI.
- ↑ Lodemann E & Wacker A. (1970). [Interferon-induction by Poly (A)-2 Poly (I)]. Naturwissenschaften , 57, 673. PMID: 5531364
- ↑ Lin ZL, Li YH, Xu YN, Wang QL, Namgoong S, Cui XS & Kim NH. (2014). Effects of growth differentiation factor 9 and bone morphogenetic protein 15 on the in vitro maturation of porcine oocytes. Reprod. Domest. Anim. , 49, 219-27. PMID: 24313324 DOI.
- ↑ Montague TG & Schier AF. (2017). Vg1-Nodal heterodimers are the endogenous inducers of mesendoderm. Elife , 6, . PMID: 29140251 DOI.
- ↑ Wei S & Wang Q. (2018). Molecular regulation of Nodal signaling during mesendoderm formation. Acta Biochim. Biophys. Sin. (Shanghai) , 50, 74-81. PMID: 29206913 DOI.
Reviews
Cunha SI, Magnusson PU, Dejana E & Lampugnani MG. (2017). Deregulated TGF-β/BMP Signaling in Vascular Malformations. Circ. Res. , 121, 981-999. PMID: 28963191 DOI.
Articles
Huminiecki L, Goldovsky L, Freilich S, Moustakas A, Ouzounis C & Heldin CH. (2009). Emergence, development and diversification of the TGF-beta signalling pathway within the animal kingdom. BMC Evol. Biol. , 9, 28. PMID: 19192293 DOI.
Gadue P, Huber TL, Paddison PJ & Keller GM. (2006). Wnt and TGF-beta signaling are required for the induction of an in vitro model of primitive streak formation using embryonic stem cells. Proc. Natl. Acad. Sci. U.S.A. , 103, 16806-11. PMID: 17077151 DOI.
Online Textbooks
- Molecular Biology of the Cell. 4th edition. Alberts B, Johnson A, Lewis J, et al.New York: Garland Science; 2002. Signal Proteins of the TGF-β Superfamily Act Through Receptor Serine/Threonine Kinases and Smads
- Molecular Cell Biology. 4th edition. Lodish H, Berk A, Zipursky SL, et al.New York: W. H. Freeman; 2000. Figure 24-20 TGFβ signaling
- Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000. TGFβ-dependent Epithelial-Mesenchymal Transition
Search PubMed
Additional Images
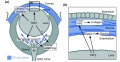
Mouse eye TGF-beta model

External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
- Abcam TGF-Beta Antibodies
- SABiosciences TGF-Beta Pathway
- RnD Systems Transforming Growth Factor Beta 1
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2024, April 19) Embryology Developmental Signals - TGF-beta. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Developmental_Signals_-_TGF-beta
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G