Book - Contributions to Embryology Carnegie Institution No.39: Difference between revisions
| Line 6: | Line 6: | ||
--[[User:Z8600021|Mark Hill]] 12:23, 14 April 2012 (EST) | --[[User:Z8600021|Mark Hill]] 12:23, 14 April 2012 (EST) Embryo No. 460 is [[Carnegie stage 20]]. | ||
Revision as of 07:56, 15 April 2012
The Cartilaginous Skull Of A Human Embryo Twenty-One Millimeters In Length
by Warren H. Lewis.
--Mark Hill 12:23, 14 April 2012 (EST) Embryo No. 460 is Carnegie stage 20.
| Historic Disclaimer - information about historic embryology pages |
|---|
 |
Introduction

The human skull has for generations attracted the attention of anatomists. From the imaginative comparative anatomists, imbued with the doctrine of the segmentation theory, we have inherited a bulky literature, gradually passing into oblivion, on the segments of an unsegmented skull. Its effects, however, are still to be noticed in the attempts to imagine a segmentation in the unsegmented brain. In the slowly accumulating literature on the cartilaginous skull of man and other vertebrates another phase is manifested. It was anticipated that in the so-called primordial or cartilaginous skull there would be found many indications of the phylogenetic relationships, and by a comparison of the cartilaginous skulls of the various vertebrates Gaupp and lus school expected to show this even more clearly. The theory expressed in the terse phrase, "ontogeny repeats phylogeny," formed the basis for such expectations. We have, however, gradually come to realize that there is more untruth than truth in it. For example, how is it possible to fit the conditions found in embryo No. 460 (Carnegie Collection), which are shown in figures 7 and 8, to the theory that "ontogeny repeats phylogeny?" Who would claim that our primitive ancestors had more brains than skull, except perhaps the few who believe in the downfall of man? And yet in this embryo the brain is enormous in size as compared with the cartilaginous skull or "primordial cranium." Even with the maximum development of the cartilaginous skull the conditions are essentially the same.
Comparative anatomy shows that in lower mammals and vertebrates the skull is relatively large as compared with. the brain, while human ontogeny shows exactly the reverse. The whole assumption that "ontogeny repeats phylogeny" was based upon the erroneous notions concerning evolution that were prevalent before the present-day conceptions of the germ-plasm were introduced. If the various steps in evolution have come about primarily through the modification of the germplasm, then we should expect changes to appear in the egg and in the subsequent stages of ontogeny, and the entire development would thus be modified as much as the adult. There undoubtedly are fleeting indications of our primitive ancestors in the development of the embryo, but they are not very numerous and are usually extremely difficult of interpretation.
It is probable that in the phlyogenetic history some sort of a membranous skull preceded the cartilaginous skull, and the latter preceded the osseous; but it is apparent from recent studies on vertebrate cartilaginous skulls that they no more form a phylogenetic series than do the adult skulls of the same species. The series of cartilaginous human skulls with which we are now familiar, modeled from embryos varying in length from 13 to 80 mm., has failed to add much or any additional phylogenetic evidence regarding the form of the skulls of our remote ancestors. In fact, these cartilaginous skulls are as characteristically human as the adult skull is human. It is becoming more and more clear, as our knowledge of the anatomy of the human embryo increases, that both it and its various organs are at all stages as characteristically human as are the adult body and its organs. One can distinguish with ease between the cartilaginous skull of man and that of the ape, the pig, the cat, the rabbit, or the mole, as each is as characterLstically formed as are the adult skulls of the same species. Homologies and similarities are to be found in the cartilaginous skulls just as in the adult skulls, but it is doubtful if much additional evidence of phylogenetic relationships will be revealed by a comparison of cartilaginous skulls.
The great need in the embryology of the human skull, as of other vertebrates, is a more complete and detailed series of the various stages, showing the gradual development from the primitive membranous stages to the adult. The present communication is concerned primarily with a particular stage that helps to fill in one of the many gaps still existing in the literature. The study of each stage is necessarily laborious, since it is practically impossible to picture such complicated forms without resort to models. The various structures in the head and neck, including the cartilaginous skull of this embryo, 21 mm. in length, were modeled with the plaster of paris technique.

Table 1 includes all of the human chondrocrania that have been reconstructed and described within recent years.
The Chondrocranium as a Whole
The chondrocranium at this stage forms a continuous mass of cartilage and precartilage. It constitutes but a small part of the brain covering, as does that part of the adult cranium which is o.ssified in cartilage. One can best understand the cartilaginous skull by comparing it with that portion of the human adult skull which is ossified in cartilage rather than with the cartilaginous skulls of the lower vertebrates. To do this with accuracy and precision an extensive series of stages intervening between the stage to be compared and the adult should be at hand. Unfortunately, but few stages are known, although in man more has been done than with any other vertebrate. We have in humaun anatomy a large literature on the ossification centers of both the cartilaginous and the membranous bones. This is of course, a great help towards understanding the cartilaginous skull. Such a literature for any of the lower vertebrates is scarcely existent, and therefore very little is now to be gained through a comparison of the cartilaginous skulls of different species.
It is well to bear in mind the great difference in size between the cartilaginous skull of the embryo and that part of the adult skull ossified in cartilage. Table 2 gives a few comparative measurements. Unfortunately, the embryonic measurements are all from the one cartilaginous skull under consideration; it would be most desirable to have several such specimens, since the size of the skull varies considerably in embryos of corresponding ages. To realize the extent of this variability one has but to compare the length of the basal plate, from the foramen magnum to the hypophysial canal, in embryos of approximately the same age. Similar differences exist in the adult, and the adult measurements here used are the averages for six skulls.

Table 2. gives a few comparative measurements from the one cartilaginous skull.
These figures serve to indicate in a general way the greater immaturity of the anterior end of the cartilaginous skull at this stage. The adult basioccipital is about 13 times as long as the cartilaginous basioccipital; the adult nasal septum about 30 times as long as the embryonic nasal septum. In agreement with this we find that the basioccipital consists of more advanced cartilage than the nasal septum. In one of the following paragraphs I have stated reasons for believing that the alar process of the sphenoid at a later stage becomes incorporated into the body of the sphenoid; this is partly substantiated by comparing the ratios for the distances between the alar processes of the embryo and between the Ungulse of the adult with the width between the outer edges of the optic foraminae or the distance between the foraminae rotundte (the latter formed in precartilage in the embryo) in both embryo and adult. These ratios are the same in each case, namely, 1:14, thus indicating that if the rate of growth in each of these regions of the sphenoid is the same the alar process must be looked upon as ultimately forming the lateral part of the body of the sphenoid. If, however, we compare the width of the basisphenoid in the embryo (not including the alar processes) with that of the adult (including the lingulse) it will he seen that the ratio is 1 :25, which corresponds exactly to the ratio for the width of the basioccipital between the otic capsules in the embryo and in the adult. One would naturally expect the two to grow at about the same rate and, if this is true, then the alar process should not be looked upon as ultimately forming part of the basisphenoid. Factors such as the possible pressure of the otic capsules on this region of the basioccipital may, however, account for the discrepancy, since the ratio for the distance between the hypoglossal foramina? (1 :15) is about the same as that for the basisphenoid, including in the embryo the alar processes and in the adult the Ungulae.
Dorsal Aspect



The most familiar aspect of the chondrocranium is the upper surface of the base, shown in figures 1 and 2. These figures should be compared with a some-what similar view of the upper surface of the base of the adult skull shown in figure 4, where the bone, ossified in cartilage, is colored blue. One is surprised, somewhat startled in fact, by the general similarity between the cartilaginous skull of the embryo and that part of the adult skull which is ossified in cartilage. The general resemblance is even more marked in later stages when the chondrocranium is more completely developed, as shown in the figures by Macklin and Hertwig of skulls from embryos 40 and 80 mm. in length. Again, if the dorsal aspect of the base of the entire skull, as shown in figure 3, is compared with that of the adult, new resemblances will appear. In figure 3 is shown not only the cartilage, but also the precartilage, blastema, and part of the dorsal membrane which enters into the formation of the brain capsule. In comparing the adult skull with figures 1 and 3, it must be borne in mind that these figures are drawn with the basioccipital horizontal and not inclined, as in the usual view of the adult skull or in the view of the cartilaginous skull shown in figure 2. At the caudal end of the cartilaginous skull is the basioccipital with the two lateral parts, or exoccipitals, one on each side. The exoccipitals are continued into the occipital squamse or nuchal plates which are not yet united in the middorsal line, so that the foramen magnum is incomplete. The basioccipital is continuous in front into the basisphenoid, and from it the two wings project on each side. The mesethmoid is continuous from the basisphenoid, without line of demarcation, to the anterior end of the chondrocranium, and connected with it are the nasal capsules. Lateral to the anterior half of the basioccipntal and fused with it are the large otic capsules corresponding to the petrous bones of the adult, each being separated from the exoccipital by the large jugular foramen and the mastoid plate.
The basioccipital consists of a flattened plate extending from the foramen magnum to the basisphenoid. The caudal edge bordering the foramen magnum is more deeply notched than in the adult. This notch is continued into a deep groove on the cranial side of the cartilage. At the apex of the groove is the opening where the notochord, after passing through the condensed mesenchyme which fills the groove, enters the body of the occipital.
Both in the embryo and in the adult each exoccipital arises from the side of the posterior half of the basioccipital and is perforated at its origin by the hypoglossal canal. In this embryo the right hypoglossal canal is divided on the cranial side into two parts by a cartilaginous bar; on the pharyngeal side there is but a single orifice, a condition not uncommon in the adult. The axis of the hypoglossal canal is nearly dorso-ventral in the chondrocranium, while in the adult it passes almost laterally.
The lateral borders of the foramen magnum are formed by the lateral parts of the occipital or exoccipitals and the squamae. There is a general similarity in direction, in position, and in form of the exoccipital in the embryo and adult. Included in the lateral occipital and constituting much of it, as seen in this view, is the occipital neural arch, which is partially separated near its dorsal end from the exoccipital and squama by a fissure (the occipital fissure) which is filled with condensed mesenchyme.
That part of the exoccipital (the alar lamina) which continues upward from the jugular process is concave on its inner surface for the large transverse sinus. The sinus continues upward and outward, as in the adult, across the exoccipital and mastoid portion of the temporal, but turns forward over the dorsal margin of the otic capsule instead of backward over the squama of the occipital, as in the adult. At the bottom of the transverse sulcus the exoccipital joins the mastoid portion of the temporal. Along the upper part of this junction j'oung cartilage is present, but towards the jugular foramen the two are joined by precartilage and at the jugular foramen by dense mesenchjone or blastema, which in older embryos changes into cartilage. There is no line of demarcation between the exoccipital and the squama. The latter continues upward from the exoccipital as a broad, curved plate of cartilage.
Above the region of the exoccipital the occipital squama is continuous with the mastoid portion of the temporal. There is no line of demarcation between the squama and mastoid, and therefore much confusion exists in the literature regarding this region. This has come about through the use of the term parietal plate for the plate of cartilage lying above the outer edge of the otic capsule and partly continuous with it. We naturally associate the word parietal with the membrane bone of the same name, and the assumption has been that the parietal plate disappeared at a later stage and was replaced by the membrane bone. Its fate, however, has never been carefully traced, and it is probable that this so-called parietal plate, which extends backward into the nuchal plate, is part of the mastoid, for in the adult the mastoid is continued upward, around the posterior and even the dorsal edge of the petrous bone.
The foramen magnum is incomplete dorsally except for a thin membrane of connective tissue, the dorsal membrane. In later stages (as shown by Le\T[, Hertwig, and Macklin) the foramen is completed by a bridge of cartilage formed by the junction of the occipital squamae, which gradually extends around the brain. This cartilaginous bridge or band, usually called the tectum posterius or tectum syyioticum, in older stages extends around the brain from the lateral occipital and mastoid plate of one side to the lateral occipital and mastoid plate of the other, and corresponds to tlio nuchal j^hite of the adult occijMtal squama, which, as is well known, ossifies in cartilage. It seems quite unnecessary to introduce into human embryology a term appUed to a somewhat similar bridge of cartilage, arching over the brain from one auditory capsule to the other, in the cartilaginous skulls of amphibia and reptiles. The nuchal plate in the human embryo is not homologous to the tectum synoticum of the reptiles, since in man it is primarily continuous with the lateral parts of the occipital, and in reptiles with the auditory capsules only. It would be better to avoid entirely the term tectum posterius in human embryology, and to use the term nuchal plate, or squatna, as Levi has done, which at once gives to it its true significance in the higher vertebrates, in man at least.

The relations about the foramen magnum can be appreciated best by comparing figures 2 and 4. Figure 2 is drawn in the same position as the adult skull and the squama is intact. In its growth around the central nervous system it precedes that of the occipital neural arch and (as has been pointed out by Macklin) fuses with its fellow of the opposite side to form the primitive foramen before the occipital neural arches meet or nearly meet.
The large jugular foramen lies between the exoccipital and the otic capsule, as in the adult, extending laterally to the junction of the exoccipital with the mastoid cartilage, and medially to the junction of the exoccipital with the otic capsule. In the embryo and in the adult the same structures pass through the foramen, but both the nerves and the vein are proportionally much larger in the embryo. These structures have, however, approximately the same relations to each other; the transverse sinus occupies the posterior and lateral part, the vagus and the accessory nerves are just anterior and medial to it, and the glossopharyngeal is still more anterior and medial and lies in the notch on the posterior edge of the otic capsule.
The temporal cartilage, as in the adult, has an otic capsule or petrous part and a mastoid plate or mastoid part. The otic capsule forms one of the most conspicuous features of the cartilaginous skull, especially at this stage, for at this time the relatively small size of the occipital squama behind and of the orbital wing of the sphenoid and the ethmoid cartilage in front gives even greater prominence to the otic capsules than at a later stage when the squama and anterior end of the chondrocranium are fully developed in cartilage.
The otic capsule consists of a medial cochlear part fused with the lateral side of the anterior half of the basioccipital, and a lateral canalicular portion that forms part of the lateral wall of the chondrocranium. The otic capsule is broader in proportion to its length than the petrous bone. The internal acoustic meatus, the opening of the aqueductus vestibuli, and the fossa subarcuata are all relatively enormous at this stage. In the infant the fossa subarcuata also forms a relatively large depression.
The direction of the axis of the otic capsule or petrous portion of the temporal is very similar to that of the adult. The upper surface of the otic capsule, as alreadj' noted, contains the internal acoustic meatus, the opening of the aqueductus vestibuli, and the large fossa subarcuata under the superior semicircular canal. In the adult these openings are on the posterior surface of the petrous bone. If the adult skull is rotated so that the cranial surface of the basioccipital is horizontal, as in figure 1, of the embryonic chondrocranium, it will be seen that the posterior surface of the petrous bone is directed upward and corresponds in position to the upper surface of the otic capsule as seen in figure 1. In the embryo one leaf of the tentorium cerebelli is attached to the anterior margin of the otic capsule, and in the adult the tentorium is likewise attached to the corresponding margin of the petrous bone, so that the upper surface of the otic capsule faces the posterior cranial fossa, as does the corresponding surface in the adult. All of the mastoid plate (except its most anterior tip) and the squama of the occipital are posterior to the tentorium and face the posterior cranial fossa, as do the mastoid and nuchal plates of the adult.
Surrounding the caudal and dorsal edges of the canalicular part, and continuous with it, is the mastoid plate or cartilage. This cartilage forms part of the outer wall of the chondrocranium and is interrupted by a large mastoid foramen. We have already' noted the fusion of the mastoid plate with the exoccipital and occipital squama. It is also grooved for the transverse sinus, and the latter covers practically all of the inner surface of the cartilage. That portion of the mastoid cartilage above the outer edge of the otic capsule corresponds, therefore, more nearly to the upper part of the mastoid bone lying caudal to the upper part of the outer end of the petrous bone. This becomes the more apparent when we consider that the upper surface of the otic capsule, as seen in this aspect of the chondrocranium, presents both the internal acoustic meatus and the opening of the aque- ductus vestibuli, and thus corresponds to the posterior surface of the petrous bone and faces the posterior cranial fossa.
The basisphenoid, which continues forward from the basioccipital without line of demarcation, occupies about the same relative position as in the adult. The dorsum sellse, although well marked, is but imperfectly developed and consists mostly of precartilage and blastema, not indicated in figures 1 and 2 but shown in figure 3. To this blastema is attached the medial part of the tentorium. Between the dorsum sellse and the tip of the otic capsule the abducens nerve passes forward towards the orbit. In front of the dorsum sellse is the large shallow sella turcica. The hypophysial canal is seen in the center of the sella and contains remnants of the h\i)ophysial stalk. Connected with the basisphenoid are the temporal and orbital wings. The greater size of the orbital wings in the cartilaginous skull is exactly what we would expect when it is taken into consideration that the part of the adult temporal wing which is ossified in cartilage is smaller than the adult orbital wing ossified in cartilage.
The temporal wings project laterally from the sides of the basisphenoid, just in front of dorsum sellse and at a lower level than the floor of the sella turcica. These wings form a small part of the very incomplete floor of the- lateral parts of what will later be the middle cranial fossa. Each lateral part of the middle cranial fossa, although indicated here by the depressed area between the orbital wings and the otic capsules, is entirely filled by the lower end of the embryonic tentorium cerebelli. The boundaries of the middle cranial fossa are more clear in figure 3, where the lateral blastemal walls are shown. The temporal wing extends into this lateral blastema and thus completes the medial portion of the floor of the middle fossa. Between the temporal wing and the otic capsule is the large middle lacerated foramen, through wliich pass the internal carotid artery and the mandibular nerve. The facial nerve, with the geniculate ganglion and the greater superficial petrosal nerve, also passes through the foramen. The large otic ganglion lies across the foramen (figs. 12 and 13). The precartilaginous tissue attached to the anterior aspect of the temporal wing completes the foramen ovale and separates it from the supraorbital fissure. Between the temporal wing and the orbital wing, with its precartilage, is the large supraorbital fissure, and through it the oculomotor, trochlear, abducens, and ophthalmic nerves pass into the orbital cavity (fig. 15).
The embryonic tentorium cerebelh consists of two lateral prismatic masses of mesenchyme connected across the midline by a band of condensed mesenchyme that extends upward from the whole breadth of the dorsum sellae. The bases of the prisms are against the lateral walls of the membranous skull and the apical edges at the membranous band that extends upward from the dorsum sellae. The lower end of each prism fills one of the lateral depressed areas of the potential middle cranial fossa, between the otic capsule in back, the orbital wing in front, and the lateral wall of the membranous skull. The posterior wall of each prismatic mass of the tentorium is composed of a thin layer of condensed mesenchjone, and its lower edge is attached along the anterior edge of the otic capsule and extends upward. The anterior wall of each prismatic mass, hkewise composed of a thin layer of condensed mesenchyme, is attached to the posterior border of the orbital wing and its precartilage and extends upward. These two membranes come together along the medial apical edge where they are continued into the medial connecting band. Laterally, the two membranes fuse with the lateral membranous wall of the skull. The interior of each prism is filled with loose mesenchyme in which are imbedded the semilunar ganglion, large blood-vessels, and nerves. The trochlear and oculomotor nerves pass from above downward through the entire length of the prism. The tentorium obUterates entireh^ the potential middle cranial fossa. Later, when this fossa develops, the anterior wall of the tentorium must be pushed down and back into the floor and against the posterior wall of the fossa.
Each ala temporalis consists of two distinct parts, a medial alar process joined to the basisphenoid by young cartilage, and a lateral part which is, more strictly speaking, the cartilaginous temporal wing. It is attached to the under surface of the alar jprocess by condensed mesenchyme and lies at a lower level (figs. 1, 2, 3, 5, 6, 10, 11, and 14). Each alar process is usually regarded as forming the hngula, but since the internal carotid artery, which enters the cranial cavity between it and the apical end of the otic capsule, passes over its cranial surface, it seems prol^ablc that it gives rise as well to that part of the basisphenoid wliich forms the carotid sulcus. The relation of the greater superficial i)etr()sal nerve to the alar process also indicates that the latter is incorporated into the body of the sphenoid since, in the passage of the nerve from the geniculate ganglion to the spheno- palatine ganglion, part of its course is just beneath the alar process medial to the medial end of the temporal wing proper. It is here that the junction of the body with the temporal wing and the pterygoid process occurs and where later develops the pterygoid canal (fig. 13). Only the lateral part of the ala temporalis of the embryo, then, corresponds to that part of the temporal wing of the adult which ossifies in cartilage. In the adult the carotid sulcus and the lingula are at a higher level than the temporal wing, a condition which e.xists in the embryo if we consider the alar process as forming a part of the body of the sphenoid.
Since most of the temporal wing and both plates of the pterj-goid process are ossified in membrane, the cartilaginous wing, even when fully developed, represents but a small part of the temporal or greater wing and jjterygoid process of the adult. The maxillary nerve lies in front and the mandibular nerve behind this cartilage. In later stages the cartilage grows around the maxillary nerve and separates it from the supraorbital fissure, thus forming the foramen rotundum (figs. 3 and 10). Cartilage does not grow around the mandibular nerve, and the foramen ovale is said to be formed by membrane bone. At this stage, therefore, the large cleft between the temporal wing and the orbital wing represents more than the supraorbital fissure.
The alar process and the temporal wing proper each has its own center of chondrification, as pointed out by Bardeen and Fawcett. The alar process chondrification unites with the basisphenoid before the temporal center unites with the alar process.
The cartilaginous orbital wing, Uke the temi;)oral wing, is very incomjjletely developed, as will be seen by comparing it with the older stages of Levi, MIacklin, and Hertwig. It consists of two parts, a proximal or basal part, and a lateral, sickle-shaped part, which springs upward, outward, and forward around the optic nerve and has much the same general position as that part of the orbital wing immediately about the optic nerve of the adult. The basal part of the orbital wing is connected to the basisphenoid by young cartilage and to the lateral part by cartilage not quite so far advanced as that in either the basal part or the lateral part. It seems probable that both parts of the orbital wing may arise from independent centers of chondrification. From the general position of this basal part and its relation to the optic nerve and to the general mesenchyme of this region, and from the fact that it gives origin to all of the muscles of the orbit except the superior oblique, I think it must ultimately become incorporated into the body of the sphenoid. The sickle-shaped lateral part of the orbital wing is intimately related to a larger precartilaginous part indicated by the green-colored structure in figure 3. This precartilaginous part shades off into the frontal blastema and there is in reality no sharp line between the two. The cartilaginous and precartilaginous parts together have somewhat the same form as the orbital wing of the 40 mm. embryo described by Macklin.
All the cartilaginous skull the optic foramen is incomplete but, as shown in figure 3, is completed by the precartilage extending from the tip of the orbital wing to the anterior part of the body of the sphenoid.
In figure 1 it can be seen how the anterior end of the basisphenoid rapidly narrows and continues into the mesethmoid. Both in figures 1 and 2 the dorsal thin edge of the mesethmoid can be seen; it corresponds in the adult to that part of the mesethmoid which bisects the posterior part of the cribriform plate. The mesethmoid extends into the large crista galli, which consists mostly of precartilage. In figure 3 the position of the future cribriform plate is indicated by the large olfactory foramina, one on either side of the short dorsal edge of the mesethmoid. On either side of the mesethmoid, and connected with the anterior edge in front of the crista galli, are the lateral wings of the nasal capsules. As seen in figures 1 and 2, the posterior edge of each capsule is presented.
The anterior cranial fossa is not evident in the cartilaginous skull. In the combined cartilaginous and membranous skull shown in figure 3 its limits are clearly indicated. The floor of the fossa is formed by the precartilage of the orbital wing, frontal blastema, and dorsal membrane, and in the center are the mesethmoid and crista galli and the olfactory and ethmoid foramina. Figure 3 shows not only the floor but also a considerable extent of the anterior wall formed by the dorsal membrane.
Median Sagittal Aspect

In order to compare the view from the median sagittal plane shown in figure 5 with a similar view of the adult skull, the latter should be rotated so that the cranial surface of the basioccipital is horizontal. In the median sagittal section of the adult skull before me the pharyngeal edge of the basioccipital measures 26 mm., the basisphenoid 24 mm., and the upper edge of the vomer (from the basis-phenoid to the anterior nasal spine of the maxilla) 54 mm. The latter distance corresponds to the ventral edge of the mesethmoid. In the cartilaginous skull the corresponding distances are 2 mm., 1.6 mm., and 1.8 mm. The ratios, then, are respectively 1:13, 1:15 and 1:30. The basisphenoid is thus proportionally shorter in the embryo at this stage than the basioccipital, and the mesethmoid much more so. This is what one would expect to find in the less differentiated regions at the anterior end of the skull.
The angles which the pharyngeal surface of the basioccipital make with the same surface of the basisphenoid, and the latter with the mesethmoid, arc practically identical with those found in the adult. Since the edge of the basisphenoid is concave, a straight line connecting the two ends was used. It is interesting to note in this connection that the angle made by the cranial surface of the basi-occipital, projected sagittally on to a line parallel with the upper edge of the zygomatic arch, is almost exactly the same in this embryo as in the adult. Such measurements serve to indicate that the cartilaginous skull is laid down from the very beginning on much the same plan as that part of the adult skull ossified in cartilage.
The basioccipital shows in its median sagittal section two parts — a thin caudal part hang beneath the notochord connecting the posterior part of the two lateral thickened masses of the body from which the exoccipitals arise, and a long wedge-shaped part in front of and above the notochord; this Ls thickest where it joins the basisphenoid. The diagonal course of the notochord through the basioccipital and its reentrance into the basal plate at or near the junction of the basioccipital with the basiphenoid is also clearly shown. The three roots of the occipital neural arch arise from the posterior half of the basioccipital and unite lateral to the hypoglossal foramina into a tapering, rod-like cartilage. The serial relationship of the occipital neural arch with the cervical neural arches is very apparent in figure 5. The dorsal tip is in line with the dorsal tips of the cervical neural arches. The occipital squama is very prominent in this view; it appears to continue upward from both the exoccipital and the otic capsule, and even to project in front of the otic capsule. There is no line of demarcation between it and the mastoid plate which encircles the posterior and upper edges of the canalicular part of the otic capsule and thus no definite limits can be given to either cartilaginous area. The anterior part of this plate is usually called the parietal plate, a term which we have already discussed and discarded, since it is not apparent that it includes anything more than what might be embodied under the term mastoid plate. In considering this region it should be borne in mind that figure 5 is drawn with the basioccipital horizontal. The large mastoid foramen (capsuloparietal fissure) lies at the bottom of the transverse sulcus and interrupts the continuity of the mastoid plate. Above the region of the otic capsule the masto- squamal plate is grooved for the endoljonphatic sac. Whether this groove lies on the mastoid or squamal part is uncertain. The fate of the various regions of the masto-squamal plate will remain obscure until we have a more complete series of stages.
The surface of the otic capsule presented in this view corresponds to the posterior surface of the petrous bone and faces the posterior cranial fossa. Its diagonal position also corresponds to the adult when the latter is seen from the same angle. The cochlear part is mostly above the level of the basioccipital, projecting but slightly below the level of the latter. The canalicular part shows the position of part of the superior semicircular canal in front and of the posterior canal in back. The endolymphatic sulcus serves to mark the position of the common duct.
The medial section of the basisphenoid extends only to the hypophysial canal and a diagonal cut in front of this cuts off the orbital wing and extends to the edge of the cartilage. It is impossible to determine the anterior and posterior limits of the basisphenoid. The posterior limit is probably at or near the notochord and the anterior limit somewhere near the line (*). The body is biconcave in the region of the sella turcica. Anteriorly, it narrows from side to side, as shown in figure 1, and increases in its sagittal thickness, as shown in figure 5. The temporal wing projects below the body and the orbital wdng above. The mesethmoid is shown intact after removal of the nasal capsule. The large crista galli and anterior end consist largely of young cartilage and precartilage.
Inferior Aspect
In comparing the inferior surface of the chondrocranium with that of the adult, only the occipital and otic regions lend themselves readily to our purpose, unless we imagine as eliminated the membranous bones, the maxillse, the palatines, the vomer, the ptorygoid process, most of the great wing of the sphenoid, and the tympanic part of the temporal. Figure (i is drawn with the pharyngeal surface of the basioccipital parallel to the plane of the paper. This is not the usual view of the adult skull, and the latter must be rotated into the corresponding plane for comparison.
The basioccipital and the basisphenoid present no new features of especial interest not already considered, except the openings of the canal for the notochord. The anterior opening where the notochord reenters the basal plate is probably near or just back of the junction of the basioccipital and basisphenoid. The exoccipital shows the large hypoglossal foramina; these are not covered by the large condj'les, as is the case in the adult. The condyles have scarcely begun to develop, and the occipital is united to the atlas by dense mesenchyme without articular surface. The tip of the occipital neural arch is quite distinct and separated from the squama. The prominent transverse or jugular process lies just caudal to the jugular foramen and, as in the adult, extends slightly lateral to the outer border of the foramen. The process is more prominent than in the adult and has more the character of the vertebral transverse processes.

The immature condition of the inferior surface of the otic capsule and mastoid region is accentuated by the fact that such structures as the carotid artery and the facial nerve are not inclosed by cartilage, but lie partly in sulci beneath the capsule. The internal carotid artery passes forward and medially in the mesenchyme beneath the cochlear part, with only the beginning of a groove formed by precartilage, not indicated in this figure. It enters the cranial cavity between the apex of the cochlear part and the alar process. The facial nerve, which enters the otic capsule through the internal acoustic meatus, soon emerges again from the facial foramen located in the anterior region of the capsular sulcus between the cochlear and canalicular parts. The geniculate ganglion lies just outside this foramen (figs. 12 and 13). The greater part of the facial nerve, which in the adult is inclosed in bone, is extracapsular and passes backward and outward in a groove on the under surface of the canalicular part. The groove lies ])etween the fossa incudis and the fenestra vestibuli or ovalis, and medial to the root of the styloid. The cartilaginous auditory ossicles lie beneath the canalicular part (fig. 14).
The serial relationship of the jugular process, the mastoid process, the upper end of the styloid, and the incus is clearly indicated in figure G, in which the jugular, mastoid, root of the styloid, and fossa incudis are .shown. It is impossible to determine at present whether this has any phylogenetic significance or not. In some of the embryos of al)out this stage there is a small separate cartilage in the blastema of the mastoid process to which are attached the digastric and stapedius muscles. The serial relationship shown in the embryo is quite different from that of the adult, where the mastoid process which gives attachment to the digastric is some little distance lateral to the jugular process, and the styloid process is immediately in front of the latter. Just in front of the jugular foramen is the opening for the aquaeductus cochlese.
The sphenoid shows the basisphenoid with the opening of the hypophysial canal near its center and the two wings on each side. Part of the orbital wing is obscured by the lateral part of the nasal capsule, as it would be in the adult from this point of view.
The line of fusion of the nasal capsule with the mesethmoid is shown, and the anterior nares directly face the observer as they do in the adult. The junction of the nasal capsules with the mesethmoid is effected along the middle third of the anterior edge of the mesethmoid by precartilage. The cut edge of this junction is shown in figure 5.
Lateral Aspect


The cartilaginous skull covers but a small area of the lateral surface of the brain, namely, part of the medulla, part of the cerebellum, and a small area in the region of the optic nerve (figs. 7 and 8). The cartilaginous skull, even in its completed form, is a very inadequate protector for the brain, never covering more than a small fraction of its surface. The condensed mesenchyme or blastema covers a much larger surface, but even the cartilage and blastema together form at this stage a very incomplete brain capsule (fig. 9). The capsule is completed by the thin dorsal membrane. The inclosing of the central nervous system by the gradual spreading of the blastema and cartilage, which invade and replace the dorsal membrane, is similar to the well-known development of the thoracic and abdominal walls and the disappearance of the ventral membrane.
The blastema covers almost all of the lateral surface of the cartilaginous skull. A small part of the occipital cartilage, including the transverse process, part of the squama and occipital neural arch, part of the orbital wing of the sphenoid, and part of the lateral surface of the nasal capsule, are uncovered (figs. 9 and 15). Into the blastema covering the squamal cartilage, rather than into the cartilage itself, are inserted the various occipital muscles (figs. 14 and 15). The blastema covering the squama and the lateral surface of the otic capsule probably fases later with the perichondrium, but at this stage it seems to be continuous with the rest of the blastemal wall which later gives rise to membrane bones. It is in the sphenoidal and frontal regions that the blastema greatly predominates over the cartilage. All of the lateral wall of the middle cranial fossa consists of blastema and the greater part of the floor (as well as all of the lateral wall of the anterior fossa) is likewise formed by blastema. The orbital walls are mostly of blastema; cartilage of the orbital wing of the sphenoid takes part in the formation of the apical region of the orbit about the optic foramen, and a portion of the medial wall of the orbit is formed by part of the lateral wall of the nasal capsule. Connected with the cartilage is considerable precartilage extending along the inner wall towards the apex (fig. 15). Part of the outer wall of the orbit, consisting of the zygomatic blastema and the zygomatic process of the frontal blastema, and also the zygomatic arch, are shown in figure 9. In lateral views of the adult skull the upper border of the zygomatic arch is horizontal, and it is therefore easier to compare figure 9 and the other lateral views with the adult if the figure.s are rotated so as to bring the zygomatic arch into a horizontal position. Attention has already been called to the fact that the angle made by projecting the line of the upper edge of the zygomatic arch on to the basioccipital is almost exactly the same in the embryo and in the adult.
By rotating figure 7 to correspond to the usual adult position it will be seen that the occipital squama projects backward from the exoccipital and mastoid, and that, since it covers only part of the medulla and cerebellum, it is concerned with the wall of the posterior cranial fossa. It is from this lateral view that one gets the impression that the squama extends into the jugular process, or more strictly, into that part of the exoccipital which continues into the lateral part of the jugular process, rather than into the occipital neural arch wliich lies in a deeper plane. The occipital neural arch has attained only a small proportion of the growth necessary to inclose the medulla. The tip of the squama, on the other hand, has grown farther around. The condition here is not as far advanced as that found by Levi in a supposedly younger embryo.
The lateral aspect of the jugular process is quite prominent, and to it are attached two muscles, the rectus capitis lateralis (serially related to the intertransversarii) and the occipitomastoid muscle (fig. 14). The former needs no special comment. The latter muscle is not found in the adult and is infrequent in embryos. Its presence would seem to indicate either that the mastoid process was separate or movable, or that the temporal and occipital cartilages were at one time movable on each other. We do find that the occipital and mastoid cartilages are separated by young cartilage or precartilage in the region below the mastoid foramen. Above this, however, no fine of separation can be found, but this does not necessarily mean that the two cartilages were articulated by a movable joint. The significance of the mastoid process is described elsewhere.
The canalicular part of the otic capsule forms a conspicuous part of the lateral wall of the chondrocranium. A slight bulging on the lower part of its lateral surface indicates the position of the lateral semicircular canal. The styloid process and incus he beneath the lateral edge of the otic capsule almost flush with the lateral surface.
Between the canalicular part and the occipital cartilage is the mastoid cartilage. It is fairly clearly defined in the region between the otic capsule and the exoccipital below the mastoid foramen. In the region above the otic capsule there is no line of demarcation between the squama and mastoid. The mastoid process projects from the lower edge of the mastoid cartilage. In this embryo it consists of blastema; in other embryos, as will be stated farther on, a small cartilaginous nodule is sometimes embedded in the blastema. To the mastoid process are attached several muscles, namely, the occipitomastoid, the atlantomastoid, the digastric, the stapedius, and the longissimus capitis (figs. fig. 14 and 15).
The anterior part of the cartilaginous skull, the sphenoid and the ethmoid, bears very little resemblance to anything ordinarily seen in the lateral view of the adult skull, partly because most of the bone ossified in cartilage becomes covered or shut out from the lateral view by membrane bone. If we can imagine all of this membrane bone stripped away, the resemblance between the two would be clearly seen.


The lateral surface of the body of the sphenoid, with the shallow sella turcica lodging the hypophysis, is to be seen in front of the apex of the cochlear part of the otic capsule. The prominent dorsum sella projects towards the great mid-brain fissure, and from it (but not shown) the thin medial membranous part of the tentorium projects into this fissure. The temporal wing lies for the most part below the level of the basisphenoid and is attached to the under surface of the alar process, while the orbital wing lies, for the most part, above the level of the basis-phenoid and is curved around the optic nerve. The two parts of the orbital wing are clearly indicated in figures 7 and 14. In the description of the middle cranial fossa and the embryonic tentorium cerebelli it was noted that the lateral prismatic mass was so placed that the apical edge extended upward from the dorsum sellse in connection with the thin medial membranous part of the tentorium lying in the great mid-brain fissure. The lower end of the embryonic tentorium, as previously noted, occupies the space between the otic capsule and the orbital wing of the sphenoid, the semilunar ganglion filling up most of the gap (fig. 8). It lies outside of the cartilaginous skull but within the membranous skull. The geniculate ganglion and adjacent sections of the facial nerve are not incased in cartilage, and they also he in this middle cranial fossa or gap (figs. fig. 12 and fig. 13).
In the lateral aspect of the chondrocranium most of the mesethmoid is shut out from view by the nasal capsule. Between the orbital wing and the nasal capsule the anterior end of the basisphenoid and a bit of the mesethmoid appear. If only the cartilaginous skull were considered, this region of the basisphenoid might be regarded as interorbital and the term inter orhital septum could be apphed. But this so-called interorbital septum exists only in the cartilaginous skull; in the more complete skull, with all the blastema and precartilage, the medial wall of the orbit is complete enough to shut it out from the orbit. The term inter orhital septum probably has no significance whatever in human embryology. The prominent crista galli projects between the cerebral hemispheres. The lateral surface of the nasal capsule forms a large proportion of the inner wall of the orbit and probably ossifies into the lamina papyracea of the ethmoid.
The Occipital Cartilage
The occipital cartilage consists of an elongated, flattened body (or basioccipital), the lateral parts (or exoccipitals), and the two nuchal plates (or squamae).
The body forms the greater part of the so-called basal plate, a term much used in descriptions of the cartilaginous skull. It continues without line of demarcation into the basisphenoid, and one can only approximate the future hne of junction between the two bones which ossify in the cartilage by the relation to the apex of the otic capsule and the entrance of the notochord. The basioccipital lies in a plane almost exactly horizontal to the long axis of the embryo. This elongated quadrilateral plate is thinner at the center, slightly concave on the cranial surface, and slightly convex on the pharygeal surface. It becomes thicker where it joins the sphenoid. The rounded caudal end is bent downward and notched in the midline at the foramen magnum. The caudal part of the basioccipital gives much more of a clue to its vertebral origin than the flattened cranial part. The caudal part is thickened and on the cranial surface is incompletely divided into two lateral masses by a deep median groove. These correspond, perhaps, to the bilateral masses which fuse to form the bodies of the cervical vertebrae. Levi finds in his 13 mm. embryo two bilateral cartilaginous centers medial to the hypoglossal roots. They probably represent, as Macklin suggests, the bilateral centers for the occipital vertebrse. Their position in the 13 mm. embryo corresponds to the position of the lateral masses in this 21 mm. embryo. These masses are united across the midline beneath the notochord, and anteriorly are continued into the broad flat plate of the basioccipital. The center of each mass contains the most highly differentiated cartilage of the entire chondrocranium.
In the 14 mm. embryo described by Levi the otic capsule and the basioccipital are apparently separated by a wide gap. Levi probably did not include all the precartilaginous tissue and blastema of the otic capsule, since in embryo No. 109 (Carnegie Collection), 11.5 mm. in length, the blastema of the otic capsule is in contact with the basioccipital. It is only in younger stages, 9 mm. length, that I find the blastema of the otic capsule separated from the basioccipital. The union of the blastema, then, precedes that of the cartilage in this region. In embryo No. 460 the anterior half of the lateral border of the basioccipital is fused with the cochlear part of the otic capsule along a crescentic line. The line of fusion can still be recognized by the differences in the degree of differentiation of the two cartilages. The cochlear part consists of young cartilage and the nuclei are closer together than in the basioccipital. The border of the basioccipital is, however, not so far advanced as the central part. The two cartilages were so completely fused in the 17 mm. embryo described by Levi that he was unable to recognize any histological border between them. Embryo No. 128 (Carnegie Collection), 20 mm. in length, although supposedly smaller and younger than the 21 mm. embryo (No. 4G0), shows a more advanced conchtion in the cartilaginous differentiation of this region, and it is impossible to find a histological border between the basioccipital and the cochlear part except at either end of the line of fusion. In another 20 mm. embryo (No. 22, Carnegie Collection) there is only the very slightest indication of the line of fusion . There is probably some variation in the rate of differentiation of the cartilages of this region in embryos of the same age and size.
Occipital Vertebra
Each exoccipital corresponds for the most part to a vertebral neural arch or hemiarch, which is so distinct in this embryo that we may properly apply the term occipital hemiarch. The roots, laminae, and transverse processes can be readily recognized. The roots are broad and thick and correspond to the roots or pedicles of the vertebral neural arches. On the right side, as already noted, there are three roots dividing the cranial end of the hypoglossal canal into two parts, a condition not uncommon in the adult. The caudal root on each side is the largest and consists of more advanced cartilage, and it is questionable whether the anterior roots should be looked upon as forming part of the occipital vertebra. It has often been suggested that they are part of an anterior, still more rudimentary vertebra. The laminae extend outward and upward from the roots, forming part of the lateral border of the foramen magnum. They are much thickened, rounded, tapering cartilages, quite distinct, and in the dorsal part entirely separate in this particular embryo from the squamal cartilage. The cartilage of the occipital arch is more differentiated than the squamal. During the further course of development, as Macklin has pointed out, the occipital neural hemiarches grow around the central nervous system and approach each other in the midline. It is not clear whether they actually meet and fuse, or whether a small part of the squamal cartilage may not intervene. The squamal cartilages, which likewise grow around the central nervous system, meet and fuse before the occipital hemiarches. The ossicle of Kerckring, which develops in this region, may represent a separate ossification center of the nuchal plate intervening between the tips of the occipital neural hemiarches, or this center may possibly be looked upon as the ossification of the fused occipital spinal epiphyses. Schultz described a skull with, two bilateral ossicles in this region, and suggests that they may correspond to the epiphyses of the spinous processes.

In this embryo the separation of the dorsal part of the lamina of the occipital vertebra from the squama of the occipital, and the more intimate relation of the latter with the transverse process, would seem to indicate that the squama is an extension upward from the transverse process rather than an outgrowth from the occipital neural lamina. The relation of the squama to the transverse process and to the occipital neural lamina is clearly shown in figures 1, 7, and 16. The occipital transverse process forms part of the caudal and lateral margins of the jugular foramen and continues up into the squama and alar lamina without line of demarcation. The alar lamina, as will be seen in figure 16, constitutes the ventral and more medial part of the squama adjoining the lamina of the occipital and the medial part of the transverse process. There is also a more gradual transition as regards the degree of differentiation from the cartilage of the transverse process into the squama than from that of the lamina.
In embryo No. 22 (Carnegie Collection), 20 mm. in length, the squama of the occipital is fused wath the lamina throughout the entire length of the latter on both sides of the embryo, although throughout most of this length the younger cartilage of the squama can easily be distinguished from the thickened, older cartilage of the lamina. Towards the apex of the arch, however, the two cartilages are of the same degree of differentiation and no distinction between them can be seen.
In embryo No. 128 (Carnegie Collection), 20 mm. in length, the resemblance which the occipital neural arch and its transverse process bear to those in the cervical region is very striking. The cartilages of the occipital and cervical neural arches show much more advanced differentiation than the thin transverse processes. The muscle intertransversarius, between the transverse processes of the second and the first cervical vertebra?, is in line with and serially related to the rectus capitis laterahs, which passes between the transverse process of the first cervical vertebra and that of the occipital. In this embryo (No. 128) it can be seen also how the squama of the occipital is a continuation upward from the transverse process rather than from the lamina. The laminae of the occipital vertebra are also fused along the entire length to the squamae, but are readily distinguishable from the latter by their much more advanced cartilaginous differentiation. The tips of these laminae are connected with the tips of the laminae of the atlas by the interspinous ligament, as in embryo No. 460.
In embryo No. 240 (Carnegie Collection), 20 mm. in length, the cartilage of the occipital neural arch on each side is likewise fused with the squama throughout its entire length. The lamina also shows more advanced differentiation than the squama. In embryo No. 431 (Carnegie Collection), 19 mm. in length, the tips of the occipital neural arches are separated from the squama on either side by the condensed mesenchyme forming the perichondrium. The e.xtent of the fusion of the laminae and the squamae is, however, more extensive than in embryo No. 460. These observations indicate that the occipital neural arch is more marked in this stage than in the adult, and that probably an extreme degree of accentuation in the embryo would precede any marked manifestation of an occipital vertebra in the adult. The manifestation of an occipital vertebra in adult skulls has been described by Gladstone, Kollman, and others. Such variations remind one of the conditions found in embryos at this stage, and we shall probably find that embryos vary as much, if not more, than adults and that such embryonic variations always precede the adult variations.
The relation and attachment of the dorsal membrane to the tips of the occipital hemiarches are similar to its relation and attachment to the tips of the neural arches of the cervical vertebrae, in that in both cases the dorsal membrane is continuous with the perichondrium on the medial side of the tip of the neural arch. Its attachment to the upper border of the squama is somewhat different, since here it is merely continuous with the thin edge. The tip of the occipital hemiarch is connected with the tips of the vertebral hemiarches by a distinct band of condensed membrane, the interspinous ligament described by Bardeen (fig. 9).
The transverse or jugular process springs from the occipital hemiarch at the junction of the roots and lamina and projects laterally back of the jugular foramen. We have already noted its serial rclationsliip with the vertebral transverse processes. The lateral extremity of the jugular process has a knob-like enlargement, and into this are inserted the rectus capitis lateralis muscle and the occipito-mastoid muscle (fig. 24).
The occipital squama or nuchal plate springs from the upper border of the lateral part of the jugular process. Near its origin it is narrow in the sagittal plane and wide m the frontal plane. It is continuous with the basal part of the lamina by a curved plate, the alar lamina (figs. 1 and 16), and rapidly widens into a broad, thin plate, which continues upward to a thin edge that runs into the dorsal membrane. Both the jugular process and the squama extend into the mastoid cartilage. At the jugular foramen these two cartilages are separated by blastema and precartilage; above this they are joined by young cartilage, which gradually changes into cartilage indistinguishable from that of the squama or mastoid. The lateral surface of the squama is covered by condensed mesenchyme The into which the edges of the cartilage merge. It is into this mesenchyme that the occipital muscles— namely, the trapezius, splenius capitis, semispinahs, rectus capitis posticus major and minor, and the obliquus capitis superior, appear to be inserted (figs. fig. 14 and 15).
The Sphenoid Cartilage


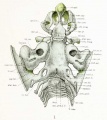
Fig. 1. Dorsal aspect of base of the cartilaginous skull with the basioccipital in the horizontal plane.
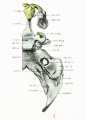
Fig. 2. Right half of dorsal aspect of the base of the cartilaginous skull.
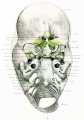
Fig. 3. Dorsal aspect of cartilaginous and membranous skull.
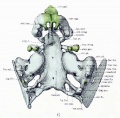
Fig. 6. Ventral aspect of base of the cartilaginous skull.

The sphenoid cartilage consists of a body with two lateral wings or processes on each side, the temporal and orbital wings (figs. 1, 2, 3, and 6). The cartilage of the body is continuous with the body of the occipital, and in front narrows and thickens as it passes into the nasal septum. The cartilaginous center from which the body develops is described as being caudal to the hypophysial canal. The chondrification spreads caudally to meet the basioccipital, upward into the dorsum sellae, and forward around the hypophysial canal where the cartilage fuses around the canal and extends forward to form the anterior part of the sphenoid and the nasal septum. Fawcett, after an examination of a 21 and a 19 mm. embryo, concluded that the dorsum sellae was an independent formation, since he found in each embryo a separate cartilaginous bar above the region of the body and separated from it by mesenchj^ne. Twenty embryos of the Carnegie Collection, ranging in length from 15.5 mm. to 24 mm., were examined, and such a cartilaginous bar was found in but one (No. 229), 19 mm. long. Twelve of the embryos were between 18 and 23 mm. Great variation in the form of the dorsum sellse was observed, but the rather rare presence of a separate center of chondrification would indicate that as a general rule the dorsum sellse develops from the basisphenoid, as described by Levi. In front of the dorsum sellse the body is biconcave and perforated near the center by the hypophysial canal. There is practically no indication in front of the sella turcica of the tuberculum sellse. The anterior part of the body is very incompletely developed, and not until the anterior tip of the orbital wing has grown medialward to meet its fellow of the opposite side and fuses with, the body is the anterior border of the chiasmatic groove apparent (fig. 3).
The temporal wing consists of two parts : the alar process, which springs from the body just in front of the level of the dorsum sellse and projects laterally and sKghtly caudally toward the apex of the cochlear part of the otic capsule, and the lateral part, attached to the under surface of the alar process and projecting laterally. Each part is supposed to have an independent center of chondrification. In this embryo the alar process is joined to the basisphenoid by j'oung cartilage and to the lateral part by blastema. We have already given sufficient consideration to the fate of the alar process and its probable incorporation into the body of the sphenoid, and also the lateral part which corresponds more strictly to that part of the temporal wing that ossifies in cartilage. This part of the cartilage is still incompletely developed; as in later stages the cartilage extends around the maxillary nerve. The orbital wing is even more immature than the temporal, as will be seen by comparing it with the older stages of Macklin and Hertwig. In figure 3 is shown the precartilaginous part, which is more extensive than the cartilaginous part. The basal part of the orbital wing lies at the apex of the orbit and gives attachment to all the muscles of the orbit except the superior oblique (figs. 10 and 15).
The Temporal Cartilage
This cartilage includes the otic capsule and the mastoid cartilage, which are intimately fused together.
The Otic Capsule
The otic capsules form the cartilaginous basis for the petrous bones of the adult. They are very prominent, being striking features of the chondrocranium, and extend on each side in a caudo-dorso-lateral direction to the lateral surfaces of the skull. Each capsule consists of two broadly united and continuous parts (a medial cochlear part and a lateral canalicular part) inclosing the cochlear and semicircular canals respectively. The two parts are more or less set off from each other by a broad shallow groove, the capsular sulcus.
The cochlear part is somewhat egg-shaped and fuses with the basioccipital. The cranial surface is rounded and presents the large, rounded, internal acoustic meatus. The ventral surface is also rounded and projects below the level of the basioccipital, forming with it a distinct groove. The cochlear part consists of somewhat younger cartilage than the canalicular part. Its relation to the cochlear duct is shown in figure 16.
The capsular sulcus extends entirely around the otic capsule. On its anterior surface is a largo foramen for the exit of the facial nerve. This foramen is separated by a narrow bar of cartilage from the internal auditory meatus. The facial nerve has but a very short course within the otic capsule; the geniculate ganglion and the nerve distal to it are outside of the capsule, but close against it (figs. 12 and 13). The capsular sulcus is broad and shallow on the inferior surface and near its center lies the fossa vestibularis or ovalis, in which is imbedded the base of the stapes. Immediately about the fossa is an area of young cartilage; the floor of the fossa is also covered with young cartilage. The posterior part of the sulcus, which borders the jugular foramen, contains the opening for the aqueductus cochloaris. At this stage only a small plexus of veins pass through it, but later it contains the perilymphatic duct.
The canalicular part of the otic capsule constitutes the lateral half and forms part of the outer wall of the chondrocranium. It is more or less oval in form and flattened in a mcdio-latoral direction. It consists for the most part of the thickened cartilaginous covering for the semicircular ducts. The locations of the latter are more or less apparent by the configuration of the cartilage. The position of the superior semicircular duct is quite clearly indicated; it lies within the prominent, rounded semicircular mass constituting the antero-dorsal portion of the canalicular half of the capsule (figs. 1, 2, 3, 5 and 15). The deep fossa subarcuata lies at the center of its curve. The lateral semicircular canal produces a distinct bulge on the lower part of the lateral surface of the capsule, while the position of the posterior duct is indicated by the configuration of the posterior portion of the canalicular part. The cranial surface shows a well marked sulcus for the endolymphatic duct which leads into the opening for the aqueductus vestibuU. The upper edge of the cartilage, where it joins the mastoid, is deeply grooved for the transverse sinus. The lower edge of this posterior portion projects ventrally in the form of a ridge, the crista parotica, to the caudal end of which is attached the styloid process. In front of the crista there is a slight depression, the fossa incudis, in which is lodged the short crus of the incus. The facial nerve lies in a groove just medial to the crista and styloid process (figs. 6, 12, and 13).
Mastoid Cartilage
The mastoid cartilage is continuous with the caudal and upper border of the canalicular part. We have already described its fusion with the exoccipital and squama of the occipital. Its inner surface is hollowed out for the great transverse sinus, and in the upper part of this sulcus is the large mastoid foramen. A distinct mass of blastema or precartilage projects ventrally from the inferior edge of the mastoid just back of the root of the styloid process. Since to this mass are attached the digastric, splenius capitis, longissimus capitis, and sternomastoid muscles, it probably represents the mastoid process. In addition to the above-mentioned muscles the stapedius also appears to arise from the inner side of the mass. At this stage the stapedius is relatively large and entirely extracapsular. Two interesting rudimentary muscles, not found in the adult, are likewise attached to this mastoid process. One, short and thick, arises from the occipital transverse process; the other, longer and more slender, extends from the transverse process of the atlas (figs. 14 and 15). I have named these muscles the occipito-masloid and the atlanto-mastoid. The occipito-mastoid is serially related to the intertransversarii.
This mastoid blastema, or mastoid-process blastema, presents interesting variations. In embryo No. 382 (Carnegie Collection), 20 mm. in length, there is a small nodule of cartilage in this region — the mastoid-process cartilage. This is attached to the occipital transverse process by a short ligament. Both the digastric and stapedius muscles are attached to it, but no trace of the occipito-mastoid or atlanto-mastoid muscles were found. In embryo No. 431 (Carnegie Collection), 19 mm. in length, a similar nodule of young cartilage was found attached in the same manner to the transverse process of the occipital by a ligament. The stapedius muscle is attached to its inner surface and the digastric to its lower surface. In both embryos the mastoid-process cartilage is closely associated with the mastoid blastema and the ligament or band of condensed mesenchyme, connecting it wdth the jugular process, occupies the same position as the occipito-mastoid muscle found in embryo No. 460. The attachment of the stapedius and digastric suggests that the mastoid process may be regarded as a remnant of the proximal end of a branchial cartilage, while its serial relation with the transverse process of the occipital and cervical vertebrae, and its connection with the former by muscle, as in embryo No. 460, or by ligament, as in embryos Nos. 431 and 382, suggest that it may be a rudimentary portion of the transverse process of an occipital or temporal vertebra. A still more speculative idea would be to consider the branchial bars as serially related to the vertebral transverse process, and the occipito-mastoid muscle could be looked upon as serially related to the intertransversarii muscles on the one hand and to the stapedius on the other.
The Ethmoid Cartilage

The ethmoid cartilage consists of the mesethmoid and the lateral nasal capsules joined to the former by precartilage. The ethmoid is very poorly developed at this stage and all the figures include more or less precartilage. The gradual transition from cartilage to young cartilage and precartilage makes it impossible to draw sharp lines between the cartilage and precartilage. The mesethmoid consists of fairly well-differentiated cartilage near its junction with the sphenoid. In this region the pharyngeal edge is much thicker than the dorsal edge. It gradually tapers towards the apex, the entire septum attaining an even thickness. The crista galli is relatively large when both cartilage and precartilage are included, as shown in the various figures.
The nasal capsules consist mostly of young cartilage and precartilage. They are relatively simple and show only slight indications of the turbinate processes. Part of the lateral surface enters into the formation of the medial wall of the orbit and part is covered by maxillary and nasal blastema, the precartilaginous tips extending beyond (fig. 15). The inner wall of the orbit is completed by frontal blastema and precartilage of the orbital wing of the sphenoid. This blastema and precartilage, as already noted, form a continuous sheet (fig. 3). The orbital surface of the sheet is more or less closely united to the upper edge of the nasal capsule (fig. 15) . On the line of junction between the cartilage and frontal blastema are found, as in the adult, the anterior and posterior ethmoidal foramina, through which pass the nasociliary nerve and the ethmoidal branches.
Bibliography
Bakdeen, C. R., 1910. MorphogcncsLs of the skeletal system. Keibel and Mall, Manual of Human Embryology.
Fawcett, 1910. Notes on the development of the human sphenoid. Jour. Anat. :iii<l Phys., vol. ) 1, p. 207.
Fisher, E., 1903. Zur EntwicklunK.sne.seliii-hte des Affen.sehiidels. Zeit.f.Morph.u.Anthropol.,Bd.5.
Gladstone, 1915. Manife.station of occipital vertebra; and fusion of the atlas with the occipital. Jour. Anat. and Phj\s., vol. 49, p. 190.
Jacoby, M., 1895. Ein Beitrag zur Kcnntnis des menschlichen Primordialcraniums. Arch. f. mikr. Anat., Bd. 44.
KoLLMAN, J., 1905. Varianten am Os occipitale, besonders in der Umbegung des Foramen occipitale magnum. Anat. Anz., Bd. 27. Verb. d. anat. Gesellsch., p. 231-2.36.
Levi, G., 1900. Beitrag Zum Studium der Entwieklung des knorpeUgen Primordialcraniums des Menschen. Arch. f. mikr. Anat. u. Entwick, Bd. .55.
Lewis, VV. II., 1915. The use of guide planes and plaster of paris for reconstructions from serial sections, etc. Anat. Rec, vol. 9.
Macklin, C. C, 1914. The skull of a human fetus of 40 mm. Am. Jour. Anat., vol. 16.
Mead, C. S., 1909. The chondrocranium of an embryo pig, Sus scrofa. Am. Jour. Anat., vol. 9.
ScHULTZ, A. IL, 1917. Ein Paariger Knocken am Unterrand der Squama occipitalis. Anat. Rec., vol. 12.
Terry, R. J., 1917. The primordial cranium of the cat. Jour. Morph., vol. 29.
Van Noorden, W., 1887. Beitrag zur An.>»tomie der knorpeligen Schiidelbasis bci menschlichen Embryonen. .Arch. f. Anat. u. Physiol, Anat. Abth.
VoiT, Max, 1909. Das Primordialcranium des Kaninchens unter Berucksichtigimg der Deckknocken. Ein Beitrag zur MorphologiedesSiiugeticrschadels. Anat. Hefte, Bd. 38, Heft. 116.
Explanation of Figures
- Lewis Links: Plate 1 | Plate 2 | Plate 3 | Plate 4 | Plate 5 | Contribution No.39 | Volume IX | Contributions to Embryology
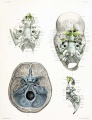
Plate 1
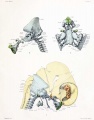
Plate 2
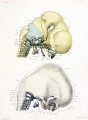
Plate 3
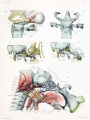
Plate 4
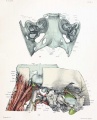
Plate 5
Plate 1

Plate 1: Fig. 1 | Fig. 2 | Fig. 3 | Fig. 4
Fig. 1. Dorsal aspect of base of the cartilaginous skull with the basioccipital in the horizontal plane.
Fig. 2. Right half of dorsal aspect of the base of the cartilaginous skull.
Fig. 3. Dorsal aspect of cartilaginous and membranous skull.
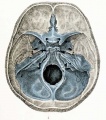
Fig. 4. Dorsal aspect of base of the adult skull.
Plate 2

Plate 2: Fig. 5 | Fig. 6 | Fig. 7
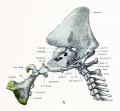
Fig. 5. Median sagittal aspect of the cartilaginous skull.
Fig. 6. Ventral aspect of base of the cartilaginous skull.
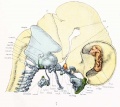
Fig. 7. Lateral aspect of cartilaginous skull and cervical vertebrae, with the brain and cervical cord and hypophysis in position.
Plate 3

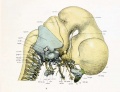
Fig. 8. Lateral aspect of cartilaginous skull and cervical vertebra; with the brain, cervical cord, and nerves.
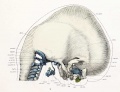
Fig. 9. Lateral view of cartilaginous skull and cervical vertebrie with the overlying membranous skull and the dorsal membrane.
Plate 4

Plate 4: Fig. 10 | Fig. 11 | Fig. 12 | Fig. 13 | Fig. 14
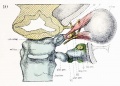
Fig. 10. Dorsal aspect of sphenoid cartilage, showing attachment of the orbital muscles to the basal part of the orbital wing.
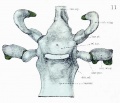
Fig. 11. Dorsal aspect of sphenoid cartilage.
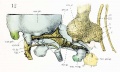
Fig. 12. Lateral view of the right otic region. Part of the malleus and incus cut away showing course of facial nerve and position of otic ganglion.
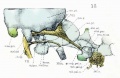
Fig. 13. Lateral view of right otic region showing relations of facial nerve.
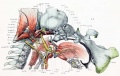
Fig. 14. Lateral view of base of cartilaginous skull with deeper muscles of occipital region and of the mouth and pharynx.
Plate 5

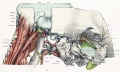
Fig. 15. Lateral view of part of cartilaginous and membranous skull.
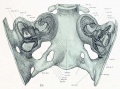
Fig. 16. Dorsal view of temporal and occipital cartilages, showing the relation of the inner ear to the otic capsule.
Abbreviations

| Historic Disclaimer - information about historic embryology pages |
|---|
|
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2024, April 17) Embryology Book - Contributions to Embryology Carnegie Institution No.39. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Book_-_Contributions_to_Embryology_Carnegie_Institution_No.39
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G