2017 Group Project 4
| 2017 Student Projects | |||
|---|---|---|---|
|
Eye Development
Mark Hill (talk) 16:09, 14 September 2017 (AEST) OK Feedback
TO DO
- Timelines
- Key discoveries
- Cellular components
- Current research areas
- Animal models compared to human development
- Central neural pathway
- Developmental Signalling Processes
- Future Questions
Other
- Images for each component of the eye
- Add pictures to Carnegie Stages
- Go through peer-reviews
- Make an introduction for the page
- Fill out Glossary List
- Fill out the missing sections in the development section
- Add videos
- REFERENCES
Introduction
Anatomy of the Adult Eye
The eye is a complex sensory structure which allows a variety of species to intake and process visual information from the world around us.
To discuss the key anatomical components of the eye, we will break down the eye in parts[1]:
Supporting Structures (Orbit, Extraoccular Muscles, Lacrimal Gland, Eyelid)

The orbit of the eye consists of a framework of bones and connective tissue which provide structural support and protection to the sensitive human eye. 7 bones contribute to the orbit of the eye: frontal, lacrimal, sphenoid, zygomtatic, ethmoid, maxilary, palatine. Several openings exist in the orbital structure, 2 key fissures are the superior orbital fissure and inferior orbital fissure and the posterior optic canal. These openings allow crainial nerves passageway.
A set of 6 extraoccular muscles allow for a strong voluntary control of the movement of the eye. These muscles include superior recuts, inferior rectus, lateral rectus, medial rectus, inferior oblique, superior oblique muscles. These muscles are some of the smallest in the human body and are designed to produce fast, controlled motion to focus on an object of interest.
The eyelid is the anterior covering of the eye, consisting of a thin fold of skin. It is controlled by the levitator palpabrae superioris muscle. Superio-laterally to the eyelid is the lacrimal gland, which serves to create a film of tear, an important component to keeping the anterior surface of the eye moist. The eyelid assists in this process by opening and closing, or blinking, which spreads the tear film across the surface of the eye. Tear film and debris caught in the tear film are excreted through the lacrimal duct which is located on the infero-medial surface of the eyelid.

Layers of the Eyeball (Sclera, Choroid, Retina)
The superficial layers of the eyeball is the sclera, a thick fibrous layer which becomes thicker as it extends posteriorly. The sclera is the visible as the white part of our eye. It's function is to provide structural stability to the eye and serves as attachment for some extraoccular insertions. Deep to the sclera is the choroid, a vascular network which provides nourishment to the layers of the eyeball and plays a large role in the growth of the eye. Deep to the choroid is the retina, a network of nerve fibers which transmit visual information. The retina contains 2 types of photoreceptors which convert visible light into nerve impulses. Rods work best in low light environments while cones work best in high light enthronements and detect colour and sharp detail.[2]: The retina does not extend to the anterior part of the eyeball, unlike the sclera and choroid layers.
Anterior structure (Lens, Cornea, Pupil, Iris, Ciliary Body, Conjunctiva Anterior Chamber)
The most anterior component of the eyeball is the cornea, which is a multilayered avascular transparent structure who's major function to the refract light to assist image formation (contributes about 75% of total refraction). Continuous with cornea superiorly and inferiorly is the conjunctiva. Deep to the cornera is the iris, or the coloured ring visible in the eye. The iris consists of 2 muscles which act to enhance and diminish the size of the pupil, a hole in the middle of the iris allowing light to enter the eye. Deep to the pupil and iris is the lens, which is a transparent structure which contributes the other 25% of the eye's refractive power. The lens changes shape through accomodation controlled by the ciliary body muscles. The ciliary body is the anterior extension of the choroid. The function of the lens is to focus images by changing refractive power based on the position and depth of focal objects. In between the anterior surface of the lens and posterior surface of the cornea is the anterior chamber. This chamber is filled with aqueous humor supplied by the ciliary body, who's function is to provide nutrients to ocular structures and maintain intraocular pressure.

Posterior structure (Optic Nerve, Optic Disc, Macula, Fovea, Posterior Chamber)
In the posterior section of the retina is the macula, a central spot in which a high concentration of cone photoreceptors are found. The direct centre of the macula is called the fovea which contains solely cone photoreceptors. The fovea is only part of the eye which can conduct 'perfect' 20/20 vision. It has a high workload and hence a strong vascular supporting structure[3]:. The most posterior part of the eye is the optic nerve (along with the central retinal artery and central retinal vein), which is responsible for transporting nerve impulses to the cerebral cortex for processing. The opening of the optic nerve is called the optic disc, which contains no photoreceptors and is a blind spot of the eye. Between the posterior surface of the lens, the retina and the optic disc is the posterior chamber. The posterior chamber contains a vitreous humour, a jelly like substance with a high viscosity.
Embryonic Contributions
Eye development is a complex process. It involves the formation of the retina and the lens, which we together call the eyeball. It also involves the formation of auxiliary eye structures which are the eyelid, lacrimal gland, and cornea. The retina is formed from the neuroectoderm, the lens from the surface ectoderm and the auxiliary tissues are formed from the head surface ectoderm, neural crest cells and the head mesoderm [4].

| Embryonic contributions | Eye component |
|---|---|
| Neuroectoderm of the forebrain | Retina, Posterior layers of the iris, The optic nerve |
| Surface ectoderm of the head | The lens of the eye, The corneal epithelium |
| Mesoderm between the neuroectoderm and the surface ectoderm | The fibrous and vascular coats of the eye |
| Neural crest cells | Choroid, Sclera, Corneal endothelium |
Timeline of embryonic development
| WEEK | DEVELOPMENT |
|---|---|
| 3-4 | Eye fields, Optic vesicles |
| 5-6 | Optic Cup, Lens Vesicle, Choroid Fissure, Hyaloid Artery |
| 7-8 | Cornea, Anterior Chamber, Pupillary Membrane, Lens, Retina |
| 8-10 | Eyelids |
| 9-15 | Iris, Ciliary Body |
Carnegie Stages
Carnegie Stages is a system of 23 stages used to describe developmental events of the vertebrate embryo. At stage 10 we see early signs related to eye development. The table below is an made of knowledge from Anthony A. Person's article [5].
| Stage | Events |
| Stage 10 (22 days) | The optic primordium has developed. Two small grooves develop on each side of the developing forebrain in the neural folds - this is the optic grooves. The optic placode has begun to develop, which is seen by a small thickening of the surface ectoderm lateral to the hindbrain. |
| Stage 11 (24 days) | Optic vesicle begins to form from the optic groove. |
| Stage 12 (26 days) | The optic vesicles have extended from the forbrain to the surface ectoderm. The optic vesicle now lies close to the surface ectoderm. |
| Stage 13 (28 days) | The optic vesicle interacts with the surface ectoderm and will induce this ectoderm to form the lens placode, which is the precursor of the lens. Optic evagination differentiation makes it possible to identiy optic parts of retina, future pigmented layer of retina and optic stalk. |
| Stage 14 (32 days) | The lens placode is indented by the lens pit and is be cup-shaped. |
| Stage 15 (33 days) | The lens pit will close and form a circle that separates from the surface ectoderm and becomes the lens vesicle. The lens vesicle and optic cub lie close to the surface ectoderm, which creates a slight elevation in the region of the eye. |
| Stage 16 (37 days ) | The lens body has grown and now has a D-shaped lens cavity. First indication of the development of the eyelids visible as eyelid grooves above and below the eye. |
| Stage 17 - 19 | Retinal pigment is visible and the retinal fissure is almost closed. The eyes are still laterally placed but starts to take a more anterior position. The lower eyelid fold develops first and then the eyelid fold above the eye. |
| Stage 18 (44 days) | Mesenchyme invades the region between the lens epithelium and the surface ectoderm. The eyes have shifted to a more anterior position. The groove above and below the eyes are deeper but have not joined yet. |
| Stage 19 - 22 | The eyelid folds develop into the eyelids and the upper and the lower eyelids now meet at the outer canthus, which is the corner of the eye. |
| Stage 20 (51 days) | The lens cavity is lost. The upper and lower lids meet laterally and medially. The eyelids now partly cover the eye. Lens suture begins to form. |
| Stage 23 (57 days) | End of embryonic period. The face begins to look human. Eyelids closure is complete. |

Development of the eye components
Introduction
<html5media width=“480" height="360">https://www.youtube.com/embed/ghHDFWlfpoQ</html5media>

The eye starts to develop at 22 days. The optic grooves (sulci) appears in the neural folds at the cranial end of the embryo. When the neural fold fuse to form the forebrain, the optic grooves will form optic vesicles. The optic vesicles are continuous cavities from the cavity of the forebrain and project from the wall of the forebrain and into the mesenchyme. The optic vesicle extends from the diencephalon and will come in contact with the surface ectoderm of the head. This induces the formation of a lens placode.
The surface ectoderm near the optic vesicles will thicken and form the lens placodes. The lens placodes will sink into the surface ectoderm and form lens pits. The edges of the lens pits will travel towards each other and fuse to form round lens vesicles, which will later lose connection with the surface ectoderm. The optic vesicles do also keep developing - they will form double-walled optic cups which are connected to the brain by the optic stalk. The two layers of the optic cup will differentiate in different directions. The cells of the outer layer will produce melanin pigment and later become the pigmented retina. The cells of the inner layer of the optic cup will proliferate fast and develop glia, ganglion cells, interneurons and light-sensitive photoreceptor neurons. These cells are in the neural retina. The ganglion cells of the retina are neurons that send signals to the brain. The axons of the ganglion cells of the neural retina will grow in the wall of the optic stalk. The cavity in the optic nerve will start disappearing, and instead, the axons of the ganglion cells will form the optic nerve. The optic stalk is now the optic nerve [6]
The optic cups will fold inwards around the lens while the lens vesicles have grown inwards so they have fully lost their connection with the surface ectoderm, which locates them in the cavities of the optic cups. The retinal fissures (linear grooves) will develop and cover the ventral surface of the optic cups and down to the optic stalk. The retinal fissures contain vascular mesenchyme and hyaloid blood vessels will develop here. The hyaloid artery supplies the structures in the eye with blood and the hyaloid vein will return the blood from these structures.
Formation of the optic vesicle
It is a specific area of the neural ectoderm that will become the optic vesicle - this happens because of a group of transcription factors - Six3, Pax6, and Rx1. These transcription factors are expressed in the most anterior tip of the neural plate. This area will split into bilateral regions and form the optic vesicles. The Pax6 protein has shown to be especially important for the development of the lens and retina. This protein is important for photoreceptive cells in all phyla. Pax 6 is also present in the murine forebrain, hindbrain, and nasal placodes, but the eyes are most sensitive its absence [6].
The sonic hedgehog gene is important for the separation of the single eye field into two fields. If this gene is inhibited, the eye field will not split which will result in cyclopia, a single eye in the center of the face [6].
Retina

The retina is the essential component of the eye with the primary function of photoreception [7]. Formation of the retina begins with the specification of retinal cells in the anterior neuroectoderm. First morphological sign of eye development is the formation of two lateral grooves in the anterior neuroectoderm, which is called the optic sulci. The cells of the optic sulci will evaginate and form the optic vesicle. The distal portion of the optic vesicle will form the retina and the proximal will form the optic stalk [8]. The retina develops from the walls of the optic cup, these walls develop into the two layers of the retina. The thin, outer layer of the optic cup will become the pigment layer of the retina and the thick, inner layer of the optic cup will differentiate into the neural retina. Forkhead transcription factors are regulating the proliferation and differentiation of the retinal precursor cells. During the embryonic and early fetal periods, the two layers of the retina are separated which makes an intraretinal space. The intraretinal space is derived from the cavity of the optic cup. The two layers of the retina will fuse and the intraretinal space will gradually disappear.
When the lens is developing the inner layer of the optic cup starts to proliferate and forms a thick neuroepithelium which will later differentiate into the neural retina. The neural retina contains photoreceptors and the cell bodies of neurons and is the light-sensitive region of the retina. The axons of the ganglion cells in the neural retina grow in the wall of the optic stalk and will form the optic nerve.
Lens
Human lens induction occurs at around 28 days and is completed around day 56. The surface ectoderm will thicken near the optic vesicle and create the lens placode and later form the lens vesicle [9]. Lens cells come from ectoderm and differentiate into either lens fibers or the lens epithelium. The anterior monolayer of epithelial cells of the lens will create the lens epithelium, which makes the sheet of cuboidal epithelium covering the anterior surface of the lens. The posterior lens vesicle cells will produce the linear primary fibre cells aligned parallel to the optic axis. These fibres will create the lens mass and form the embryonic lens nucleus. The lens epithelial cells will keep proliferating and produce new cells which generate a secondary lens fiber cells. This rows of cell will form the outer shells and keep the lens growing throughout life. This makes the eye lens unique - it will have an addition of new cells inside the surrounding capsule all the time [10].
Ciliary Body
The ciliary body is a muscular and secretory tissue and is located directly behind the lens. The ciliary body forms part of the anterior segment of the eye and is an important regulator of eye physiology and the vision. The ciliary body produces the aqueous fluid which fills the eyes and nourishes the lens and cornea - this aqueous fluid function to keep the eye in a pressurized and inflated state, which is important the vision.The ciliary body also synthesizes collagenIX and tenasin-C [11].
The ciliary body extends from the iris root (anteriorly) to the ora serrata (posteriorly). It consist of ciliary muscles and ciliary processes. Each ciliary process (fold) is covered by a double-layered secretory epithelium; the outer pigmented and the inner unpigmented (closes to the lens). The epithelial layers of the ciliary body comes from the retina of the optic cup. We see two different ciliary body epithelium: the inner non-pigmented ciliary epithelium which is connected with the neural retina and the outer pigmented ciliary epithelium which is connected with the retinal pigmented epithelium. The epithelium of the iris is the further anterior extension. The epithelial layers are associated with a stroma containing the ciliary muscle. The ciliary muscle is complex, but can be divided into three portions; anterior, posterior and internal [12].
In the chick eye development, we see the mesenchyme grow together on the margin of the optic cup and will form the stroma of the ciliary body and the iris, which is located more anteriorly. It is unknown how if the lens has a role in inducing the secretory ciliary body epithelium and the muscular iris epithelium [13].
Iris
NEED MORE INFO
The Iris develops at the end of the third month of development. The iris is a thin layer and derives from the anterior rim of the optic cup. In the anterior of the eye, the optic epithelium is nonneural and matures as ciliary body and iris epithelia.
Cornea
The cornea is the unique, transparent and avascular component located at the most anterior section of the eye. It is composed of five layers stemming from the 3 main layers of epithelium, stroma and endothelium:
- Corneal epithelium (most anteriorly) – consisting of multiple layers of cells which rest on a basement membrane.
- Bowman’s membrane – a clear, acellular homogenous layer. This layer is a modified portion of the stroma and has no regenerative capacity as a response to damage [14].
- Corneal stroma – largest portion of the retina consisting of collagenous tissue.
- Descemet’s membrane – a homogenous, elastic true basement layer with a regenerative capacity.
- Corneal endothelium – a single layer of cuboidal cells responsible for the maintenance of the dehydrative nature of corneal tissue. Cite error: The opening
<ref>tag is malformed or has a bad name.
Corneal development involves interactions between ectoderm-derived tissues. More specifically, the corneal epithelium arises from interactions between the cranial ectoderm and optic vesicles derived from the neural ectoderm. [15]. The neural tube and the adjacent ectoderm give rise to the neural crest cells which are multipotent with a migratory capacity. These cells contribute to the formation of corneal epithelium and the stromal cells. Innervation of the cornea stroma and epithelium is derived from both the neural crest and the ectodermal placode Cite error: The opening <ref> tag is malformed or has a bad name. The endothelium is comprised of differentiated neural crest cells which contribute to the later formation of Descemet’s membrane [16].
Aqueous Chambers
Choroid and Sclera
The choroid and sclera are both surround the eye and are located adjacent to each other (the choroid lines the inner portion of the sclera). The sclera is the outer, fibrous white layer of the eye which functions as a supportive wall of the eye ball and is continuous with the cornea [17]. The heavy vasculature of the choroid provides a major supply of oxygen and nourishment to the retina [18]. They are derived from interactions between the neural crest and mesoderm-derived mesenchyme. The sclera forms as a result of condensation of the mesenchyme outside the optic cup [19]. The choroid arises from the mesenchyme surrounding the optic vesicle and the cranial neural crest cells. The first vasculature structures appear in the choroid during week 15 in the form of arterioles and venules; veins and arteries become distinguishable by week 22 [20].
Extraocular muscles
The extraocular muscles are responsible for eye movement within the eye orbit – a critical aspect of sight and communication. 6 main muscles contribute to eye movement. These include: superior rectus, inferior rectus, lateral rectus, medial rectus, superior oblique and inferior oblique.
The development of these muscles is dependent on interactions between the eye, its muscles and the neural crest. [21]. Somites supplied by cranial nerve III forms 5 of the 6 extraocular muscles which form an interlocking tendinous ring made of fibrous connective tissue. The 5 muscles include the lateral, medial, superior and inferior rectus, and the superior oblique muscle. This common tendinous ring is known as the annulus of Zinn and surrounds the entry of the optic nerve. The remaining sixth muscle is the inferior oblique muscle and is separate from the aforementioned tendinous ring due to its distinct point of origin. [22].
Eyelids
The eyelids are upper and lower folds of skin functioning as a barrier protecting the anterior surface of the eye from direct local injury. Eyelids also regulate light exposure, distribute precorneal tear film over the corneal surface when blinking and are involved in functions involving tear flow [23]. No sign of eyelid fold development is evident at week 5 however at this stage the lens pit begins to invaginate from the surface ectoderm [24]. Week 6 initiates the timeline of eyelid development as small depressions emerge in the surface ectoderm both above and below the developing eye – these rapidly deepen to form the folds of the eyelids [25]. The lower eyelid fold develops first and, by the end of week 6, the lower fold is more distinct than the upper eyelid fold [26]. Week 7 marks the point at which both the upper and lower folds are distinctly developed and during this stage, the upper lid assumes its more dominant role [24]. The eyelid is not fully closed - there is a gap between the folds and the cornea is slightly visible – they eventually undergo fusion in week 8 [24].
Lacrimal Glands
The lacrimal glands are exocrine glands located in the upper lateral region of each eye orbit which secrete the aqueous potion of the tear film. This film is continuously secreted and functions to clean and protect the external surface of the eye, keeping the environment constantly moist and preventing possible injury from dust and other particles [27]</pubmed>. Lacrimal gland morphogenesis is identified in three stages. • The first is the presumptive glandular stage. This stage involves the thickening of the conjunctival superior fornix epithelium and the condensing of the surrounding mesenchyme [28]. • The second is the bud stage. This stage is considered the initial sign of glandular formation due to the development of nodular structures and lumina within epithelia buds [28]. • The third is the glandular maturity stage (week 9-16). In this stage, the lacrimal gland begins to morphologically mirror the adult gland [28].
Developmental signaling processes
Eye formations is a complicated process, since the different components of the eye are formed from different tissues. Therefore there is a lot of key questions in how these processes are coordinated.
- The proliferation and differentiation of retinal precursor cells are regulated by forkhead transcription factors
- Fibroblast growth factor signaling regulates retinal ganglion cell differentiation
Eye formation in the absence of the retina
The gene Rx is a key component in the formation of retinal structures. Studies have shown that mice lacking Rx function do not form optic sulci or optic vesicles and do not have retina-specific gene expression. There has been made studies in human, medaka, zebrafish and Xenopus which suggest that Rx genes are required for the formation of the vertebrate retina. It is also shown that in Rx-deficient mouse embryos the mature lens does not develop. This indicates that retinal cells are necessary for lens formation, and if the mouse is lacking Rx gene, then it will not display any retina-specific gene expression [29].
It is thought that signaling from the optic vesicle is important to activate the lens-specific gene and the formation of the lens placode. If there are no retinal cells generated, then the morphogenesis of the optic cup does not take place and the lens will not form. The study also showed that elimination of β-catenin expression in the head surface ectoderm in Rx-deficient embryos would develop a lens-like structure even though there were no optic vesicle/cup. Elimination of β-catenin lead to upregulation of Pax6, which is a gene that has a key role in lens formation. This indicates that the optic vesicle/cup might not be required for lens formation. The lens developed smaller than in wild type embryos, which means that the optic cup/vesicle might have an effect on a determination of the size of the lens. It could be that the developing optic vesicle/cup shields the developing lens placode from signals for other tissues that could disturb the lens formation.
It is not fully understood if the formation of the auxiliary eye structures is dependent on the development of the eyeball and its components, the retina and the lens. A study made my Eric C. Swindell and al. showed that auxiliary eye structures form even in the absence of retinal morphogenesis and retina-specific gene expression. This indicates that there are two separate developmental processes involved in the formation of the eye and its associated structures. In the absence of the retina and the lens, we still see specific gene expression in the surface ectoderm initiating the formation of auxiliary eye structures like eyelids and lacrimal glands [30].
Current research
Iris epithelium cells as a potential source for regenerative medicine for retinal pigment epithelium degeneration
The retinal pigment epithelium (RPE) is a monolayer of neural-crest-derived cells and is located between the photoreceptors and the choroid. There are many disorders that involve dysfunctional RPE and leads to retinal degenerative diseases. There is no treatment for these diseases, therefore, one of the most promising future therapy for RPE related disorders is to replace the cells of the dysfunctional RPE.
Studies have focused on the development and use of induced pluripotent stem cells (iPSC) for cell replacement therapy. It is possible to differentiate iPSC in vitro towards RPE cells and use them for experimental transplantation studies in animal models. But an alternative strategy for a retinal replacement is based on transdifferentiation (direct conversion) and involves transforming an adult somatic cell into another adult somatic cell. Use of overexpression of cell-lineage specific genes can convert one cell into another cell type and skip the pluripotent state. Iris epithelium (IE) cells might be able to be transdifferentiated into the RPE since both RPE and IE are neuro-epithelia and has a common embryological origin - they both derive from neuroectoderm of the developing optic cup.
A study made by Anna Bennis et al. [31] has shown that the canonical pathways for the most highly expressed genes of the IE and the RPE were very similar. Wnt signaling pathways has shown to be active in the IE but not in the RPE. The Wnt signaling pathway consists of a group of signal transduction pathways that has an influence on cell fate determination and cell proliferation during embryonic development. Activation of Wnt SP makes it possible to reprogram somatic cells (also retinal neurons) into iPSC and is important for the differentiation of pluripotent stem cells to RPE cells. The high expression of Wnt SP genes in the iE suggests that the IE has a multipotent character during life and therefore could be transformed into RPE cells used for RPE replacement.
Animal Models
Congenital Abnormalities
Although quite rare, abnormalities can occur during the embryonic development that causes serious effects to the features of the eyes, its function as well as the further normal development after birth. These abnormalities can occur unilaterally or bilaterally and it is also common for most reported cases to have more than one type of eye abnormalities at a time. Thus, the estimated prevalence statistics listed in the table can overlap.
| Congenital Anomalies | Description | Epidemiology |
|---|---|---|
| Anopthalmia | The absence of the eyes. | Prevalence: Anophthalmia-Microphthalmia Syndrome was estimated to be 5.3 per 100 000 cases [32]
Cause: SOX2 gene mutation [33] |
| Microphtalmia | Abnormal reduction in size of the eyeballs. | |
| Coloboma | A condition where parts of the eye is missing and failed to develop normally. | Prevalence: Occular coloboma was estimated to be 8.0 per 100 000 cases [32] |
| Aniridia | Complete or partial absence of the iris. | Prevalence: Estimated to be 1.5 per 100 000 cases [32] |
| Optic Nerve Hypoplasia | A condition where the optic nerves are underdeveloped. | Prevalence: Reported to be the most common abnormality [34]
|
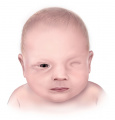
Figure 2. Anopthalmia
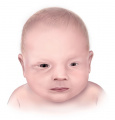
Figure 3. Microphthalmia

Figure 4. Iris Coloboma

Figure 5. Partial Aniridia




- Incomplete Information
Future questions
Glossary
| Term | Definition |
| Choroid | The pigmented vascular layer of the eye located between the retina and sclera. |
| Ciliary body | The connection between the iris and choroid consisting of the ciliary muscle, ciliary processes and the ciliary ring. Alters the shape of the lens and ciliary epithelium in order to focus on an image. |
| Cornea | The transparent anterior part of the external coat of the eye covering the iris and pupil and continuous with the sclera. |
| Ectoderm | The outermost primary germ cell layer in the early embryo. |
| Endoderm | The innermost primary germ cell layer in the early embryo. |
| Ectodermal placode | A neurogenic placode consisting of an area of thickened epithelium in the embryonic head ectoderm layer that gives rise to neurons and other structures of the sensory system. |
| Extraocular muscles | Muscles controlling eye movement. |
| Iris | The coloured muscular ring surrounding the pupil which manipulates the diameter and size of the pupil and controls the amount of light entering the eye. |
| Lens | The transparent biconvex structure posterior to the iris with the main function of focusing light onto the retina in order to form clear images of objects at various distances. |
| Lens pit | A pit-like depression in the fetal head where the lens develops. |
| Lumina | the central cavity of a tubular structure. |
| Neural crest | A temporary group of embryonic ectodermal cells. |
| Neural tube | A hollow tubular structure as a result of neural plate folding which later develops into the brain and spinal cord. |
| Pupil | The central opening in the iris, allowing and regulating the entry of light into the eye. |
| Retina | The sensory membrane that forms the inner lining of the eye containing rods and cones with the main function of photoreception. Nerve impulses in response to visual information are transmitted to the brain via the optic nerve. |
| Sclera | The dense, white, fibrous membrane that, along with the cornea, forms the external protective covering of the eye |
| Tear film | A layer of tears which spreads over the eye, ultimately maintaining the health of the ocular surface by making the external surface smooth and clear.
|
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
References
- ↑ Chen, J., 2011. Handbook of Visual Display Technology. Springer Berlin Heidelberg. Chapter 2.1
- ↑ Curcio CA, Hendrickson AE. Organization and development of the primate photoreceptor mosaic. Prog Ret Ret. 1991;10:89–120.
- ↑ <pubmed>6462623</pubmed>
- ↑ <pubmed>18675797 </pubmed>
- ↑ <pubmed>7364662</pubmed>
- ↑ 6.0 6.1 6.2 Gilbert SF. Developmental Biology. 6th edition. Sunderland (MA): Sinauer Associates; 2000. Available from: https://www.ncbi.nlm.nih.gov/books/NBK9983/
- ↑ <pubmed>7084144</pubmed>
- ↑ <pubmed>18675797 </pubmed>
- ↑ <pubmed>20171212 </pubmed>
- ↑ <pubmed>25406393</pubmed>
- ↑ <pubmed>17275804</pubmed>
- ↑ <pubmed>12127103</pubmed>
- ↑ <pubmed>17275804</pubmed>
- ↑ <pubmed>7084144</pubmed>
- ↑ <pubmed>26310148</pubmed>
- ↑ <pubmed>6511224</pubmed>
- ↑ Cite error: Invalid
<ref>tag; no text was provided for refs named’’7084144’’ - ↑ <pubmed>20044062</pubmed>
- ↑ <pubmed>1628748</pubmed>
- ↑ <pubmed>2199235</pubmed>
- ↑ <pubmed>21482859</pubmed>
- ↑ <pubmed>21482859</pubmed>
- ↑ <pubmed>1993591</pubmed>
- ↑ 24.0 24.1 24.2 <pubmed>27124372</pubmed>
- ↑ <pubmed>7364662</pubmed>
- ↑ <pubmed>21416630</pubmed>
- ↑ <pubmed>17001201
- ↑ 28.0 28.1 28.2 <pubmed>14635806</pubmed>
- ↑ <pubmed>18675797 </pubmed>
- ↑ <pubmed>18675797 </pubmed>
- ↑ <pubmed>28827822</pubmed>
- ↑ 32.0 32.1 32.2 Orphanet Report Series – Rare Diseases Collection. (2017). Prevalence of rare diseases: Bibliographic data (no. 2). Paris. France: Author. Retrieved from: [[1]]
- ↑ Mauri, L., Franzoni, A., Scarcello, M., Sala, S., Garavelli, L. Modugno, A., ... Penco, S. (2015). SOX2, OTX2 and PAX6 analysis in subjects with anophthalmia and microphthalmia. European Journal of Medical Genetics, 58, 66-70. doi: 10.1016/j.ejmg.2014.12.005
- ↑ Ryabets-Lienhard, A., Stewart, C., Borchert, M., & Geffner, M. E. (2016). The Optic Nerve Hypoplasia Spectrum - Review of the Literature and Clinical Guidelines. Advances in Pediatrics, 63, 127-146. doi: 10.1016/j.yapd.2016.04.009
External links
z5075309 - <pubmed>26956898</pubmed>